")
Back to Journals » Infection and Drug Resistance » Volume 14
Unraveling Mechanisms and Epidemic Characteristics of Nitrofurantoin Resistance in Uropathogenic Enterococcus faecium Clinical Isolates
Authors Zhang Y, Wang L, Zhou C, Lin Y, Liu S, Zeng W, Yu K, Zhou T , Cao J
Received 13 January 2021
Accepted for publication 8 April 2021
Published 22 April 2021 Volume 2021:14 Pages 1601—1611
DOI https://doi.org/10.2147/IDR.S301802
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Ying Zhang,1 Lingbo Wang,2 Cui Zhou,2 Yishuai Lin,1 Shixing Liu,2 Weiliang Zeng,1 Kaihang Yu,1 Tieli Zhou,2 Jianming Cao1
1Department of Medical Lab Science, School of Laboratory Medicine and Life Science, Wenzhou Medical University, Wenzhou, Zhejiang Province, 325035, People’s Republic of China; 2Department of Clinical Laboratory, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang Province, 325035, People’s Republic of China
Correspondence: Jianming Cao
Department of Medical Lab Science, School of Laboratory Medicine and Life Science, Wenzhou Medical University, Wenzhou, Zhejiang Province, People’s Republic of China
Tel +86-0577-88069595
Email [email protected]
Tieli Zhou
Department of Clinical Laboratory, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang Province, People’s Republic of China
Tel +86-0577-86689885
Email [email protected]
Purpose: Multidrug-resistant (MDR) Enterococcus faecium is an important nosocomial pathogen causing urinary tract infection, and the reapplication of nitrofurantoin (NIT) in the clinic has attracted great attention. This study aims to explore the NIT resistance mechanisms and epidemiological characteristics of E. faecium clinical isolates.
Patients and Methods: A total of 633 E. faecium clinical isolates was obtained from urine samples in a clinical teaching hospital during 2017– 2018. Among them, 40 NIT-resistant strains, and a similar number of -intermediate and -susceptible strains were isolated. The minimum inhibitory concentrations (MICs) of NIT were detected by agar dilution method. The prevalence and mutations of nitroreductase-encoding genes ef0404 and ef0648 were explored by polymerase chain reaction (PCR), followed by efflux pump inhibition test and quantitative real-time PCR (qRT-PCR) to investigate the resistance mechanisms of NIT. Furthermore, the epidemiological characteristics were detected by multilocus sequence typing (MLST).
Results: The carrying rates of nitroreductase in NIT-susceptible, -intermediate, and -resistant isolates were 100%, 50%, and 20%, respectively. After exposure to the efflux pump inhibitor, the MIC of 12 E. faecium decreased by ≥ 4-fold. However, the efflux pump genes efrAB, emeA, and oqxAB were not overexpressed in NIT-resistant E. faecium isolates. Moreover, MLST analysis revealed that all the NIT-resistant isolates belonged to CC17, of which 30 (75%) were associated with ST78.
Conclusion: This study has established for the first time that the absence of EF0404 and EF0648 is the main mechanism of NIT resistance in E. faecium. Our findings are likely to fill the knowledge gap pertaining to the NIT resistance mechanism in E. faecium and provide important insights for molecular epidemiological characteristics analysis.
Keywords: Enterococcus faecium, nitrofurantoin, nitroreductase, resistance mechanism, epidemiology
Introduction
As a ubiquitous group of Gram-positive bacteria, Enterococcus is a leading cause of hospital-acquired infections and has therefore posed a serious threat to public health around the world.1,2 The important infections most commonly caused by Enterococcus are urinary tract, device-associated, and soft-tissue infections as well as bacteremia.3 In recent years, Enterococcus faecium has become the foremost Gram-positive pathogen responsible for urinary tract infections.4,5 In addition to intrinsic resistance and genetic diversity, its ability to recruit and express antimicrobial resistance determinants contributes to the rapid increase of multidrug resistance (MDR) in E. faecium.6,7 The limited availability of novel antimicrobial agents has posed serious challenges to the clinical treatment of infectious diseases.8,9
As an age-old synthetic drug, nitrofurantoin (NIT) has been used for the prevention of urinary tract infection (UTI) for over 60 years.10 Because of its considerable bactericidal activity and low resistance rate, the drug has attracted renewed clinical interest. The efficacy of NIT against high level of aminoglycoside-resistant (HLAR) Enterococcus and vancomycin-resistant Enterococcus (VRE) has led to it being considered as the last resort for the first-line therapy of uncomplicated lower urinary tract infection caused by several bacteria.11–13
However, the extensive use of NIT has resulted in the increased drug resistance of E. faecium. Resistance mechanisms of NIT previously studied are limited to Enterobacteriaceae. For instance, mutations in the nitroreductase-encoding gene nfsAB and the overexpression of the efflux pump gene oqxAB play important roles in NIT resistance in Escherichia coli and Klebsiella pneumoniae.14–17 In addition, deletion in ribE (encoding lumazine synthase involved in the biosynthesis of flavin mononucleotide) is a key NIT resistance mechanism in E. coli.13,18 However, the mechanism of NIT resistance in E. faecium remains poorly understood.
Previous studies have reported that EF0404, EF0648, EF0655, and EF1181 exhibit nitroreductase activity in Enterococcus faecalis V583.19 Besides, Fatemeh Raci (2003) has suggested that the antimicrobial effect of nitro drugs is mediated by microbial nitroreductases that reduce the drug to a cytotoxic nitro radical and result in DNA damage.20 Nonetheless, there is no evidence to establish that nitroreductase is responsible for NIT resistance in E. faecium.
Hence, we aimed to investigate the main mechanism of NIT resistance in E. faecium, which is mediated by nitroreductases and multidrug resistance efflux pumps. Besides, multilocus sequence typing (MLST) was performed to reveal the homology among the NIT-resistant isolates.
Materials and Methods
Bacterial Strains
A total of 633 non-duplicated E. faecium strains was isolated from the First Affiliated Hospital of Wenzhou Medical University (Wenzhou, China) in 2017–2018. Bacterial identification was performed using Matrix-Assisted Laser Desorption/Ionization Time of Flight Mass Spectrometry (MALDI-TOF MS; BioMérieux, Lyons, France). E. faecium ATCC 29212 was used as a quality control strain in antimicrobial susceptibility testing experiments.
