Back to Journals » OncoTargets and Therapy » Volume 13
TUG1 Promoted Tumor Progression by Sponging miR-335-5p and Regulating CXCR4-Mediated Infiltration of Pro-Tumor Immunocytes in CTNNB1-Mutated Hepatoblastoma
Authors Xie F, Zhang L, Yao Q, Shan L, Liu J, Dong N, Liang J
Received 16 October 2019
Accepted for publication 6 March 2020
Published 14 April 2020 Volume 2020:13 Pages 3105—3115
DOI https://doi.org/10.2147/OTT.S234819
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Gaetano Romano
Fujing Xie,1,* Lianhai Zhang,2,* Qing Yao,3 Liyu Shan,3 Jike Liu,2 Nanhai Dong,4 Jun Liang1
1Department of Pediatrics, Liaocheng People’s Hospital, Liaocheng 252000, Shandong Province, People’s Republic of China; 2Department of Pediatric Surgery, Liaocheng People’s Hospital, Liaocheng 252000, Shandong Province, People’s Republic of China; 3Medical College of Xi’an Jiaotong University, Xi’an 710061, Shaanxi Province, People’s Republic of China; 4Department of Clinical Laboratory, Liaocheng People’s Hospital, Liaocheng 252000, Shandong Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jun Liang; Nanhai Dong West Dongchang Road 67, Liaocheng 252000, People’s Republic of China
Tel +86 13346255358
; +86 13563016897
Email [email protected]; [email protected]
Introduction: HB presents with the highest frequency of CTNNB1 mutations, resulting in activation of Wnt signaling pathway. A number of studies have demonstrated CTNNB1 mutation contributed to the development of HB. However, limited research explored the function of lncRNAs in HB with CTNNB1 mutation.
Methods: We screened lncRNA expression profiles in CTNNB1-mutated HB samples and identified lncRNAs associated with malignant phenotype in HB. The association between lncRNA and immune microenvironment was investigated. The biological function of lncRNA was further explored using in vitro experiments.
Results: TUG1 was identified as onco-lncRNA in CTNNB1-mutated HB. TUG1 was shown to be associated with the infiltration of pro-tumor immunocytes via regulating the expression of CXCR4, a chemokine receptor playing a critical role in regulation of immune microenvironment. Inhibiting TUG1 could increase endogenous levels of miR-335-5p and consequently downregulating CXCR4, a direct target of miR-335-5p.
Conclusion: Our findings provide evidence for TUG1 mediating infiltration of pro-tumor immunocytes in HB patients carrying CTNNB1 mutation. TUG1-miR-335-5p-CXCR4 axis might be a promising immunological target for the treatment of HB patients.
Keywords: hepatoblastoma, TUG1, CXCR4, immunocyte
Introduction
Hepatoblastoma (HB) is the most common liver primary malignant tumor of the children. It is mostly at advanced stage when diagnosed, accounting for nearly 80% of primary liver malignant tumor of the children. The incidence of HB among children under 15 years old reaches one in a million. HB is derived from liver embryo tissue, which is mainly composed of immature epithelial cells similar to the fetal liver.1 Factors such as patient age, histological type of tumor, and clinical stage have been shown to affect the prognosis of patients. The conclusions of global HB clinical studies suggest that these risk factors may also be considered to the clinical differentiation and prognostic variability.2 However, there is no biomarker or international consensus for risk stratification in HB patients.
CTNNB1 is the gene encoding β-catenin, a crucial regulator of Wingless-Int (Wnt) signaling pathway, that is essential for cell proliferation, differentiation, and function, and plays an important role in stem cell renewal and organ regeneration under normal physiological condition.3 In “unstimulated” cells, β-catenin is present in the cytoplasm and is constantly targeted by phosphorylation and degradation, thereby preventing β-catenin from translocating to the nucleus to activate target gene expression. When a specific signaling pathway is stimulated (e.g., Wnt), β-catenin is stabilized and shuttles to the nucleus where it binds to the coactivators such as T cell factor (TCF)/lymphokine, resulting in β-catenin accumulation in nuclear. Aberrant expression of β-catenin activates specific genes related to tumorigenesis, and its abnormal activation has been reported in many cancer types.4 Previous study showed that activation rate of β-catenin was up to 90% in HCC.5 CTNNB1 mutation is an important pathway to β-catenin activation, conducing proliferation of abnormal cells, expressing cancer-associated membrane proteins and other characteristics of cancer cells.6 Recently, several studies reported that CTNNB1 mutation frequently associated with poor spontaneous T cell infiltration across most human cancers.7,8 Apart from T cells, whether CTNNB1 mutation affects other immunocytes in HB, such as macrophages (MΦ), neutrophils, and dendritic cells (DCs), was still largely unknown.
Long non-coding RNAs (lncRNAs) are a class of RNAs with more than 200 nucleotides in length but no protein-coding function mostly.9 Recent studies have shown that IncRNA can regulate the expression or mode of action of certain molecules by binding to certain molecules of the Hippo signaling pathway, thereby promoting or inhibiting the development and progression of tumors. In the pathologic and physiological processes of HB, as studies have indicated, lncRNA reveals association with proliferation and angiogenesis. LUCAT1 and CRNDE have been described in the biological function of hepatoblastoma.10,11 LncRNA is expected to become a new biomarker and therapeutic target for hepatoblastoma, and research on lncRNA will bring new hope to patients with hepatoblastoma or other liver diseases.12
To obtain more information, this study aims to select the lncRNA associated with HB carrying CTNNB1 mutation by using a comprehensive analysis of microarray data. Firstly, lncRNA was annotated based on gene expression profiling data to identify differential expression of lncRNA (DELs) between different prognostic subgroups. Secondly, the predicted values of DEL identified in HB patients were evaluated. The potential biological function of the identified DELs was predicted using weighted gene correlation network analysis (WGCNA). Thirdly, we conducted an in vitro experiment.
