Back to Journals » Infection and Drug Resistance » Volume 12
Virulence-associated genes and drug susceptibility patterns of uropathogenic Escherichia coli isolated from patients with urinary tract infection
Authors Farajzadah Sheikh A, Goodarzi H, Yadyad MJ, Aslani S ![]() , Amin M
, Amin M ![]() , Jomehzadeh N
, Jomehzadeh N ![]() , Ranjbar R
, Ranjbar R ![]() , Moradzadeh M, Azarpira S, Akhond MR, Hashemzadeh M
, Moradzadeh M, Azarpira S, Akhond MR, Hashemzadeh M
Received 28 December 2018
Accepted for publication 3 May 2019
Published 17 July 2019 Volume 2019:12 Pages 2039—2047
DOI https://doi.org/10.2147/IDR.S199764
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Joachim Wink
Ahmad Farajzadah Sheikh,1,2 Hamed Goodarzi,1–3 Mohammad Jaafar Yadyad,1 Sajad Aslani,4 Mansoor Amin,1,2 Nabi Jomehzadeh,5 Reza Ranjbar,3 Mina Moradzadeh,1 Samireh Azarpira,1 Mohamad Reza Akhond,6 Mohamad Hashemzadeh1,2
1Infectious and Tropical Diseases Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 2Department of Microbiology, School of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 3Molecular Biology Research Center, Baqiyatallah University of Medical Science, Tehran, Iran; 4Student Research Committee, Kerman University of Medical Sciences, Kerman, Iran; 5Abadan Faculty of Medical Sciences, Abadan, Iran; 6Department of Statistics, Mathematical Science and Computer Faculty, Shahid Chamran University, Ahvaz, Iran
Background: Different Escherichia coli phylogenetic groups, such as A, B1, B2, and D, have four functional groups – adhesins, microcins, toxins, and capsules – which can cause urinary tract infections (UTIs). A phylogenetic group with a high virulence content becomes a worldwide health concern. Resistance to antimicrobial agents increasingly complicates the management of E. coli extraintestinal infections, as a major source of illness, death, and increased health care costs. The aim of this study was to determine the virulence content and the antimicrobial susceptibility pattern of different uropathogenic E. coli (UPEC) phylogenetic groups in Ahvaz, Iran.
Methods: Phylogenetic groups, virulence-associated genes (VAGs), and antimicrobial susceptibility tests were detected by molecular and phenotypic methods in a total of 232 clinically well-characterized E. coli strains, isolated from two collections of patients with hospital-acquired (HA) and community-acquired (CA) UTIs.
Results: Our results revealed that among 232 UPEC strains, the most frequent phylogenetic group was phylogroup D (58%) with the greatest content in virulence factors, including kpsM (23%), neuA (76.3%, capsule), cnf (29.6%, toxin), and Pap (54.8%, adhesin). Phylogroups D and, to a lesser extent, B2 were the most drug-resistant phylogroups. In addition, phylogroup D was responsible for the majority of HA (64.7%) and CA (48.4%) infections.
Conclusion: Among UPEC strains causing UTIs, different phylogroups, through different VAGs, could cause severe infection. Knowledge about the distribution of the four functional groups and VAGs belonging to these phylogroups would significantly help to confine and prevent the development of lethal infection caused by these strains.
Keywords: uropathogenic Escherichia coli, drug resistance, virulence factors, Iran
Introduction
Urinary tract infection (UTI) is one of the most common bacterial infections, occurring in healthy people and immunocompromised patients.1 Approximately, 70–95% of community-acquired (CA) cases and 50% of all nosocomial infections related to UTI are caused by Escherichia coli.2 Uropathogenic E. coli (UPEC) strains are an economic burden for both communities and hospital resources. It has been estimated that UTIs cost about $6 billion for national health resources per year.3 Pathogenicity is the ability of an agent to cause disease, and pathogenic bacteria possess several factors that enable them to enhance their virulence. UPEC is a member of extraintestinal pathogenic E. coli (ExPEC) and through a variety of virulence-associated genes (VAGs) can produce extraintestinal diseases, such as pyelonephritis, cystitis, sepsis, and neonatal meningitis.1,2 This bacterium has several virulence-related functions. These virulence-related functional groups are adhesins, microcins, toxins, and capsules, which overcome the host defenses, invade host tissues, and finally trigger a local inflammatory response in the host. Based on many studies of the virulence content and phylogeny of UPEC, four phylogenetic groups have been described (A, B1, B2, and D), and also a significant heterogeneity has been shown among these strains.4 Among these phylogroups, phylogroup B2 strains are usually the most common causing UTI. Different phylogroups have a variety of VAGs.5,6 However, phylogroups A and B1 usually lack a distinct virulence profile.5,6
Antimicrobial resistance is defined as the ability of an organism to resist the action of an antimicrobial agent to which it was previously susceptible. The frequent antimicrobial resistance to bacterial infections occurs through indiscriminate use of antibiotics, default in treatment, and microbial characteristics. UTI can be controlled using an antimicrobial agent, but the increase in antimicrobial resistance has become a major concern, which promotes multiple drug resistance in UPEC, in both CA and hospital-acquired (HA) infections.4,7 In addition, according to clinical studies, antibiotic resistance and VAGs have been induced in some E. coli strains in order to become virulent. Then, study of strain characterization, phylogenetic properties, and antibiotic susceptibility would improve our understanding of the epidemiology and virulence of this pathogen, and will allow the development of a rapid assay for monitoring UPEC.7 The aim of this study was to phylotype and understand the virulence and antibiotic susceptibility pattern of E. coli causing UTI in Ahvaz, Iran. Thus, we studied 232 clinically well-described E. coli isolates from patients with UTI to determine the phylogroups, VAGs, and resistance to antimicrobial agents, and also to find the correlation of these factors to HA and CA infection.