Antimicrobial Susceptibility Testing
Agar dilution method was applied to determine the minimum inhibitory concentration (MIC) of NIT, and the results were interpreted according to the recommendations of the Clinical and Laboratory Standards Institute (CLSI, 2020) guidelines. The breakpoints of NIT for the agar dilution method were as follows: susceptible ≤32 mg/L; intermediate = 64 mg/L; and resistant ≥128 mg/L. All strains were tested in three biologically independent experiments.
PCR Amplification and DNA Sequencing
Genomic DNA of the experimental strains was extracted using the Bioflux Bacterial DNA Extraction Kit (Bioflux, Tokyo, Japan) as per the instructions of the manufacturer. Polymerase chain reaction (PCR) was employed to detect NIT resistance-related genes (ef0404, ef0648, ef0655, and ef1181) and efflux pump genes (oqxA, oqxB, efrA, efrB, and emeA). The positive PCR products were sent to Shanghai Majorbio Bio-Pharm Technology Co. (Shanghai, China) for sequencing. The sequences were compared with the standard strain E. faecium DO (Accession number: CP003583) deposited in the National Center for Biotechnology Information (NCBI) database using BLASTn and BLASTx programs (http://blast.ncbi.nlm.nih.gov/Blast.cgi). The online PROVEAN platform (http://provean.jcvi.org/seq_submit.php) was used to predict the alterations in the biological functions of the proteins. Primers used for amplification and sequencing are furnished in Table 1.
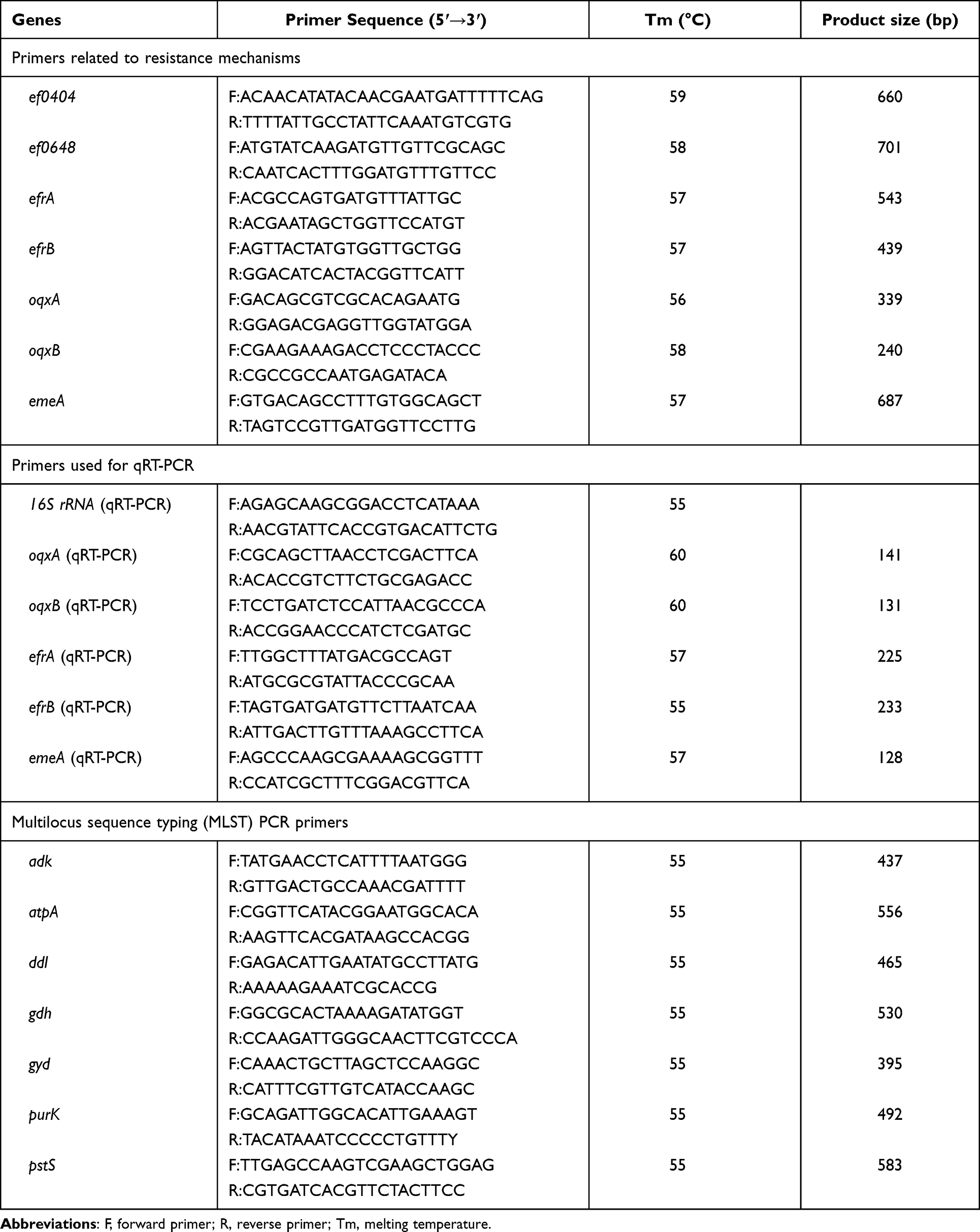 |
Table 1 Primers Used in This Study |
Effect of Efflux Pump Inhibitor
Several studies have revealed the presence of 34 potential drug-efflux genes in the E. faecalis genome, and the pumps have been shown to exhibit differing but somewhat overlapping broad substrate profiles.21 Therefore, the efflux pump inhibition test was performed to identify which efflux pump is responsible for NIT resistance in E. faecium. The MICs of E. faecium with or without the efflux pump inhibitors carbonyl cyanide m-chlorophenylhydrazone (CCCP, 6 μg/mL), verapamil (100 μg/mL), chlorpromazine (20 μg/mL), reserpine (20 μg/mL), omeprazole (100 μg/mL), and Phe-Arg-β-naphthylamide (PAβN) (20 μg/mL) were compared to measure the efflux activities. The phenotype test is regarded to be positive when the MIC of the strain decreases by ≥4-fold after the supplementation of the efflux pump inhibitor.16 NIT-susceptible and -intermediate E. faecium strains were used as negative controls.