Materials and Methods
Weighted Correlation Network Analysis (WGCNA)
A microarray dataset of HB patients (GSE75271 based on Affymetrix Human Genome U133 Plus 2.0 Array platform13) was selected for WGCNA analysis. The annotation of noncoding RNA transcripts, including 8240 lncRNA transcripts, was obtained according to our previous study.14 We selected HB samples carrying CTNNB1 mutation for WGCNA.15 The clinical features included race, age, gender, HB subtype, Cairo Classification, tumor stage and histology, and prognostic characteristics. The dynamic tree cut algorithm was used to select modules of clustered lncRNAs.
TIMER Database Analysis
The correlation between lncRNA expression and immunocyte infiltration was calculated using TIMER. TIMER is a web-based tool containing enrichment of immunocytes, including B cells, CD4+ T cells, CD8+ T cells, neutrophils, macrophages, and dendritic cells. TIMER employs the deconvolution method to estimate the immunocyte infiltration and tumor purity-based multiple profiling data of 10,897 cancer samples from TCGA dataset.
Cell Culture and in vitro Transfection
An HB cell line (HepG2) was used to validate the function of candidate lncRNA.16 The HepG2 cells were purchased commercially from the Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). The cells were cultured in DMEM having 10% fetal bovine serum. The small RNA interference (siRNA) in which target candidate lncRNA was synthesized by GenePharma (Shanghai, China). We designed two siRNA sequences (siRNA #1 and #2) to select the most efficient sequence for the following experiment. The transfections of siRNA or negative control (si-Ctrl) were conducted using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA). Similarly, miR-335-5p mimic and control mimic were also transfected into cultured HepG2 cells by using Lipofectamine 2000 according to the manufacturer’s instructions.
RNA Isolation and Quantitative Real-Time PCR Analysis
RNAfast200 Kit (Fastagen Biotech, Shanghai, China) was used to isolate total RNA, and cDNA was synthesized from the total RNA (2 μg). The mRNA expression was determined by quantitative real-time PCR using the SYBR green dye (TaKaRa). And miRNA expression levels were detected applying the SYBR® PrimeScriptTM miRNA RT-PCR Kit and SYBR® Premix Ex Taq™ (TaKaRa). The mRNA expression was normalized to β-actin and the miRNA expression was normalized to U6. The relative mRNA and miRNA levels analyzed using the Comparative-Ct Method (ΔΔCt method). The primers for qRT-PCR are as follows: TUG1:5ʹ-TAGCAGTTCCCCAATCCTTG-3ʹ (sense) and 5ʹ-CACAAATTCCCATCATTCCC-3ʹ (antisense); CXCR4: 5ʹ-CTCAGACCACCGGTCTCTTC-3ʹ (sense) and 5ʹ-ATAGTCCCCTGAGCCCATTT-3ʹ (antisense); β-actin: 5ʹ- ATCGTGCGTGTGACATTAAGGAG −3ʹ (sense) and 5ʹ- AGGAAGGAAGGCTGGAAGAGTG −3ʹ (antisense). The primers of miRNAs were synthesized by TaKaRa Biotechnology according to their corresponding sequences.
Western Blotting Assay
HepG2 whole cell lysate was homogenized in a lysis buffer containing protease inhibitor mixture (Sigma-Aldrich). The protein concentration was detected by using BCA method and equal amounts of total proteins were separated by SDS-PAGE. After electrophoresis, the proteins were transferred to a polyvinylidene difluoride (PVDF) membrane. After blocking the membrane, the membranes were immunoblotted with the appropriate first antibodies (goat anti-CXCR4 antibody, ab1670, Abcam) at 4°C for 12 hrs, and incubated with corresponding secondary antibodies. Signals were monitored by chemiluminescence staining method (BioRad).
Immunofluorescence Analysis
The immunofluorescence analysis was used to determine the expression levels of CXCR4 in siRNA or si-Ctrl transfected HepG2 cells. In brief, the cells were firstly fixed and blocked, and then incubated with 1% skimmed milk for 1 hr at room temperature to block non-specific binding sites. Samples were then incubated with anti-CXCR4 polyclonal antibody (Abcam) overnight at 4°C. After being washed in PBS, after incubating with rhodamine (red)-conjugated secondary antibody, the binding sites were visualized by a laser scanning microscope (Olympus, Tokyo, Japan).
Statistical Analysis
The TUG1 expression of each subgroup in GSE75271 was presented as box and whiskers plot. The CXCR4 expression in hepG2 cells was presented as mean ± standard deviation (SD). Comparison between two subgroups was conducted by Student’s t-test and comparison between three subgroups was conducted by ANOVA analysis. Expression correlation between two genes was calculated by Pearson correlation analysis. The P value more than 0.05 was considered statistically significant.
Results
Identification of Differentially Expressed Genes That Were Associated with Clinical Features in HB Carrying CTNNB1 Mutation
We firstly employed the gene profiling data of HB samples carrying CTNNB1 mutation to identify the DEGs that were associated with clinical features (including race, age, sex, HB subtype, Cairo Classification, tumor stage, NFE2L2 mutation, TERT mutation, histology, prognosis, and survival year). The dendrogram of enrolled samples is presented as Figure 1A. By using WGCNA method, the genes were divided into 26 modules (Figure 1B). We tried to explore whether any of the groups of genes from each of the identified modules were correlated with the clinical variables of HCC. We calculated the PCCs between the modules and clinical features. Among these modules, the purple module (124 probe sets) yielded significant PCCs with HB subtypes, Cairo classification and tumor stage (HB subtypes: r=0.78, p=2e-10, Cairo classification: r=0.57, p=3e-5, tumor stage: r=0.55, p=7e-5) (Figure 1C). Previous studies have demonstrated that the above three features affected patients’ prognosis.14 The highly coexpressed genes in the purple module have potential biological significance in CTNNB1-mutated HB. Therefore, we selected the genes in MEpurple module for further investigations.
