")
Back to Journals » Infection and Drug Resistance » Volume 17
Vitamin D Improves Klebsiella-Induced Severe Pneumonia in Rats by Regulating Intestinal Microbiota
Authors Hu JJ, Lin YS, Zhang JC , Wang YH
Received 27 September 2023
Accepted for publication 17 January 2024
Published 8 February 2024 Volume 2024:17 Pages 475—484
DOI https://doi.org/10.2147/IDR.S442330
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Sandip Patil
Jia-Jia Hu,* Yu-Sen Lin,* Jing-Cong Zhang, Yan-Hong Wang
Medical Intensive Care Unit, The Third Affiliated Hospital, Sun Yat-sen University, Guangzhou, Guangdong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yan-Hong Wang, Email [email protected]
Background: In the context of progressively uncontrolled drug resistance of bacteria, the difficulty of treating Klebsiella (KP)-induced pneumonia increases. Searching for drugs other than antibiotics has become an urgent task. Vitamin D (VD), meanwhile, is shown to be capable of treating pneumonia. Therefore, we aimed to explore the effects and mechanisms of VD on KP-infected rats.
Methods: Male Sprague Dawley rats were divided into the Control, VD, KP and KP+VD groups. A rat pneumonia model was induced using an intratracheal drop of 2.4× 108 CFU/mL KP. VD treatment was performed by gavage using 5 μg/kg. Subsequently, the survival of the rats was recorded, and the lungs, bronchoalveolar lavage fluid, and feces of the rats were collected 4 days after KP infection. Next, the water content of lung tissues was measured by the wet-to-dry weight ratio. Histopathological changes of lung tissues were observed by Hematoxylin and Eosin staining and the levels of inflammatory factors (TNF-α, IL-1β, MCP1) were detected using ELISA. The feces of rats in each group were also subjected to 16S rDNA gene analysis of intestinal microbiota.
Results: Compared with the KP group, the KP+VD group showed a significant increase in survival, a significant decrease in water content and bacterial counts in the lungs, a significant improvement in lung injury, and a significant decline in the levels of TNF-α, IL-1β, and MCP1. According to the 16S rDNA sequencing, VD altered the structure of the intestinal bacterial community in the KP-infected rats and made the species richness similar to that of healthy rats. Additionally, the abundance of Anaeroglobus was significantly increased in the KP+VD group.
Conclusion: VD modulates intestinal microbiota to increase the resistance of rats to pneumonia caused by Klebsiella infection.
Keywords: severe pneumonia, Klebsiella, intestinal microbiota, vitamin D
Introduction
Klebsiella, a group of gram-negative bacteria belonging to the Enterobacteriaceae family, is recognized as the leading cause of pneumonia.1 In healthy populations, Klebsiella colonizes the surface of oropharyngeal, nasopharyngeal and gastrointestinal mucosa.2 Moreover, Klebsiella has also long been identified as a pathogen of disease and remains one of the most common pathogens in hospitals worldwide.3 Antibiotics have always been first choice against Klebsiella, such as beta-lactams, aminoglycosides, and tetracyclines.4 However, the use of antibiotics may lead to a serious consequence—antibiotic resistance.5 Antibiotic resistance is now the major contributor to global death. In 2019, 1.27 million people died directly from antibiotic resistance, and another 4.95 million deaths were linked to antibiotic-resistant infections.6 It is predicted that by 2050, antibiotic-resistant bacteria will cause 10 million deaths per year and result in a global economic loss of up to 100 trillion US dollars.7 The World Health Organization listed extended-spectrum beta-lactamase (ESBL)-producing and carbapenemase-resistant Klebsiella pneumoniae (KP) as a serious public health threat back in 2017.8 Such Klebsiella causes more than 900,000 infections and more than 70,000 deaths annually in Europe alone, accounting for 25% of deaths or disabilities caused by multidrug-resistant bacterial infections.9 Therefore, there is an urgent need to find new drugs to treat bacterial infections, especially Klebsiella infections.
In previously recognition, vitamin D (VD) is considered to play a role in maintaining calcium and phosphorus levels in the blood. It enhances calcium absorption in the small intestine by increasing the expression of calcium transporting proteins, the epithelial calcium channel TRPV6, and calbindin in the small intestinal epithelial cells.10 Later studies have suggested that VD signaling is also integral to the regulation of the immune system, mediating the expression of the immune-related protein CYP27B1.11 VD can activate the transcription of innate immune-related genes, including TLR2 or TLR4, by binding to the VD nuclear receptor.12 Additionally, Liao et al directly demonstrated that VD was able to promote epithelial tissue repair and host defense responses against H1N1 influenza virus and Staphylococcus aureus infections.13 Hence, VD is believed to function in activating the body’s antimicrobial capacity. Furthermore, VD also regulates intestinal microbiota, and dysregulation of intestinal microbiota is associated with the occurrence of a variety of bacterial infections, including respiratory and pulmonary infections.14 Li et al found that administration of oral Bifidobacterium tetravaccine tablets (Live) changed the structure of the intestinal flora and significantly improved the symptoms of recurrent respiratory tract infections in children.15 In addition, Schuijt et al revealed that the gut microbiota protected the primary alveolar macrophage function during pneumococcal pneumonia.16 It is evident that gut flora is particularly important for lung tissues against bacterial infections. Based on these studies, we tried to observe the therapeutic effect and mechanism of VD on rats infected with Klebsiella.
