Back to Journals » Cancer Management and Research » Volume 11
Urinary 8-hydroxydeoxyguanosine in relation to XRCC1 rs25487 G/A (Arg399Gln) and OGG1 rs1052133 C/G (Ser326Cys) DNA repair genes polymorphisms in patients with chronic hepatitis C and related hepatocellular carcinoma
Authors Mahmoud AA, Hassan MH ![]() , Ghweil AA, Abdelrahman A
, Ghweil AA, Abdelrahman A ![]() , Mohammad AN
, Mohammad AN ![]() , Ameen HH
, Ameen HH
Received 22 March 2019
Accepted for publication 7 May 2019
Published 10 June 2019 Volume 2019:11 Pages 5343—5351
DOI https://doi.org/10.2147/CMAR.S209112
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Harikrishna Nakshatri
Aida A Mahmoud,1 Mohammed H Hassan,2 Ali A Ghweil,3 Amany Abdelrahman,4 Asmaa N Mohammad,5 Hesham H Ameen6
1Medical Biochemistry Department, Faculty of Medicine, Sohag University, Sohag, Egypt; 2Medical Biochemistry Department, Faculty of Medicine, South Valley University, Qena, Egypt; 3Tropical Medicine and Gastroenterology Department, Faculty of Medicine, South Valley University, Qena, Egypt; 4Medical Physiology Department, Faculty of Medicine, Sohag University, Sohag, Egypt; 5Tropical Medicine and Gastroenterology Department, Faculty of Medicine, Sohag University, Sohag, Egypt; 6Clinical Pathology, Faculty of Medicine, Al-Azhar University (Assiut Branch), Assiut, Egypt
Background and aim: DNA repair represents a protective mechanism against cell injury and cancer. 8-hydroxy-deoxyguanosine (8-OHdG) is the main ROS-induced DNA mutation. The current study aimed to evaluate urinary 8-OHdG levels in patients with chronic hepatitis C virus (HCV) and its related hepatocellular (HCC) and correlate its level to XRCC1 rs25487 G/A and OGG1 rs1052133 C/G gene polymorphisms.
Materials and methods: Urinary 8-OHdG assays were performed using HPLC technique, and XRCC1 rs25487 G/A and OGG1 rs1052133 C/G gene polymorphisms were analyzed by PCR using confronting two-pair primer method (PCR-CTPP) in 200 subjects allocated into 50 chronic HCV patients, 50 HCV-related HCC patients, and 100 controls.
Results: There were significantly increased urinary 8-OHdG levels in HCV-related HCC and chronic HCV patients when compared with the controls (P<0.05 for all). Urinary 8-OHdG was associated with the tumor spread. Regarding, XRCC1 (Arg399Gln), AA (Gln/Gln) genotype and A-allele were more frequent in HCC and chronic HCV patients than in the controls (P<0.05). ORs (95%CI) using the dominant and the recessive genetic models were; 2.1 (1.1–4.1), P=0.032 and 1.9 (1–3.6), P=0.043 respectively. For OGG1 (Ser326Cys), GG (Cys/Cys) genotype and G-allele were increased significantly in chronic HCV and HCC patients compared to the controls (P<0.05). ORs (95%CI) under the dominant and the recessive genetic models were; 2.1 (1.1–4.1), P=0.032 and 1.9 (1–3.8), P=0.049 respectively. Additionally, XRCC1 (AA) and OGG1 (GG) genotypes had significantly increased urinary 8-OHdG levels among patients (P<0.05).
Conclusions: XRCC1 (AA) and OGG1 (GG) could be considered as possible genotypic risk factors for HCV- related HCC development which were associated with significantly high urinary 8-hydroxy-deoxyguanosine levels, thus urinary 8-OHdG could be considered as non-invasive marker in follow-up chronic HCV progression into HCC.