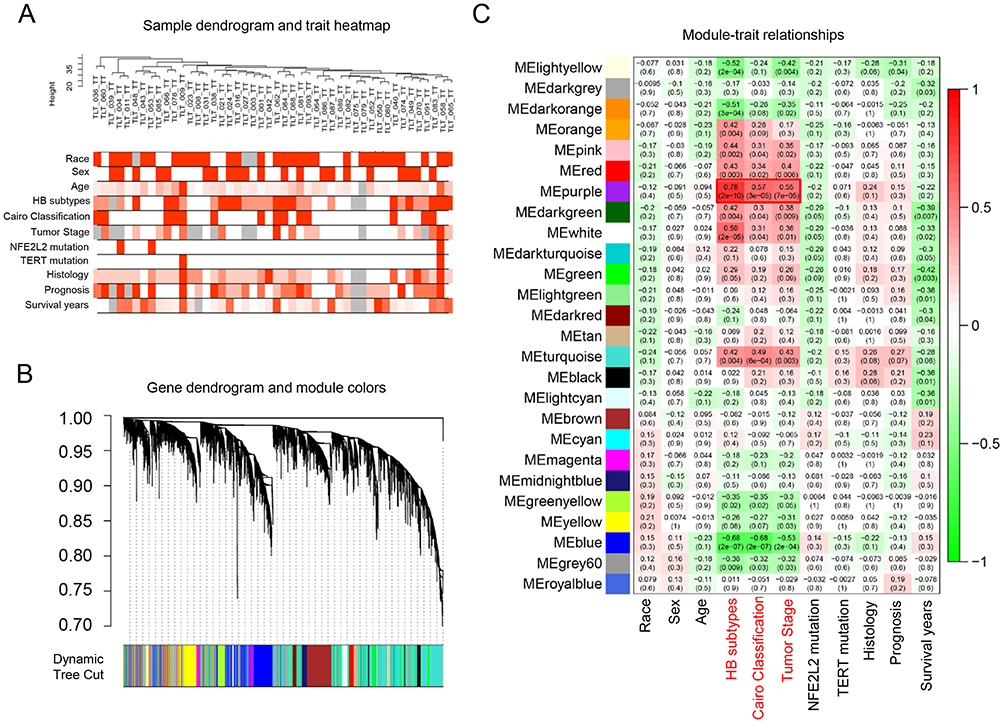 |
Figure 1 Identification of DELs that were associated with clinical features in CTNNB1-mutated HB using WGCNA. (A) Dendrogram of samples and heatmap of clinical traits. (B) Dendrogram of selected genes. (C) Module–trait relationship. |
TUG1 Was Associated with Malignant Phenotype in HB Patients Carrying CTNNB1 Mutation
According to our reported method, we annotated a total of seven lncRNAs from MEpurple module, including TUG1, LOC150776, LOC101929841, LOC101926887, LOC100289561, LOC100287015, and LINC00657. From these seven lncRNAs, TUG1 was found to be most significantly associated with the malignant features of HB patients carrying CTNNB1 mutation. We investigated the expression levels of TUG1 in different subgroups. Our results showed that the expression of TUG1 was significantly elevated in patients of HB2 and HB3 (Figure 2A). Similarly, we also found that patients with advanced tumors (Cairo C2 or stage 3/4) had higher TUG1 expression (Figure 2B and C). Moreover, those patients carrying high TUG1 expression had shorter survival time than those with low TUG1 expression (Figure 2D). The above results suggested that TUG1 might play a critical role in tumor progression in CTNNB1-mutated HB.
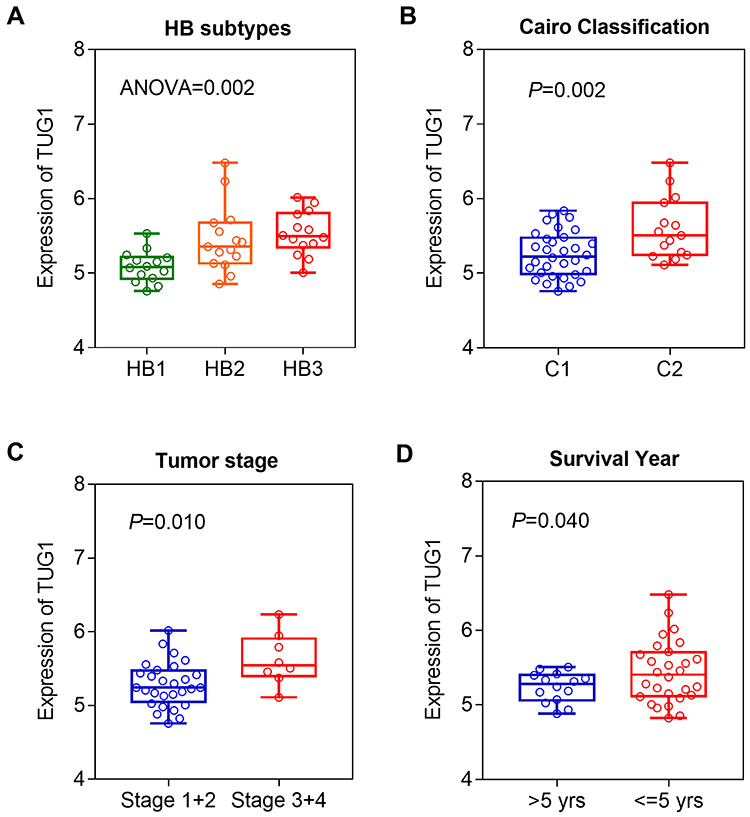 |
Figure 2 The expression levels of TUG1 in different subgroups. (A) Comparison of TUG1 expression HB1, HB2, and HB3 patients. (B) TUG1 expression in C1 and C2 subgroups. (C) TUG1 expression in patients with tumor stage 1+2 and stage 3+4. (D) TUG1 expression in long- (>5yrs) and short-time (≤5yrs) survival subgroups. |
TUG1 Was Associated with the Infiltration of Pro-Tumor Immunocytes in Liver Cancer Tissues
In our study, we confirmed that higher counts of tumor-infiltrating macrophages, neutrophils, and dendritic cells were associated with poor prognosis in liver cancer patients (Figure 3A). Interestingly, the expression of TUG1 was positively correlated with the infiltration levels of macrophages, neutrophils, and dendritic cells in liver cancer tissues (Figure 3B). Moreover, we also calculated the expression correlation between TUG1 and several immune biomarkers. The results further confirmed that TUG1 expression was positively correlated with the expression of COX2 (a macrophage marker), CCR7 (a neutrophil marker), CD1c, NRP1, and CD11c (three dendritic cell marker) (Figure 3C). The above findings suggested a correlation between high TUG1 expression and infiltration of pro-tumor immunocytes in liver cancer tissue.
