")
Back to Journals » Infection and Drug Resistance » Volume 17
Whole-Genome Sequencing of an Escherichia coli ST69 Strain Harboring blaCTX-M-27 on a Hybrid Plasmid
Authors Wang L, Guan Y, Lin X , Wei J, Zhang Q, Zhang L, Tan J, Jiang J, Ling C, Cai L, Li X , Liang X, Wei W, Li RM
Received 17 August 2023
Accepted for publication 29 December 2023
Published 1 February 2024 Volume 2024:17 Pages 365—375
DOI https://doi.org/10.2147/IDR.S427571
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Sandip Patil
Ling Wang,1,* Yuee Guan,2,* Xu Lin,3,* Jie Wei,4 Qinghuan Zhang,4 Limei Zhang,5 Jing Tan,1 Jie Jiang,1 Caiqin Ling,1 Lei Cai,1 Xiaobin Li,5 Xiong Liang,1 Wei Wei,6 Rui-Man Li7
1Department of Obstetrics, Zhuhai Hospital Affiliated with Jinan University (Zhuhai People’s Hospital), Zhuhai, 519000, People’s Republic of China; 2Department of Cardiology, Zhuhai Hospital Affiliated with Jinan University (Zhuhai People’s Hospital), Zhuhai, 519000, People’s Republic of China; 3Department of Gastrointestinal Surgery, Zhuhai Hospital Affiliated with Jinan University (Zhuhai People’s Hospital), Zhuhai, 519000, People’s Republic of China; 4Department of Clinical Laboratory, Zhuhai Hospital Affiliated with Jinan University (Zhuhai People’s Hospital), Zhuhai, 519000, People’s Republic of China; 5Guangdong Provincial Key Laboratory of Tumor Interventional Diagnosis and Treatment, Zhuhai Hospital Affiliated with Jinan University (Zhuhai People’s Hospital), Zhuhai, 519000, People’s Republic of China; 6Department of Cardiothoracic Surgery, Zhuhai Hospital Affiliated with Jinan University (Zhuhai People’s Hospital), Zhuhai, 519000, People’s Republic of China; 7Department of Gynaecology and Obstetrics, The First Affiliated Hospital of Jinan University, Guangzhou, 510630, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Rui-Man Li; Wei Wei, Email [email protected]; [email protected]
Objective: Escherichia coli is a common Gram-negative human pathogen. The emergence of E. coli with multiple-antibiotic-resistant phenotypes has become a serious health concern. This study reports the whole-genome sequences of third-generation cephalosporin-resistant (3GC-R) and multidrug-resistant (MDR) E. coli EC6868 and explores the acquired antibiotic-resistance genes (ARGs) as well as their genetic contexts.
Methods: E. coli EC6868 was isolated from a vaginal secretion sample of a pregnant patient in China. The antimicrobial susceptibility was assessed, and whole-genome sequencing was conducted. The acquired ARGs, insertion sequence (IS) elements, and integrons within the genome of E. coli EC6868 were identified, and the genetic contexts associated with the ARGs were analyzed systematically.
Results: E. coli EC6868 was determined to belong to ST69 and harbored a 144.9-kb IncF plasmid (pEC6868-1) with three replicons (Col156, IncFIBAP001918, and IncFII). The ESBL gene blaCTX-M-27 was located on the structure “∆ISEcp1-blaCTX-M-27-IS 903B”, which was widely present in the species of Enterobacteriales. Other ARGs carried by plasmid pEC6868-1 were mainly located on the 18.9-kb IS 26-composite transposon (five copies of intact IS 26 and one copy of truncated IS 26) composing of IS 26-mphA-mrx(A)-mphR(A)-IS 6100, ∆TnAs3-eamA-tet(A)-tetR(A)-aph(6)-Id-aph(3”)-Ib-sul2-IS 26, and a class 1 integron, which was widely present on IncF plasmids of E. coli, mainly distributed in ST131, ST38, and ST405. Notably, pEC6868 in our study was the first report on a plasmid harboring the 18.9-kb structure in E. coli ST69 in China.
Conclusion: The 3GC-R E. coli ST69 strain with an MDR IncF plasmid carrying blaCTX-M-27 and other ARGs, conferring resistance to aminoglycosides, macrolides, sulfonamides, tetracycline, and trimethoprim, was identified in a hospital in China. Mobile genetic elements including ISEcp1, IS 903B, IS 26, Tn 3, IS 6100 and class 1 integron were found within the MDR region, which could play important roles in the global dissemination of these resistance genes.
Keywords: Escherichia coli, vaginal secretion, plasmid, blaCTX-M-27, genetic context
Introduction
Both Gram-negative and Gram-positive bacteria are responsible for urogenital tract infections. However, among Gram-negative bacterial agents, Escherichia coli is the most common causative agent.1,2 Based on data from the China Antimicrobial Surveillance Network (CHINET), in 2022, E. coli had the highest detection rate, comprising 18.97% of clinical isolates. Uropathogenic E. coli strains were recognized as typical bacterial agents for urinary tract infections, these groups of infectious diseases are more common among women rather than men, as well as female reproductive tract.3,4 In 2021, a meta-analysis of 82 studies, with a sample size of 33,118, indicated that the prevalence of E. coli in vaginal samples from pregnant women is 32% and that of extended-spectrum-lactamase (ESBL)-producing E. coli in pregnant women is 34%.5 Perinatal infections caused by E. coli can result in premature birth, a low birth weight, and an increased risk of neonatal infection and death.6,7 Notably, due to the misuse or overuse of antibiotics, antimicrobial resistance (AMR) is considered to feature among the top 10 threats to global health.8 The emergence and global spread of antibiotic-resistant E. coli have caused difficulties in clinical treatment and have become a public health threat.9 Third-generation cephalosporins are the most commonly used class of antibiotics in gynecology/obstetrics and pediatrics departments to treat infections caused by E. coli;10,11 however, the prevalence of E. coli resistant to third-generation cephalosporins is increasing.12
Enterobacteriales bear different antibiotic resistance genes encoding Extended-Spectrum-ß-Lactamases (ESBLs) and Metallo-ß-Lactamases (MBLs) on their chromosomes and mobile genetic elements such as plasmids and transposons.13 In E. coli (typical representative of Enterobacteriales), ESBLs mainly confer third-generation cephalosporin resistance.14 TEM- and SHV-type ESBLs were initially described in the 1980s.15,16 The first CTX-M-type ESBL (CTX-M-1), with higher hydrolytic activity against cefotaxime than against ceftazidime, emerged in 1995.17 Since then, several variants of CTX-M-1 have been found,18 and these are now the most prevalent ESBLs all over the world, with E. coli as their major bacterial host.19
Mobile genetic elements (eg plasmid, insertion sequence (IS), transposon, and integron) play a major role in facilitating the spread of resistance genes.20 ESBL genes are mainly located on plasmids, and among the ESBL-carrying plasmids from E. coli, the most common replicon types are IncF, IncI1, IncN, IncHI1, and IncHI2.21 ESBL genes are always associated with many ISs (such as IS26, ISCR1, ISEcp1, and IS10), transposons (such as Tn2), and integrons.22–24 Here, we report a multidrug resistant (MDR) E. coli ST69 strain carrying an IncF plasmid harboring blaCTX-M-27 and other antibiotic resistance genes (ARGs) and present a comparative analysis of the MDR region of the IncF plasmid.