Keywords: urinary 8-hydroxy-deoxyguanosine, high performance liquid chromatography, confronting two-pair primers, XRCC1 and OGG1 SNPs, DNA repair, hepatocellular carcinoma
Introduction
About 50–80% of patients infected by hepatitis C virus (HCV) develop chronic inflammatory disease process and hepatocellular carcinoma (HCC).1 Chronic hepatitis C is the leading indication for liver transplantation in many parts of the world.2,3 The number of people who are positive for HCV RNA was estimated as 1% (range: 0.8–1.14%) or approximately 71 million individuals (range: 62–79 million) worldwide.4
HCC commonly occurs in the setting of chronic injury and inflammation that promote DNA damage and chromosomal aberrations. DNA damage response (DDR) pathways coordinate DNA repair, cell cycle arrest, and cell death or senescence. Aberrations of DDR may affect the genes integrity, leading to hepatic injury and HCC development.5
Base excision repair and the nucleotide excision repair are two main DNA repair pathways. X-ray repair cross-complementing group 1 (XRCC1) and 8-oxoguanine DNA glycosylase (OGG1) are genes involved in the base excision repair pathway. Single nucleotide polymorphisms (SNPs) in the genes involved in DNA repair pathways could influence susceptibility to oxidative damage and carcinogenesis.6
The XRCC1 gene is located on chromosome 19 (q13.2-13.3) and spans a genetic distance of 33 kb. It has17 exons, encoding a protein consisting of 633 amino acids and its molecular weight about 70kDa.7 This protein interacts with three main enzymes involved in DNA single-strand break repair; poly(ADP-ribose) polymerase-1, DNA polymerase h and DNA ligase III. XRCC1 rs25487 G/A gene polymorphism is located at the C terminus of the BRCT I domain and results in two different amino acids (Arg399Gln). The A-allele encodes for glutamine while the G-allele encodes for arginine.8
The OGG1 gene is located on the short arm of chromosome 3 (3p25) and encodes 8-hydroxyguanine glycosylase that excises 8-Oxodeoxyguanosine (8-OHdG) that results from exposure to reactive oxygen species.9 An elevated 8-OHdG content in DNA has been shown to increase the cancer risk.10 OGG1 gene polymorphism rs1052133 C/G (Ser326Cys) codes for two different amino acids at this site; C-allele codes for serine (Ser) and G-allele codes for cysteine (Cys). OGG1 326 Cys/Cys was reported as a low-expression genotype for 8-hydroxyguanine glycosylase and associated with human cancer susceptibility.11 Urinary 8-OHdG is a sensitive marker of the DNA damage and a prognostic factor in HCC. A strong association was found between 8-OHdG expression and poor patient survival and clinical staging in HCC patients.12
As DNA repair plays a protective mechanism against cellular injury and cancer, the present study aimed to investigate the oxidative DNA damage in the form of urinary 8-OHdG levels in chronic HCV infection and HCV-related HCC, in addition to, two DNA repair enzymes genes polymorphisms rs25487 G/A (Arg399Gln) of XRCC1 and rs1052133 C/G (Ser326Cys) of OGG1 genes, since there is no literature that directly points to correlation between oxidative damage levels and polymorphisms in HCC.
Materials and methods
Participants
The current cross-sectional, case-control, hospital-based study has been conducted on 200 subjects categorized into three groups: Group A included 50 chronic HCV patients without cirrhosis, group B included 50 patients with HCV-related HCC recruited from the Tropical Medicine Departments, Sohag and South Valley University Hospitals, Egypt. Group C included 100 healthy individuals who were unrelated to patients, with no history of liver disease (negative for HBsAg and HCV antibody with normal liver function tests and normal abdominal ultrasound), age and sex-matched to the patients as a control group. Selected controls should not take any type of medications through the last three months to avoid any confounding factors that may affect the results. All enrollees gave written informed consent and the ethical committee of Sohag and Qena Faculties of Medicine, Egypt approved this study, and it was conducted in accordance with the Declaration of Helsinki. We adjusted the size of the sample to achieve 80% power and 5% confidence of significance (type I error).
Diagnosis of chronic HCV was based on persistence of HCV antibodies and HCV RNA in the serum of the patients for more than 6 months. Cirrhosis was diagnosed with liver fibroscan, abdominal sonography and biochemical evidence of parenchymal damage with or without endoscopic evidence of esophageal or gastric varices.13 The HCC diagnostic criteria were based on the guidelines proposed by European Association for the Study of the Liver (EASL).14 Patients with associated other comorbidities eg, kidney disease, were excluded from the study.
Laboratory workup
Routine laboratory investigations including HCV antibody, HBsAg, ALT, AST, serum albumin and prothrombin concentration were done for each participant.
Biochemical analysis of urinary 8-OHdG
Biochemical assays of of 8-OHdG in human urine samples were performed using high performance liquid chromatography (HPLC; Agilent Technologies 1200 Series, G1315D DAD; Agilent Technologies, Santa Clara, CA, USA). The urinary 8-OHdG levels were measured according to Kawai et al,15 with slight modifications, as done in a previously published work.16 Frozen urine samples were defrosted at room temperature and properly mixed. A 50 μL aliquot of urine was mixed with the same volume of diluents containing the ribonucleoside marker 8-OHdG (120 µg mL−1) and 4% acetonitrile (ACN) in 130 mM sodium acetate (pH 4.5) and 0.6 mM sulphuric acid. The pH of the diluted urine solution should be below 7 to obtain good separation. The urine solutions were incubated in a refrigerator at 5°C for 2–3 hours. The diluted urine samples were centrifuged at 13,000 rpm for 5 minutes using a cooling centrifuge. After centrifugation, the sample was injected automatically on an anion exchange column of HPLC (Zorbax Extend-C18 Analytical 4.6×150 mm 5-µ, Agilent Technologies, Santa Clara, CA, USA). The detector diode array detector (DAD) was at wavelength of 254 nm. The HPLC conditions for the HLC were as follows: mobile phase: 100% (NaOAC 50 mM), pH 4.6; column temperature: 35ºC; flow rate: 1.5 ml/min. The concentration of 8-OHdG was calculated from the obtained standard curves (Figure 1 and the Figure S1).
 | Figure 1 Calibration curves for high performance liquid chromatographic (HPLC) assay of 8-OHdG at concentrations 1, 5, 10, 15 and 20 mg/l. |
 | Figure S1 High performance liquid chromatography analysis curves of 8-hydroxy-deoxyguanosine for some included subjects. |
Blood collection and DNA extraction
Venous blood samples were collected in EDTA tubes, centrifuged at 3000 rpm. DNA was extracted from the puffy coat of blood samples using DNA extraction kit supplied by Jena Bioscience (Jena, Germany) as describedby the enclosed instructions.