Materials and Methods
Strains and Culture
KP (43816; serotype 2) was purchased from the American Type Culture Collection (Manassas, VA, USA) and cultured at 37°C using Luria-Bertani (LB) medium. To obtain KP, firstly, LB solid medium was streaked, and after an overnight incubation at 37°C, single colonies were seeded into a 5 mL of LB liquid medium and incubated again in a gas bath shaker at 200 rpm for 16 h. Then, they were transferred to a fresh LB medium at a ratio of 1:100 until OD600=1.0. After that, the fluids containing bacteria were centrifuged at 4000 rpm and the precipitate was collected. Subsequently, three times of saline washes and centrifugation at 4000 rpm were carried out. Again, the precipitate was collected. Finally, the bacteria were resuspended to 2.4×108 CFU/mL with normal saline17 for subsequent use.
Rat Feeding
Male Sprague Dawley rats weighing 180 g–220 g were housed in cages (6 rats/cage) in an environment with a 12 h day-night cycle, a room temperature of 22°C, a relative humidity of 50%, and free access to food and water. The animal experiments in this study were approved by the Animal Ethics Committee of the Third Affiliated Hospital, Sun Yat-sen University. Besides, all procedures were strictly adhere to the internationally recognized basic principles of the “three R’s” of laboratory animal welfare, including Replace, Reduce and Refine.18
Rat Model of Pulmonary Infection
The rats were divided into Control, VD, KP, and KP+VD groups. Briefly, the rats in the Control and KP groups were administered intragastrically with saline (once a day for 8 days), whereas the other two groups were given equal amount of VD (once a day for 8 days). Besides, rats in the KP and KP+VD groups were given KP infection on the 7th day. A rat model of pulmonary bacterial infection was established by intratracheal instillation of the bacterial fluid, and the specific steps were shown as follows. After anaesthesia by CO2, the rats were fixed on a wooden board and kept at a tilt of 60°. The incisors of the rats were fixed, the tongues were pulled out with forceps, and the pharyngeal secretions were wiped clean. The position of the rats was adjusted to the point that the fissure of glottis could be clearly observed through the oral cavity. Subsequently, a catheter was inserted into the airway, followed by slow intratracheal instillation of 50 μL bacterial suspension. During this process, the rats were kept in a head-high, foot-low position and rotated from side to side so that the suspension could be evenly distributed in their right and left lungs. On completion of the instillation, the rats were placed in the lateral position. After awakening, the rats were put in sterile cages and allowed to eat freely. The rats were observed daily for their feeding, hair, mental status, and survival status.17,19,20 In the Control and VD groups, intratracheal instillation of same amount of sterile saline was performed as described above. Subsequently, the survival rate of rats in each group was recorded. The rats were euthanized 4 days after infection. Then, the lung tissues, bronchoalveolar lavage fluid (BALF), and feces were collected from rats in each group.
Measurement of Lung Water Content
The obtained fresh lung was placed on weighing paper, and the excess blood and water on the surface of the lung were wiped off using the weighing paper. The wet weight of the lung tissues was subsequently weighed in a Cubis® II analytical balance (Sartorius, Germany) as wet weight. Next, the lung tissues of each group of rats were placed in beakers and dried in a thermostat at 70°C until the weight of the lung was constant (dry weight). Lastly, lung water content was measured by calculating wet-to-dry (W/D) weight ratios.
Calculation of Bacterial Colony Count in Lung Tissue
Firstly, fresh lung tissues were weighed, and then 50 mg lung from each group was collected in a homogenizing tube. Upon adding homogenizing beads and 1 mL of saline, the tube was placed in a tissue grinder (Tissuelyser-24L, Shanghai Jingxin, China). After the lung tissues were completely disrupted, 100 μL of homogenizing solution was subjected to five consecutive 10-fold dilutions, and all gradients of solution (5 μL) was dropped on the Mueller-Hinton agar. The droplets were air-dried, and the colonies were counted after 18-h incubation in a 37°C thermostat. Additionally, the number of colonies was calibrated by organ weight.
Hematoxylin and Eosin (H&E) Staining
Lung tissue samples were fixed in 4% paraformaldehyde and embedded in paraffin. The embedded samples were cut into multiple 4 μm-thick tissue sections using a slicer (RM2125RTS, Leica, Germany). The sections were then baked, deparaffinized in xylene, and hydrated in gradient ethanol solution (high to low concentration). Next, they were stained in hematoxylin solution for 1 min, followed by differentiation in 1% acid alcohol for 15 sec and washing step using running water. Subsequently, the sections were incubated in eosin solution for 3 min. They were then dehydrated in gradient ethanol solution (high to low concentration), cleared in xylene, and cover-slipped with neutral balsam. Ultimately, the samples were observed under a microscope (CKX41, Olympus, Japan) and photographed.