Materials and methods
Ethics statement
The study was approved by the Research Ethics Committee of the Ahvaz Jundishapur University of Medical Sciences (No. IR.AJUMS.REC.1395.222.), Ahvaz, Iran. Written informed consent was obtained from all UTI patients.
A total of 232 UPEC strains, obtained from patients with UTI determined as HA and CA infection in Ahvaz, were collected and stored between 2013 and 2014. E. coli colonies were kept at −80 °C in trypticase soy broth, supplemented with 5% glycerol, until further use.3
Antimicrobial resistance
Antimicrobial susceptibility testing for 11 antibiotics, including ampicillin (10 μg), ceftazidime (30 μg), ceftriaxone (30 μg), ciprofloxacin (5 μg), gentamycin (10 μg), nalidixic acid (30 μg), trimethoprim–sulfamethoxazole (1.25+23.75 µg), amikacin (30 μg), nitrofurantoin (300 μg), tetracycline (30 μg), and imipenem (10 μg), was performed using the disk diffusion method, as recommended and specified by the Clinical and Laboratory Standards Institute.8 Commercial antimicrobial disks from Roscoe Co. (Denmark) were used in this study. In addition, E. coli ATCC 25922 was used for quality control.8,9 Six groups of antibiotics (MAST, CO, UK), including amino-penicillin, carbapenem, third generation of cephalosporin, tetracycline, aminoglycoside, and quinolone, were selected for susceptibility testing using the disk diffusion method. All 232 isolates were analyzed, E. coli ATCC 25922 was used for quality control, and the results were interpreted according to the Clinical and Laboratory Standards Institute.8,9
Phylotyping of UPEC by triplex PCR
ChuA, YjaA, and TspE4, three genetic markers, were amplified by the triplex PCR assay for classification of phylogroups of UPEC isolates.10 Four phylogroups of E. coli (A, B1, B2, and D) can be distinguished by this triplex PCR method. The primer sequences used for PCR have been taken from studies by Clermont et al.10 PCR products were run on 2% agarose gel. Based on the results obtained from the triplex PCR assay, all isolates are classified into the four phylogroups, based on the presence or absence of specific PCR-amplified fragment patterns, as follows: group B2 (chuA+, yjaA+, TSPE4±), group B1 (chuA–, yjaA±, TSPE4+), group D (chuA+, yjaA–, TSPE4±), and group A (chuA–, yjaA , TSPE4–).10,11
Virulence genotyping
In this study, using gene-specific primer and PCR techniques, we analyzed four functional groups – adhesins (draD, type 1 fimbriae (fimH), P fimbriae (papC and papG Allele I, II and III), and S fimbriae (sfa A)), microcins (mcmA, mchB, mciA, mceA, and cvaC), capsules (kpsM and neuA), and toxins (cnf (cytotoxic necrotizing factor), hlyA (alpha hemolysin), and sat (secreted auto transporter toxin)) – in UPEC strains against wild-type E. coli K12. Amplification of the VAGs was carried out using primer sets selected from previous published works (Table1).4,12–16 The multiplex PCR assay was designed for 15 VAGs (papG allele III, sfa, and kpsM; papC, papG allele I, and papG allele II; mciA, neuA, and draD; Cnf-1, mcmA, mchB, and fimH; mceA and cvaC) while the uniplex PCR assay was performed for hlyA and sat, and reactions were performed in a final volume of 25 µl, containing 50 ng template DNA, 10 pmol of each oligonucleotide primer pair (CinaGen, Germany), and 15 µl of master mix (CinaGen PCR master kit). Reaction mixtures were subjected to the following conditions in a thermal cycler (Eppendorf, Germany): 5 min at 94 ºC, 30 cycles of 30 s at 94 ºC, and 30 s at 60 ºC for neuA, fimH, mchB, cnf, mcmA, mciA, draD, and Sat; 67 ºC for PapG Allele I, PapG Allele II, PapG Allele III, kpsM, papC, hlyA, and sfa; and 52 ºC for cvaC and mceA, followed by 30 s at 72 ºC, with a final cycle of 7 min at 72 ºC, and a final hold at 10 ºC. The PCR products were run on 2% agarose gel. Amplicons were stained with ethidium bromide and photographed using the Gel Doc system, and their size was determined by comparison to a 100-bp DNA size marker (Invitrogen; Thermo Fisher Scientific, Waltham, MA, USA).