Genotyping
DNA was amplified by the polymerase chain reaction (PCR) using the confronting two-pair primer method (PCR-CTPP).17 This method involve the using of confronting pairs of primers (four primers in all) to determine the genotype of a SNP according to the different sizes of the generated products, without the need for sequencing or restriction enzymes, thus saving time and cost. In addition,PCR-CTPP is better than PCR-RFLP in terms of the possibility of genotyping misclassification.17 The primersequences were shown in (Table 1). For the amplification, 25 µL reaction mixture containing 100–200 ng genomic DNA, 0.5 unit of Taq DNA polymerase, 2.5 of 10 x PCR buffer, 1 µL of 15 mM MgCl2, 3 µL of dNTPs mix (10 mM each) and 10 pM of each primer was used.
 | Table 1 Primers used for the target genomic DNA amplification |
The cycling conditions of XRCC1 rs25487 genotyping were 5 min of initial denaturation at 94°C, followed by 30 cycles of 30 seconds at 94°C, 1 minute at 64°C 45 seconds at 72°C, and a final extension of 10 minutes at 72°C. For OGG1 Ser326Cys genotyping, the cycling conditions were 10 minutes of initial denaturation at 95°C, followed by 30 cycles of 1 minute at 95°C, 1 minute at 62°C, 1 minute at 72°C and a final extension of 5 minutes at 72°C. The amplified DNA was visualized on a 2% agarose gel stained with 2 µL/100 mL ethidium bromide. The XRCC1 Arg399Gln polymorphism was genotyped as a 447-bp band for the G (Arg) allele, a 222-bp band for the A (Gln) allele, and a 630-bp common band. The OGG1 rs1052133 polymorphism was genotyped as a 252-bp band for the C (Ser) allele, a 194-bp band for the G (Cys) allele, and a 406-bp common band (Figures 2 and 3).
 | Figure 2 Detection of XRCC1 rs25487 A/G (Arg399Gln) single nucleotide polymorphism using the confronting two-pair primer method (PCR-CTPP). |
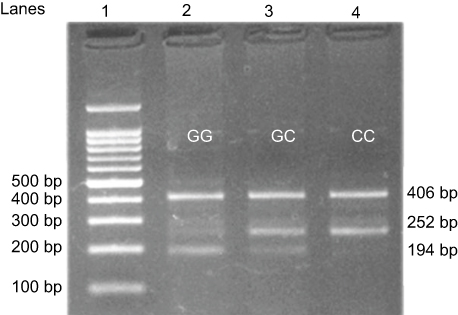 | Figure 3 Detection of OGG1 rs1052133 C/G (Ser326Cys) single nucleotide polymorphism using the confronting two-pairprimer method (PCR-CTPP). |
Statistical analysis
All statistical calculations were performed using the IBM SPSS version 20.0 software (IBM corporation, Armonk, NY, USA). The data were normally distributed. We represented the data as: mean ±SD or number and percent. Deviation from HWE (Hardy–Weinberg equilibrium) was tested by chi-squared analysis. The chi-squared test was also used to test the statistical significance of the association. Continuous data were compared using ANOVA with Tukey’s post-hoc test. We considered a two-tailed P-value of less than 0.05 statistically significant. Odds ratio (95%CI) was calculated using the dominant and the recessive genetic models.