Enzyme-Linked Immunosorbent Assay (ELISA)
The lungs of euthanized rats were exposed, and 3 mL of saline was pushed into the lungs from the tracheal cannula. BALF was suctioned and collected after repeated lavage for three times. BALF was then centrifuged at 3000 r/min at 4°C for 15 min, and the levels of inflammation-related factors (TNF-α, IL-1β, MCP1) in the supernatant were detected. According to the instructions of the corresponding ELISA kits for TNF-α (EK382, Multi sciences, China), IL-1β (EK301B, Multi sciences, China), and MCP1 (EK387, Multi sciences, China), incubation, reaction, and color development were carried out. The levels of each factor was measured at the wavelength of 450 nm using a microplate reader (800 TS, Agilent BioTek, USA).
Sequencing of Intestinal Microbiota
The fecal genomic DNA in each group of rats collected was extracted using the Stool Genomic DNA Extraction Kit (DP328, TIANGEN, China). The quality and purity of DNA were observed by 1% agarose gel electrophoresis. The concentration of DNA was adjusted to 1 ng/μL. Bacterial-specific 16S rDNA amplification primers (341F: CCTAYGGGRBGCASCAG; 806R: GGACTACNNGGGGTATCTAAT) were employed to amplify 16S rDNA of bacteria in feces, and then 1% agarose gel electrophoresis was performed to observe the quality and purity of 16S rDNA. The size of the bacterial 16S rDNA was 400–450 bp. The PCR products were then purified using the TIANquick Mini Purification Kit (DP203, TIANGEN, China). After generating sequencing libraries with the TruSeq® DNA PCR-Free Sample Preparation Kit (Illumina, USA), the quality of the libraries was assessed on an Agilent Bioanalyzer 2100 system. Finally, the libraries were sequenced on the Illumina NovaSeq 6000 platform and 250 bp paired-end reads were generated. Sequences without primers were removed using Cutadapt (v1.18), and fully sequenced primers were cut from the reads. The clustering was performed in Q30.Usearch (version 11.0.667) based on 97% similarity. Alpha diversity (Chao1 and Shannon) was calculated using Mothur v1.1.42, whereas beta diversity was calculated using vegan package. Additionally, species differences were analyzed using deseq2 package.21
Data Analysis
All quantitative results in this study were expressed as mean ± standard error and statistically analyzed using SPSS.23. Unpaired two-tailed t-test was used to assess the differences between the two groups and one-way analysis of variance was used to compare more than two groups. P-value less than 0.05 was considered significant.
Results
Vitamin D Promotes Survival and Reduces Lung Water Content and Bacteria Count in Klebsiella pneumoniae-Induced Rats
VD deficiency is thought to be one of the most important factors driving the progression of lung disease,22 but whether VD affects KP infection is unknown. To observe their relationship, KP-infected rats were treated with VD. Briefly, the survival rate of rats in the KP groups was significantly lower compared with the controls. However, the survival rate of rats in the KP+VD group was significantly higher than that in the KP group (Figure 1A). Based on the W/D weight ratios, the water content of lung tissues of KP-infected rats was significantly increased, while it was reduced after VD treatment (Figure 1B). Meanwhile, the number of bacteria in the lung tissues of rats in each group was counted, and the results showed that the bacteria count was significantly increased in KP-infected rats while markedly decreased after VD treatment (Figure 1C). These results indicated that VD could resist lung infection caused by KP.
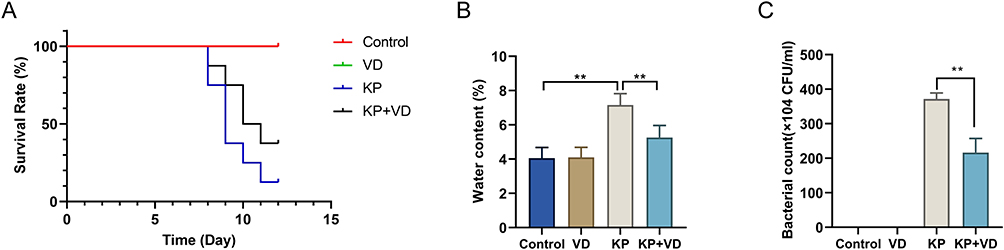 |
Figure 1 Vitamin D promotes survival and reduces lung water content and bacteria count in Klebsiella pneumoniae-induced rats. (A) Survival rate of rats in Control, VD, KP, and KP+VD groups within 15 days after pulmonary infection; (B) Water content of lung tissues of rats in each group; (C) Number of bacteria in lung tissues of rats in each group within 15 days after lung infection. **p < 0.01. Abbreviations: VD, Vitamin D; KP, Klebsiella pneumoniae. |
Vitamin D Attenuates Pathological Lung Injury in Klebsiella pneumoniae-Induced Rats
In order to explore whether VD would affect the destruction of lung tissues by KP, the lung histopathology of rats in each group was observed by H&E staining. The Control groups showed normal alveolar structure with intact alveolar walls. The lung tissues of rats in the KP group were significantly damaged, with thickening of the alveolar walls, infiltration of a large number of inflammatory cells in the alveolar spaces, alveolar dilatation, and congestion in the pulmonary alveoli and parenchyma. However, after VD treatment, the lung injury of KP-infected rats was significantly improved, and the inflammatory infiltration and congestion were alleviated (Figure 2). These results suggested that VD protected lung tissues from KP-caused damage.