Quantitative Real-Time PCR (qRT-PCR) of Efflux Pump Gene
To evaluate the relative expression levels of efflux pump genes efrAB, emeA, and oqxA, qRT-PCR was performed on NIT-resistant and -susceptible E. faecium isolates before and after induction with NIT of 1/2 MIC based on a 7500 RT-PCR system (Thermo Fisher Scientific, Marsiling, Singapore) with an SYBRTM Green RT-PCR Kit (TOYOBO, Osaka, Japan). The primers used are listed in Table 1. To extract the RNA of the experimental strains, a single colony was selected and inoculated overnight in blood agar plates and added to fresh Luria broth (LB) medium with shaking at 180 rpm to logarithmic phase (OD600 value of approximately 0.5) at 37°C. The bacterial culture (3 mL) was centrifuged at 14,000 × g for 5 min, and the supernatant was discarded. Total RNA was extracted using the RNeasy Mini Kit (Qiagen, Valencia, CA, USA) according to the manufacturer’s protocol. The concentration and purity of the extracted RNA were estimated, and the samples were stored at −80°C for further experiments. The purified RNA was then reverse transcribed to cDNA for qRT-PCR analysis with a cDNA Synthesis Kit (Takara, Japan) based on the manufacturer’s manual. The 16S rRNA gene was used as the reference gene to normalize the data. The 2–ΔΔCt method was utilized for determining the expressions of efrAB, emeA, and oqxA.
MLST Typing of NIT-Resistant Isolates
In this experiment, clone correlation analysis of the 40 NIT-resistant isolates was carried out with MLST by amplifying seven housekeeping loci (adk, atpA, ddl, gdh, ggd, pstS, and purK) of E. faecium. The database available at Institut Pasteur’s E. faecium MLST website (http://efaecium.mlst.net/) was checked to obtain the corresponding allelic profiles and subsequently their sequence type (ST). The primers used are given in Table 1.
Statistical Analysis
All data were analyzed using the GraphPad Prism v8.01 statistical software package (GraphPad Software, La Jolla, CA, USA). The Chi-Square test was used to compare the significance of gene carriage rate. Unpaired Student’s t-test (two-tailed) was performed for comparing the significance of gene expression in qRT-PCR. p-values of <0.05 were considered to be statistically significant. For all analyses, * = p < 0.05, ** = p < 0.01, and *** = p < 0.001.
Results
Antimicrobial Susceptibility Testing