Results
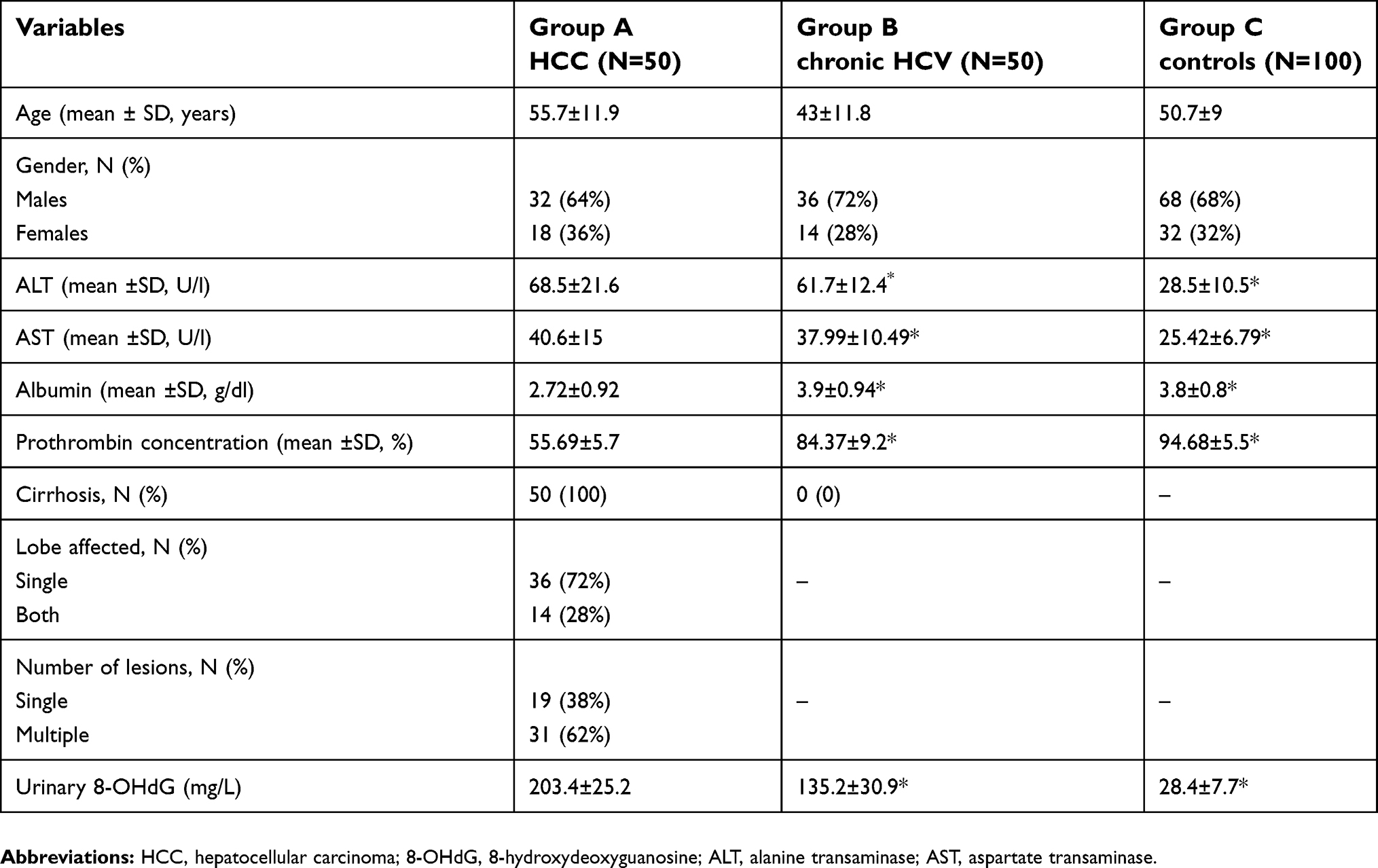
The current study has been carried out on 200 subjects who were allocated into three groups; group A included 50 patients with HCC: 32 patients (64%) were males and 18 (36%) were females; group B included 50 patients with chronic HCV: 72% were males and 28% were females. They were comparable with group C which included 100 healthy controls: 68 (68%) subjects were males and 32 (32%) subjects were females. The mean ages of the three groups were 55.7 years ±11.9, 43 years ±11.8 and 50.7 years ±9 respectively with non-significant differences regardingage and sex, indicating age and gender matching (Table 2). Their serum levels of liver enzymes, serum albumin and prothrombin concentration were presented in (Table 2)
 | Table 2 Demographic, clinical and laboratory data of the studied groups |
There were significantly higher mean ±SD urinary levels of 8-OHdG among group A (203.4 mg/L ±25.2) and group B (135.2 mg/L ±30.9) when compared with group C (28.4±7.7) (P<0.05 for both) with significantly higher levels among group A vs group B (<0. 05), (Table 2).
Genotypes and alleles distribution study revealed no deviation from HWE for XRCC1 rs25487 and OGG1 rs1052133 gene polymorphisms. XRCC1 rs25487 genotypes distribution showed a significantly higher frequency of AA and significantly lower GG genotypic frequency in chronic HCV and HCC patients vs controls (x2=7.4, P=0.024), with non-significant differences between the three groups regarding the AG genotype (P˃0.05). Additionally there were non-significant differences in genotypes distribution between HCC and chronic HCV patients (x2=0.24, P=0.89). Frequency of G-allele decreased significantly with significant increase in A-allele frequency in patients’ groups when compared to the controls (x2=7.3, P=0.007). Regarding OGG1 rs1052133 genotypes distribution, the GG genotype and G-allele were more prevalent, with significant lower CC genotype and C allele, in patients than in the controls (P<0.05). Additionally there were nonsignificant differences in genotypes or allelic distribution between HCC and chronic HCV patients. ORs (95%CI) in all patients versus the controls using the dominant and the recessive genetic models were shown (Table 3). There were nonsignificant differences as regards the genotypic or allelic frequencies of both SNPs in relation to the gender or liver function tests (ALT, AST, albumin and prothrombin concentration) or tumor spread.