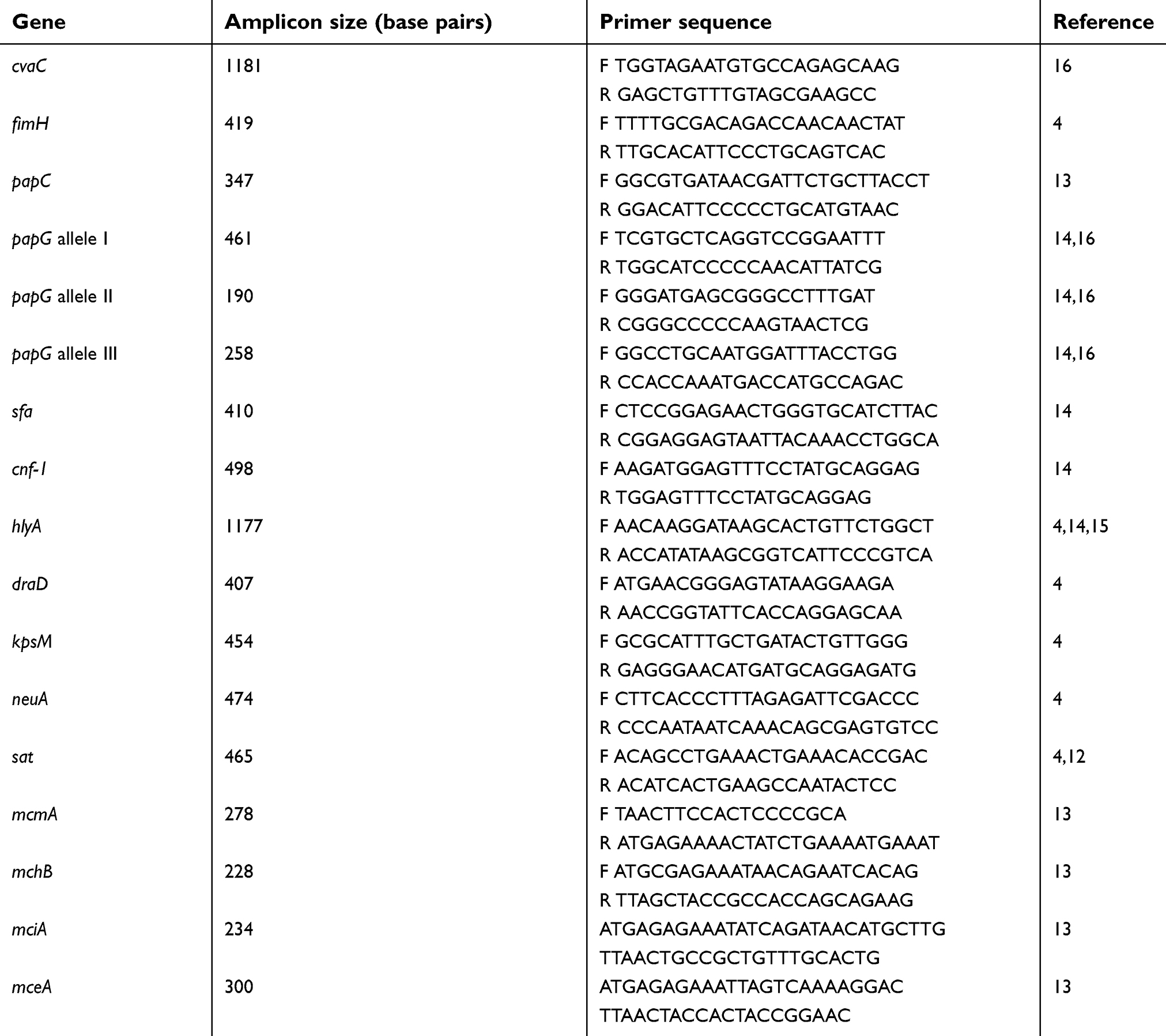 |
Table 1 The primer sequences used for the PCR-based amplification of the VAGs in Escherichia coli isolated from UTI patients |
Statistical analysis
In order to identify significant associations between phylogroups of UPEC isolates and the presence of VAGs, and to detect any significant correlation among E. coli VAGs, Fisher’s exact test (SPSS version 19.0; IBM Corporation, Armonk, NY, USA) was performed. p<0.05 was considered statistically significant.
Results
Antibiotic susceptibility test
Among 232 UPEC strains, 47% were completely susceptible to antimicrobial agents. Antimicrobial resistance for all antibiotics, except ceftriaxone, was less frequent among isolates from the first collection (HA isolates) than among those from the second (CA isolates). For most antibiotics, except ampicillin, ceftriaxone, trimethoprim–sulfamethoxazole, and amikacin, no significant differences in antimicrobial resistance rates were found, either among CA or HA isolates. However, resistance rates among CA isolates were greater than those for HA isolates. The results of drug resistance in different phylogenetic groups of UPEC isolates are cited in Table 2.
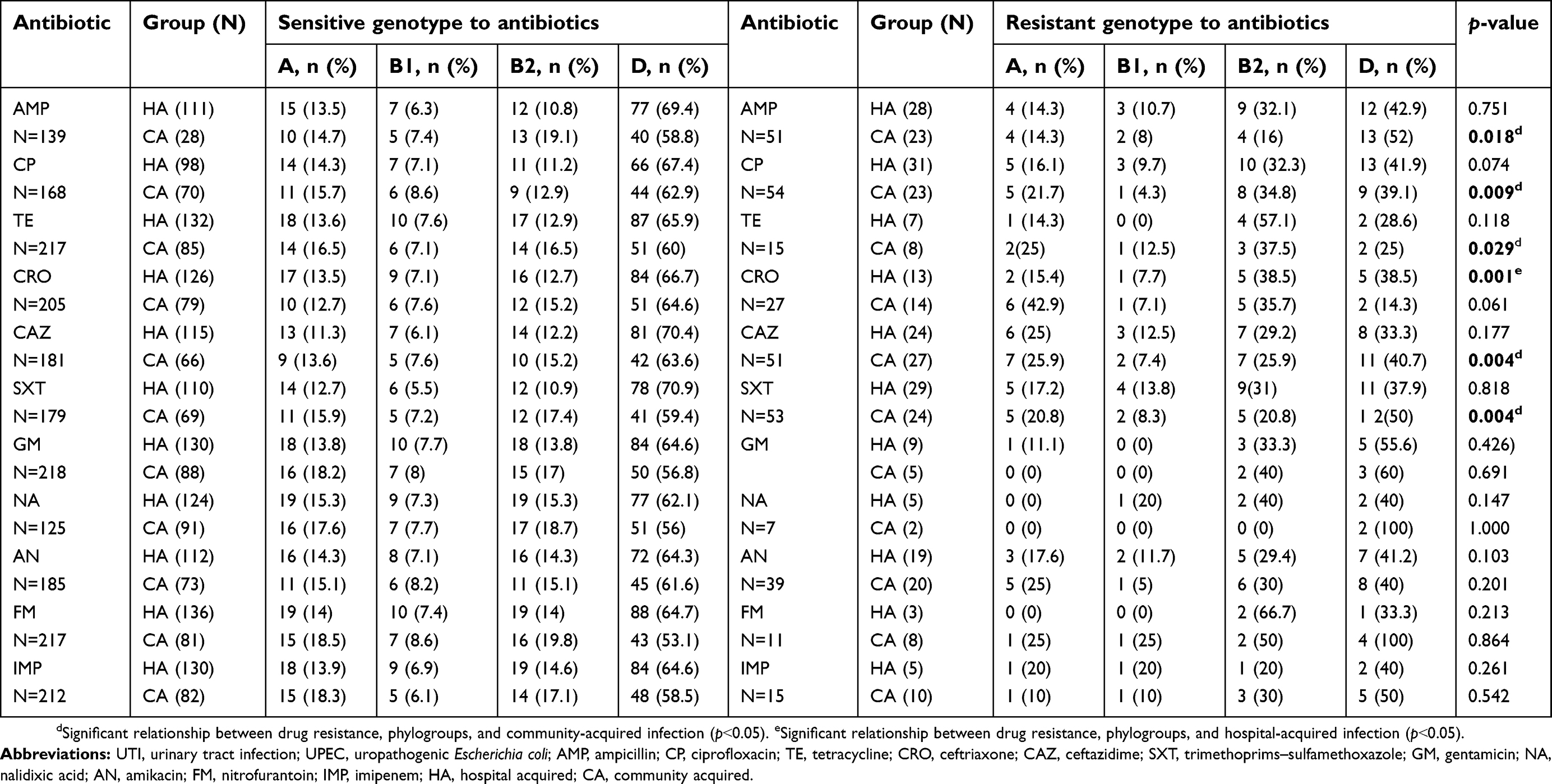 |