Among the 633 E. faecium isolates collected from the First Affiliated Hospital of Wenzhou Medical University (Wenzhou, China) in 2017–2018, 50.9% (322/633) were resistant to NIT. As shown in Table 2, the NIT-resistant strains had higher drug resistance rates than the NIT-intermediate and -susceptible strains. Based on the results of antimicrobial susceptibility testing, 40 each of NIT-resistant, -intermediate, and -susceptible E. faecium strains isolated from the urine samples were selected for further research. The NIT-resistant isolates demonstrated high resistance rates toward ampicillin, penicillin, fluoroquinolones (ciprofloxacin and levofloxacin), and tetracycline. Besides, all strains were highly susceptible to linezolid and glycopeptides (vancomycin and teicoplanin) (Table 3).
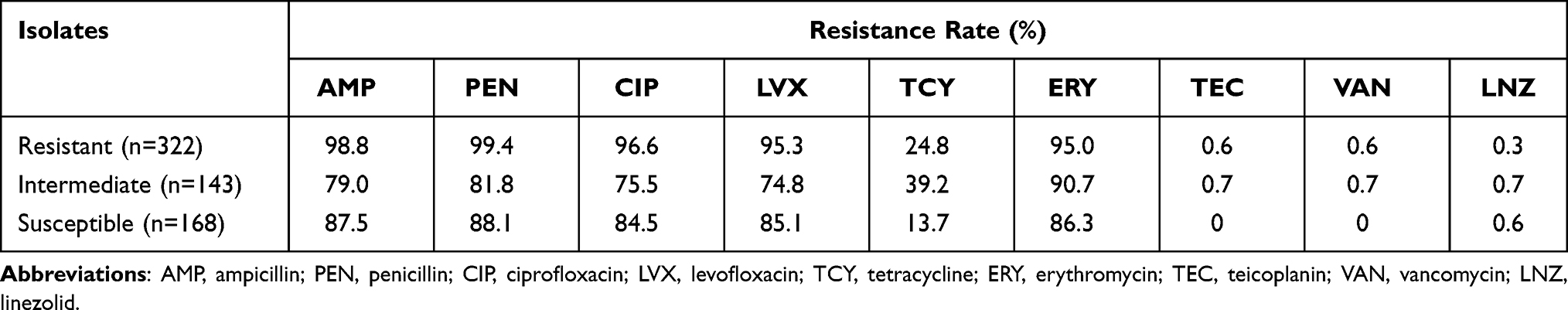 |
Table 2 Percentage of Antimicrobial Agent Resistance Rates Among Nitrofurantoin-Resistant, -Intermediate and -Susceptible E. Faecium Isolates |
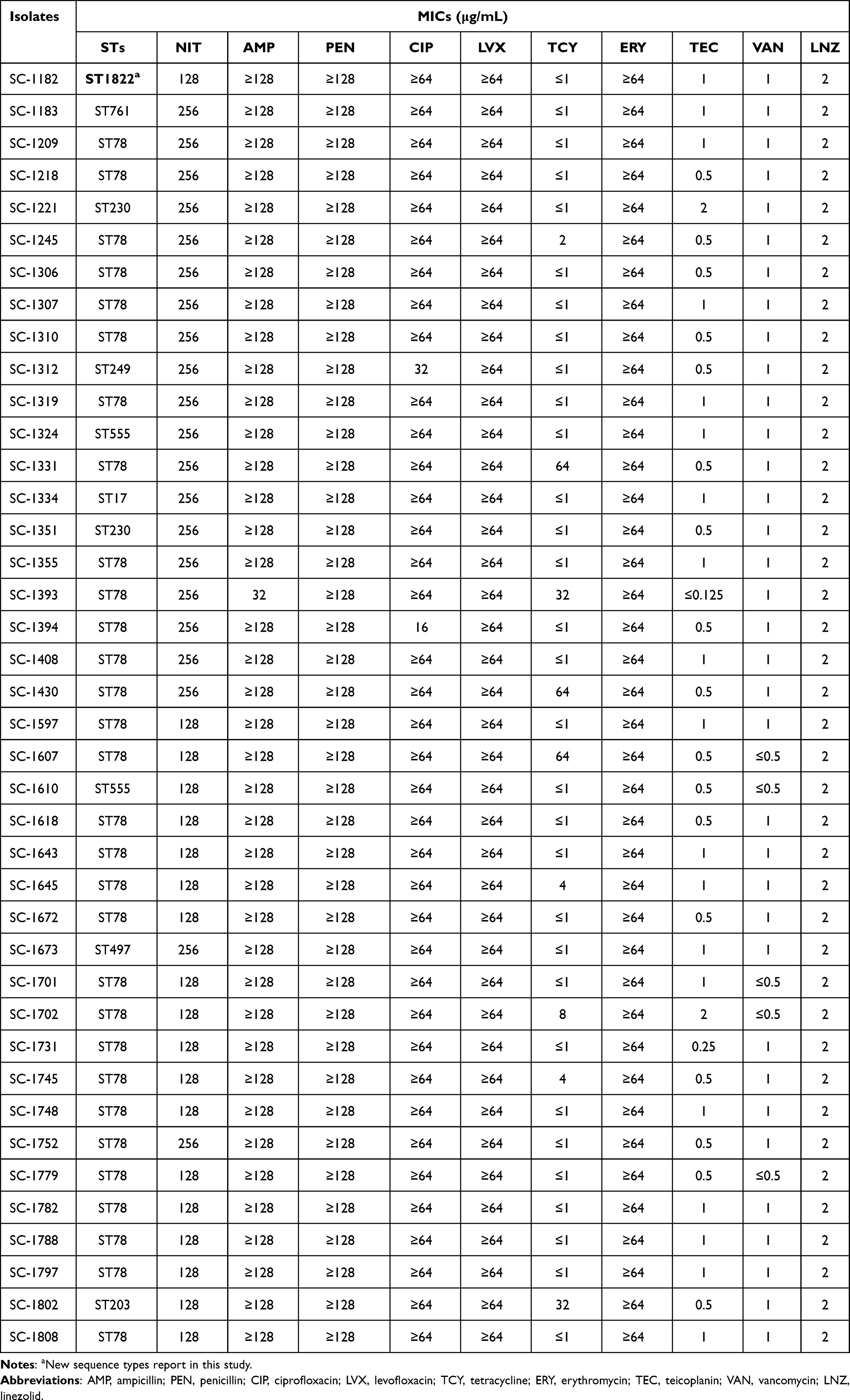 |
Table 3 Minimum Inhibitory Concentrations (MICs) and Sequence Types (STs) of 40 Nitrofurantoin-Resistant E. Faecium Isolates |
Molecular Mechanisms of NIT Resistance
A significant positive correlation was observed between NIT resistance and the prevalence of ef0404 and ef0648 genes. In our study, all the susceptible isolates were found to carry at least one nitroreductase gene (ef0404 75.0% and ef0648 72.5%), and the carriage rates of the nitroreductase genes among the NIT-intermediate (ef0404 20.0% and ef0648 35.0%) and -resistant isolates (ef0404 5.0% and ef0648 17.5%) were 50% and 20%, respectively (Table 4). Furthermore, our study revealed that 20.8% (25/120) of the E. faecium isolates carried efrAB and 10.0% (12/120) carried emeA. One unanticipated finding was that the oqxA gene was present in only two (SC-1245 and SC-1325) out of 120 E. faecium isolates, and none had the oqxB gene (Figure S1).
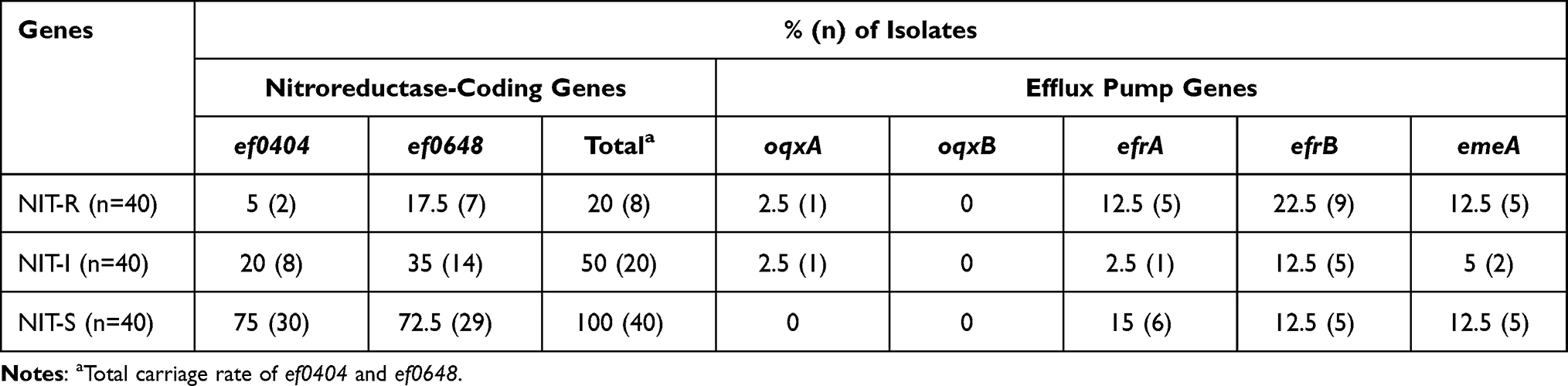 |
Table 4 Distribution of Resistance-Related Genes in Nitrofurantoin Resistant (NIT-R), -Intermediate (NIT-I) and -Susceptible (NIT-S) Enterococcus Faecium Isolates |
While further exploring the resistance mechanisms of NIT, mutations of the resistance determinants ef0404 and ef0648 were detected in 12 NIT-resistant E. faecium isolates. Most of these were nonsense mutations, while missense mutation 478G > A and 52C > T of the nitroreductase-encoding gene ef0648 was observed in four E. faecium strains. Moreover, base insertion (69–70insC) and amino acid mutations (52C > T) in ef0648 were considered deleterious by PROVEAN. No missense mutation was found in the nitroreductase-encoding gene ef0404 (Table 5).