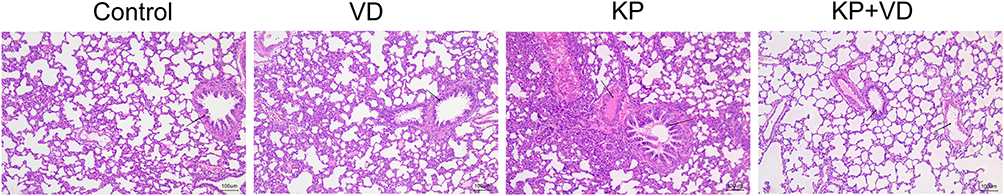 |
Figure 2 Vitamin D attenuates pathological lung injury in Klebsiella pneumoniae-induced rats. H&E staining was used to detect pathological changes in the lung tissues of rats in the Control, VD, KP, and KP+VD groups. Abbreviations: VD, Vitamin D; KP, Klebsiella pneumoniae. |
Vitamin D Attenuates the Inflammation Levels in Klebsiella pneumoniae-Induced Rats
VD has been proved to be a factor involved in the regulation of immune response.23 We examined the levels of inflammatory factors (TNF-α, IL-1β, MCP1) in BALF to determine the effect of VD on inflammation of lung tissues in KP-infected rats. The results showed that KP infection caused a significant increase in the levels of TNF-α, IL-1β, and MCP1 in BALF, but their levels were markedly decreased after VD treatment (Figure 3A–C). These results suggested that VD could reduce the inflammatory response of lung tissues induced by KP.
 |
Figure 3 Vitamin D attenuates inflammation in Klebsiella pneumoniae-induced rats. (A–C) ELISA was performed to detect the levels of TNF-α (A), IL-1β (B), and MCP1 (C) in the bronchoalveolar lavage fluid of the Control, VD, KP, and KP+VD groups. **p < 0.01. Abbreviations: D, Vitamin D; KP, Klebsiella pneumoniae. |
Vitamin D Changes Intestinal Bacterial Community Structure in Klebsiella pneumoniae-Induced Rats
Pulmonary inflammation caused by KP infection is associated with alterations in the intestinal microbiome,24 and VD has been demonstrated to modulate the intestinal microbiota in inflammatory bowel diseases.14 Therefore, we hypothesized that VD might affect KP-induced pulmonary inflammation by modulating the intestinal microbiota. The feces were collected from each group of rats and subject to 16S rDNA sequencing to observe the structure of the intestinal microbiota in each group. At the portal level, the intestinal microbial composition of the KP group was different from that of the other three groups. The abundance of Bacteroidetes and Proteobacteria was significantly increased in KP-infected rats but decreased after VD treatment (Figure 4A). At the family level, we observed a significant increase in the abundance of Enterobacteriaceae and a marked decrease in the abundance of Lachnospiraceae in KP-infected rats. Compared with the KP group, the abundance of Enterobacteriaceae was reduced, whereas the abundance of Veillonellaceae was significantly increased in the KP+VD group (Figure 4B). At the genus level, the intestinal microbiota of rats in the KP group showed a significant decrease in the proportion of Veillonella and a significant increase in the abundance of Klebsiella. VD treatment decreased the abundance of Klebsiella and increased the abundance of Veillonella in the KP-infected rats (Figure 4C). Finally, at the species level, a significant increase in the abundance of Escherichia coli and a significant decrease in Bacteroides_fragilis could be observed in the KP group. The abundance of both Lactobacillus_salivarius and Fusobacterium_nucleatum was significantly increased in the KP+VD group compared to the KP group (Figure 4D). Collectively, VD significantly altered the structure of the intestinal bacterial community in KP-induced rats.
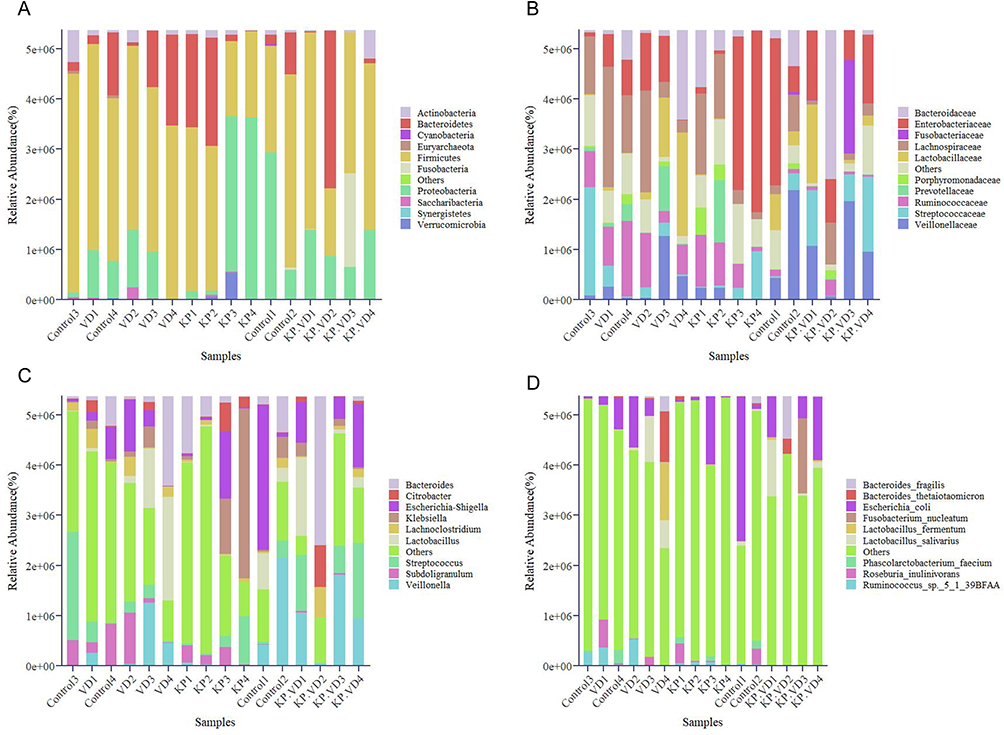 |
Figure 4 Vitamin D changes intestinal bacterial community structure in Klebsiella pneumoniae-induced rats. (A–D) Bacterial community structure in the Control, VD, KP, and KP+VD group samples was assessed based on phylum (A), family (B), genus (C), and species (D) levels. Abbreviations: VD, Vitamin D; KP, Klebsiella pneumoniae. |