Table 2 Antibiotic resistance of different UPEC phylogroups in CA and HA UTI |
Phylogenetic groups
Based on the results obtained from the triplex PCR assay, the prevalence of the four main phylogroups (A, B1, B2, and D) differed in E. coli isolates responsible for UTI. The frequency of phylogroup D (58%) was higher than other phylogroups, followed by 16% for group A, 18% for group B2, and 7.75% for group B1. Phylogroup data were analyzed and are presented in Table 3.
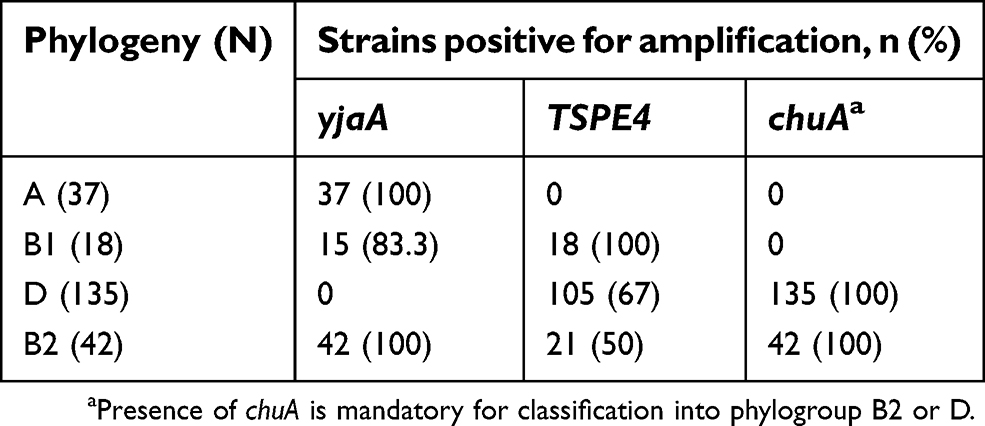 |
Table 3 Phylogeny distribution among the isolates from the urinary tract infection patients |
Phylogenetic and VAG distribution in CA and HA isolates
The distribution of phylogeny and related VAGs, selected for the characterization of the UPEC isolates, showed different and distinctive patterns in the two collections including HA and CA infections (Table 4). The rate of phylogroup D among HA isolates was greater than for CA isolates. The type of VAGs is categorized into four groups, including adhesin/fimbria, microcin genes, cytotoxins, and protectins. Table 3 highlights the role of these virulence factors in UPEC strains. Among the aforementioned factors in Table 4, adhesins factors such as sfa and draD were detected more in the CA population, compared to the HA population.