Materials and Methods
For this study, the data collection and analysis are shown in Figure S1.
Bacterial Strains and Identification
The strain EC6868 was isolated from the vaginal secretions of a 39-year-old pregnant woman at the Zhuhai People’s Hospital in December 2021. Species identification and antimicrobial susceptibility testing were conducted using a fully automatic VITEK 2 COMPACT system (BioMérieux, France) according to the manufacturer’s instructions. The results of antimicrobial sensitivity were interpreted based on the Clinical and Laboratory Standards Institute guidelines (M100-S24). The identity of EC6868 was further confirmed through 16S rRNA gene sequencing by using bacterial universal primers (27F/1492R).
Whole-Genome Sequencing, Assembly, and Annotation
Whole-genome sequencing of strain EC6868 was conducted by Genewiz Biotechnology Co. Ltd. (Suzhou, China) by using the Illumina paired-end sequencing and PacBio long reads sequencing. PacBio reads were assembled using HGAP4.0/Falcon 0.3 of WGS-Assembler 8.2.25 The genome assembly was then polished with the software Pilon 1.2226 using Illumina reads. The assembled genome (both chromosome and plasmids) of strain EC6868 was submitted to the GenBank database27 and annotated using the NCBI Prokaryotic Annotation Pipeline (PGAP).28
Bioinformatics Analysis of the Genome of Strain EC6868
Acquired ARGs in the genome of strain EC6868 were identified using the websever ResFinder 4.1,29 with a minimum coverage of 60% and a minimum identity of 90%. Multilocus sequence typing (MLST) of strain EC6868 was performed using MLST 2.0,30 selecting the database as “Escherichia coli #1” and using seven housekeeping genes, including adk, fumC, gyrB, icd, mdh, purA, and recA. Replicon types of the plasmids contained by strain EC6868 were determined using PlasmidFinder 2.1,31 with the database “Enterobacteriales”, minimum coverage of 60%, and minimum identity of 95%. The ISs adjacent to the ARGs in the EC6868 genome were identified using ISfinder.32 Sequence similarity search was performed using MegaBLAST33 scans against the GenBank nr database. BLAST Ring Image Generator (BRIG) 0.9534 and Easyfig2.2.535 were used for plasmid comparisons.
Nucleotide Sequence Accession Numbers
The genome sequence of strain EC6868, which contained a chromosome and two plasmids, was submitted to GenBank under the accession numbers CP095083–CP095085.
Results
Identification and Antimicrobial Susceptibility Testing of Strain EC6868
Strain EC6868 isolated from vaginal secretions of a pregnant woman, was identified as E. coli by the automatic VITEK 2 COMPACT system, which was then confirmed via 16SrRNA gene sequencing. E. coli strain EC6868 showed resistance to cephalosporins, including cefuroxime, cefuroxime axetil, and ceftriaxone, but conferred intermediate-level ceftazidime resistance (Table S1). In addition, it showed resistance to trimethoprim/sulfamethoxazole and intermediate-level resistance to levofloxacin (Table S1).
Genomic Features of the E. coli Strain EC6868
Genomic analysis revealed that the genome of E. coli strain EC6868 comprised a 5.25-Mb chromosome and two plasmids with sizes of 144,934 bp (pEC6868-1) and 7919 bp (pEC6868-2). MLST analysis indicated that E. coli strain EC6868 belonged to sequence type (ST) 69. PlasmidFinder results indicated that plasmid pEC6868-1 contained three replicons (Col156, IncFIBAP001918, and IncFII), whereas plasmid pEC6868-2 was an untypeable plasmid.
ResFinder results indicated that E. coli EC6868 carried multiple ARGs located on both chromosomes and the plasmid pEC6868-1. The chromosome of E. coli EC6868 was found to harbor disinfectant-resistance genes (sitABCD). Further, the multidrug-resistance plasmid pEC6868-1 harbored ARGs conferring resistance to extended-spectrum cephalosporins (blaCTX-M-27), aminoglycosides (aph(6)-Id, aph(3”)-Ib, and aadA5), macrolides (erm(B) and mph(A)-mrx(A)-mphR(A)), sulphonamides (sul1 and sul2), tetracycline (tet(A)-tetR(A)), and trimethoprim (dfrA17). Plasmid pEC6868-1 also harbored the antiseptic-resistance gene ΔqacE.
Comparative Analysis of the MDR Region of Plasmid pEC6868-1
The ARGs, as well as the adjacent IS elements carried by plasmid pEC6868-1, were located on the ~31.2-kb fragment of the plasmid, forming a large MDR region. The MDR region of pEC6868-1, flanked by ∆IS1R and ∆IS1R-∆IS26, which was nearly identical to part of the E. coli plasmid pB16EC0698-3 (100.00% coverage and 99.78% identity; Figure 1). In the MDR region of pEC6868-1, IS26 was found to be located upstream of the macrolide-resistance gene ermB and its leading peptide gene ermB(L) (Figure 1). The ESBL gene blaCTX-M-27 was located on an ISEcp1-mediated transposition unit, with a truncated ISEcp1 (∆ISEcp1) located upstream of blaCTX-M-27 and IS903B located downstream of blaCTX-M-27, forming the genetic structure ∆ISEcp1-blaCTX-M-27-IS903B (Figure 1). Based on the region ∆ISEcp1-blaCTX-M-27-IS903B, the BLAST search hit from the database of GenBank (on November 16th, 2023), with the minimum coverage of 100% and minimum identity of 99%, showed that 1109 strains contain the structure (Figure 2). The structure was widely present in the species of Enterobacteriales, including Klebsiella pneumoniae (549), E. coli (347), Salmonella enterica (106), Proteus mirabilis (44) and the other 23 species (63) (Figures 2 and S2).
