Back to Journals » Journal of Pain Research » Volume 15
Z-Guggulsterone Relieves Neuropathic Pain by Inhibiting the Expression of Astrocytes and Proinflammatory Cytokines in the Spinal Dorsal Horn
Authors Dang SJ, Wei WB, Li RL ![]() , Song CX
, Song CX ![]() , Xu J
, Xu J
Received 2 February 2022
Accepted for publication 2 April 2022
Published 5 May 2022 Volume 2022:15 Pages 1315—1324
DOI https://doi.org/10.2147/JPR.S360126
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Qi Fang
Sha-Jie Dang,1,2,* Wen-Bo Wei,3,* Rui-Li Li,4,* Can-Xu Song,5 Jin Xu1
1The Key Laboratory of Biomedical Information Engineering of Ministry of Education, Institute of Health and Rehabilitation Science, School of Life Science and Technology, Xi’an Jiaotong University, Xi’an, 710049, People’s Republic of China; 2Department of Anesthesiology, Shaanxi Provincial Cancer Hospital, Xi’an, 710061, People’s Republic of China; 3Department of Orthopedics, Shaanxi Provincial People’s Hospital, Xi’an, 710068, People’s Republic of China; 4Department of Pharmacy, Xijing Hospital, Fourth Military Medical University, Xi’an, 710032, People’s Republic of China; 5Department of Ultrasound, Shaanxi Provincial Cancer Hospital, Xi’an, 710061, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jin Xu, The Key Laboratory of Biomedical Information Engineering of Ministry of Education, Institute of Health and Rehabilitation Science, School of Life Science and Technology, Xi’an Jiaotong University, Xi’an, 710049, People’s Republic of China, Email [email protected]
Objective: The study objective was to investigate whether Z-guggulsterone can relieve neuropathic pain in sciatic nerve chronic constriction injury (CCI) mice by inhibiting the expression of astrocytes and proinflammatory cytokines in the spinal dorsal horn.
Methods: Neuropathic pain was induced and assessed in CCI mice. Z-guggulsterone was administered multiple times via intraperitoneal injection. Pain behaviour assessments were made by conducting paw withdrawal mechanical threshold (PWMT) and thermal withdrawal latency (TWL) tests. The expression level of the glial fibrillary acidic protein (GFAP) in the spinal dorsal horn was observed by immunofluorescence. The levels of the proinflammatory cytokines, IL-1β, IL-6 and TNF-α in the spinal cord were measured by ELISA. Data were analysed using one-way ANOVA or two-way ANOVA.
Results: The PWMT and TWL were higher on the 5th, 7th, 10th and 14th days after CCI, the expression level of GFAP in the spinal dorsal horn was lower, and the levels of IL-1β, IL-6 and TNF-α in the spinal cord were lower in the CCI+Z-GS-L, CCI+Z-GS-M and CCI+Z-GS-H groups than in the CCI+Veh group in a dose-dependent manner (P < 0.05).
Conclusion: Z-guggulsterone can relieve neurological pain in CCI mice, which may be related to the inhibition of astrocytes and proinflammatory cytokines in the spinal dorsal horn.
Keywords: neuropathic pain, Z-guggulsterone, spinal dorsal horn, astrocytes, proinflammatory cytokines
Introduction
Neuropathic pain (NP) is a chronic pain disease with a high incidence rate and complicated pathogenesis.1 Its treatment is difficult and urgently needs to be addressed by the medical community.2 The mechanism of NP is very complex and not yet fully understood. Multiple studies have shown that inflammation plays an important role in neuropathic pain.3 The key factors for the formation of neuroinflammation are the activation of astrocytes in the spinal dorsal horn and the release of inflammatory cytokines, which are involved in NP through different signalling pathways.3 The spinal dorsal horn, which is the primary centre of the transmission, processing and integration of pain signals, plays a key role in NP.4 Nerve injury or inflammation causes the activation of astrocytes in the spinal dorsal horn, which regulates the release of proinflammatory cytokines and then increases the excitability of neurons, finally triggering pain and its maintenance.5–7 Proinflammatory cytokines, such as interleukin (IL)-6, IL-1β, and tumour necrosis factor (TNF)-α, in the spinal dorsal horn cause pain hypersensitivity through central sensitization.8 Therefore, targeting astrocytes activation and proinflammatory cytokines action in the spinal dorsal horn can be a potential therapeutic strategy to treat NP.
Neuropathic pain belongs to the “Bi syndrome” category in traditional Chinese medicine.9 Traditional Chinese medicine has unique advantages in the prevention and treatment of NP because it possesses fewer complications and side effects than those of synthetic drugs.10 Myrrh can promote qi, eliminate swelling, disperse blood stasis and relieve pain; therefore, it is used to treat pain.11 However, its analgesic effects and mechanism of action are still unclear. Z-guggulsterone is the main active component of myrrh, which exerts various pharmacological effects.12,13 Several studies have shown that the molecular target of Z-guggulsterone is closely related to pain through its anti-inflammatory effect.14–16 At present, there are few reports on the application of Z-guggulsterone to treat NP.