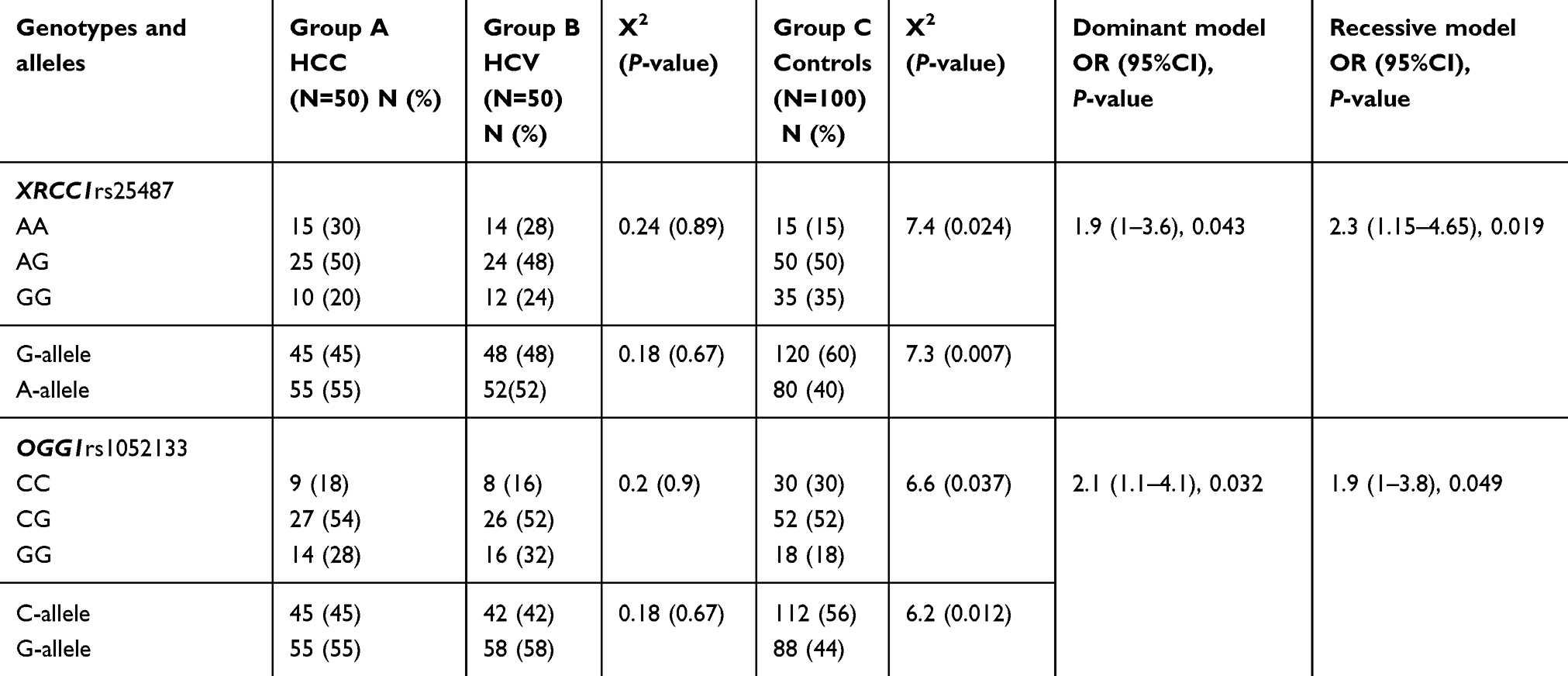 | Table 3 Genotypic and allelic frequencies of XRCC1rs25487 and OGG1rs1052133 among the studied groups |
Regarding patients with HCC who exhibited AA genotype for XRCC1 rs25487, they showed significant higher mean ±SD of urinary levels of 8-OHdG than those with AG or GG genotypes (229 mg/L ±28, 192 mg/L ±8 and 191 mg/L ±13 respectively) with similar findings among chronic HCV (149 mg/L ±30, 107 mg/L ±5, 102 mg/L ±7 respectively), P˂0.05 for all. Regarding mean ±SD of urinary levels of 8-OHdG in relation to OGG1 rs1052133 genotypes distribution, GG genotype revealed significant higher urinary excretion of 8-OHdG among HCC patients (239 mg/L ±12) and patients with chronic HCV (171 mg/L ±9) when compared with CG (204 mg/l ±18 and 138 mg/l ±26 respectively) and CC genotypes (201 mg/l ±27 and 134 mg/L ±31 respectively), P˃0.05 for all with nonsignificant differences among the controls (Table 4).
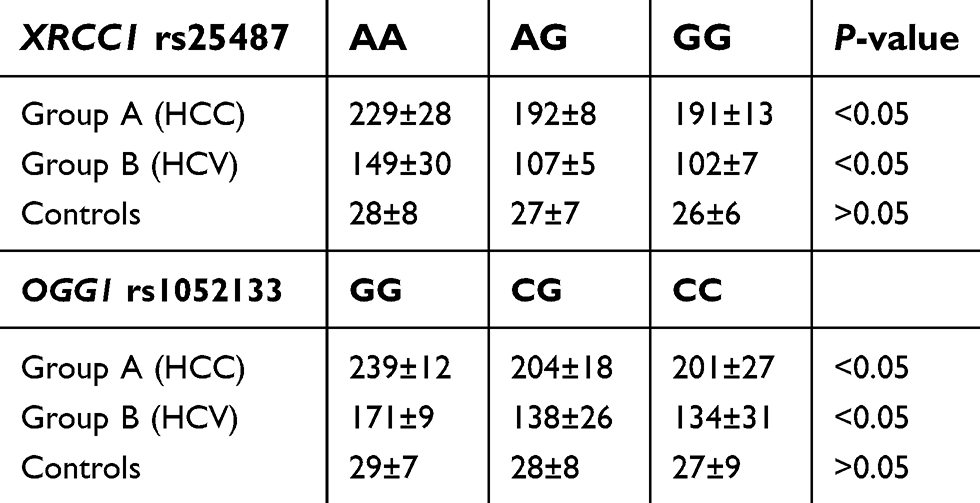 | Table 4 Mean ±SD of urinary 8-OHdG levels (mg/L) according to genotypes among the studied groups |
In addition, mean ±SD of urinary levels of 8-OHdG was significantly higher among HCC patients with both lobes affected (232.2±27) than one lobe (191.7±12), and in those with multiple malignant hepatic lesions (211±28) than those with single lesion (190±11), P<0.05 for both, (Table 5).