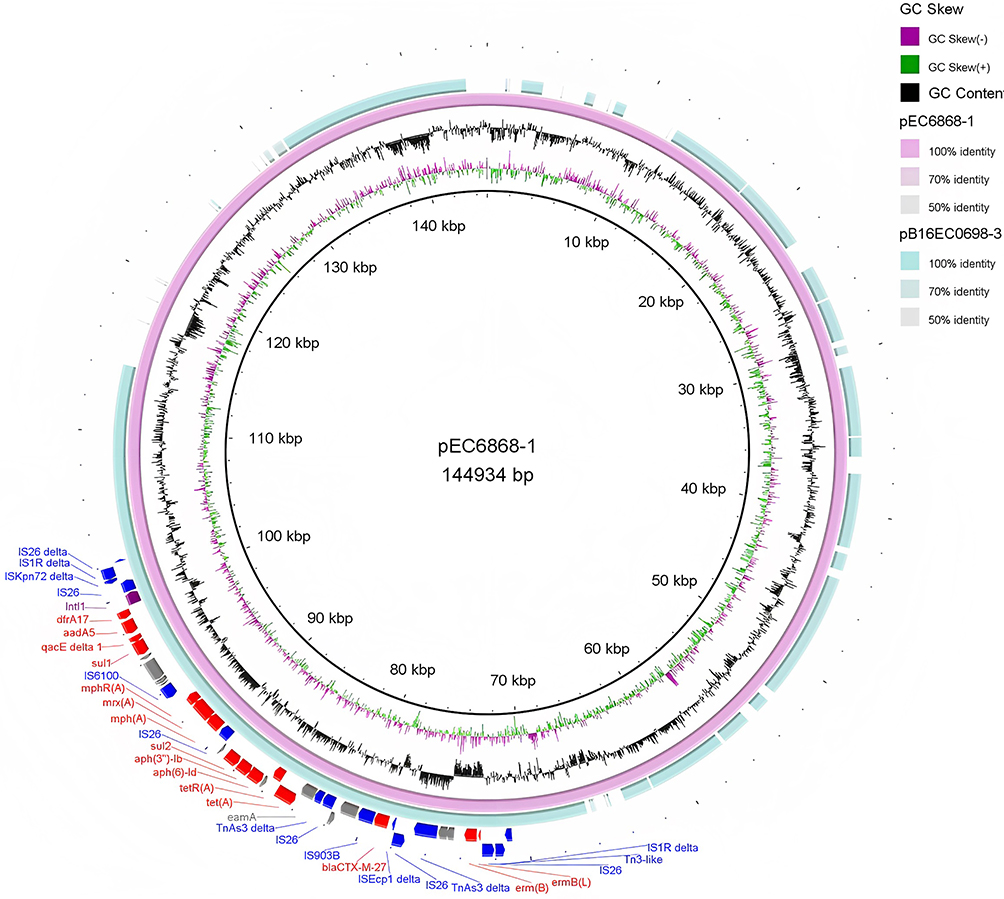 |
Figure 1 Comparative analysis of multidrug-resistance (MDR) region of plasmid pEC6868-1 and that of Escherichia coli plasmid pB16EC0698-3 generated using BRIG. Resistance, transposase, and integrase genes are shown in red, blue, and purple, respectively. |
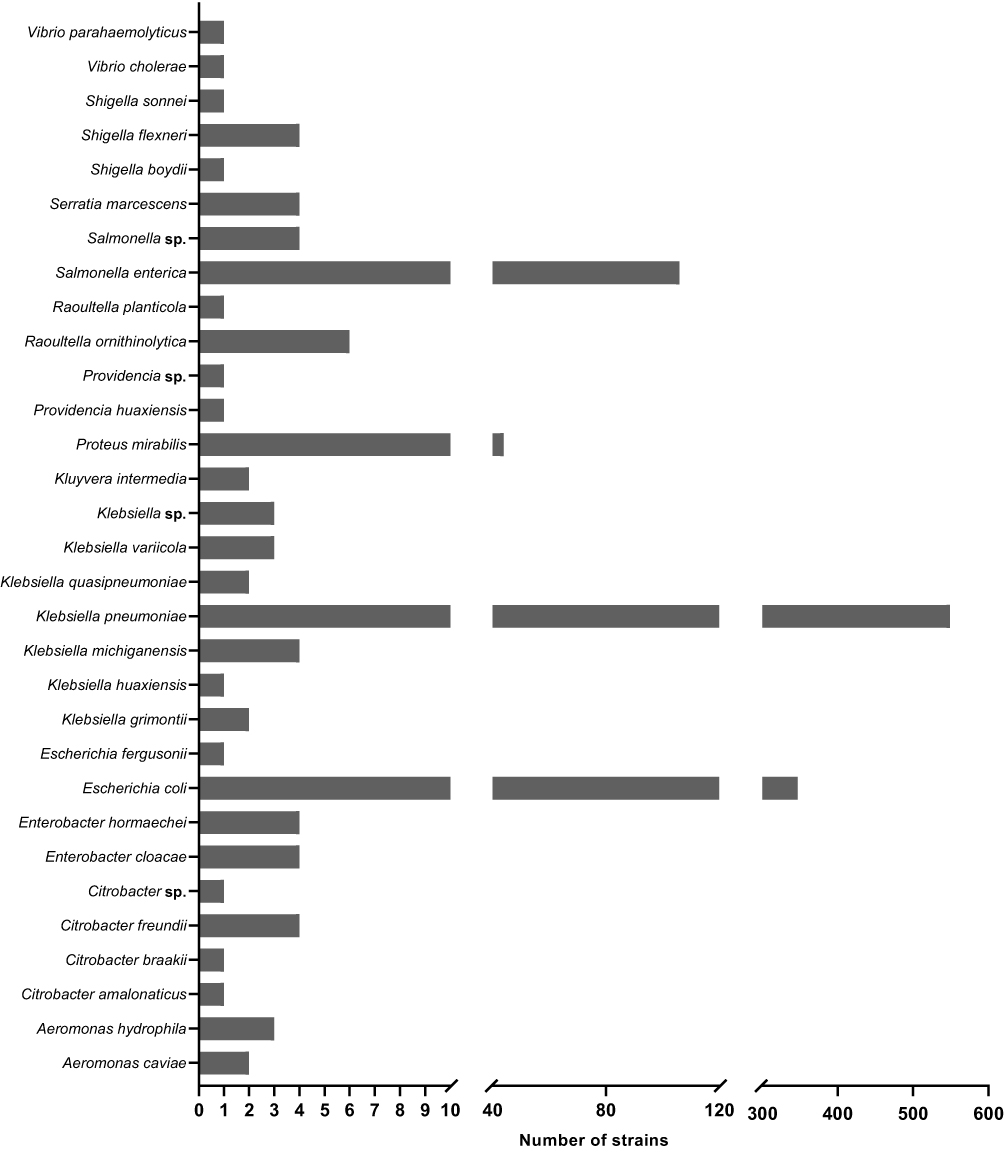 |
Figure 2 An overview of the species of Enterobacteriales harboring the structure “∆ISEcp1-blaCTX-M-27-IS903B.”. |
Interestingly, except for ermB-ermB(L) and blaCTX-M-27, other ARGs carried by plasmid pEC6868-1 were located on the 18.9-kb IS26-composite transposon. Overall, the 18.9-kb IS26-composite transposon was composed of three parts as follows: (1) the mphA-mrx(A)-mphR(A) operon conferring macrolide resistance was flanked by IS26 and IS6100, forming the IS26-mphA-mrx(A)-mphR(A)-IS6100 transposable structure (Figure 1); (2) one fragment of pEC6868-1 containing tet(A)-tetR(A), aph(6)-Id, aph(3”)-Ib, and sul2 located adjacent to IS26-∆TnAs3-eamA and IS26-mphA-mrx(A)-mphR(A)-IS6100 (Figure 1); (3) notably, a complex class 1 integron, including intI1, dfrA17, aadA5, ∆qacE, and sul1, was also found in the MDR region of plasmid pEC6868-1, located adjacent to IS26-∆ISKpn72-∆IS1R-∆IS26 (Figure 1).