Vitamin D Alters the Diversity of Intestinal Bacterial Structure in Klebsiella pneumoniae-Induced Rats
Subsequently, α-diversity analysis was performed on the 16S rDNA sequencing data. The results showed that the chao1 value, Shannon value, Simpson value, and Observed value of the KP group were higher than those of the Control group, whereas these values were significantly lower in the KP+ group compared to the KP group (Figure 5A–D). In addition, ANOSIM analysis revealed a significant increase in the Bray-Curtis index of the other three groups of rats relative to the Control group (Figure 5E). According to PCoA analysis, the bacterial colony richness of the KP+VD group, rather than the KP group, was closest to that of the Control group (Figure 5F). Taken together, the bacterial community in the KP group had a higher species richness and species evenness than the Control or KP+VD groups. VD significantly reduced the disturbance in the intestinal microbiota richness and made the richness similar to that of healthy rats.
 |
Figure 5 Vitamin D alters the diversity of intestinal bacterial structure in Klebsiella pneumoniae-induced rats. (A–D) α-diversity analysis of chao1 value (A), Shannon value (B), Simpson value (C), and Observed value (D) of bacterial communities in samples of the Control, VD, KP, and KP+VD groups. (E) Bray-Curtis anosim of bacterial colonies in the Control, VD, KP, and KP+VD groups. (F) PCoA analysis of the weighting of bacterial colonies in the Control, VD, KP, and KP+VD groups. Abbreviations: VD, Vitamin D; KP, Klebsiella pneumoniae. |
Composition of Differential Species in Bacterial Community
We then analyzed the key flora mediating gut microbial remodeling in KP-infected lung tissues as well as in KP-infected rats with VD treatment. LEfSe analysis and linear discriminant analysis were used to reveal the composition of differential species in gut microbiome in each group. The results showed that the relative abundance of Prevotella, Lachnospiraceae, and Ruminiclostridium was significantly higher in the KP group than in the KP+VD and Control groups. Besides, Anaeroglobus had the highest relative abundance in the KP+VD group, and Lachnospira and Alcaligenaceae had the highest relative abundance in the VD group (Figure 6A and B). Collectively, Prevotella, Lachnospiraceae, and Ruminiclostridium might be the key flora mediating KP-induced lung infection, while Anaeroglobus abundance was critical for the treatment of KP-induced lung infection.
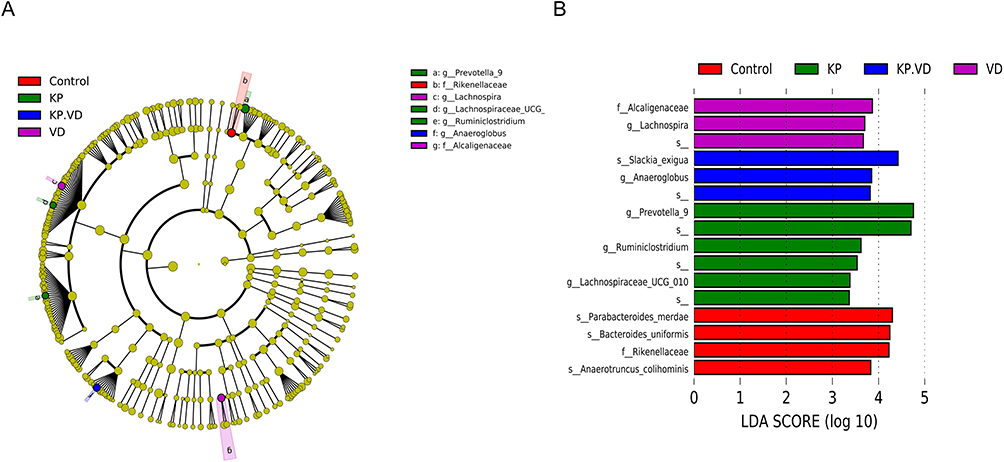 |
Figure 6 Composition of differential species in bacterial community. (A) LEfSe analysis for the composition of differential species in microbial communities in the Control, VD, KP, and KP+VD groups. (B) Linear discriminant analysis (LDA) of microbiota that differed significant ly among the Control, VD, KP, and KP+VD groups. |
Discussion
Alarmingly, the detection rate of Klebsiella in clinical isolates shows an upward trend, with KP-caused pneumonia accounting for 17.2% of community-acquired pneumonia and 6.5–11.6% of hospital-acquired pneumonia.25 The biggest limiting factor for the treatment of KP-induced infection is the development of antibiotic resistance.26 Relative to other strains, KP is highly susceptible to developing resistance to most of the commonly used first-line antibiotics, such as meropenem, cephalosporins, and polymyxins (known as the last line of defense against gram-negative bacteria).27 It is evident that KP poses a great threat to human health. However, here we discovered the potential of VD to treat rats infected by KP.