Therefore, this paper describes an investigation of the effect of Z-guggulsterone on pain behaviour, astrocytes and proinflammatory cytokines in the spinal dorsal horn in mice subjected to sciatic nerve chronic constriction injury (CCI) to explore whether Z-guggulsterone relieves NP and whether it is associated with the inhibition of astrocytes and proinflammatory cytokines in the spinal dorsal horn. This study will provide new ideas for the treatment of NP and provide a theoretical basis for the clinical application of Z-guggulsterone.
Materials and Methods
Animal Preparation and Ethics Statement
Adult male C57BL/6J mice, which were purchased from the Fourth Military Medical University and weighed 18~22 g, were placed in cages at room temperature (22~25 °C) and 40%-60% humidity with free access to food and tap water under a 12/12 h light/dark cycle. The experimental procedures carried out on animals were approved and reviewed by the Ethical Committee of the Fourth Military Medical University. All animal experiments were conducted according to the Guidelines of the NIH for the welfare of laboratory animals.
Drug and Reagents
Z-guggulsterone (>98%, CAS: 39025–23-5) was obtained from Yuanye Biotechnology Co., Ltd. (Shanghai, China). TNF-α, IL-1β and IL-6 ELISA kits were provided by Abcam Technology (Cambridge, The United Kingdom). Mouse anti-GFAP (GA5) was obtained from Cell Signaling Technology (Danvers, USA). Alexa 594-labelled donkey anti-mouse IgG was obtained from Thermo Fisher Scientific (Waltham, USA). A BCA Kit was obtained from Jingcai (Xi’an, China). RIPA lysis buffer and protease inhibitor were purchased from Kangwei (Jiangsu, China). Primary antibody and secondary antibody dilutions for Western blotting and immunofluorescence were purchased from Biyuntian (Shanghai, China). DMSO (the vehicle for Z-guggulsterone) was obtained from Sigma (Saint Louis, USA).
Experimental Design
Thirty-six C57BL/6J mice aged 8~10 weeks were selected and randomly divided into 6 groups (n = 6/group): Sham+Veh (sham treated with vehicle), Sham+Z-GS-H (sham treated with 50 mg/kg Z-guggulsterone), CCI+Veh (CCI model treated with vehicle), CCI+Z-GS-L (CCI model treated with 10 mg/kg Z-guggulsterone), CCI+Z-GS-M (CCI model treated with 30 mg/kg Z-guggulsterone), and CCI+Z-GS-H (CCI model treated with 50 mg/kg Z-guggulsterone). The sham group and CCI group received the same volume of vehicle. Z-guggulsterone was administered multiple times via intraperitoneal injection once daily from the 5th to the 14th day after CCI. Pain behaviour assessments, including the PWMT and TWL tests, were conducted before CCI and on the 1st, 3rd, 5th, 7th, 10th, and 14th days after CCI. On the 14th day after CCI, three mice were randomly selected from each group, and the expression level of GFAP in the spinal dorsal horn was determined by immunofluorescence. The other three mice in each group were used to examine the levels of IL-1β, IL-6 and TNF-α in the spinal cord by ELISA (Figure 1).
 |
Figure 1 Z-guggulsterone and experimental design. (A) Chemical structure of Z-guggulsterone. (B) Detailed timeline of the experiments. Abbreviations: CCI, sciatic nerve chronic constriction injury; PWMT, paw withdrawal mechanical threshold; TWL, thermal withdrawal latency; IF, immunofluorescence; ELISA, enzyme-linked immunosorbent assay; i.p. intraperitoneal. |
CCI Model
The sciatic nerve chronic constriction injury (CCI) model is a classic animal model of NP.17 Thus, this model was used in this study. Briefly, mice were anaesthetized by continuous inhalation of sevoflurane, and the left common sciatic nerve of each mouse was exposed at the middle of the thighbone in the lateral position. Close to the trifurcation, the sciatic nerve was separated from the adhered tissue, and 3 ligatures (5–0 chronic gut) were loosely tied around it with approximately 1 mm between each, and the ligatures were tightened to the point where the left hind limb slightly contracted. Then, the skin layers and muscle were sutured, and iodine was used to disinfect the surgical area. After the operation, the mice were placed in an incubator to monitor their recovery from anaesthesia.
Behavioural Assessments of Mechanical and Thermal Hyperalgesia
All manipulations were performed under quiet conditions by the same experimenter in a test room to prevent stress. Before the behavioural experiment, animals were placed in the test environment for 30 min each day for a total of 3 days.