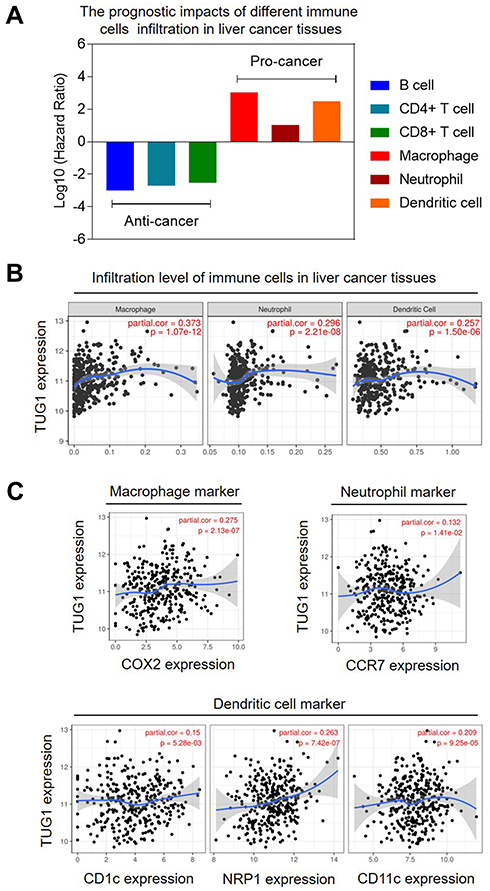 |
Figure 3 TUG1 expression was positively associated with pro-cancer immunocytes infiltration in liver cancer tissues. (A) Infiltration of macrophage, neutrophil, and dendritic cells was associated with poor prognosis. (B) TUG1 expression was positively associated with infiltration of macrophage, neutrophil, and dendritic cells. (C) TUG1 expression was positively associated with biomarkers of macrophage, neutrophil, and dendritic cells. |
TUG1 Promoted the Infiltration of Pro-Tumor Immunocytes by Regulating CXCR4 Expression
To investigate the molecular mechanism of TUG1 inducing immunocyte infiltration, the genetic correlation analysis was performed using GSE75271. We calculated the correlation coefficient between TUG1 and 57 chemokine genes (including 82 probes). The results showed that the three top probes were all targeting CXCR4 (Figure 4A). The correlation coefficients between TUG1 and three probes of CXCR4 were significantly positive (Figure 4B). Furthermore, we confirmed the expression correlation between TUG1 and CXCR4 in liver cancer tissues (TCGA dataset). After adjusted by tumor purity, TUG1 was positively correlated with CXCR4 expression (Figure 4C).