Because VD is thought to have anti-infective properties, it has been used as an adjunct to the treatment of childhood pneumonia in early studies. For instance, one meta-analysis showed that high doses of VD administered as an adjunct to the treatment of pneumonia significantly boosted the immune system of patients to reduce the incidence of pneumonia recurrence.28 Furthermore, Manaseki-Holland et al observed in clinical trials that VD supplementation contributed to increasing the survival rate of children with pneumonia and preventing recurrent episodes of the disease.29 Also, some clinical studies did not observe significant preventive or therapeutic effect of VD supplementation in children with pneumonia or severe pneumonia,30,31 but the children in these studies only received a single or short-term use of VD. The efficacy of VD in the treatment of pneumonia is controversial, as emphasized in the review by Das et al.32 Additionally, the efficacy of VD in the treatment of pneumonia caused by KP infection has not been reported. Here, our study first revealed that VD significantly increased the survival of KP-infected rats, reduced lung edema and bacterial counts, and ameliorated lung tissue damage. VD receptor and VD metabolizing enzymes are widely expressed in immune cells, including lymphocytes, monocytes, and macrophages, participating in the development of various immune-related diseases.33,34 Therefore, vitamin D has been classified as a candidate for the prevention and treatment of immune-related diseases. Here, we observed that VD modulated the immune response in lung tissues, significantly reducing the levels of TNF-α, IL-1β, and MCP1. Although our study suggested that vitamin D may be useful in treating pneumonia caused by KP, the specific effects of VD as an independent treatment for pneumonia still remain to be studied and discussed. Pneumonia is often considered as an integral part of a comprehensive treatment strategy, rather than a separate treatment. Therefore, exploring the role of VD+other drugs in the treatment of pneumonia caused by KP is an important research direction in the future.
Recently, a growing number of studies have proved a definite link between the homeostasis of gut microbiota and the development of pneumonia.35 Lung diseases are often associated with dysbiosis and an immune-inflammatory response.36 Jiang et al disclosed significant changes in the intestinal microbiota in patients with pneumonia.37 Deng et al demonstrated that altering the intestinal flora could alleviate the symptoms of infectious pneumonia.38 Rastogi et al emphasized again the importance of intestinal flora for the immune response and gut-lung homeostasis.39 Meanwhile, Battistini et al determined the role of VD in regulating intestinal flora.14 Based on these studies, we hypothesized that VD may resist KP infection by affecting the intestinal flora of rats. Therefore, 16S rDNA gene analysis was performed on the intestinal flora of rats from each group, and the analysis results revealed a significant increase in the abundance of Prevotella, Lachnospiraceae, Ruminiclostridium in the KP-infected rats. Pettigrew et al also reported a strong association between Prevotella and the development of pneumonia.40 The abundance of Anaeroglobus was significantly increased in the KP+VD group, which may be a key factor in stopping lung infections.41 Moreover, VD was able to restore the structure and abundance of the intestinal flora in KP-infected rats close to that of the Control group. It is known from a previous study that VD is associated with increased levels of beneficial intestinal bacteria and decreased levels of pathogenic bacteria.42 On the one hand, VD regulates the intestinal microbiota by maintaining the integrity of the intestinal barrier, and on the other hand, it influences the distribution of fecal microbiota by promoting the innate immune system through the VD receptor.43 Therefore, we believe that VD fights against KP-induced infection by regulating the intestinal flora. However, we did not further explore the mechanisms VD regulating the intestinal microbiota and the intestinal microbiota influencing KP infection. Additionally, we did not rule out the mechanism VD directly regulating the immune system of the lungs to resist KP infection. All of these questions require the development of an experimental plan for further investigation.
Conclusion
To sum up, VD regulates intestinal microbiota, increases the body resistance to KP infection, reduces mortality caused by KP, and alleviates KP-induced lung damage and inflammatory response. Therefore, we believe that VD may be used to treat pneumonia caused by KP infection. However, further research is still needed in the future to validate and establish our findings.
Data Sharing Statement
The datasets used during the current study are available from the corresponding author on reasonable request.