 | Table 5 Mean ±SD of urinary 8-OHdG levels (mg/L) in terms of tumor extension |
Discussion
Egypt is considered among the countries with highly prevalent HCV infection with 5.3 million subjects with HCV antibodies positive, of whom 3.7 million were positive for HCV-RNA.18 Furthermore, HCC account for 11.75% of digestive tract malignancies and 1.68% of cancer in general among Egyptians.19 HCC has an increasing incidence in Egypt due to the high prevalence of HBV and HCV as primary risk factors.20
8-hydroxy-deoxyguanosine is a key marker for measuring the effect of endogenous oxidative damage to DNA and plays a role in initiation and promotion of cancer.21 Although oxidative stress occurs frequently in chronic hepatitis C patients, but its role in further progression into HCC is not well known.22 The results of the current study showed an increase in urinary 8-OHdG level in patients with HCC and HCV patients than the controls with higher levels in HCC than in HCV group.
In the current study, AA genotype and A-allele of XRCC1 rs25487 G/A increased significantly in the patients with HCC when compared to the controls. The XRCC1 gene encodes one of the major repair factors involved in base excision repair, as it fixes base damage and DNA single strand breaks caused by alkylating agents and ionizing radiation and XRCC1 gene polymorphisms were found to be associated with the risk of several cancers. Many studies reported the association between XRCC1 rs25487 G/A gene polymorphism and different types of cancer. XRCC1 rs25487 G/A was reported to be associated with nonmelanoma skin cancer,23 childhood lymphoblastic leukemia in Asians,24 Hodgkin's disease,25 and HCC.26 The amino acid changes from Arg to Gln at codon 399 (exon 10) of XRCC1 protein may alter the protein function. It has been shown that these changes are associated with significant alterations in DNA repair capacity and increase the susceptibility to several diseases.27 In contrary, Mattar et al,28 reported XRCC1Arg194Trp but not XRCC1 Arg399Gln as associated risk factor for HCV related HCC development in Egypt that could be explained by the ethnic, geographic differences and the complex heterogeneity of HCC.
The findings of the current study also revealed risk association between OGG1 rs1052133 C/G (Ser326Cys) gene polymorphism and chronic HCV related HCC. G/G (Cys/Cys) genotype and G- allele increased significantly in HCC patients when compared to the controls. OGG1 rs1052133 C/G (Ser326Cys) gene polymorphism was found involved in base excision repair and was found associated with the risk of bladder,29 gallbladder,30 lung,31 and breast32 cancers. OGG1 Ser326Cys polymorphism may alter OGG1 mRNA levels leading to delayed repair of oxidative DNA damage. In silicocomputational biology tools reported that OGG1 Ser326Cys polymorphism may change the functional and structural characteristics of the OGG1 protein, so the 326Cys variant is smaller and more hydrophobic than the wild type. That change in the shape of OGG1 protein leads to deleterious effects on its function.33
In addition,our results revealed that AA genotype and A-allele of XRCC1 rs25487 G/A and G/G (Cys/Cys) genotype and G- allele of OGG1 rs1052133 C/G were increased significantly in the patients with chronic HCV which was in line with Almeida et al.34 This could be explained by the possible effects of HCV infection in preventing many DNA repair processes to enhance chromosomal instability in both hepatocytes and monocytes, which could explain the immunological perturbation and oncogenicity of HCV infection.35 Chronic inflammation induces reactive oxygen species production that cause damage of the cellular molecules including DNA, which if not repaired will resultin malignant transformation.
Moreover, patients having AA genotype of XRCC1 rs25487 G/A, and G/G (Cys/Cys) genotype of OGG1 rs1052133 C/G, exhibited significant higher urinary 8-OHdG levels.This could be explained by the impaired DNA repair ability of the proteins coded by these two genes as a result of the mutation with subsequent accumulation of the oxidation induced damaged DNA molecules (8-OHdG). OGG1 can removes 8-OHG by cleaving the N-glycosyl bonds between the oxidized guanine and the deoxyribose backbone, leaving an apurinic/apyrimidinic site and the presence of G allele cause downregulation of OGG1 with subsequent increase in the 8-OHdG levels.36,37 8-OHdG was found associated with increased risk for HCC and hepatic 8-OHdG levels are useful as markers to identify the high-risk individuals.22 Furthermore, urinary levels of 8-OHdG were significantly higher among HCC patients with both lobes affected rather than one lobe and in those with multiple malignant hepatic lesions than those with single lesion, indicating its association with the tumor spread and with a need for further evaluation.