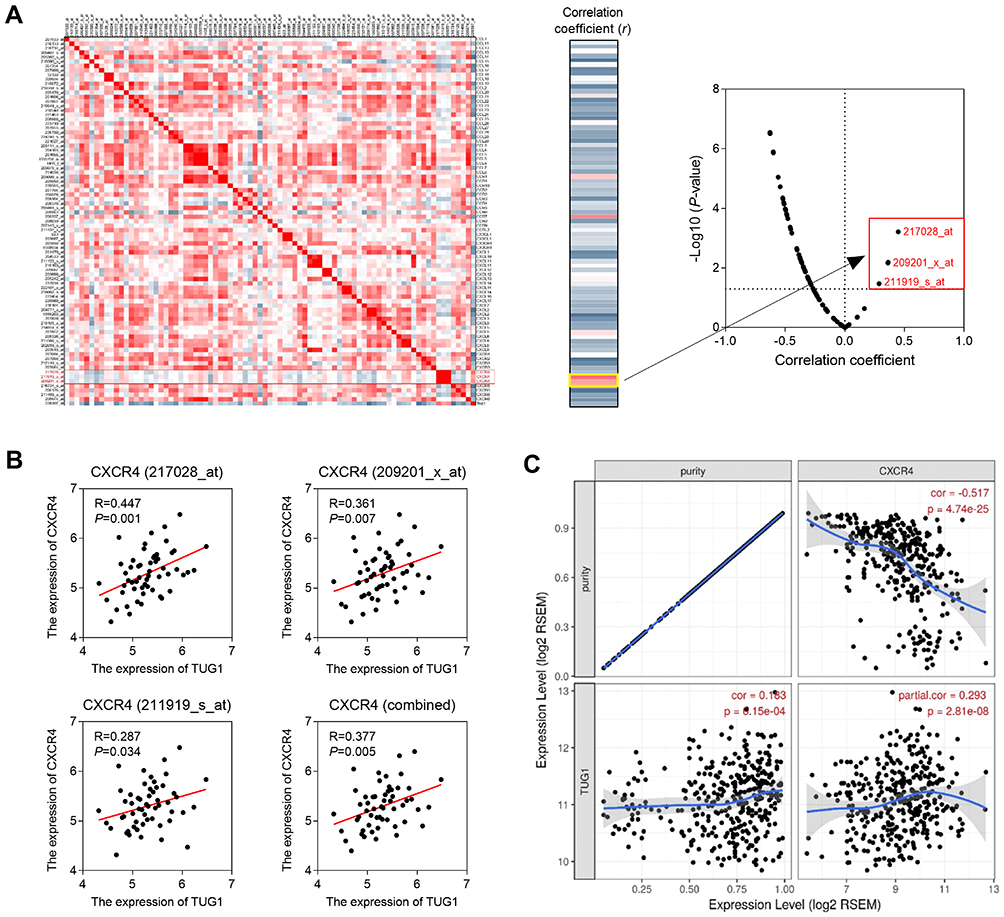 |
Figure 4 TUG1 expression was co-expressed with CXCR4. (A) The correlation coefficient between TUG1 and 57 chemokine genes (82 probes). CXCR4 expression was positively correlated with TUG1 in GSE75271 (B) and TCGA dataset (C). |
Previous studies have demonstrated that CXCR4 is involved in the immune microenvironment of the tumor.17 Targeting CXCR4 inhibited the growth and migration of malignant tumors by affecting immunocyte infiltration.18,19 We then calculated the correlation between CXCR4 and immunocyte infiltration. We found that CXCR4 expression was significantly associated with the infiltration levels of macrophages, neutrophils, and dendritic cells in liver cancer tissues (Figure 5A). Moreover, the CXCR4 expression was also positively correlated with biomarkers of macrophages (COX2), neutrophils (CCR7), and dendritic cells (CD1c, NRP1, and CD11c) (Figure 5B).
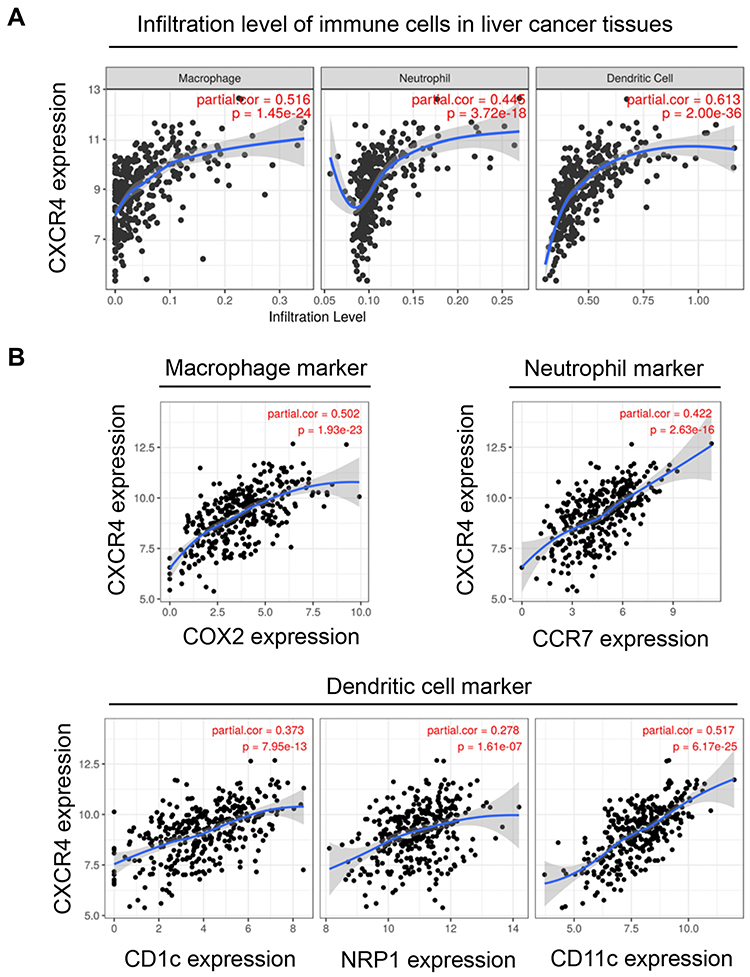 |
Figure 5 CXCR4 expression was positively associated with pro-cancer immunocytes infiltration. (A) CXCR4 expression was positively associated with infiltration of macrophage, neutrophil, and dendritic cells. (B) CXCR4 expression was positively associated with biomarkers of macrophage, neutrophil, and dendritic cells. |
Considering the critical role of CXCR4 in mediating immunocyte infiltration, we performed experiments to validate the regulation between TUG1 and CXCR4. By knockdown TUG1 expression using siRNA in HepG2, an HB cell line, the mRNA expression of CXCR4 was significantly decreased (Figure 6A). Western blotting and IF analysis also confirmed that TUG1-knockdown attenuated CXCR4 protein expression (Figure 6B and C). We also explored whether TUG1 knockdown could affect CXCL12 production in HB cells. Our data showed a non-significant link between TUG1 and CXCL12 (data not shown).