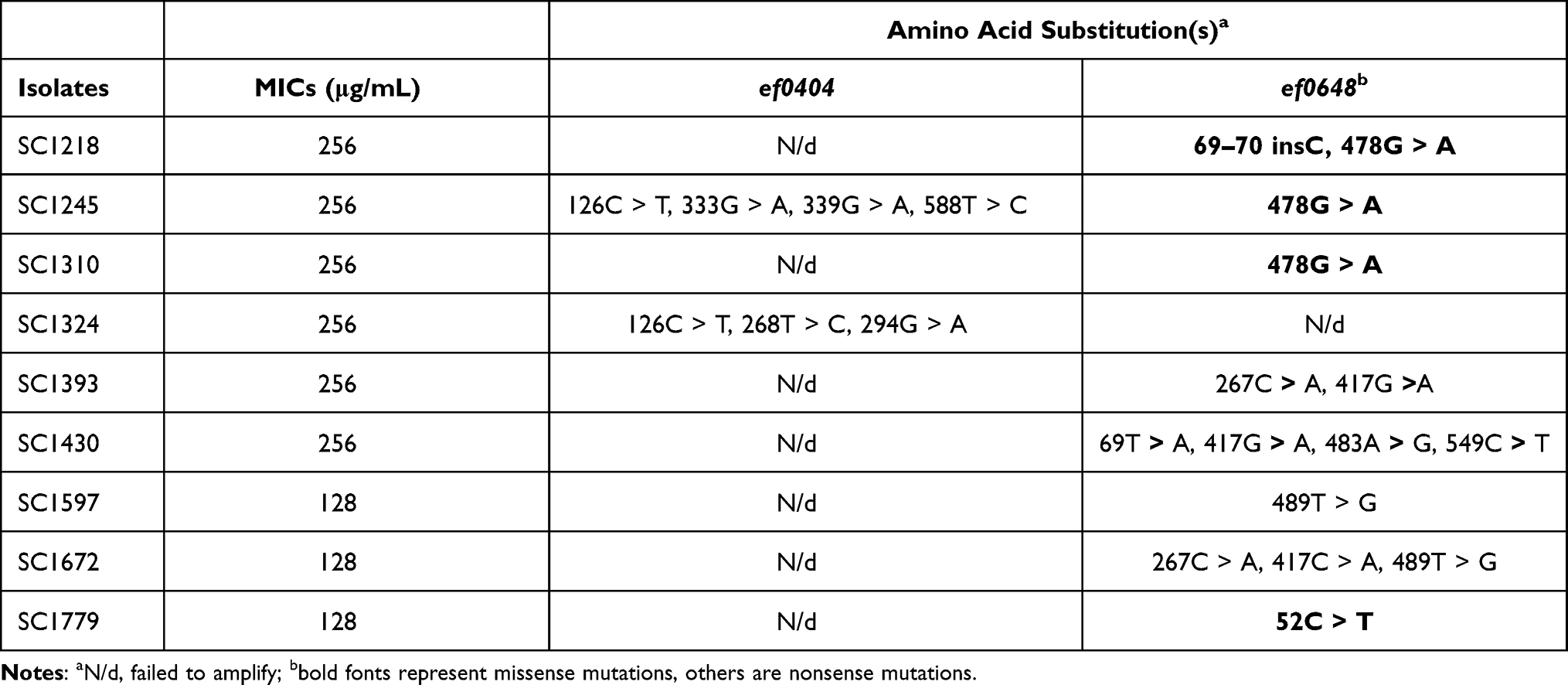 |
Table 5 Analysis the Mutations of Nitroreductase in Nitrofurantoin-Resistant Isolates |
The Potential Effect of the Efflux Pump Mechanism
The correlation between NIT resistance and efflux pump was tested. The results suggested that after exposure to the efflux inhibitors CCCP, verapamil, and chlorpromazine, the MICs of the 12 NIT-resistant E. faecium strains decreased by ≥4-fold. However, when used in combination with reserpine, omeprazole, and PAβN, the efflux pump inhibition test was negative. The MIC showed either no change or <4-fold decrease in NIT-susceptible (SC-1177), and NIT-intermediate (SC-1178) E. faecium strains (Table 6). Our findings allude that the overexpression of the efflux pump may influence NIT resistance in E. faecium.
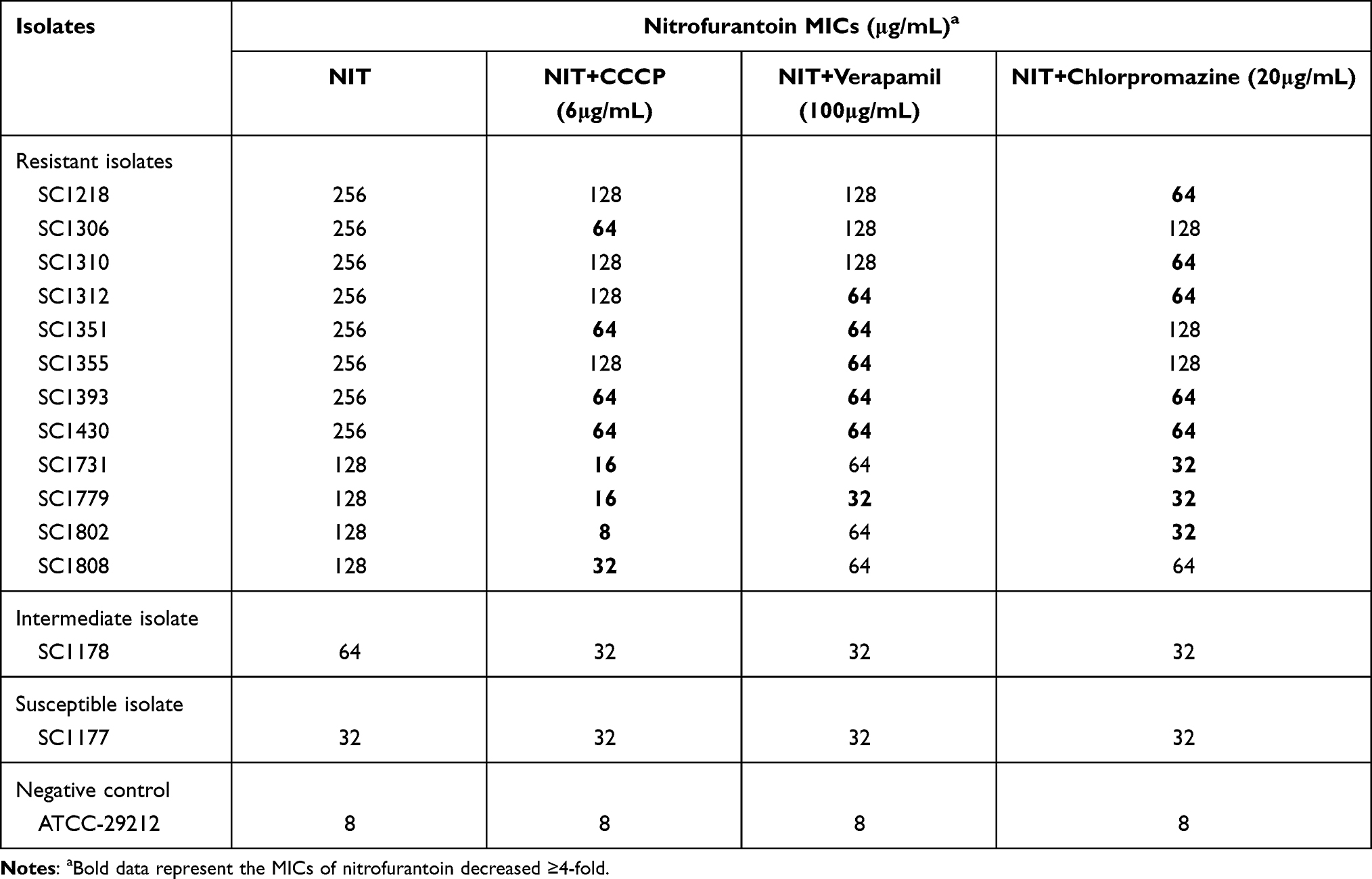 |
Table 6 Minimum Inhibitory Concentrations (MICs) of Nitrofurantoin (NIT) in Nitrofurantoin-Resistant (n = 12) -Intermediate (n = 1), and -Susceptible (n = 1) Isolates with or without the Efflux Pump Inhibitors |
Analysis of the Expression Level of the Efflux Pump Gene
Previous studies have indicated that efflux pumps EfrAB and EmeA are present in E. faecium and lead to multidrug resistance.22,23 PCR results exposed that efflux pumps EfrAB and EmeA existed in 12 and 5 NIT-resistant E. faecium strains, respectively. The OqxAB efflux pump was present in only one NIT-resistant and one NIT-intermediate E. faecium strain. We examined the effect of efflux pump overexpression on NIT resistance. As depicted in Figure 1, the expression levels of efrAB, emeA, and oqxA in four NIT-resistant strains after induction by NIT of 1/2 MIC were not significantly increased in comparison with the control strain ATCC 29212. Hence, our results indicate that the efflux pumps EfrAB, EmeA, and OqxAB do not play a significant role in NIT resistance in the isolated E. faecium strains.
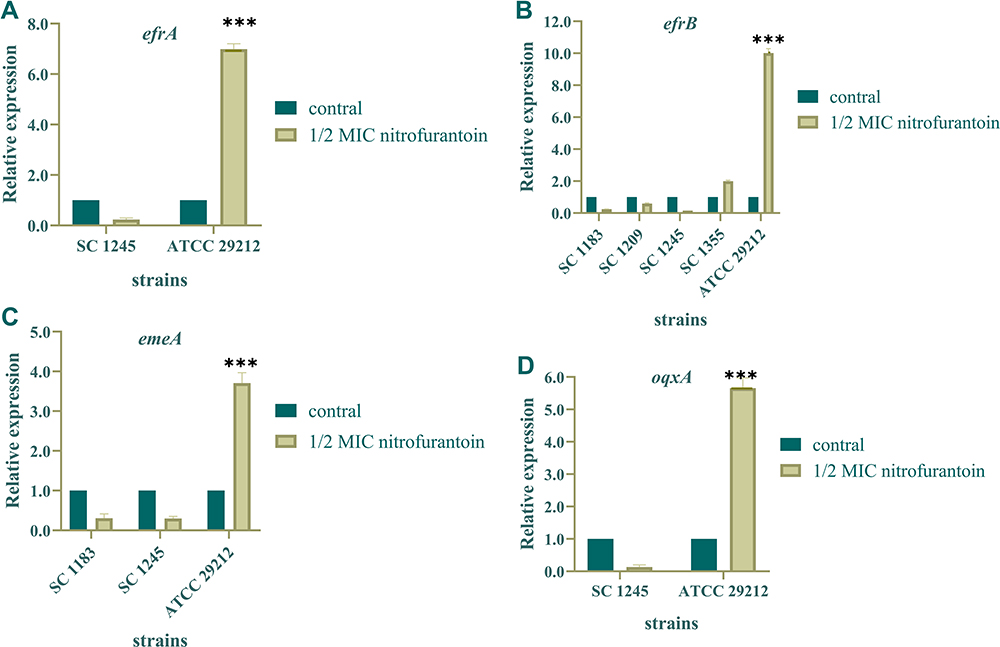 |
Figure 1 Relative expression levels of the efflux pump encoding genes in 4 nitrofurantoin-resistant E. faecium isolates. (A–D) The relative expression levels of efflux pump genes efrA, efrB, emeA, and oqxA in 4 nitrofurantoin-resistant E. faecium isolates before and after induced by nitrofurantoin of 1/2 MIC. P < 0.05 were considered to be statistically significant. ***P < 0.001 (Student’s t-test). |