Conclusion
XRCC1 rs25487 G/A, and OGG1 rs1052133 C/G polymorphisms are more prevalent in HCV and HCC patients than in controls, indicating impaired base excision repair induced HCV infection which may play a role in HCV related HCC development via impaired DNA repair protein activities as proved by the raised urinary 8-OHdG levels. Urinary 8-OHdG might be a noninvasive marker for the prediction of HCC in chronic HCV patients.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Manns MP, Buti M, Gane E, et al. Hepatitis C virus infection. Nat Rev Dis Primers. 2017;3:17006. doi:10.1038/nrdp.2017.6
2. Mohd Hanafiah K, Groeger J, Flaxman AD, Wiersma ST. Global epidemiology of hepatitis C virus infection: new estimates of age-specific antibody to HCV seroprevalence. Hepatology. 2013;57(4):1333–1342. doi:10.1002/hep.26141
3. Gower E, Estes C, Blach S, Razavi-Shearer K, Razavi H. Global epidemiology and genotype distribution of the hepatitis C virus infection. J Hepatol. 2014;61(1 Suppl):S45–S57. doi:10.1016/j.jhep.2014.07.027
4.
5. Chung YL. Defective DNA damage response and repair in liver cells expressing hepatitis B virus surface antigen. Faseb J. 2013;27:2316–2327. doi:10.1096/fj.12-226639
6. Takeshita H, Fujihara J, Yasuda T, Kimura-Kataoka K. Worldwide distribution of four SNPs in X-ray and repair and cross-complementing group 1 (XRCC1). Clin Transl Sci. 2015;8(4):347–350. doi:10.1111/cts.12237
7. Li Y, Liu F, Tan SQ, Wang Y, Li SW. X-ray repair cross-complementing group 1 (XRCC1) genetic polymorphisms and cervical cancer risk: a huge systematic review and meta-analysis. PLoS One. 2012;7(9):e44441. doi:10.1371/journal.pone.0044441
8. Huang Y, Li L, Yu L. XRCC1 Arg399Gln, Arg194Trp and Arg280His polymorphisms in breast cancer risk: a meta-analysis. Mutagenesis. 2009;24:331–339. doi:10.1093/mutage/gep013
9. Zou H, Li Q, Xia W, Liu Y, Wei X, Wang D. Association between the OGG1 Ser326Cys polymorphism and cancer risk: evidence from 152 case-control studies. J Cancer. 2016;7(10):1273–1280. doi:10.7150/jca.15035
10. Pan L, Zhu B, Hao W, et al. Oxidized guanine base lesions function in 8-oxoguanine DNA glycosylase-1-mediated epigenetic regulation of nuclear factor κB-driven gene expression. J Biol Chem. 2016;291(49):25553–25566. doi:10.1074/jbc.M116.751453
11. Kasymov RD, Grin IR, Endutkin AV, et al. Excision of 8-oxoguanine from methylated CpG dinucleotides by human 8-oxoguanine DNA. glycosylase. FEBS Lett. 2013;587(18):3129–3134. doi:10.1016/j.febslet.2013.08.008
12. Arauz J, Ramos-Tovar E, Muriel P. Redox state and methods to evaluate oxidative stress in liver damage: from bench to bedside. Ann Hepatol. 2016;15(2):160–173. doi:10.5604/16652681.1193701
13. Tsai JF, Chang WY, Jeng JE, Ho MS, Lin ZY, Tsai JH. Hepatitis B and C virus infection as risk factors for liver cirrhosis and cirrhotic hepatocellular carcinoma: a case-control study. Liver. 1994;14(2):98–102.
14. Bruix J, Sherman M, Llovet JM, et al. EASL panel of experts on HCC. Clinical management of hepatocellular carcinoma. Conclusions of the Barcelona-2000 EASL conference. European association for the study of the liver. J Hepatol. 2001;35(3):421–430.
15. Kawai K, Li YS, Kasai H. Accurate measurement of 8-OH-dG and 8-OH-Gua in mouse DNA, urine and serum: effects of X-ray irradiation. Genes Environ. 2007;29:107–114. doi:10.3123/jemsge.29.107
16. Saleem TH, Abo El-Maali N, Hassan MH, et al. Comparative protective effects of N-acetylcysteine, N-acetyl methionine, and N-acetyl glucosamine against paracetamol and phenacetin therapeutic doses-induced hepatotoxicity in rats. Int J Hepatol. 2018;2018:7603437. doi:10.1155/2018/7603437
17. Hamajima N, Saito T, Matsuo K, Kozaki K, Takahashi T, Tajima K. Polymerase chain reaction with confronting two-pair primers for polymorphism genotyping. Jpn J Cancer Res. 2000;91(9):865–868.
18. Kandeel A, Genedy M, El-Refai S, Funk AL, Fontanet A, Talaat M. The prevalence of hepatitis C virus infection in Egypt 2015: implications for future policy on prevention and treatment. Liver Int. 2017;37(1):45–53. doi:10.1111/liv.13450
19. Mokhtar N, Gouda I, Adel I. Cancer pathology registry 2003-2004 and time trend analysis. In: Mokhtar N, Gouda I, Adel I, editors. Malignant Digestive System Tumors. Cairo: Elsheraa Press; 2007:55–67.