Based on the 18.9-kb IS26-composite transposon of plasmid pEC6868-1, the BLAST searches against the nr database of GenBank showed that the 18.9-kb region was widely present on plasmids of E. coli (77 plasmids), mainly distributed in ST131, ST38, and ST405 (Figure 3). Notably, pEC6868 in our study was the first report on a plasmid harboring the 18.9-kb IS26-composite transposon in E. coli ST69 in China. Another ST69 strain of E. coli containing the 18.9-kb structure was found in the United States, 2017 (Table S2). Surprisingly, 75 of the 77 plasmids harboring the 18.9-kb IS26-composite transposon were found to be the IncF plasmids, including IncFIA, IncFIB and IncFII subtypes, accounting for 97.4% of all the 18.9-kb structure-harboring plasmids of E. coli. In addition, four plasmids harboring the 18.9-kb IS26-composite transposon were also found in K. pneumoniae (Figures 3 and S2).
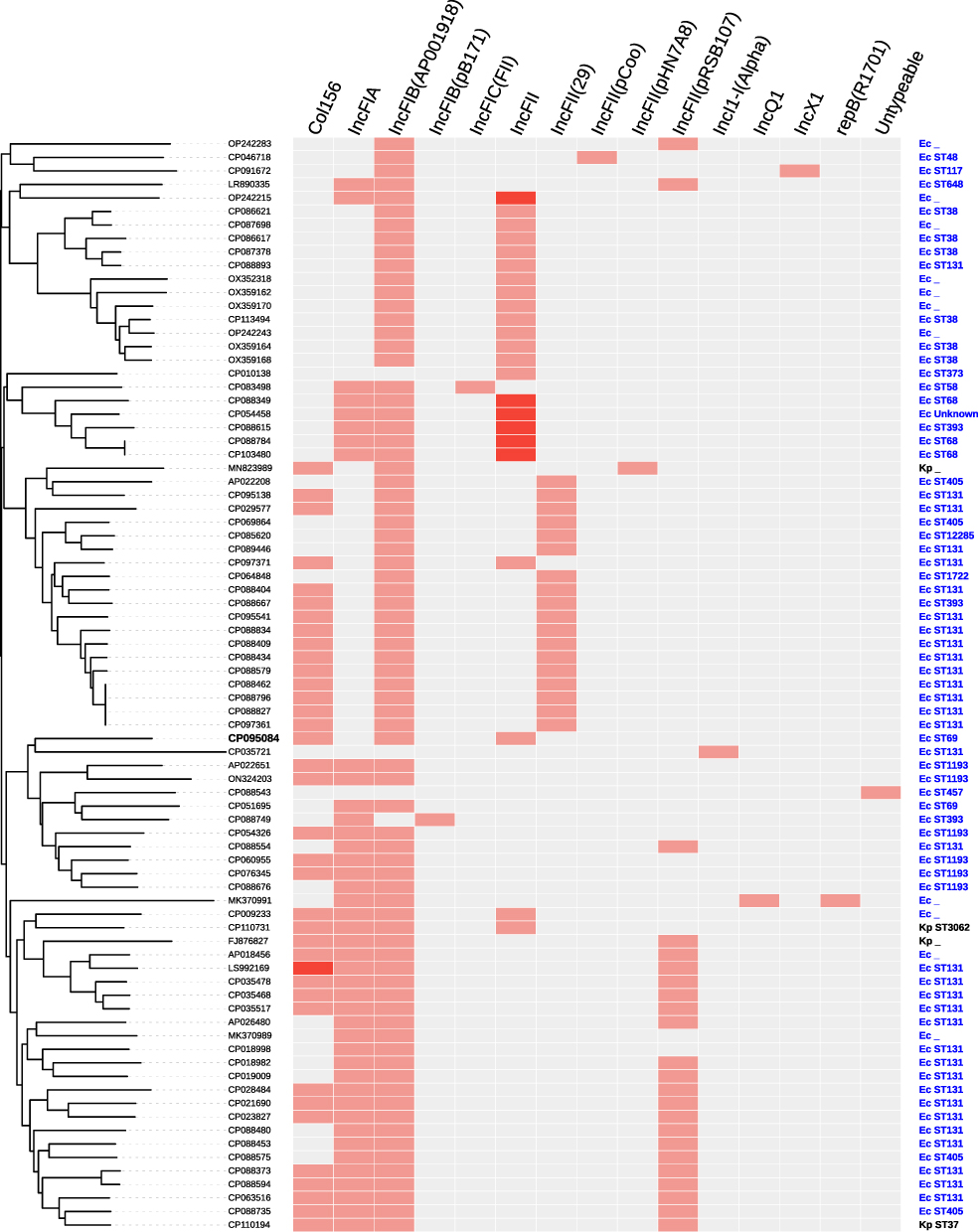 |
Figure 3 Details of replicon types and STs of host strains of the 81 plasmids harboring 18.9-kb IS26-composite transposon. Phylogenetic cladogram based on the presence/absence of orthologous gene families of the 81 plasmids harboring 18.9-kb IS26-composite transposon were constructed. Ec and Kp represent E. coli and K. pneumoniae, respectively. |
Moreover, we found that the region within this 18.9 kb IS26-composite transposon, containing IS26-mphA-mrx(A)-mphR(A)-IS6100 and the complex class 1 integron, was not only present on the plasmids but also on the chromosomes of some pathogens, such as E. coli, K. pneumoniae, Klebsiella michiganensis, Proteus mirabilis, Shigella flexneri, and Morganella morganii (Figure 4).