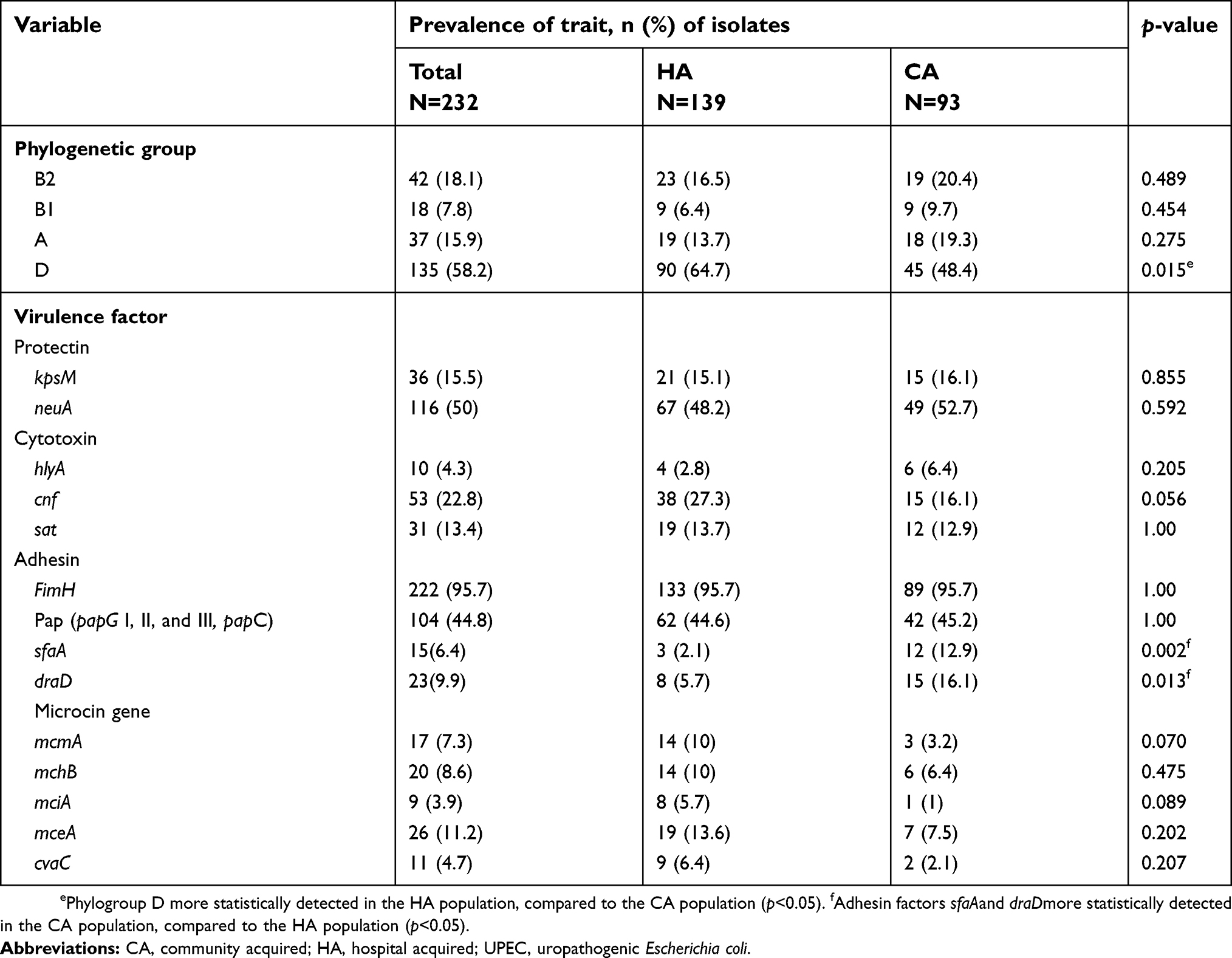 |
Table 4 Distribution of phylogenetic groups and virulence factors among the 232 UPEC isolates in HA and CA infections |
Correlations between phylogroups and VAGs
The frequency of VAGs among UPEC ranged from 3.9% (kpsM) to 95.7% (fimH). In each UPEC strain, 17 VFGs were detected. Based on data from the multiplex and uniplex PCR assays for amplifying the total 17 VAGs in 232 UPEC samples, the distribution and correlation of the phylogenetic groups and VFsare described in detail in Table 5. Some VAGs, including kpsM, hlyA, and sfaA, were absent in phylogenetic groups A and B1, while these VAGs were present in phylogenetic groups D and B2. In addition, other VAGs with high frequency were observed more in phylogenetic groups B2 and D.
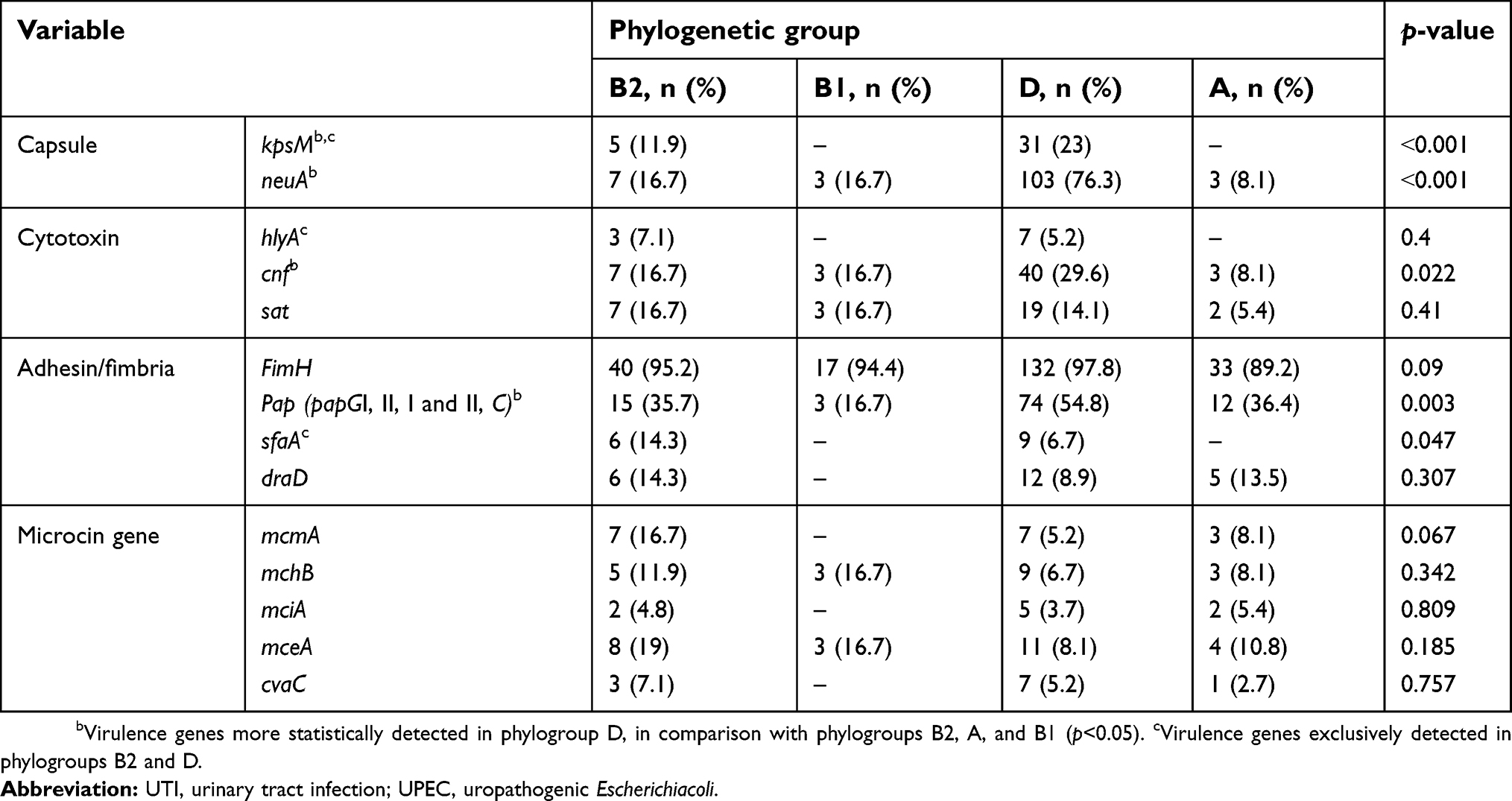 |
Table 5 Distribution of virulence genes/phylogenetic groups among the UPEC strain in the UTI patients |
Discussion
Pathogenic and commensal strains of E. coli based on phylogenetic analyses are differently sorted into four major phylogenetic groups (phylogroups), named A, B1, B2, and D.17 The VAG content (four functional groups) in UPEC is usually varied. The distribution and diversity of the VAGs are usually related to the phylogenetic groups. This work was the first study to characterize the phylogenetic group and VAG content in UPEC strains in Ahvaz, Iran. In the present study, phylogenetic groups were evaluated by a multiplex PCR technique, and so the antimicrobial susceptibility, virulence-associated traits, and correlation of these factors with their infection source were investigated in 232 UPEC strains. Based on the recent phylogenetic studies, contrary to commensal strains (phylogroups A and B1), UTIs are mostly derived from phylogroup B2 and phylogroup D of E. coli strains. Approximately, the distribution of some initial colonization factors, such as fimH, was equal in the all phylogenetic groups, but some VAGs in UPEC strains, including the genes encoding fimbria P (pap, pyelonephritis-associated pilus), sfa, and draD, play a significant role in UTI prevalence, since they promote the colonization of proximal and distal tubular cells from human kidney.7,18–21 In the present study, phylogroup D (58%) was detected as the most common phylotype of E. coli strains isolated from patients with UTI, while phylogroups B2, A, and B1 were detected in 18.1%, 15.9%, and 7.8% of UPEC strains, respectively. Nevertheless, unlike our study, in the study conducted by Lara et al22phylogroups B2 (43%) and D (38%) were the most common phylogenetic groups responsible