Paw Withdrawal Mechanical Threshold (PWMT)
Calibrated von Frey filaments (Stoelting, Kiel, WI, USA) were used to assess claw contraction following mechanical stimulation. Mice were placed on an elevated mesh grid (aperture of approximately 0.5 cm×0.5 cm) with a glass frame (20 cm×20 cm×25 cm). A series of von Frey filaments with stimulation intensities of 0.04, 0.07, 0.16, 0.4, 0.6, 1.0 and 1.4 g were used to stimulate the central plantar area of each hind paw. The force was applied for approximately 5 s until the filament adopted an “s” shape and was then repeated 10 times on each hind paw at 3~5 s intervals. The strength of the filaments with more than five paw retraction reflexes was recorded as the mechanical stimulus retraction threshold (PWMT). If the maximum stimulation intensity (1.4 g) did not produce a foot reflex, then the threshold was recorded as 1.4 g.
Thermal Withdraw Latency (TWL)
Paw withdrawal in response to noxious thermal stimuli was assessed using a Full-Automatic Plantar Analgesia Tester (IITC Life Science, Woodland Hills, California, USA) that produces radiant heat by directing a beam of light onto the plantar surface of the hind paw. Mice were placed in clear plastic boxes on a 2 mm thick glass plate (20 cm×20 cm×25 cm). The mice were placed on a 30 cm holder. A set voltage of 10 mV and a 100 W halogen projector lamp were applied as the sources of heat pain. By adjusting the distance between the glass plate and the light source, thermal radiation with a diameter of 5 mm was applied to the plantar surface of the paw of each mouse. The time from the initiation of thermal radiation to the reduction in foot reflex time was recorded in seconds, with 10 s for each test interval. The measurements were repeated five times, and the average number of Hargreaves test values was recorded as the thermal withdrawal latency. A cut-off time of 20 s was adopted to prevent possible tissue damage, and the average value was recorded as 20 s if this cut-off was reached.
Immunofluorescence
Mice were deeply anaesthetized with 1% pentobarbital sodium (50 mg/kg) by intraperitoneal (i.p.), immediately perfused with 30 mL of precooled saline followed by 50 mL of 4% paraformaldehyde (PFA) through the aorta. The whole spinal cord was removed and immersed in 4% PFA for 4 h at room temperature (RT). Then, the spinal cords were dehydrated overnight with 20% and 30% sucrose solutions at 4 °C. The lumbar enlargement of the spinal cord was trimmed and sliced into 25 μm-thick sections with a Cryostat Microtome (Leica, Wetzlar, Germany). The samples were mounted on 3-aminopropyl-triethoxysilane-coated glass slides. Spinal cord sections were stored at −20 °C. Immunofluorescence was performed as follows. The sections were air-dried and circled with an immunity staining guard pen. The slices were permeabilized with 0.2% Triton X-100 in PBS at RT for 20 min. The sections were washed with PBS (3×10 min) and blocked with donkey serum (5%) at RT for 2 h. The primary antibody (mouse anti-GFAP antibody, 1:500) was added for incubation overnight at 4 °C. The sections were then washed with PBS (3×10 min). The secondary antibody (donkey anti-mouse IgG with Alexa Fluor 594, 1:1000) was added for incubation in the dark for 2 h at RT. The sections were washed with PBS (3×10 min) and cover slipped using mounting liquid. A confocal laser scanning microscope (FV1000, Olympus, Tokyo, Japan) was used to observe the SDH, and images were collected with a CCD camera system. Digital images were captured using FV10-ASW-1.6 software (Olympus, Tokyo, Japan). The relative fluorescence densities of the target proteins within a fixed area of 250 μm × 250 μm in the SDH were analysed by ImageJ (ImageJ Software, USA).
Enzyme-Linked Immunosorbent Assay (ELISA)
Mice were deeply anaesthetized with intraperitoneal injection of 1% pentobarbital sodium (50 mg/kg) followed by immediate perfusion with 30 mL of precooled saline through the aorta. The spinal cords on the injured side, including the lumbar enlargement and approximately 1 cm in length, were obtained. Protein extraction was performed as follows. The obtained spinal cords were placed into a centrifuge tube, and RIPA lysis buffer and protease inhibitor were added. The samples were homogenized intermittently for 4 min on ice and centrifuged at 1000 rpm at 4 °C for 15 min. The precipitate was discarded, and the supernatant was transferred to another centrifuge tube. Then, the protein concentrations in the samples were analysed with a BCA kit. ELISA was performed according to the directions of the kits. The absorbance was measured at 450 nm with an enzyme labelling instrument (Thermo Fisher Scientific, Waltham, USA), and standard curves were constructed. According to the sample OD value, the levels of IL-1β, IL-6 and TNF-α in the spinal cord were calculated.
Statistical Analysis
All data are expressed as the means ± S.D. Statistical analysis was performed with SPSS 18.0 (SPSS, Inc., IBM). Graphic images were generated using GraphPad Prism 9 (GraphPad Software, La Jolla, CA, USA). One-way ANOVA followed by least significant difference (LSD) for post hoc analysis was used for comparisons between groups. Two-way repeated-measures ANOVA with Bonferroni confidence interval adjustment was used to compare two groups at different time points. Values of P< 0.05 were considered statistically significant.