Ethics Approval and Consent to Participate
The animal experiments in this study were approved by the Animal Ethics Committee of The Third Affiliated Hospital, Sun Yat-sen University, and animal welfare was strictly adhered to.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Marr CM, Russo TA. Hypervirulent Klebsiella pneumoniae: a new public health threat. Expert Rev Anti Infect Ther. 2019;17(2):71–73. doi:10.1080/14787210.2019.1555470
2. Piperaki ET, Syrogiannopoulos GA, Tzouvelekis LS, Daikos GL. Klebsiella pneumoniae: virulence, biofilm and antimicrobial resistance. Pediatr Infect Dis J. 2017;36(10):1002–1005. doi:10.1097/INF.0000000000001675
3. Wang G, Zhao G, Chao X, Xie L, Wang H. The characteristic of virulence, biofilm and antibiotic resistance of Klebsiella pneumoniae. Int J Environ Res Public Health. 2020;17(17). doi:10.3390/ijerph17176278
4. Bhardwaj S, Mehra P, Dhanjal DS, et al. Antibiotics and antibiotic resistance- flipsides of the same coin. Curr Pharm Des. 2022;28(28):2312–2329. doi:10.2174/1381612828666220608120238
5. Stojowska-Swedrzynska K, Lupkowska A, Kuczynska-Wisnik D, Laskowska E. Antibiotic Heteroresistance in Klebsiella pneumoniae. Int J Mol Sci. 2021;23(1). doi:10.3390/ijms23010449
6. Antimicrobial Resistance C. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet. 2022;399(10325):629–655. doi:10.1016/S0140-6736(21)02724-0
7. Hardie KR. Antimicrobial resistance: the good, the bad, and the ugly. Emerg Top Life Sci. 2020;4(2):129–136. doi:10.1042/ETLS20190194
8. Wyres KL, Lam MMC, Holt KE. Population genomics of Klebsiella pneumoniae. Nat Rev Microbiol. 2020;18(6):344–359. doi:10.1038/s41579-019-0315-1
9. Cassini A, Hogberg LD, Plachouras D, et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: a population-level modelling analysis. Lancet Infect Dis. 2019;19(1):56–66. doi:10.1016/S1473-3099(18)30605-4
10. Wongdee K, Chanpaisaeng K, Teerapornpuntakit J, Charoenphandhu N. Intestinal calcium absorption. Compr Physiol. 2021;11(3):2047–2073. doi:10.1002/cphy.c200014
11. Voutsadakis IA. Vitamin D receptor (VDR) and metabolizing enzymes CYP27B1 and CYP24A1 in breast cancer. Mol Biol Rep. 2020;47(12):9821–9830. doi:10.1007/s11033-020-05780-1
12. Rizvi I, Garg RK, Jain A, et al. Vitamin D status, vitamin D receptor and toll like receptor-2 polymorphisms in tuberculous meningitis: a case-control study. Infection. 2016;44(5):633–640. doi:10.1007/s15010-016-0907-x
13. Liao S, Huang Y, Zhang J, et al. Vitamin D promotes epithelial tissue repair and host defense responses against influenza H1N1 virus and Staphylococcus aureus infections. Respir Res. 2023;24(1):175. doi:10.1186/s12931-023-02477-4
14. Battistini C, Ballan R, Herkenhoff ME, Saad SMI, Sun J. Vitamin D modulates intestinal microbiota in inflammatory bowel diseases. Int J Mol Sci. 2020;22(1). doi:10.3390/ijms22010362
15. Li KL, Wang BZ, Li ZP, Li YL, Liang JJ. Alterations of intestinal flora and the effects of probiotics in children with recurrent respiratory tract infection. World J Pediatr. 2019;15(3):255–261. doi:10.1007/s12519-019-00248-0
16. Schuijt TJ, Lankelma JM, Scicluna BP, et al. The gut microbiota plays a protective role in the host defence against pneumococcal pneumonia. Gut. 2016;65(4):575–583. doi:10.1136/gutjnl-2015-309728
17. Zhang Y, Zhu C, Zhao H, Sun Z, Wang X. Anti-inflammatory effect of chlorogenic acid in Klebsiella pneumoniae-induced pneumonia by inactivating the p38MAPK pathway. Int J Med Microbiol. 2023;313(2):151576. doi:10.1016/j.ijmm.2023.151576
18. Ohl F, van der Staay FJ. Animal welfare: at the interface between science and society. Vet J. 2012;192(1):13–19. doi:10.1016/j.tvjl.2011.05.019
19. Al-Hassan S, Attia H, Alomar H, Arafa M, Ali RA. The inhibitory mechanisms of losartan and vitamin D on amiodarone-induced lung inflammation in rats: role of mitogen-activated protein kinases/activator protein-1. J Biochem Mol Toxicol. 2021;35(12):e22923. doi:10.1002/jbt.22923
20. Zhang M, Dong M, Liu W, et al. 1alpha,25-dihydroxyvitamin D3 ameliorates seawater aspiration-induced acute lung injury via NF-kappaB and RhoA/Rho kinase pathways. PLoS One. 2014;9(8):e104507. doi:10.1371/journal.pone.0104507
21. Du J, Zhang P, Luo J, et al. Dietary betaine prevents obesity through gut microbiota-drived microRNA-378a family. Gut Microbes. 2021;13(1):1–19. doi:10.1080/19490976.2020.1862612