for UTI caused by ExPEC strains.22 Also, our results were not consistent with the studies performed by Johnson et al and Moreno et al concerning phylogroups in UPEC, showing the highest prevalence in phylogroup B2.6,21 Our findings are in agreement with those of previous studies performed by Poey et al4 to detect the virulence profile of UPECin pregnant women and children with UTI, reporting that phylogroup D was responsible for the majority of UTI.4
Overall, in this work based on PCR analysis, the frequency of VAGs ranged from 3.9% for mciA to 95.7% for FimH in various phylogenetic groups. Our data demonstrated that some VAGs, including adhesion factors (kpsM, sfaA, draD), hlyA, and microcin genes (mcmA, mciA, and cvaC), were absent in phylogroup B1, while kpsM, hlyA, and sfaA were absent in phylogroup A (Table 4). Among various phylogroups, phylogroup D strains demonstrated the highest virulence scores, and consequently the most diverse virulence content. In this phylogroup, significant differences were observed in some virulence scores, such as kpsM, neuA, cnf, and pap with p<0.001, p<0.001, p=0.022, and p=0.003, respectively (Table 4). The greatest distributions of adhesion factors, fimH, pap, draD, and sfaA, were seen in phylogroup D strains, which may justify the highest frequency of UTI caused by this phylogroup. Similar to our study, in the study performed by Lara et al22 the frequency of pap in phylogroups B2 and D was 48% and 53%, respectively.22
According to studies conducted by Tiba et al and Usein et al, high frequencies in some VAGs, including sfa, hlyA, and papC were observed in UPEC strains.23,2424 In our study, in agreement with those of previous studies performed by Farshad et al and Dormanesh et al in Iran, cnf1 with a frequency of 22.8% was more prevalent than sat and hlyA.25–2727 In addition, in the study performed by Johnson et al in 243 ExPEC isolates from human, phylogroups B2 and D were responsible for most of the infection, with a frequency of 63% and 26%, respectively, and the VAG prevalence ranged from 0.4% to 98%.2727 Our results (Table 4) demonstrated a frequency of kpsM, neuA, cnf1, and pap in all phylogroups and their significant association with phylogroup D, while sfa was more related to phylogroup B2. However, other VAGs (eg, papC, fimH, cnf, sat, draD, mceA, papG and C, mcmA, mciA, mchB, and neuA) had more dispersed incidence in four phylogroups with different prevalences. Then, the majority of VAGs were concentrated within phylogroup D. Then, in this work, the pathogenicity of phylogenetic groups was as follows: D˃B2>A>B1.