Results
Z-Guggulsterone Attenuates Mechanical and Thermal Hyperalgesia in CCI Mice
In the CCI+Veh group, the PWMT began to decrease on the 1st day after CCI (0.87 ± 0.21 g), continued to decrease until the 7th day after CCI (0.19 ± 0.11 g) and was then maintained until the 14th day after CCI (0.12 ± 0.49 g). These results were significantly different from those in the Sham+Veh group (P<0.05, Figure 2A). In the Sham+Veh group and Sham+Z-GS-H group, the PWMT was unchanged before and after surgery, and there was no significant difference in the PWMT at each time point between the two groups (P>0.05, Figure 2A). In the CCI+Z-GS-L, CCI+Z-GS-M and CCI+Z-GS-H groups, 10, 30 and 50 mg/kg Z-guggulsterone were injected intraperitoneally from the 5th day to the 14th day after CCI, respectively, and the PWMT increased on the 5th, 7th, 10th and 14th days after CCI. The results from these groups were significantly different from those of the CCI+Veh group and showed dose dependence (P<0.05, Figure 2A).
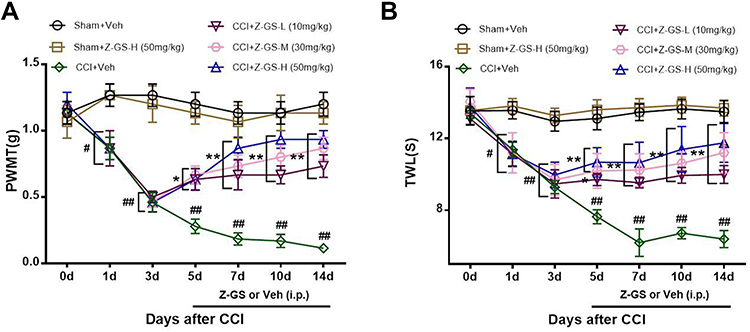 |
Figure 2 Effects of Z-guggulsterone on the PWMT and TWL induced by CCI in mice. (A) PWMT. (B) TWL. #p<0.05, ##p<0.01 vs Sham+Veh group; *p<0.05, **p<0.01 vs CCI+Veh group; n = 6. All data are presented as the means ±S.D. Two-way repeated analyses of variance (ANOVAs) were used for statistical analysis. Abbreviations: PWMT, paw withdrawal mechanical threshold; TWL, thermal withdrawal latency; CCI, sciatic nerve chronic constriction injury; Z-GS, Z-guggulsterone; Z-GS-L, Z-guggulsterone low dose (10 mg/kg); Z-GS-M, Z-guggulsterone middle dose (30 mg/kg); Z-GS-H, Z-guggulsterone high dose (50 mg/kg); Veh, vehicle. |
In the CCI+Veh group, the TWL began to decrease on the 1st day after CCI (11.37 ± 1.01 s), continued to decrease until the 7th day after CCI (6.20 ± 1.87 s) and was then maintained until the 14th day after CCI (6.40 ± 1.16 s). These results were significantly different from those of the Sham+Veh group (P<0.05, Figure 2B). In the Sham+Veh group and Sham+Z-GS-H group, the TWL was unchanged before and after surgery, and there was no significant difference in the TWL at each time point between the two groups (P>0.05, Figure 2B). In the CCI+Z-GS-L, CCI+Z-GS-M and CCI+Z-GS-H groups, the TWL increased on the 5th, 7th, 10th and 14th days after CCI. The results from these groups were significantly different from those of the CCI+Veh group and showed dose dependence (P<0.05, Figure 2B).
Z-Guggulsterone Inhibits the Expression of GFAP in the Spinal Dorsal Horn of CCI Mice
There was no difference in the expression level of GFAP in the spinal dorsal horn between the Sham+Z-GS-H group and the Sham+Veh group (P>0.05, Figure 3). The expression level of GFAP in the spinal dorsal horn on the injured side was significantly higher in the CCI+Veh group than in the Sham+Veh group (P<0.01, Figure 3). The expression levels of GFAP in the spinal dorsal horn on the injured side were lower in the CCI+Z-GS-L, CCI+Z-GS-M and CCI+Z-GS-H groups than in the CCI+Veh group in a dose-dependent manner (P<0.05, Figure 3).