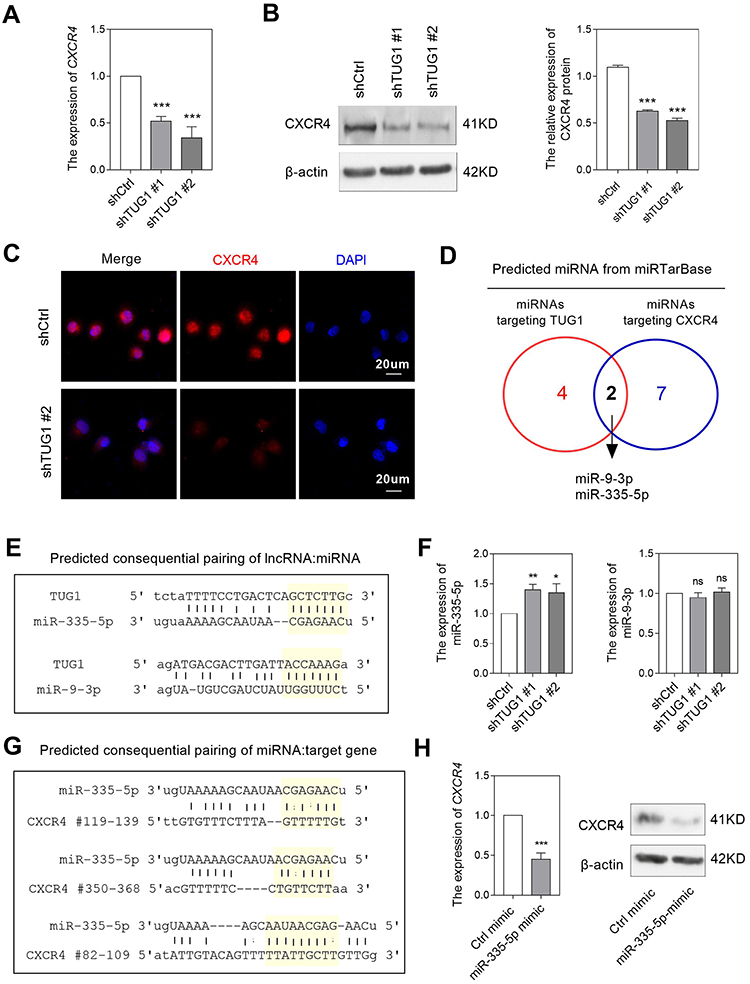 |
Figure 6 Downregulation of TUG1 inhibited CXCR4 expression. (A) The mRNA levels of CXCR4 affected by TUG1 downregulation. The effects of TUG1 downregulation on the protein levels of CXCR4 were detected by using Western blotting (B) and immunofluorescence method (C). (D) Predicted miRNAs that had capacity of binding with TUG1 and CXCR4. (E) Predicted consequential pairing of TUG1 and two miRNAs. (F) Inhibiting TUG1 increased miR-335-5p, but not miR-9-3p expression. (G) Predicted consequential pairing of miR-335-5p and CXCR4. (H) Exogenous miR-335-5p decreased CXCR4 expression at both the mRNA and protein levels. *P<0.05, **P<0.01, ***P<0.001. Abbreviation: ns, non-significant. |
TUG1 Regulated CXCR4 Expression via Sponging miR-335-5p
Previous studies showed that TUG1 could serve as ceRNA to sponge miRNA and consequently attenuate the inhibition effect of miRNAs on their downstream target.20 By using miRTarBase, we predicted two miRNAs, miR-9-3p and miR-335-5p, which had the capacity of binding both TUG1 and CXCR4 (Figure 6D and E). We further compared the expression of two miRNAs between TUG1-knockdown and control HB cells. Our data showed that miR-335-5p, but not miR-9-3p, was significantly increased in TUG1-knockdown cells (Figure 6F), suggesting miR-335-5p might be the key downstream molecule of TUG1, consistent with the work reported by Wang et al.21
Furthermore, we investigated whether miR-335-5p could affect CXCR4 expression in HB cells. By using miRanda website, miR-335-5p was predicted to bind three different 3-UTR sites of CXCR4 (Figure 6G), suggesting miR-335-5p might target CXCR4 mRNA. When we transferring miR-335-5p mimic to HB cells, we found both mRNA and protein expression of CXCR4 were decreased, comparing with the control group (Figure 6H). The above findings confirmed that TUG1 might upregulate CXCR4 expression by sponging miR-335-5p.
Discussion
In the present study, we performed a comprehensive analysis of lncRNA expression profiles in HB patients carrying CTNNB1 mutation. And TUG1 was found to be associated with malignant phenotypes. Furthermore, our study demonstrated that TUG1 promoted the infiltration of pro-tumor immunocytes via sponging miR-335-5p and regulating CXCR4 expression.
During the past decade, lncRNAs have attracted increasing attention and were found to play critical roles in multiple malignancies.22 For the past years, only few published studies focused on the relationships between lncRNAs and HB. Our previous study identified numerous differentially expressed lncRNAs between HB tissues and normal tissues, and verified LINC01314 as a tumor-suppressing lncRNA involved in Hippo signaling pathway in HB.14 Nevertheless, the functions of lncRNAs in CTNNB1-mutated HB have not been completely demonstrated so far. In the present study, we identified one lncRNA, TUG1, which was associated with the malignant phenotypes of CTNNB1-mutated HB by comparing lncRNA expression profiles between different prognostic subgroups. Our finding of positive association between TUG1 and HB was similar to Dong et al’s study,23 which found that TUG1 overexpression contributed to HB progression by regulation angiogenesis.