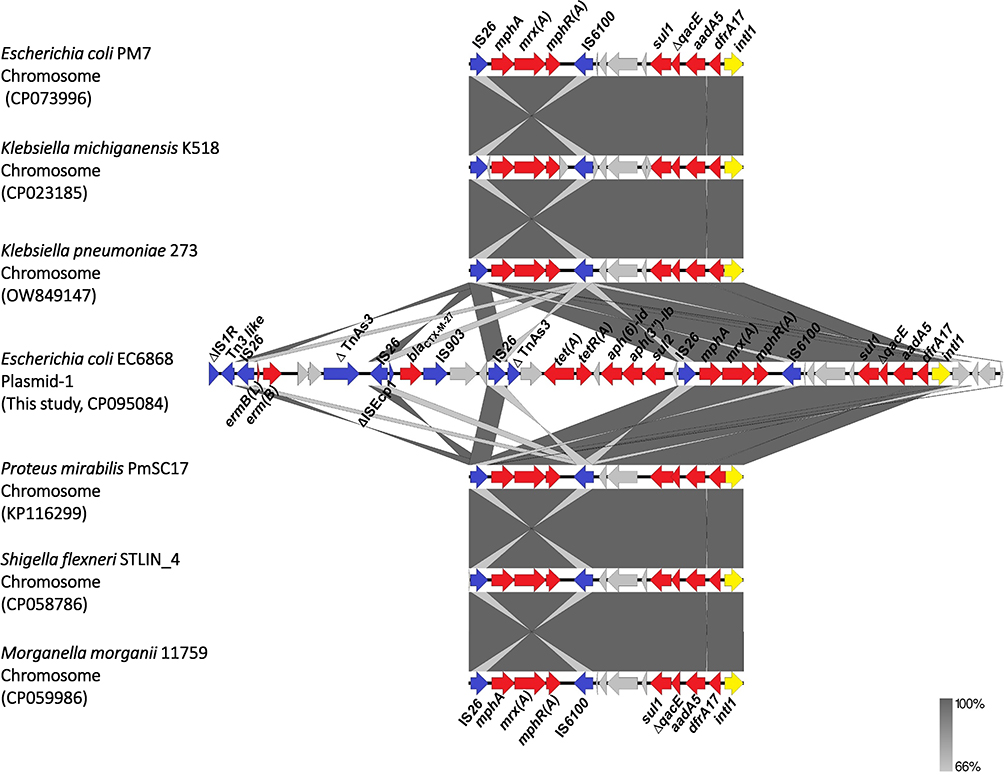 |
Figure 4 Comparison of the gene cluster containing IS26-mphA-mrx(A)-mphR(A)-IS6100 and the complex class 1 integron carried by plasmid pEC6868-1with chromosomes of other pathogenic bacteria other than Escherichia coli. Resistance, transposase, and integrase genes are shown in red, blue, and yellow, respectively. |
Discussion
In this study, we report a third-generation cephalosporin-resistant strain, E. coli ST69 (EC6868), isolated from the vaginal secretions of a 39-year-old pregnant woman. E. coli ST69 was determined to be one of the multidrug resistant E. coli clones of phylogenetic group D, which are widespread among different hosts, often causing urinary tract infections and exhibiting resistance to antibiotics.36–38 ST69 E. coli strains have been reported to carry the blaVIM-harboring IncA plasmid,39 blaNDM-1-harboring IncI1 plasmid,40 and mcr-1-harboring IncHI2 plasmid.41 In this work, we report that the ST69 E. coli strain carries one hybrid plasmid, with three replicons (Col156, IncFIBAP001918, and IncFII), harboring blaCTX-M-27. IncF plasmids, comprising one of the most common incompatibility types, have been identified globally in Enterobacteriales. Moreover, they vary in size (50–200 kb) and replicon types, which harbor huge amount of ARGs and confer resistance to all major classes of antibiotics, such as beta-lactams, chloramphenicol, aminoglycosides, quinolones, and tetracyclines.42–45 Notably, the IncF-blaCTX-M association found in E. coli EC6868 in our study has been widely observed in E. coli isolates of both human and animal origins; for example, the IncF plasmid R100 is responsible for the spread of blaCTX-M-14 in the UK and France.46,47
Strain EC6868 in our study was found to be a CTX-M-type ESBL-producing E. coli isolate. CTX-M-14 and CTX-M-15 are the most common variants of CTX-M, and CTX-M-27 has increased rapidly in terms of its prevalence.48 Notably, the detection of CTX-M-27 in E. coli isolates from patient has been rising and is particularly alarming due to its presence in clonal groups including ST10, ST69, and ST131.49,50 The ∆ISEcp1 marker was located upstream of blaCTX-M-27 and carried by pEC6868-1 (ΔISEcp1-blaCTX-M-27-IS903B) was found in another study, which points that the conserved region responsible for transmission of blaCTX-M-27 was ISEcp1-blaCTX-M-27-IS903B.51 ISEcp1 belongs to the IS1380 family,52 and the ISEcp1-like element is associated with several blaCTX-M gene types in Enterobacteriaceae, including blaCTX-M-14, blaCTX-M-24, blaCTX-M-22, and blaCTX-M-79.53 IS903B, located downstream of blaCTX-M-27 carried by pEC6868-1, is a small insertion element of 1057 bp that is transposed predominantly via a conservative “cut-and-paste” mechanism.54
Five copies of intact IS26 and one ∆IS26 were found within the MDR region of plasmid pEC6868-1. The 820-bp IS26 was first described in 198355 and was determined to be present in numerous early antibiotic-resistant bacteria, playing a critical role in disseminating ARGs among Gram-negative bacteria.56 IS26 is frequently associated with genes encoding antibiotic resistance factors,57 and it can contribute to the expression of the ARGs by supplying a promoter −35 region which can be coupled with a −10 region in the adjacent ARG.58
In this study, two translocatable units conferring macrolide resistance, IS26-ermB(L)-erm(B) and IS26-mphA-mrx(A)-mphR(A)-IS6100, were detected in plasmid pEC6868-1. In IS26-ermB(L)-erm(B), the ermB encodes a ribosomal methylase which can reduce the affinity of macrolides for the ribosome, resulting in high level of macrolide resistance.59 Macrolide antibiotics promote ribosome stalling on the ErmB(L) (regulatory leader peptide), ultimately inducing the expression of ermB.60 In the gene cluster mphA-mrx(A)-mphR(A) for high-level macrolide inactivation, mphA encodes a phosphotransferase that inactivates erythromycin and mrx(A) encodes a protein required for expression of MphA, whereas MphR(A) encoded by mphR(A) negatively regulates the expression of mphA at the transcriptional level.61 The IS26-mph(A)-mrx(A)-mph(R)(A)-IS6100 unit, which is linked to the global dissemination of macrolide-resistance genes,62 is the most common form in plasmids carrying mphA.63