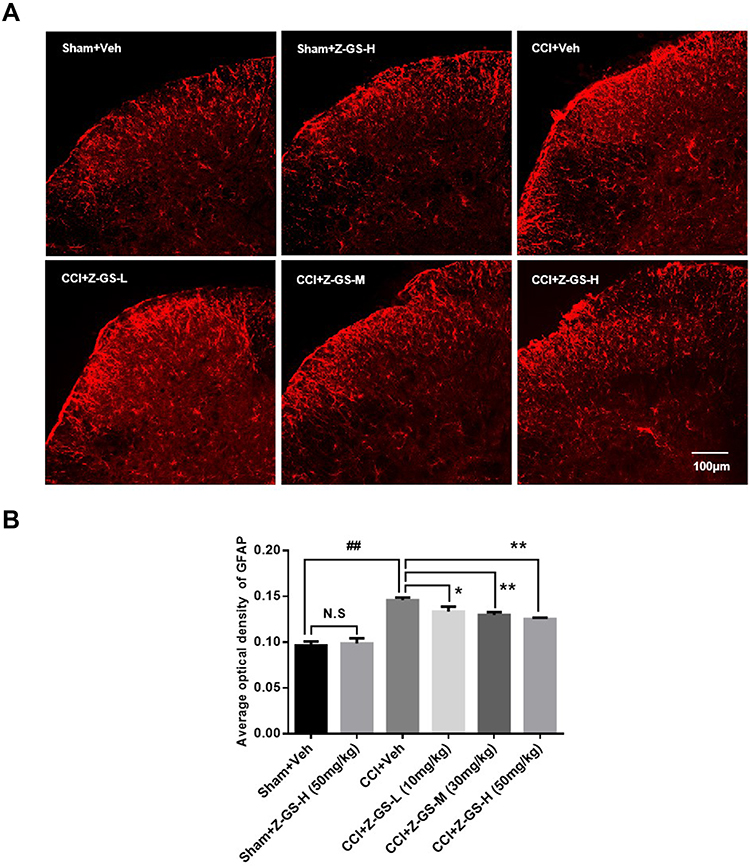 |
Figure 3 Effect of Z-guggulsterone on the expression of GFAP in the spinal dorsal horn of CCI mice. (×20) (A) Immunofluorescence analysis. (B) Histogram of the average optical density (AOD). ##p<0.01 vs Sham+Veh group; *p<0.05, **p<0.01 vs CCI+Veh group; n = 3. All data are presented as the means ±S.D. Scale bar = 100 µm. One-way ANOVA followed by least significant difference (LSD) for post hoc analysis was used for statistical analysis. Abbreviations: GFAP, glial fibrillary acidic protein; CCI, sciatic nerve chronic constriction injury; Z-GS, Z-guggulsterone; Z-GS-L, Z-guggulsterone low dose (10 mg/kg); Z-GS-M, Z-guggulsterone middle dose (30 mg/kg); Z-GS-H, Z-guggulsterone high dose (50 mg/kg); Veh, vehicle. |
Z-Guggulsterone Attenuates the Levels of IL-1β, IL-6 and TNF-α in the Spinal Cord of CCI Mice
There was no difference in the level of IL-1β in the spinal cord between the Sham+Z-GS-H group and the Sham+Veh group (P>0.05, Figure 4A). The level of IL-1β in the spinal cord on the injured side was significantly higher in the CCI+Veh group (118.77 ± 9.96) than in the Sham+Veh group (31.69 ± 4.74) (P<0.01, Figure 4A). The levels of IL-1β in the spinal cord on the injured side were lower in the CCI+Z-GS-L group (73.68 ± 3.80), CCI+Z-GS-M group (53.22 ± 5.37) and CCI+Z-GS-H group (39.76 ± 5.22) than in the CCI+Veh group in a dose-dependent manner (P<0.05, Figure 4A).