As we know, the infiltration of immune cells in tumor microenvironment played important and complex roles in tumor formation and development. Among these processes, innate immune cells like tumor-associated macrophages (TAMs), neutrophils (TANs), and dendritic cells mainly exert the role of promoting tumor progression,24,25 as we confirmed that higher amount of tumor-infiltrating macrophages, neutrophils, and dendritic cells were found to be associated with poor prognosis in liver cancer patients. Interestingly, in our study, we found the expression of TUG1 was positively correlated with the infiltration levels of macrophages, neutrophils, and dendritic cells in liver cancer tissues. We also confirmed that TUG1 expression was positively correlated with the expression of COX2 (a macrophage marker), CCR7 (a neutrophil marker), CD1c, NRP1, and CD11c (three dendritic cell marker). These findings suggested a correlation between high TUG1 expression and high levels of infiltration of pro-tumor immunocytes in liver cancer tissue leading to the malignant progression of CTNN1-mutated HB.
After further investigate the molecular mechanism of TUG1 inducing immunocyte infiltration, we found and confirmed that TUG1 was positively correlated with CXCR4 expression by calculating correlation coefficients. CXCR4, binding and interacting with its ligand CXCL12, could activate many downstream signaling pathways and play a critical role in mediating infiltration of immune cells like lymphocytes, neutrophils, and monocytes. Previous studies have shown that CXCR4 is involved in the immune microenvironment of the tumor26 and therefore exerts effects on cell survival, proliferation, angiogenesis, and metastasis.19 In our study, we confirmed that in liver cancer tissues, CXCR4 expression was not only significantly associated with the infiltration levels of immunocytes but also positively correlated with biomarkers of macrophages (COX2), neutrophils (CCR7), and dendritic cells (CD1c, NRP1, and CD11c). Western blotting and IF analysis confirmed that both the mRNA expression and protein expression of CXCR4 was significantly decreased in HB cells with TUG1-knockdown. Moreover, we found that miR-335-5p mediated the regulatory mechanism between TUG1 and CXCR4. Inhibiting TUG1 could increase endogenous levels of miR-335-5p and consequently downregulating CXCR4, a direct target of miR-335-5p. Taken above results together, we figured out that through sponging miR-335-5p and positively regulating the expression of CXCR4, TUG1 promotes the infiltration of pro-tumor immunocytes, thus leading to the progression and malignant phenotypes of HB.
There were several limitations in this study. In this study, we did not obtain enough evidence to confirm the effects of CTNNB1 mutation on CXCR4 expression. But several recent studies provided evidence to support our hypothesis. Firstly, CXCR4 expression was found to accompany with the activity of β-catenin.27 In HB cells, CTNNB1 mutation always caused activation of β-catenin, and consequently might induce a high level of CXCR4. Secondly, miR-335-5p, which was investigated in our additional study, was found to regulate Wnt/β-catenin signaling pathway and contribute to inhibit cancer stem cell phenotype.28 Finally, a recently published study focusing on Wilms’ tumor, another pediatric tumor with recurrently CTNNB1 mutation,29,30 reported a link of Wnt/β-catenin signaling, miR-335-5p, and TUG1.31 The above evidence supported that TUG1-miR-335-5p-CXCR7 played a critical role in CTNNB1-mutated HB.
In conclusion, our study identified the lncRNA TUG1 associated with malignant phenotypes of CTNNB1-mutated HB by performing a comprehensive analysis of lncRNA expression profiles. Furthermore, we demonstrated the mechanism that TUG1 promoted the infiltration of pro-tumor immunocytes by sponging miR-335-5p and regulating CXCR4 expression. Our findings provide evidence that TUG1 might be a promising target for antitumor immune therapy in HB patients carrying CTNNB1 mutation.
Disclosure
None of the authors have any conflict of interest to disclose.
References
1. Rowland JM. Hepatoblastoma: assessment of criteria for histologic classification. Med Pediatr Oncol. 2002;39(5):478–483. doi:10.1002/(ISSN)1096-911X
2. Maibach R, Roebuck D, Brugieres L, et al. Prognostic stratification for children with hepatoblastoma: the SIOPEL experience. Eur J Cancer. 2012;48(10):1543–1549. doi:10.1016/j.ejca.2011.12.011
3. Perugorria MJ, Olaizola P, Labiano I, et al. Wnt-beta-catenin signalling in liver development, health and disease. Nat Rev Gastroenterol Hepatol. 2019;16(2):121–136. doi:10.1038/s41575-018-0075-9
4. Litten JB, Chen TT, Schultz R, et al. Activated NOTCH2 is overexpressed in hepatoblastomas: an immunohistochemical study. Pediat Dev Pathol. 2011;14(5):378–383. doi:10.2350/10-09-0900-OA.1
5. Vilchez V, Turcios L, Marti F, Gedaly R. Targeting Wnt/beta-catenin pathway in hepatocellular carcinoma treatment. World J Gastroenterol. 2016;22(2):823–832. doi:10.3748/wjg.v22.i2.823
6. Rebouissou S, Franconi A, Calderaro J, et al. Genotype-phenotype correlation of CTNNB1 mutations reveals different ss-catenin activity associated with liver tumor progression. Hepatology. 2016;64(6):2047–2061. doi:10.1002/hep.v64.6