Class 1 integrons are prevalent genetic elements that play a key role in the spread of antibiotic resistance,64 allowing bacteria to capture and exchange ARGs embedded in the gene cassettes. Moreover, acquisition of gene cassettes is catalyzed by integrase encoded by int1, which is a site-specific recombinase.65 Class 1 integrons are structurally composed of three core elements, specifically an integrase gene (intI1), a primary recombination site (attI1), and a common promoter enabling the transcription of gene cassettes.66 In all integrons, detection rate of class 1 integrons in clinical isolates is highest at 50–70%, which is linked to the production and spread of antibiotic-resistant bacteria.67,68
Conclusion
Here, we describe the genomic characteristics of a 3GC-R and ESBL-producing E. coli strain EC6868 belonging to ST69 harboring blaCTX-M-27 isolated from a pregnant patient in China. The strain carried multiple-resistance genes conferring resistance to aminoglycosides, macrolides, sulfonamides, tetracycline, and trimethoprim. The blaCTX-M-27 gene located on a hybrid plasmid pEC6868-1 (IncFII/IncFIB/Col156). The ESBL gene blaCTX-M-27 was flanked by the truncated ISEcp1 (∆ISEcp1) and the IS903B (∆ISEcp1-blaCTX-M-27-IS903B), which was widely present in the species of Enterobacteriales, especially in K. pneumoniae, E. coli, and Salmonella enterica. Except for blaCTX-M-27, other ARGs carried by plasmid pEC6868-1 were mainly located on the 18.9-kb IS26-composite transposon (five copies of intact IS26 and one copy of truncated IS26). The 18.9-kb structure was widely present on IncF plasmids of E. coli, mainly distributed in ST131, ST38, and ST405. Notably, pEC6868 in our study was the first report on a plasmid harboring the 18.9-kb structure in E. coli ST69 in China. In addition, Tn3, IS6100 and class 1 integron also play important role in the dissemination of acquired ARGs carried by the plasmid pEC6868-1 of E. coli strain EC6868.
Ethical Approval Statement
This study has been approved by the Ethics Committee of Zhuhai People’s Hospital. The present study was a study focusing on bacteria and did not contain any sensitive personal information. Therefore, informed consent was not required according to “Measures for the Ethical Review of Biomedical Research Involving Humans” (https://www.gov.cn/gongbao/content/2017/content_5227817.htm).
Funding
This work was supported financially by the grants from the Science and Technology Projects of Social Development in Zhuhai (Grant No. 2220004000129).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Behzadi P, García-Perdomo HA, Autrán Gómez AM, et al. Editorial: uropathogens, urinary tract infections, the host-pathogen interactions and treatment. Front Microbiol. 2023;14:1183236. doi:10.3389/fmicb.2023.1183236
2. Paitan Y. Current Trends in Antimicrobial Resistance of Escherichia coli. Curr Top Microbiol Immunol. 2018;416:181–211. doi:10.1007/82_2018_110
3. Issakhanian L, Behzadi PJCPD. Antimicrobial agents and urinary tract infections. Current Pharmaceutical Design. 2019;25(12):1409–1423. doi:10.2174/1381612825999190619130216
4. Behzadi PJFM. Classical chaperone-usher (CU) adhesive fimbriome: uropathogenic Escherichia coli (UPEC) and urinary tract infections (UTIs). Int J Med. 2020;65(1):45–65.
5. Moradi Y, Eshrati B, Motevalian SA, et al. A systematic review and meta-analysis on the prevalence of Escherichia coli and extended-spectrum β-lactamase-producing Escherichia coli in pregnant women. Arch Gynecol Obstet. 2021;303(2):363–379. doi:10.1007/s00404-020-05903-w
6. Denkel LA, Schwab F, Kola A, et al. The mother as most important risk factor for colonization of very low birth weight (VLBW) infants with extended-spectrum β-lactamase-producing Enterobacteriaceae (ESBL-E). J Antimicrob Chemother. 2014;69(8):2230–2237. doi:10.1093/jac/dku097
7. Page JM, Bardsley T, Thorsten V, et al. Stillbirth Associated With Infection in a Diverse U.S. Cohort Obstet Gynecol. 2019;134(6):1187–1196. doi:10.1097/AOG.0000000000003515
8. Algammal A, Hetta HF, Mabrok M, et al. Editorial: emerging multidrug-resistant bacterial pathogens ”superbugs”: a rising public health threat. Front Microbiol. 2023;14:1135614. doi:10.3389/fmicb.2023.1135614
9. Serwecińska LJW. Antimicrobials and antibiotic-resistant bacteria: a risk to the environment and to public health. Water. 2020;12(12):3313.