Molecular Epidemiological Analysis
Amplification of the seven housekeeping genes of the 40 NIT-resistant E. faecium isolates by PCR and MLST analysis revealed that they belonged to the same subtype, CC17. ST78 (30/40) was the predominant ST, accounting for 75.0%, followed by ST230 (2/40) and ST555 (2/40). Moreover, our investigation revealed a new sequence type (ST1822) for the first time (yet to be registered in the MLST database), and four individual isolates were assigned to ST761, ST17, ST249, ST497, and ST203 (Table 3). These results are consistent with Djahmi’s findings, which showed that the most important STs present in E. faecium belong to the clonal complex CC17 and that the major ST of E. faecium is ST78 in Europe and Asia.24 These results prove that the clonal cluster CC17 associated with nitrofurantoin-resistant E. faecium might originate from a single clonal lineage, which is likely to provide important insights for molecular epidemiological analysis.
Discussion
Prior studies have documented that E. faecium is an important pathogen causing urinary tract infection.4,5 Its Intrinsic resistance and the capability to acquire resistance genes pose restrictions on therapeutic options.25 NIT is an effective antimicrobial agent used for the treatment of urinary tract infections, and its application has increased exponentially in recent years.26 More importantly, NIT can also be used in extended spectrum beta-lactamase-producing and carbapenem-resistant bacterial infections.27,28 The European Association of Urology guidelines recommend the use of NIT as a first-line treatment for acute uncomplicated cystitis, especially in women.29 In this study, majority of the NIT-resistant strains were isolated from urine samples, which might be explained by the fact that NIT is mainly used for the treatment of urinary tract infections.30 The drug possesses several antibacterial mechanisms, the most important of which is the reduction of its nitro group by the bacterial nitroreductases, producing toxic products and thus affecting the cell’s metabolism.31 However, the resistance mechanisms of NIT in E. faecium are yet to be well understood, and further investigations are needed to prevent the spread of NIT resistance.