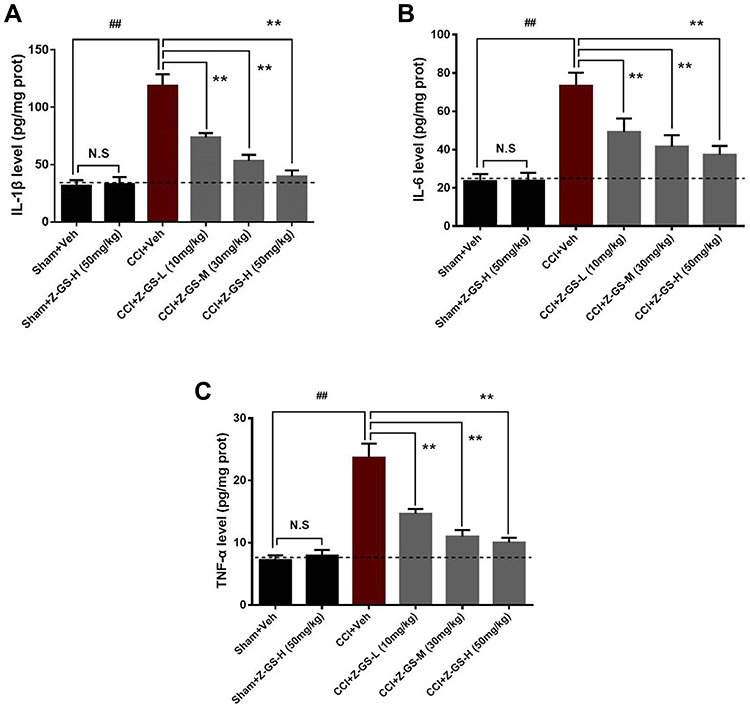 |
Figure 4 Effect of Z-guggulsterone on the levels of IL-1β, IL-6 and TNF-α in the spinal cord of CCI mice. (A) IL-1β. (B) IL-6. (C) TNF-α. ##p<0.01 vs Sham+Veh group; **p<0.01 vs CCI+Veh group; n = 6. All data are presented as the means ±S.D. One-way ANOVA followed by least significant difference (LSD) for post hoc analysis was used for statistical analysis. Abbreviations: IL-1β, interleukin 1β; IL-6, interleukin 6; TNF-α, tumour necrosis factor-α; CCI, sciatic nerve chronic constriction injury; Z-GS, Z-guggulsterone; Z-GS-L, Z-guggulsterone low dose (10 mg/kg); Z-GS-M, Z-guggulsterone middle dose (30 mg/kg); Z-GS-H, Z-guggulsterone high dose (50 mg/kg); Veh, vehicle. |
There was no difference in the level of IL-6 in the spinal cord between the Sham+Z-GS-H group and the Sham+Veh group (P>0.05, Figure 4B). The level of IL-6 in the spinal cord on the injured side was significantly higher in the CCI+Veh group (73.28 ± 6.83) than in the Sham+Veh group (23.64 ± 3.60) (P<0.01, Figure 4B). The levels of IL-6 in the spinal cord on the injured side were lower in the CCI+Z-GS-L group (49.13 ± 7.19), CCI+Z-GS-M group (41.58 ± 5.96) and CCI+Z-GS-H group (37.26 ± 4.74) than in the CCI+Veh group in a dose-dependent manner (P<0.05, Figure 4B).
There was no difference in the level of TNF-α in the spinal cord between the Sham+Z-GS-H group and the Sham+Veh group (P>0.05, Figure 4C). The level of TNF-α in the spinal cord on the injured side was significantly higher in the CCI+Veh group (23.66 ± 2.28) than in the Sham+Veh group (7.25 ± 0.76) (P<0.01, Figure 4C). The levels of TNF-α in the spinal cord on the injured side were lower in the CCI+Z-GS-L group (14.67 ± 0.77), CCI+Z-GS-M group (10.98 ± 1.06) and CCI+Z-GS-H group (10.03 ± 0.78) than in the CCI+Veh group in a dose-dependent manner (P<0.05, Figure 4C).
Discussion
In this study, we demonstrated that Z-guggulsterone significantly alleviated mechanical and thermal hyperalgesia induced by CCI in mice. We observed that Z-guggulsterone inhibited astrocytes activation and the upregulation of the proinflammatory cytokines IL-1β, IL-6 and TNF-α in the spinal dorsal horn of CCI mice.
Currently, NP remains a difficult problem that urgently needs to be solved. In recent years, traditional Chinese medicine has been widely used to treat NP. In recent years, traditional Chinese medicine has been widely used to treat NP.10 A large amount of Chinese medicine literature and substantial modern research have reported the analgesic effects of myrrh, of which guggulsterone is the main active component. Previous studies have found that guggulsterone has antioxidant, anti-inflammatory, hypolipidaemic, antitumour and other effects.12,13 Tianlong Liu14 found that Z-guggulsterone attenuated astrocyte-mediated neuroinflammation after ischaemia by inhibiting the Toll-like receptor 4 pathway. Feng-Guo Liu15 showed that Z-guggulsterone had an antidepressant effect by activating the BDNF signalling pathway in mice. Huang16 confirmed that Z-guggulsterone negatively controlled microglia-mediated neuroinflammation by blocking IκB-α-NF-κB signalling. These studies show that the molecular targets of Z-guggulsterone are closely related to pain, and we speculate that this compound has a potential analgesic effect.
In this study, we examined the mechanical pain threshold and thermal pain threshold by performing paw withdrawal mechanical threshold (PWMT) and thermal withdrawal latency (TWL) tests in CCI mice at different time points. The results showed that the PWMT and TWL decreased on the first day after CCI, continued to decline over 7 days, and then remained stable until the 14th day. This result is in line with those of a previous report.18 The CCI model was therefore established successfully and could meet the needs of subsequent experiments. Furthermore, the results showed that Z-guggulsterone had no effect on the PWMT and TWL in normal mice. From the 5th day to the 14th day after CCI, 10, 30 and 50 mg/kg Z-guggulsterone was injected intraperitoneally (CCI+Z-GS-L, CCI+Z-GS-M and CCI+Z-GS-H groups, respectively), and the PWMT and TWL showed an increasing trend, which was significantly different from that observed in the CCI+Veh group. With increasing Z-guggulsterone concentration, the PWMT and TWL increased in a dose-dependent manner. This finding suggests that Z-guggulsterone can relieve neuropathic pain in CCI mice and that relief is related to the dose of Z-guggulsterone.