10. Ansaldi Y, Martinez de Tejada Weber B. Urinary tract infections in pregnancy. Clin Microbiol Infect. 2022;29(10):1249–1253. doi:10.1016/j.cmi.2022.08.015
11. Zhang JS, Liu G, Zhang W-S, et al. Antibiotic usage in Chinese children: a point prevalence survey. World J Pediatr. 2018;14(4):335–343. doi:10.1007/s12519-018-0176-0
12. Roer L, Hansen F, Thomsen MCF, et al. WGS-based surveillance of third-generation cephalosporin-resistant Escherichia coli from bloodstream infections in Denmark. J Antimicrob Chemother. 2017;72(7):1922–1929. doi:10.1093/jac/dkx092
13. Behzadi P, García-Perdomo HA, Karpiński TM, et al. Metallo-ß-lactamases: a review. Mol Biol Rep. 2020;47(8):6281–6294. doi:10.1007/s11033-020-05651-9
14. Paterson DL, Bonomo RA. Extended-spectrum beta-lactamases: a clinical update. Clin Microbiol Rev. 2005;18(4):657–686. doi:10.1128/CMR.18.4.657-686.2005
15. Medeiros AA. Beta-lactamases. Br Med Bull. 1984;40(1):18–27. doi:10.1093/oxfordjournals.bmb.a071942
16. Jacoby GA, Medeiros AA. More extended-spectrum beta-lactamases. Antimicrob Agents Chemother. 1991;35(9):1697–1704. doi:10.1128/AAC.35.9.1697
17. Tzouvelekis LS, Tzelepi E, Tassios PT, et al. CTX-M-type beta-lactamases: an emerging group of extended-spectrum enzymes. Int J Antimicrob Agents. 2000;14(2):137–142. doi:10.1016/S0924-8579(99)00165-X
18. Bauernfeind A, Grimm H, Schweighart S. A new plasmidic cefotaximase in a clinical isolate of Escherichia coli. Infection. 1990;18(5):294–298. doi:10.1007/BF01647010
19. Livermore DM, Canton R, Gniadkowski M, et al. CTX-M: changing the face of ESBLs in Europe. J Antimicrob Chemother. 2007;59(2):165–174. doi:10.1093/jac/dkl483
20. Partridge SR, Kwong SM, Firth N, et al. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin Microbiol Rev. 2018;31(4). doi:10.1128/CMR.00088-17
21. Carattoli A. Plasmids and the spread of resistance. Int J Med Microbiol. 2013;303(6–7):298–304. doi:10.1016/j.ijmm.2013.02.001
22. Schink AK, Kadlec K, Kaspar H, et al. Analysis of extended-spectrum-β-lactamase-producing Escherichia coli isolates collected in the GERM-Vet monitoring programme. J Antimicrob Chemother. 2013;68(8):1741–1749. doi:10.1093/jac/dkt123
23. Cantón R, González-Alba JM, Galán JC. CTX-M Enzymes: origin and Diffusion. Front Microbiol. 2012;3:110. doi:10.3389/fmicb.2012.00110
24. Poirel L, Naas T, Nordmann P. Genetic support of extended-spectrum beta-lactamases. Clin Microbiol Infect. 2008;14 Suppl 1:75–81. doi:10.1111/j.1469-0691.2007.01865.x
25. Myers EW, Sutton GG, Delcher AL, et al. A Whole-Genome Assembly of Drosophila. Science. 2000;287(5461):2196–2204. doi:10.1126/science.287.5461.2196
26. Walker BJ, Abeel T, Shea T, et al. Pilon: an integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS One. 2014;9(11):e112963. doi:10.1371/journal.pone.0112963
27. Sayers EW, Cavanaugh M, Clark K, et al. GenBank. Nucleic Acids Res. 2020;48(D1):D84–d86. doi:10.1093/nar/gkz956
28. Tatusova T, DiCuccio M, Badretdin A, et al. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016;44(14):6614–6624. doi:10.1093/nar/gkw569
29. Bortolaia V, Kaas RS, Ruppe E, et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J Antimicrob Chemother. 2020;75(12):3491–3500. doi:10.1093/jac/dkaa345
30. Larsen MV, Cosentino S, Rasmussen S, et al. Multilocus sequence typing of total-genome-sequenced bacteria. J Clin Microbiol. 2012;50(4):1355–1361. doi:10.1128/JCM.06094-11
31. Carattoli A, Zankari E, García-Fernández A, et al. In Silico Detection and Typing of Plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob Agents Chemother. 2014;58(7):3895–3903. doi:10.1128/AAC.02412-14
32. Siguier P, et al. ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006;34(Database issue):D32–6. doi:10.1093/nar/gkj014
33. Morgulis A, Coulouris G, Raytselis Y, et al. Database indexing for production MegaBLAST searches. Bioinformatics. 2008;24(16):1757–1764. doi:10.1093/bioinformatics/btn322
34. Alikhan NF, Petty NK, Ben Zakour NL, et al. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics. 2011;12(1):402. doi:10.1186/1471-2164-12-402
35. Sullivan MJ, Petty NK, Beatson SA. Easyfig: a genome comparison visualizer. Bioinformatics. 2011;27(7):1009–1010. doi:10.1093/bioinformatics/btr039
36. Novais Â, Vuotto C, Pires J, et al. Diversity and biofilm-production ability among isolates of Escherichia coli phylogroup D belonging to ST69, ST393 and ST405 clonal groups. BMC Microbiol. 2013;13(1):144. doi:10.1186/1471-2180-13-144
37. Behzadi P, Najafi A, Behzadi E, et al. Microarray long oligo probe designing for Escherichia coli: an in-silico DNA marker extraction. Cent European J Urol. 2016;69(1):105–111. doi:10.5173/ceju.2016.654
38. Khonsari MS, Behzadi P, Foroohi FJMG. The prevalence of type 3 fimbriae in Uropathogenic Escherichia coli isolated from clinical urine samples. Water. 2021;28:100881.