Significantly, we proved for the first time that nitroreductases EF0404 and EF0648 were responsible for NIT resistance in E. faecium and that deletion in the nitroreductase-encoding gene is the main mechanism involved. As expected, while the nitroreductase-encoding gene was found to exist in 100% of the NIT-sensitive E. faecium strains, it occurred in only 20% of the NIT-resistant strains. However, previous studies have asserted that four nitroreductases (EF0404, EF1181, EF0648, and EF0655) are present in E. faecalis.19 However, only EF0404 and EF0648 were detected in our study. This may be the reason for the huge difference in the NIT resistance rates between E. faecalis and E. faecium (0.6% vs 50.9%). Besides, to the best of our knowledge, this is the first time that the plasmid-borne NIT resistance gene oqxAB has been detected in clinical E. faecium isolates, which mediates resistance to NIT in E. coli and K. pneumonia.15–17 However, our result disagrees with the findings of Li Yuan et al,32 who reported that the carriage rates of oqxA and oqxB (79.3% and 65.5%, respectively) in Enterococcus were significantly higher than that in this study (1.7% and 0, respectively). We speculate that this variation might have been caused by the difference in sample sources since the strains were isolated from swine manure in the earlier study. Although the prevalence of the oqxAB gene was low in our study, screening for the gene should be implemented to prevent its spread among E. faecium.33
Based on the results of antimicrobial susceptibility testing, many E. faecium isolates were found to be resistant to most of the antimicrobial agents in clinical use, probably due to genes encoding MDR efflux pumps in Enterococcus.22 To ascertain the role of efflux pumps in the NIT-resistant phenotype of the 40 E. faecium isolates, the most extensively studied efflux pumps EmeA (a member of the major facilitator superfamily) and EfrAB (belonging to the ATP-binding cassette (ABC) superfamily) were amplified by PCR.24,34 The results revealed that 12 and 5 NIT-resistant E. faecium strains exhibited the efflux pumps EfrAB and EmeA, respectively. Moreover, the effect of efflux inhibitors (including CCCP, verapamil, chlorpromazine, reserpine, omeprazole, and PAβN) on the activity of NIT against E. faecium was investigated. It was discerned that 12 of the 40 (40%) NIT-resistant isolates demonstrated a 4-fold decrease in the MIC of NIT in the presence of the efflux pump inhibitors CCCP, verapamil, and chlorpromazine. Moreover, according to qRT-PCR, efflux pump genes efrAB, emeA, and oqxA in the NIT-resistant strains were not overexpressed after induction by NIT. Our results imply that the overexpression of the pump leads to NIT resistance in the E. faecium isolates. However, which efflux pump plays a role warrants further research. Furthermore, the results of MLST alluded that the distribution of the STs was concentrated, suggesting that the NIT-resistant strains may spread horizontally among the nosocomial population. Although we have illuminated the main mechanisms of NIT resistance in E. faecium, more research needs to be undertaken to completely elucidate the concept.
Conclusions
Collectively, the findings of the present investigation provide convincing evidence that deletions in the nitroreductases-encoding genes ef0404 and ef0648 and the overexpression of the efflux pump genes are the main reasons for NIT resistance in E. faecium. Our results would be helpful in establishing a theoretical basis for the rational use of the drug and in adopting appropriate control measures to curb the increase in antimicrobial resistance.
Ethical Statement
The whole investigation protocols in this study were approved by The Ethics Committee of the First Affiliated Hospital of Wenzhou Medical University. There are no studies with humans or animals performed by any of the authors in this article. Informed consent was waived because this study with observational nature mainly focused on bacteria and did no interventions to patients.
Acknowledgments
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Arias CA, Murray BE. The rise of the Enterococcus: beyond vancomycin resistance. Nat Rev Microbiol. 2012;10(4):266–278. doi:10.1038/nrmicro2761
2. García-Solache M, Rice LB. The Enterococcus: a Model of Adaptability to Its Environment. Clin Microbiol Rev. 2019;32(2):e00058–18.
3. Raza T, Ullah SR, Mehmood K, Andleeb S. Vancomycin resistant Enterococcus: a brief review. J Pak Med Assoc. 2018;68(5):768–772.