Analysis of phylogeny and VAGs was also performed in two HA and CA collections, showing that phylogroup D and cnf1 encoding for cytotoxin necrotizing factor are more related to HA isolates, while sfaA encoding for S fimbriae and draD encoding for adhesion factors were found to be associated with CA rather than HA isolates (Table 5). Similar to other studies, our study showed that pap encoding for P fimbriae was found to be related to CA rather than HA isolates.3 Thus, overall, the adherence potency of UPEC for urinary tract cells in the CA isolates was greater than that in HA isolates, due to adhesion factors. Moreover, our findings indicate that both HA and CA infections can be due to different virulent strains. Resistance to some antibiotics, such as ampicillin, ceftazidime, trimethoprim–sulfamethoxazole, tetracycline, and ciprofloxacin, were associated with CA rather than HA isolates. Statistically significant differences were seen in relation to drug resistance and phylogroups in the CA populations. The high level of resistance among CA isolates, compared to the HA isolates, showed that antibiotic resistance in UPEC is correlated with and due to the uncontrolled use of antibiotics outside the hospital.28 The results of this study revealed that phylogenetic groups D and B2 of UPEC isolates are resistant to antibiotics rather than other phylogenetic groups. VAGs were found to be associated with the two phylogroups D and B2. The eminent presence of VAGs associated with phylogroups B2 and D confirms that phylogroup D and, to a lesser extent, phylogroup B2 are two phylogroups that contain potent human ExPEC isolates causing UTI. Thus, the potential virulence characteristics of different phylogroups could help the bacteria to overcome host defense mechanisms and ultimately cause infection.29 Lactobacillus products, as nonantibiotic alternatives, could be used to prevent recurrent UTIs. This could help to reduce the rising rates of antibiotic resistance.30 An interesting and important area for future work would be to study E. coli isolates in women with frequent/recurrent UTIs that can be caused by adherence of vaginal isolates in different E. coli strains or the survival of E. coli in the bladder through a progression of intracellular bacterial communities.31,32
Conclusion
Among UPEC strains causing UTI, different phylogroups, through different VAGs, could cause severe infection. Knowledge about the distribution of the four functional groups and VAGs belonging to these phylogroups would significantly help to confine and prevent the development of lethal infection caused by these strains.
Abbreviations list
AMP, ampicillin; CP, ciprofloxacin; GM, gentamicin; NA, nalidixic acid; SXT, trimethoprim–sulfamethoxazole; TE, tetracycline; CAZ, ceftazidime; CRO, ceftriaxone; GM, gentamicin; AN, amikacin; IMP, imipenem; CA, community acquired; HA, hospital acquired; VAG, virulence-associated gene; UTI, urinary tract infection; ExPEC, extra-intestinal pathogenic Escherichia coli.
Acknowledgments
This work is approved by the Health Research Institute, Infectious and Tropical Diseases Research Center (No. 92106). Special thanks to the Research Affairs, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran, for financial support.
Author contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to accountable for all aspects of the works.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Vander Bij AK, Peirano G, Pitondo-Silva A, Pitout JDD. The presence of genes encoding for different virulence factors in clonally related Escherichia coli that produce CTX-Ms. Diagn Microbiol Infect Dis. 2012;72:297–302. doi:10.1016/j.diagmicrobio.2011.12.011
2. Lau SH, Reddy S, Cheesbrough J, et al. Major UPEC strain isolated in the northwest of England identified by multilocus sequence typing. J Clin Microbiol. 2008;46(3):1076–1080. doi:10.1128/JCM.02065-07
3. Rasmussen LS, Ejrnaes K, Lundgren B, Hammerum AM, Moller NF. Virulence factors and phylogenetic grouping of Escherichia coli isolates from patients with bacteraemia of urinary tract origin relate to sex and hospital- vs. community-acquired origin. Int J Med Microbiol. 2012;302(3):129–134. doi:10.1016/j.ijmm.2012.03.002
4. Poey ME, Albini M, Saona G, Lavina M. Virulence profiles in uropathogenic Escherichia coli isolated from pregnant women and children with urinary tract abnormalities. Microb Pathog. 2012;52:292–301. doi:10.1016/j.micpath.2012.02.006
5. Leopold SR, Sawyer SA, Whittam TS, Tarr PI. Obscured phylogeny and possible recombinational dormancy in Escherichia coli. BMC Evol Biol. 2011;11:183. doi:10.1186/1471-2148-11-183
6. Moreno E, Johnson JR, Perez T, Prats G, Kuskowski MA, Andreu A. Structure and urovirulence characteristics of the fecal Escherichia coli population among healthy women. Microb Infect. 2009;11:274–280. doi:10.1016/j.micinf.2008.12.002
7. Clermont O, Olier M, Hoede C, et al. Animal and human pathogenic Escherichia coli strains share common genetic backgrounds. Infect Genet Evol. 2011;11:654–662. doi:10.1016/j.meegid.2011.02.005
8. Reller LB, Weinstein M, Jorgensen JH, Ferraro MJ. Antimicrobial susceptibility testing: a review of general principles and contemporary practices. Clin Infect Dis. 2009;49(11):1749–1755. doi:10.1086/647952
9. Ranjbar R, Haghi-Ashtiani MT, Jafari NJ, Abedini M. The prevalence and antimicrobial susceptibility of bacterial uropathogens isolated from pediatric patients. Iran J Publ Health. 2009;38(2):134–138.
10. Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl Environ Microbiol. 2000;66:4555–4558. doi:10.1128/AEM.66.10.4555-4558.2000
11. Gordon DM, Clermont O, Tolley H, Denamur E. Assigning Escherichia coli strains to phylogenetic groups: multi-locus sequence typing versus the PCR triplex method. Environ Microbiol. 2008;10:2484–2496. doi:10.1111/j.1462-2920.2008.01669.x
12. Ejrnaes K, Stegger M, Reisner A, et al. Characteristics of Escherichia coli causing persistence or relapse of urinary tract infections: phylogenetic groups, virulence factors and biofilm formation. Virulence. 2011;2:528–537. doi:10.4161/viru.2.6.18189
13. Azpiroz MF, Poey ME, Lavina M. Microcins and urovirulence in Escherichia coli. Microb Pathog. 2009;47:274–280. doi:10.1016/j.micpath.2009.09.003
14. Moreno E, Planells I, Prats G, Planes AM, Moreno G, Andreu A. Comparative study of Escherichia coli virulence determinants in strains a using urinary tract bacteremia versus strains causing pyelonephritis and other sources of bacteremia. Diagn Microbiol Infect Dis. 2005;53:93–99. doi:10.1016/j.diagmicrobio.2005.05.015
15. Ewers C, Li G, Wilking H, et al. Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: how closely related are they? Int J Med Microbiol. 2007;297:163–176. 33. doi:10.1016/j.ijmm.2007.01.003
16. Momtaz H, Karimian A, Madani M, et al. Uropathogenic Escherichia coli in Iran: serogroup distributions, virulence factors and antimicrobial resistance properties. Ann Clin Microbiol Antimicrob. 2013;12(1):8. doi:10.1186/1476-0711-12-8
17. Doumith M, Day MJ, Hope R, Wain J, Woodford N. Improved multiplex PCR strategy for rapid assignment of the four major Escherichia coli phylogenetic groups. J Clin Microbiol. 2012;50(9):3108–3110. doi:10.1128/JCM.01468-12
18. Russo TA, Johnson JR. Proposal for a new inclusive designation for extraintestinal pathogenic isolates of Escherichia coli: ExPEC. J Infect Dis. 2000;181(5):1753–1754. doi:10.1086/315418
19. Korhonen TK, Parkkinen J, Hacker J, et al. Binding of Escherichia coli S fimbriae to human kidney epithelium. Infect Immun. 1986;54(2):322–327.
20. Marre R, Kreft B, Hacker J. Genetically engineered S and F1C fimbriae differ in their contribution to adherence of Escherichia coli to cultured renal tubular cells. Infect Immun. 1990;58:3434–3437.
21. Johnson JR, Kuskowski MA, Gajewski A, et al. Extended virulence genotypes and phylogenetic background of Escherichia coli isolates from patients with cystitis, pyelonephritis, or prostatitis. J Infect Dis. 2005;19(1):46–50. doi:10.1086/426450
22. Lara FBM, Nery DR, de Oliveira PM, et al. Pereira virulence markers and phylogenetic analysis of Escherichia coli strains with hybrid EAEC/UPEC genotypes recovered from sporadic cases of extraintestinal infections. Front Microbiol. 2017;8:146. doi:10.3389/fmicb.2017.00146
23. Tiba MR, Yano T, Leite Dda S. Genotypic characterization of virulence factors in Escherichia coli strains from patients with cystitis. Rev Inst Med Trop Sao Paulo. 2008;50(5):255–260. doi:10.1590/S0036-46652008000500001
24. Usein CR, Damian M, Tatu-Chitoiu D, et al. Prevalence of virulence genes in Escherichia coli strains isolated from Romanian adult urinary tract infection cases. J Cell Mol Med. 2001;5(3):303–310. doi:10.1111/j.1582-4934.2001.tb00164.x
25. Moreno E, Andreu A, Pigrau C, Kuskowski MA, Johnson JR, Prats G. Relationship between Escherichia coli strains causing acute cystitis in women and the fecal E. coli population of the host. J Clin Microbiol. 2008;46:2529–2534. doi:10.1128/JCM.00813-08
26. Farshad S, Ranjbar R, Japoni A, Hosseini M, Anvarinejad M, Mohammadzadegan R. Microbial susceptibility, virulence factors, and plasmid profiles of uropathogenic Escherichia coli strains isolated from children in Jahrom, Iran. Arch Iran Med. 2012;15(5):312–316.
27. Dormanesh B, Safarpoor Dehkordi F, Hosseini S, et al. Virulence factors and o-serogroups profiles of uropathogenic Escherichia coli isolated from Iranian pediatric patients. Iran Red Crescent Med J. 2014;16(2):e14627. doi:10.5812/ircmj.14627
28. Johnson JR, Sannes MR, Croy C, et al. Antimicrobial drug–resistant Escherichia coli from humans and poultry products, Minnesota and Wisconsin, 2002–2004. Emerg Infect Dis. 2007;13(6):838–846. doi:10.3201/eid1306.061576
29. Monitoring DIAR 2016. Use of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from food animals, food, and humans in Denmark.
30. Ng QX, Peters C, Venkatanarayanan N, Goh YY, Ho CY, Yeo WS. Use of Lactobacillus spp. to prevent recurrent urinary tract infections in females. Med Hypotheses. 2018;114:49–54. doi:10.1016/j.mehy.2018.03.001
31. Schaeffer AJ, Jones JM, Falkowski WS, Duncan JL, Chmiel JS, Plotkin BJ. Variable adherence of uropathogenic Escherichia coli to epithelial cells from women with recurrent urinary tract infection. J Urol. 1982;128(6):1227–1230.
32. Mulvey MA, Schilling JD, Hultgren SJ. Establishment of a persistent Escherichia coli reservoir during the acute phase of a bladder infection. Infect Immun. 2001;69(7):4572–4579. doi:10.1128/IAI.69.7.4572-4579.2001
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.