In general, inflammation is a pathophysiological condition associated with neuropathic pain.4 Previous studies have shown that proinflammatory cytokines such as TNF-α, IL-1β and IL-6 have potent neuromodulatory effects, which lead to neuropathic pain.19 TNF-α is a key proinflammatory factor during the inflammatory response that can activate various inflammatory cells, further accelerating the cascading release of other inflammatory mediators and greatly increasing the degree of the inflammatory response.20 IL-1β receptors mainly exist in the central and peripheral nervous systems, and the interaction between IL-1β and its receptors induces a variety of responses, which are mainly intercellular responses mediated by IL-6. Moreover, IL-1β is closely related to COX-2 and type II phospholipase A2.21 IL-6 can induce glial cell activation and participate in the maintenance of NP.22 In this study, the proinflammatory cytokines IL-1β, IL-6 and TNF-α were selected for examination by ELISA. The results of this study showed that the levels of IL-1β, IL-6 and TNF-α in the spinal cord on the injured side in CCI mice were significantly higher than those in normal mice. Z-guggulsterone had no effect on the levels of IL-1β, IL-6 and TNF-α in the spinal cord of normal mice. Z-guggulsterone decreased the levels of IL-1β, IL-6 and TNF-α in the spinal cord on the injured side in CCI mice in a dose-dependent manner. This study indicated that Z-guggulsterone inhibited the expression of proinflammatory cytokines TNF-α, IL-1β and IL-6 in the spinal cords of CCI mice, thereby relieving neuropathic pain.
In recent years, spinal astrocytes have been favoured by pain researchers as an important target. Studies have shown that activated astrocytes produce numerous mediators, such as proinflammatory cytokines, which result in the activation of a cascade in the surrounding glia that enhances neuronal activity. Furthermore, these proinflammatory cytokines released by activated astrocytes play an important role in the development of neuropathic pain following peripheral nerve injury.23,24 A previous study showed that during NP, astrocytes in the spinal dorsal horn are activated, and the level of GFAP, which can specifically label astrocytes, is significantly increased. This causes the release of a large number of proinflammatory cytokines, such as IL-1β, IL-6 and TNF-α, through a series of signalling pathways, and the invasion of nearby neurons or microglia, finally resulting in a state of pain hypersensitivity.25 Therefore, whether the mechanism by which Z-guggulsterone alleviates neuropathic pain in CCI mice is related to the inhibition of spinal astrocyte activation is a key question. Our results showed that the expression level of GFAP was significantly higher in the spinal dorsal horn on the injured side in CCI mice than in normal mice. Z-guggulsterone had no effect on the expression level of GFAP in the spinal dorsal horn of normal mice. Z-guggulsterone dose-dependently reduced the expression of GFAP in the spinal dorsal horn on the injured side in CCI mice. Therefore, these results suggest that Z-guggulsterone alleviates NP in CCI mice by inhibiting the activation of astrocytes in the spinal dorsal horn.
Increased astrocytes activation and levels of IL-1β, IL-6 and TNF-α were detected in the spinal dorsal horn of CCI mice. Conversely, Z-guggulsterone injection decreased astrocytes activation and the levels of these proinflammatory cytokines in the spinal dorsal horn, thus exerting an analgesic effect. Although more in-depth research is needed in the future to explore the relationship between Z-guggulsterone and these pain-related cytokines by multiple interventions, the current study may reveal effective analgesic properties and promising therapeutic targets for Z-guggulsterone to attenuate NP.
Conclusion
Z-guggulsterone can relieve neurological pain in CCI mice, and its analgesic effect is related to inhibiting the expression of GFAP and the proinflammatory cytokines IL-1β, IL-6 and TNF-α in the spinal dorsal horn. This study provides a novel method to treat neuropathological pain and a reliable theoretical basis for the further application of Z-guggulsterone in clinical medicine.
Data Sharing Statement
The authors will allow the sharing of participant data. The data will be available for anyone who wishes to access for any purpose. The data will be accessible from immediately following publication to 6 months after publication, and contact should be made to the first author by email ([email protected]).
Author Contributions
All authors made a significant contribution to the work reported, whether in the conception, study design, execution, acquisition of data, analysis and interpretation, or all of these areas. All authors took part in drafting, revising or critically reviewing the article, gave final approval of the version to be published, have agreed on the journal to which the article has been submitted, and agree to be accountable for all aspects of this work.
Funding
This work was supported by the Scientific Research Project of Shaanxi Provincial Administration of Traditional Chinese Medicine (no. 2021-GJ-JC003).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Finnerup NB, Kuner R, Jensen TS. Neuropathic Pain: from Mechanisms to Treatment. Physiol Rev. 2021;101(1):259–301.