7. Luke JJ, Bao R, Sweis RF, Spranger S, Gajewski TF. WNT/beta-catenin pathway activation correlates with immune exclusion across human cancers. Clin Cancer Res. 2019;25(10):3074–3083. doi:10.1158/1078-0432.CCR-18-1942
8. Shimada S, Mogushi K, Akiyama Y, et al. Comprehensive molecular and immunological characterization of hepatocellular carcinoma. EBioMedicine. 2019;40:457–470. doi:10.1016/j.ebiom.2018.12.058
9. Ling H, Fabbri M, Calin GA. MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat Rev Drug Discovery. 2013;12(11):847–865. doi:10.1038/nrd4140
10. Anderson DM, Anderson KM, Chang CL, et al. A micropeptide encoded by a putative long noncoding RNA regulates muscle performance. Cell. 2015;160(4):595–606. doi:10.1016/j.cell.2015.01.009
11. Fatica A, Bozzoni I. Long non-coding RNAs: new players in cell differentiation and development. Nat Rev Genet. 2014;15(1):7–21. doi:10.1038/nrg3606
12. Liu S, Xie F, Xiang X, et al. Identification of differentially expressed genes, lncRNAs and miRNAs which are associated with tumor malignant phenotypes in hepatoblastoma patients. Oncotarget. 2017;8(57):97554–97564. doi:10.18632/oncotarget.22181
13. Sumazin P, Chen Y, Trevino LR, et al. Genomic analysis of hepatoblastoma identifies distinct molecular and prognostic subgroups. Hepatology. 2017;65(1):104–121. doi:10.1002/hep.v65.1
14. Lv B, Zhang L, Miao R, et al. Comprehensive analysis and experimental verification of LINC01314 as a tumor suppressor in hepatoblastoma. Biomed Pharmacother. 2018;98:783–792. doi:10.1016/j.biopha.2018.01.013
15. Xu X, Zhou Y, Miao R, et al. Transcriptional modules related to hepatocellular carcinoma survival: coexpression network analysis. Front Med. 2016;10(2):183–190. doi:10.1007/s11684-016-0440-4
16. Hsiao CC, Chen PH, Cheng CI, et al. Toll-like receptor-4 is a target for suppression of proliferation and chemoresistance in HepG2 hepatoblastoma cells. Cancer Lett. 2015;368(1):144–152. doi:10.1016/j.canlet.2015.08.004
17. Yang M, Zeng C, Li P, et al. Impact of CXCR4 and CXCR7 knockout by CRISPR/Cas9 on the function of triple-negative breast cancer cells. Onco Targets Ther. 2019;12:3849–3858. doi:10.2147/OTT.S195661
18. Han AR, Lee JY, Kim HJ, Min WS, Park G, Kim SH. A CXCR4 antagonist leads to tumor suppression by activation of immune cells in a leukemia-induced microenvironment. Oncol Rep. 2015;34(6):2880–2888. doi:10.3892/or.2015.4297
19. Pollino S, Palmerini E, Dozza B, et al. CXCR4 in human osteosarcoma malignant progression. The response of osteosarcoma cell lines to the fully human CXCR4 antibody MDX1338. J Bone Oncol. 2019;17:100239. doi:10.1016/j.jbo.2019.100239
20. Zhou H, Sun L, Wan F. Molecular mechanisms of TUG1 in the proliferation, apoptosis, migration and invasion of cancer cells. Oncol Lett. 2019;18(5):4393–4402. doi:10.3892/ol.2019.10848
21. Wang Y, Yang T, Zhang Z, et al. Long non-coding RNA TUG1 promotes migration and invasion by acting as a ceRNA of miR-335-5p in osteosarcoma cells. Cancer Sci. 2017;108(5):859–867. doi:10.1111/cas.13201
22. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi:10.1158/0008-5472.CAN-16-2634
23. Dong R, Liu GB, Liu BH, et al. Targeting long non-coding RNA-TUG1 inhibits tumor growth and angiogenesis in hepatoblastoma. Cell Death Dis. 2016;7(6):e2278. doi:10.1038/cddis.2016.143
24. Albini A, Bruno A, Noonan DM, Mortara L. Contribution to tumor angiogenesis from innate immune cells within the tumor microenvironment: implications for immunotherapy. Front Immunol. 2018;9:527. doi:10.3389/fimmu.2018.00527
25. Gonzalez H, Hagerling C, Werb Z. Roles of the immune system in cancer: from tumor initiation to metastatic progression. Genes Dev. 2018;32(19–20):1267–1284. doi:10.1101/gad.314617.118
26. Susek KH, Karvouni M, Alici E, Lundqvist A. The role of CXC chemokine receptors 1–4 on immune cells in the tumor microenvironment. Front Immunol. 2018;9:2159. doi:10.3389/fimmu.2018.02159
27. Yang CM, Ji S, Li Y, Fu LY, Jiang T, Meng FD. Beta-Catenin promotes cell proliferation, migration, and invasion but induces apoptosis in renal cell carcinoma. Onco Targets Ther. 2017;10:711–724. doi:10.2147/OTT.S117933
28. Chen JH, Huang WC, Bamodu OA, Chang PM, Chao TY, Huang TH. Monospecific antibody targeting of CDH11 inhibits epithelial-to-mesenchymal transition and represses cancer stem cell-like phenotype by up-regulating miR-335 in metastatic breast cancer, in vitro and in vivo. BMC Cancer. 2019;19(1):634. doi:10.1186/s12885-019-5811-1
29. Wirths O, Waha A, Weggen S, et al. Overexpression of human Dickkopf-1, an antagonist of wingless/WNT signaling, in human hepatoblastomas and Wilms’ tumors. Lab Invest. 2003;83(3):429–434. doi:10.1097/01.LAB.0000059926.66359.BD
30. Gadd S, Huff V, Walz AL, et al. A children’s oncology group and TARGET initiative exploring the genetic landscape of Wilms tumor. Nat Genet. 2017;49(10):1487–1494. doi:10.1038/ng.3940
31. Chen W, Zhuang J, Gong L, Dai Y, Diao H. Investigating the dysfunctional pathogenesis of Wilms’ tumor through a multidimensional integration strategy. Ann Transl Med. 2019;7(7):136. doi:10.21037/atm
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.