4. Shrestha LB, Baral R, Poudel P, Khanal B. Clinical, etiological and antimicrobial susceptibility profile of pediatric urinary tract infections in a tertiary care hospital of Nepal. BMC Pediat. 2019;19(1):36. doi:10.1186/s12887-019-1410-1
5. O’Brien VP, Hannan TJ, Nielsen HV, Hultgren SJ. Drug and Vaccine Development for the Treatment and Prevention of Urinary Tract Infections. Microbiol Spectr. 2016;4(1):
6. Cattoir V, Leclercq R. Twenty-five years of shared life with vancomycin-resistant Enterococcus: is it time to divorce? J Antimicrob Chemother. 2013;68(4):731–742. doi:10.1093/jac/dks469
7. Miller WR, Munita JM, Arias CA. Mechanisms of antibiotic resistance in. Enterococcus Expert Rev Anti Infect Ther. 2014;12(10):1221–1236. doi:10.1586/14787210.2014.956092
8. Kilbas I, Ciftci IH. Antimicrobial resistance of Enterococcus isolates in Turkey: a meta-analysis of current studies. J Glob Antimicrob Resist. 2018;12:26–30. doi:10.1016/j.jgar.2017.08.012
9. Yim J, Smith JR, Rybak MJ. Role of Combination Antimicrobial Therapy for Vancomycin-Resistant Enterococcus faecium Infections. Review of the Current Evidence. Pharmacotherapy. 2017;37(5):579–592. doi:10.1002/phar.1922
10. Wijma RA, Huttner A, Koch BCP, Mouton JW, Muller AE. Review of the pharmacokinetic properties of nitrofurantoin and nitroxoline. J Antimicrob Chemother. 2018;73(11):2916–2926. doi:10.1093/jac/dky255
11. Zayyad H, Eliakim-Raz N, Leibovici L, Paul M. Revival of old antibiotics: needs, the state of evidence and expectations. Int J Antimicrob Agents. 2017;49(5):536–541. doi:10.1016/j.ijantimicag.2016.11.021
12. Vervoort J, Xavier BB, Stewardson A, et al. An in vitro deletion in ribE encoding lumazine synthase contributes to nitrofurantoin resistance in Escherichia coli. Antimicrob Agents Chemother. 2014;58(12):7225–7233. doi:10.1128/AAC.03952-14
13. Meena S, Mohapatra S, Sood S, Dhawan B, Das BK, Kapil A. Revisiting Nitrofurantoin for Vancomycin-resistant Enterococcus. J Clin Diagn Res. 2017;11(6):DC19–DC22. doi:10.7860/JCDR/2017/25140.10140
14. Osei Sekyere J. Genomic insights into nitrofurantoin resistance mechanisms and epidemiology in clinical Enterobacteriaceae. Future Sci OA. 2018;4(5):FSO293. doi:10.4155/fsoa-2017-0156
15. Xu Q, Jiang J, Zhu Z, et al. Efflux pumps AcrAB and OqxAB contribute to nitrofurantoin resistance in a uropathogenic Klebsiella pneumoniae isolate. Int J Antimicrob Agents. 2019;54(2):223–227. doi:10.1016/j.ijantimicag.2019.06.004
16. Zhang X, Zhang Y, Wang F, et al. Unravelling mechanisms of nitrofurantoin resistance and epidemiological characteristics among Escherichia coli clinical isolates. Int J Antimicrob Agents. 2018;52(2):226–232. doi:10.1016/j.ijantimicag.2018.04.021
17. Amladi AU, Abirami B, Devi SM, et al. Susceptibility profile, resistance mechanisms & efficacy ratios of fosfomycin, nitrofurantoin & colistin for carbapenem-resistant Enterobacteriaceae causing urinary tract infections. Indian J Med Res. 2019;149(2):185–191. doi:10.4103/ijmr.IJMR_2086_17
18. Mortl S, Fischer M, Richter G, Tack J, Weinkauf S, Bacher A. Biosynthesis of riboflavin: lumazine synthase of. Escherichia Coli J Biol Chem. 1996;271:33201–33207. doi:10.1074/jbc.271.52.33201
19. Chalansonnet V, Mercier C, Orenga S, Gilbert C. Identification of Enterococcus faecium enzymes with azoreductases and/or nitroreductase activity. BMC Microbiol. 2017;17(1):126. doi:10.1186/s12866-017-1033-3
20. Rafii F, Wynne R, Heinze TM, Paine DD. Mechanism of metronidazole-resistance by isolates of nitroreductase-producing Enterococcus gallinarum and Enterococcus casseliflavus from the human intestinal tract. FEMS Microbiol Lett. 2003;225(2):195–200. doi:10.1016/S0378-1097(03)00513-5
21. Li XZ, Nikaido H. Efflux-mediated drug resistance in bacteria. Drugs. 2004;64(2):159–204.
22. Shiadeh SMJ, Hashemi A, Fallah F, Rashidan M. First detection of efrAB, an ABC multidrug efflux pump in Enterococcus faecium in Tehran, Iran. Acta Microbiol Immunol Hung. 2019;66(1):57–68. doi:10.1556/030.65.2018.016
23. Sánchez Valenzuela A, Lavilla Lerma L, Benomar N, et al. Phenotypic and molecular antibiotic resistance profile of Enterococcus faecalis and Enterococcus faecium isolated from different traditional fermented foods. Foodborne Pathog Dis. 2013;10:143–149. doi:10.1089/fpd.2012.1279
24. Djahmi N, Boutet-Dubois A, Nedjai S, Dekhil M, Sotto A, Lavigne JP. Molecular epidemiology of Enterococcus sp. isolated in a university hospital in Algeria. Scand J Infect Dis. 2012;44(9):656–662. doi:10.3109/00365548.2012.673232
25. Zhang X, Bi W, Chen L, et al. Molecular mechanisms and epidemiology of fosfomycin resistance in Enterococcus isolated from patients at a teaching hospital in China, 2013–2016. J Glob Antimicrob Resist. 2020;20:191–196. doi:10.1016/j.jgar.2019.08.006
26. Cassir N, Rolain JM, Brouqui P. A new strategy to fight antimicrobial resistance: the revival of old antibiotics. Front Microbiol. 2014;5:551. doi:10.3389/fmicb.2014.00551
27. Matthews PC, Barrett LK, Warren S, et al. Oral fosfomycin for treatment of urinary tract infection: a retrospective cohort study. BMC Infect Dis. 2016;16(1):556. doi:10.1186/s12879-016-1888-1
28. Beckford-Ball J. Management of suspected bacterial urinary tract infection. Nurs Times. 2006;102(36):25–26.
29. Cunha BA, Cunha CB, Lam B, et al. Nitrofurantoin safety and effectiveness in treating acute uncomplicated cystitis (AUC) in hospitalized adults with renal insufficiency: antibiotic stewardship implications. Eur J Clin Microbiol Infect Dis. 2017;36(7):1213–1216. doi:10.1007/s10096-017-2911-1
30. Huttner A, Verhaegh EM, Harbarth S, Muller AE, Theuretzbacher U, Mouton JW. Nitrofurantoin revisited: a systematic review and meta-analysis of controlled trials. J Antimicrob Chemother. 2015;70(9):2456–2464. doi:10.1093/jac/dkv147
31. McOsker CC, Fitzpatrick PM. Nitrofurantoin: mechanism of action and implications for resistance development in common uropathogens. J Antimicrob Chemother. 1994;33:23–30. doi:10.1093/jac/33.suppl_A.23
32. Yuan L, Zhai YJ, Wu H, et al. Identification and prevalence of RND family multidrug efflux pump oqxAB genes in Enterococci isolates from swine manure in China. J Med Microbiol. 2018;67(6):733–739. doi:10.1099/jmm.0.000736
33. Li J, Zhang H, Ning J, et al. The nature and epidemiology of oqxAB, a multidrug efflux pump. Antimicrob Resist Infect Control. 2019;8:44.
34. Jonas BM, Murray BE, Weinstock GM. Characterization of emeA, a NorA homolog and multidrug resistance efflux pump, in Enterococcus faecalis. Antimicrob Agents Chemother. 2001;45(12):3574–3579. doi:10.1128/AAC.45.12.3574-3579.2001
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.