20. Gomaa AI, Hashim MS, Waked I. Comparing staging systems for predicting prognosis and survival in patients with hepatocellular carcinoma in Egypt. PLoS One. 2014;9:e90929. doi:10.1371/journal.pone.0090929
21. Valavanidis A, Vlachogianni T, Fiotakis C. 8-hydroxy-2’ -deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev. 2009;27(2):120–139. doi:10.1080/10590500902885684
22. Tanaka H, Fujita N, Sugimoto R, et al. Hepatic oxidative DNA damage is associated with increased risk for hepatocellular carcinoma in chronic hepatitis C. Br J Cancer. 2008;98(3):580–586. doi:10.1038/sj.bjc.6604204
23. Nelson HH, Kelsey KT, Mott LA, Karagas MR. The XRCC1 Arg399Gln polymorphism, sunburn, and non-melanoma skin cancer: evidence of gene-environment interaction. Cancer Res. 2002;62(1):152–155.
24. Wang F, Zhao Q, He HR, et al. The association between XRCC1 Arg399Gln polymorphism and risk of leukemia in different populations: a meta-analysis of case-control studies. Onco Targets Ther. 2015;8:3277–3287. doi:10.2147/OTT.S92752
25. El-Zein R, Monroy CM, Etzel CJ, et al. Genetic polymorphisms in DNA repair genes as modulators of Hodgkin disease risk. Cancer. 2009;115(8):1651–1659. doi:10.1002/cncr.24205
26. Jung SW, Park NH, Shin JW, et al. Polymorphisms of DNA repair genes in Korean hepatocellular carcinoma patients with chronic hepatitis B: possible implications on survival. J Hepatol. 2012;57(3):621–627. doi:10.1016/j.jhep.2012.04.039
27. Duman N, Aktan M, Ozturk S, et al. Investigation of Arg399Gln and Arg194Trp polymorphisms of the XRCC1 (x-ray cross-complementing group 1) gene and its correlation to sister chromatid exchange frequency in patients with chronic lymphocytic leukemia. Genet Test Mol Biomarkers. 2012;16(4):287–291. doi:10.1089/gtmb.2011.0152
28. Mattar MM, Zekri AN, Hussein N, Morsy H, Esmat G, Amin MA. Polymorphisms of base-excision repair genes and the hepatocarcinogenesis. Gene. 2018;675:62–68. doi:10.1016/j.gene.2018.06.056
29. Arizono K, Osada Y, Kuroda Y, Repair DNA. Gene hOGG1 codon 326 and XRCC1 codon 399 Polymorphisms And Bladder Cancer Risk In a Japanese population. Japanese J ClinOncol. 2008;38:186–191. doi:10.1093/jjco/hym176
30. Jiao X, Huang J, Wu S, et al. hOGG1 Ser326Cys polymorphism and susceptibility to gallbladder cancer in a Chinese population. Int J Cancer. 2007;121(3):501–505. doi:10.1002/ijc.22748
31. Le Marchand L, Donlon T, Lum-Jones A, Seifried A, Wilkens LR. Association of the hOGG1 Ser326Cys polymorphism with lung cancer risk. Cancer Epidemiol Biomarkers Prev. 2002;11(4):409–412.
32. Alanazi M, Pathan AAK, Shaik JP, et al. The hOGG1 Ser326Cys gene polymorphism and breast cancer risk in Saudi population. Pathol Oncol Res. 2017;23(3):525–535. doi:10.1007/s12253-016-0146-6
33. Kang SW, Kim SK, Park HJ, Chung JH, Ban JY. Human 8-oxoguanine DNA glycosylase gene polymorphism (Ser326Cys) and cancer risk: updated meta-analysis. Oncotarget. 2017;8(27):44761–44775. doi:10.18632/oncotarget.16226
34. Almei Da Pereira Leite ST, Marques-Guimarães N, Silva-Oliveira JC, Dutra-Souto FJ, Alves-dos-Santos R, Bassi-Branco CL. The X-ray repair cross complementing protein 1 (XRCC1) rs25487 polymorphism and susceptibility to cirrhosis in Brazilian patients with chronic viral hepatitis. Ann Hepatol. 2013;12(5):733–739.
35. Machida K, McNamara G, Cheng KT, et al. Hepatitis C virus inhibits DNA damage repair through reactive oxygen and nitrogen species and by interfering with the ATM-NBS1/Mre11/Rad50 DNA repair pathway in monocytes and hepatocytes. J Immunol. 2010;185(11):6985–6998. doi:10.4049/jimmunol.1000618
36. Cooke MS, Evans MD. 8-Oxo-deoxyguanosine: reduce, reuse, recycle. Proc Natl Acad Sci U S A. 2007;104:13535–13536. doi:10.1073/pnas.0706878104
37. Peng T, Shen HM, Liu ZM, et al. Oxidative DNA damage in peripheral leukocytes and its association with expression and polymorphisms of hOGG1: A study of adolescents in a high risk region for hepatocellular carcinoma in China. World J Gastroenterol. 2003;9:2186–2193. doi:10.3748/wjg.v9.i10.2186
Supplementary materials
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.