22. Martineau AR, Jolliffe DA, Greenberg L, et al. Vitamin D supplementation to prevent acute respiratory infections: individual participant data meta-analysis. Health Technol Assess. 2019;23(2):1–44. doi:10.3310/hta23020
23. Ao T, Kikuta J, Ishii M. The effects of vitamin D on immune system and inflammatory diseases. Biomolecules. 2021;11(11). doi:10.3390/biom11111624
24. Jiang Q, Xu Q, Kenez A, Chen S, Yang G. Klebsiella pneumoniae infection is associated with alterations in the gut microbiome and lung metabolome. Microbiol Res. 2022;263:127139. doi:10.1016/j.micres.2022.127139
25. Teramoto S, Yoshida K, Hizawa N. Update on the pathogenesis and management of pneumonia in the elderly-roles of aspiration pneumonia. Respir Investig. 2015;53(5):178–184. doi:10.1016/j.resinv.2015.01.003
26. Effah CY, Sun T, Liu S, Wu Y. Klebsiella pneumoniae: an increasing threat to public health. Ann Clin Microbiol Antimicrob. 2020;19(1):1. doi:10.1186/s12941-019-0343-8
27. Bassetti M, Righi E, Carnelutti A, Graziano E, Russo A. Multidrug-resistant Klebsiella pneumoniae: challenges for treatment, prevention and infection control. Expert Rev Anti Infect Ther. 2018;16(10):749–761. doi:10.1080/14787210.2018.1522249
28. Yang C, Lu Y, Wan M, et al. Efficacy of high-dose vitamin D supplementation as an adjuvant treatment on pneumonia: systematic review and a meta-analysis of randomized controlled studies. Nutr Clin Pract. 2021;36(2):368–384. doi:10.1002/ncp.10585
29. Manaseki-Holland S, Qader G, Isaq Masher M, et al. Effects of vitamin D supplementation to children diagnosed with pneumonia in Kabul: a randomised controlled trial. Trop Med Int Health. 2010;15(10):1148–1155. doi:10.1111/j.1365-3156.2010.02578.x
30. Gupta P, Dewan P, Shah D, et al. Vitamin D supplementation for treatment and prevention of pneumonia in under-five children: a randomized double-blind placebo controlled trial. Indian Pediatr. 2016;53(11):967–976. doi:10.1007/s13312-016-0970-5
31. Choudhary N, Gupta P. Vitamin D supplementation for severe pneumonia--a randomized controlled trial. Indian Pediatr. 2012;49(6):449–454. doi:10.1007/s13312-012-0073-x
32. Das RR, Singh M, Naik SS. Vitamin D as an adjunct to antibiotics for the treatment of acute childhood pneumonia. Cochrane Database Syst Rev. 2023;1(1):CD011597. doi:10.1002/14651858.CD011597.pub3
33. Fakhoury HMA, Kvietys PR, AlKattan W, et al. Vitamin D and intestinal homeostasis: barrier, microbiota, and immune modulation. J Steroid Biochem Mol Biol. 2020;200:105663. doi:10.1016/j.jsbmb.2020.105663
34. Sirbe C, Rednic S, Grama A, Pop TL. An update on the effects of vitamin D on the immune system and autoimmune diseases. Int J Mol Sci. 2022;23(17). doi:10.3390/ijms23179784
35. Thibeault C, Suttorp N, Opitz B. The microbiota in pneumonia: from protection to predisposition. Sci Transl Med. 2021;13(576). doi:10.1126/scitranslmed.aba0501
36. Ma PJ, Wang MM, Wang Y. Gut microbiota: a new insight into lung diseases. Biomed Pharmacother. 2022;155:113810. doi:10.1016/j.biopha.2022.113810
37. Jiang Y, Bao C, Zhao X, Chen Y, Song Y, Xiao Z. Intestinal bacteria flora changes in patients with Mycoplasma pneumoniae pneumonia with or without wheezing. Sci Rep. 2022;12(1):5683. doi:10.1038/s41598-022-09700-0
38. Deng L, Shi Y, Liu P, et al. GeGen QinLian decoction alleviate influenza virus infectious pneumonia through intestinal flora. Biomed Pharmacother. 2021;141:111896. doi:10.1016/j.biopha.2021.111896
39. Rastogi S, Mohanty S, Sharma S, Tripathi P. Possible role of gut microbes and host’s immune response in gut-lung homeostasis. Front Immunol. 2022;13:954339. doi:10.3389/fimmu.2022.954339
40. Pettigrew MM, Kwon J, Gent JF, et al. Comparison of the respiratory resistomes and microbiota in children receiving short versus standard course treatment for community-acquired pneumonia. mBio. 2022;13(2):e0019522. doi:10.1128/mbio.00195-22
41. Miyashita N. Atypical pneumonia: pathophysiology, diagnosis, and treatment. Respir Investig. 2022;60(1):56–67. doi:10.1016/j.resinv.2021.09.009
42. Cantorna MT, Lin YD, Arora J, et al. Vitamin D regulates the microbiota to control the numbers of RORgammat/FoxP3+ Regulatory T cells in the colon. Front Immunol. 2019;10:1772. doi:10.3389/fimmu.2019.01772
43. Kellermann L, Jensen KB, Bergenheim F, et al. Mucosal vitamin D signaling in inflammatory bowel disease. Autoimmun Rev. 2020;19(11):102672. doi:10.1016/j.autrev.2020.102672
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.