39. Mattioni Marchetti V, Bitar I, Piazza A, et al. Genomic Insight of VIM-harboring IncA Plasmid from a Clinical ST69 Escherichia coli Strain in Italy. Microorganisms. 2020;8(8):1232. doi:10.3390/microorganisms8081232
40. Soliman AM, Ramadan H, Sadek M, et al. Draft genome sequence of a bla(NDM-1)- and bla(OXA-244)-carrying multidrug-resistant Escherichia coli D-ST69 clinical isolate from Egypt. J Glob Antimicrob Resist. 2020;22:832–834. doi:10.1016/j.jgar.2020.07.015
41. Hammad AM, Hoffmann M, Gonzalez-Escalona N, et al. Genomic features of colistin resistant Escherichia coli ST69 strain harboring mcr-1 on IncHI2 plasmid from raw milk cheese in Egypt. Infect Genet Evol. 2019;73:126–131. doi:10.1016/j.meegid.2019.04.021
42. Yang QE, Sun J, Li L, et al. IncF plasmid diversity in multi-drug resistant Escherichia coli strains from animals in China. Front Microbiol. 2015;6:964. doi:10.3389/fmicb.2015.00964
43. Mathers AJ, Peirano G, Pitout JD. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin Microbiol Rev. 2015;28(3):565–591. doi:10.1128/CMR.00116-14
44. Liao XP, Liu B-T, Yang Q-E, et al. Comparison of plasmids coharboring 16S rRNA methylase and extended-spectrum β-lactamase genes among Escherichia coli isolates from pets and poultry. J Food Prot. 2013;76(12):2018–2023. doi:10.4315/0362-028X.JFP-13-200
45. Hozzari A, Behzadi P, Kerishchi Khiabani P, et al. Clinical cases, drug resistance, and virulence genes profiling in Uropathogenic Escherichia coli. J Appl Genet. 2020;61(2):265–273. doi:10.1007/s13353-020-00542-y
46. Woodford N, Carattoli A, Karisik E, et al. Complete Nucleotide Sequences of Plasmids pEK204, pEK499, and pEK516, Encoding CTX-M Enzymes in Three Major Escherichia coli Lineages from the United Kingdom, All Belonging to the International O25:H4-ST131 Clone. Antimicrob Agents Chemother. 2009;53(10):4472–4482. doi:10.1128/AAC.00688-09
47. Dahmen S, Madec JY, Haenni M. F2:A-:B- plasmid carrying the extended-spectrum β-lactamase bla(CTX-M-55/57) gene in Proteus mirabilis isolated from a primate. Int J Antimicrob Agents. 2013;41(6):594–595. doi:10.1016/j.ijantimicag.2013.02.004
48. Bevan ER, Jones AM, Hawkey PM. Global epidemiology of CTX-M β-lactamases: temporal and geographical shifts in genotype. J Antimicrob Chemother. 2017;72(8):2145–2155. doi:10.1093/jac/dkx146
49. Matsumura Y, Johnson JR, Yamamoto M, et al. CTX-M-27- and CTX-M-14-producing, ciprofloxacin-resistant Escherichia coli of the H 30 subclonal group within ST131 drive a Japanese regional ESBL epidemic. J Antimicrob Chemother. 2015;70(6):1639–1649. doi:10.1093/jac/dkv017
50. Rohde AM, Zweigner J, Wiese-Posselt M, et al. Prevalence of third-generation cephalosporin-resistant Enterobacterales colonization on hospital admission and ESBL genotype-specific risk factors: a cross-sectional study in six German university hospitals. J Antimicrob Chemother. 2020;75(6):1631–1638. doi:10.1093/jac/dkaa052
51. Zhang Y, et al. A Novel Structure Harboring bla(CTX-M-27) on IncF Plasmids in Escherichia coli Isolated from Swine in China. Antibiotics. 2021;10(4).
52. Poirel L, Decousser JW, Nordmann P. Insertion sequence ISEcp1B is involved in expression and mobilization of a bla(CTX-M) beta-lactamase gene. Antimicrob Agents Chemother. 2003;47(9):2938–2945. doi:10.1128/AAC.47.9.2938-2945.2003
53. Poirel L, Lartigue M-F, Decousser J-W, et al. IS Ecp1B -Mediated Transposition of bla CTX-M in Escherichia coli. Antimicrob Agents Chemother. 2005;49(1):447–450. doi:10.1128/AAC.49.1.447-450.2005
54. Tavakoli NP, DeVost J, Derbyshire KM. Defining functional regions of the IS903 transposase. J Mol Biol. 1997;274(4):491–504. doi:10.1006/jmbi.1997.1410
55. Mollet B, Iida S, Shepherd J, et al. Nucleotide sequence of IS 26, a new prokaryotic mobile genetic etement. Nucleic Acids Res. 1983;11(18):6319–6330. doi:10.1093/nar/11.18.6319
56. Harmer CJ, Hall RM. IS26 cannot move alone. J Antimicrob Chemother. 2021;76(6):1428–1432. doi:10.1093/jac/dkab055
57. Harmer CJ, Moran RA, Hall RM. Movement of IS26-associated antibiotic resistance genes occurs via a translocatable unit that includes a single IS26 and preferentially inserts adjacent to another IS26. mBio. 2014;5(5):e01801–14. doi:10.1128/mBio.01801-14
58. Lee KY, Hopkins JD, Syvanen M. Direct involvement of IS26 in an antibiotic resistance operon. J Bacteriol. 1990;172(6):3229–3236. doi:10.1128/jb.172.6.3229-3236.1990
59. Leclercq R, Courvalin P. Resistance to macrolides and related antibiotics in Streptococcus pneumoniae. Antimicrob Agents Chemother. 2002;46(9):2727–2734. doi:10.1128/AAC.46.9.2727-2734.2002
60. Arenz S, Ramu H, Gupta P, et al. Molecular basis for erythromycin-dependent ribosome stalling during translation of the ErmBL leader peptide. Nat Commun. 2014;5(1):3501. doi:10.1038/ncomms4501
61. Noguchi N, Takada K, Katayama J, et al. Regulation of Transcription of the mph (A) Gene for Macrolide 2′-Phosphotransferase I in Escherichia coli: characterization of the Regulatory Gene mphR (A). J Bacteriol. 2000;182(18):5052–5058. doi:10.1128/JB.182.18.5052-5058.2000
62. Nusrin S, Asad A, Hayat S, et al. Multiple Mechanisms Confer Resistance to Azithromycin in Shigella in Bangladesh: a Comprehensive Whole Genome-Based Approach. Microbiol Spectr. 2022;10(4):e0074122. doi:10.1128/spectrum.00741-22
63. Xiang Y, Wu F, Chai Y, et al. A new plasmid carrying mphA causes prevalence of azithromycin resistance in enterotoxigenic Escherichia coli serogroup O6. BMC Microbiol. 2020;20(1):247. doi:10.1186/s12866-020-01927-z
64. Gillings M, Boucher Y, Labbate M, et al. The evolution of class 1 integrons and the rise of antibiotic resistance. J Bacteriol. 2008;190(14):5095–5100. doi:10.1128/JB.00152-08
65. Baltazar M, Bourgeois-Nicolaos N, Larroudé M, et al. Activation of class 1 integron integrase is promoted in the intestinal environment. PLoS Genet. 2022;18(4):e1010177. doi:10.1371/journal.pgen.1010177
66. Gillings MR. Integrons: past, present, and future. Microbiol Mol Biol Rev. 2014;78(2):257–277. doi:10.1128/MMBR.00056-13
67. Fluit AC, Schmitz FJ. Class 1 integrons, gene cassettes, mobility, and epidemiology. Eur J Clin Microbiol Infect Dis. 1999;18(11):761–770. doi:10.1007/s100960050398
68. Collis CM, Hall RM. Site-specific deletion and rearrangement of integron insert genes catalyzed by the integron DNA integrase. J Bacteriol. 1992;174(5):1574–1585. doi:10.1128/jb.174.5.1574-1585.1992
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.