2. Witt EA, Kenworthy J, Isherwood G, et al. Examining the association between pain severity and quality-of-life, work-productivity loss, and healthcare resource use among European adults diagnosed with pain. J Med Econ. 2016;19(9):858–865.
3. Sommer C, Leinders M, Üçeyler N. Inflammation in the pathophysiology of neuropathic pain. Pain. 2018;159(3):595–602.
4. Peirs C, Williams SG, Zhao X, et al. Mechanical Allodynia Circuitry in the Dorsal Horn Is Defined by the Nature of the Injury. Neuron. 2021;109(1):73–90 e77.
5. Old EA, Clark AK, Malcangio M. The role of glia in the spinal cord in neuropathic and inflammatory pain. Handb Exp Pharmacol. 2015;227:145–170.
6. Gwak YS, Hulsebosch CE, Leem JW. Neuronal-Glial Interactions Maintain Chronic Neuropathic Pain after Spinal Cord Injury. Neural Plast. 2017;2017:2480689.
7. Ueda H, Neyama H, Nagai J, et al. Involvement of lysophosphatidic acid-induced astrocyte activation underlying the maintenance of partial sciatic nerve injury-induced neuropathic pain. Pain. 2018;159(11):2170–2178.
8. Clark AK, Old EA, Malcangio M. Neuropathic pain and cytokines: current perspectives. J Pain Res. 2013;6:803–814.
9. Zhang EQ. Bi Syndrome (Arthralgia Syndrome). J Tradit Chin Med. 2010;30(2):145–152.
10. Li SH, Li L, Yang RN, et al. Compounds of traditional Chinese medicine and neuropathic pain. Chin J Nat Med. 2020;18(1):28–35.
11. Abdul-Ghani RA, Loutfy N, Hassan A. Myrrh and trematodoses in Egypt: an overview of safety, efficacy and effectiveness profiles. Parasitol Int. 2009;58(3):210–214.
12. Yamada T, Sugimoto K. Guggulsterone and Its Role in Chronic Diseases. Adv Exp Med Biol. 2016;929:329–361.
13. Ahmad MA, Mujeeb M, Akhtar M, et al. Guggulipid: a Promising Multi-Purpose Herbal Medicinal Agent. Drug Res. 2020;70(4):123–130.
14. Liu T, Liu M, Zhang T, et al. Z-Guggulsterone attenuates astrocytes-mediated neuroinflammation after ischemia by inhibiting toll-like receptor 4 pathway. J Neurochem. 2018;147(6):803–815.
15. Liu FG, Hu WF, Wang JL, et al. Z-Guggulsterone Produces Antidepressant-Like Effects in Mice through Activation of the BDNF Signaling Pathway. Int J Neuropsychopharmacol. 2017;20(6):485–497.
16. Huang C, Wang J, Lu X, et al. Z-guggulsterone negatively controls microglia-mediated neuroinflammation via blocking IκB-α-NF-κB signals. Neurosci Lett. 2016;619:34–42.
17. Challa SR. Surgical animal models of neuropathic pain: pros and Cons. Int J Neurosci. 2015;125(3):170–174.
18. Zhang W, Suo M, Yu G, et al. Antinociceptive and anti-inflammatory effects of cryptotanshinone through PI3K/Akt signaling pathway in a rat model of neuropathic pain. Chem Biol Interact. 2019;305:127–133.
19. Al-Mazidi S, Alotaibi M, Nedjadi T, et al. Blocking of cytokines signalling attenuates evoked and spontaneous neuropathic pain behaviours in the paclitaxel rat model of chemotherapy-induced neuropathy. Eur J Pain. 2018;22(4):810–821.
20. Leung L, Cahill CM. TNF-alpha and neuropathic pain–a review. J Neuroinflammation. 2010;7:27.
21. Del Rey A, Apkarian AV, Martina M, et al. Chronic neuropathic pain-like behavior and brain-borne IL-1β. Ann N Y Acad Sci. 2012;1262(1):101–107.
22. Hung AL, Lim M, Doshi TL. Targeting cytokines for treatment of neuropathic pain. Scand J Pain. 2017;17:287–293.
23. Donnelly CR, Andriessen AS, Chen G, et al. Central Nervous System Targets: glial Cell Mechanisms in Chronic Pain. Neurotherapeutics. 2020;17(3):846–860.
24. Lee JH, Kim W. The Role of Satellite Glial Cells, Astrocytes, and Microglia in Oxaliplatin-Induced Neuropathic Pain. Biomedicines. 2020;8(9):545.
25. Oh JY, Hwang TY, Jang JH, et al. Muscovite nanoparticles mitigate neuropathic pain by modulating the inflammatory response and neuroglial activation in the spinal cord. Neural Regen Res. 2020;15(11):2162–2168.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.