Back to Journals » Infection and Drug Resistance » Volume 17
Treatment and Management of Acinetobacter Pneumonia: Lessons Learned from Recent World Event
Authors Rangel K ![]() , De-Simone SG
, De-Simone SG ![]()
Received 26 October 2023
Accepted for publication 23 January 2024
Published 8 February 2024 Volume 2024:17 Pages 507—529
DOI https://doi.org/10.2147/IDR.S431525
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Karyne Rangel,1,2 Salvatore Giovanni De-Simone1– 4
1Center for Technological Development in Health (CDTS)/National Institute of Science and Technology for Innovation in Neglected Population Diseases (INCT-IDPN), Oswaldo Cruz Foundation (FIOCRUZ), Rio de Janeiro, RJ, 21040-900, Brazil; 2Epidemiology and Molecular Systematics Laboratory (LEMS), Oswaldo Cruz Institute, Oswaldo Cruz Foundation (FIOCRUZ), Rio de Janeiro, RJ, 21040-900, Brazil; 3Program of Post-Graduation on Science and Biotechnology, Department of Molecular and Cellular Biology, Biology Institute, Federal Fluminense University, Niterói, RJ, 22040-036, Brazil; 4Program of Post-Graduation on Parasitic Biology, Oswaldo Cruz Institute, Oswaldo Cruz Foundation (FIOCRUZ), Rio de Janeiro, RJ, 21040-900, Brazil
Correspondence: Salvatore Giovanni De-Simone, Center for Technological Development in Health (CDTS)/National Institute of Science and Technology for Innovation in Neglected Population Diseases (INCT-IDPN), Oswaldo Cruz Foundation (FIOCRUZ), Av. Brasil, 4365 - Manguinhos, Rio de Janeiro, 21040-900, Tel +55 2138658181, Email [email protected]
Abstract: Acinetobacter pneumonia is a significant healthcare-associated infection that poses a considerable challenge to clinicians due to its multidrug-resistant nature. Recent world events, such as the COVID-19 pandemic, have highlighted the need for effective treatment and management strategies for Acinetobacter pneumonia. In this review, we discuss lessons learned from recent world events, particularly the COVID-19 pandemic, in the context of the treatment and management of Acinetobacter pneumonia. We performed an extensive literature review to uncover studies and information pertinent to the topic. The COVID-19 pandemic underscored the importance of infection control measures in healthcare settings, including proper hand hygiene, isolation protocols, and personal protective equipment use, to prevent the spread of multidrug-resistant pathogens like Acinetobacter. Additionally, the pandemic highlighted the crucial role of antimicrobial stewardship programs in optimizing antibiotic use and curbing the emergence of resistance. Advances in diagnostic techniques, such as rapid molecular testing, have also proven valuable in identifying Acinetobacter infections promptly. Furthermore, due to the limited availability of antibiotics for treating infections caused A. baumannii, alternative strategies are needed like the use of antimicrobial peptides, bacteriophages and their enzymes, nanoparticles, photodynamic and chelate therapy. Recent world events, particularly the COVID-19 pandemic, have provided valuable insights into the treatment and management of Acinetobacter pneumonia. These lessons emphasize the significance of infection control, antimicrobial stewardship, and early diagnostics in combating this challenging infection.
Keywords: Acinetobacter baumannii, ventilator-associated pneumonia, VAP, hospital-acquired pneumonia, HAP, carbapenem-resistant A. baumannii, CRAB, pneumonia, COVID-19
Introduction
Nosocomial pneumonia (NP) or healthcare-associated pneumonia (HCAP) is typically categorized into two types: hospital-acquired pneumonia (HAP), defined as pneumonia acquired within 48 hours of hospital admission or within 14 days after hospital discharge, and ventilator-associated pneumonia (VAP), defined as pneumonia that develops 48 to 72 h after endotracheal intubation or tracheostomy1 These types of pneumonia constitute common challenges in healthcare, responsible for 25% of all healthcare-associated infections (HAIs) in both high-income and low/middle-income countries.1,2
Mechanical Ventilation (MV) is a non-curative supportive method with specific indications, potential hemodynamic implications, and associated complications.3 This type of support is essential for patients with altered respiratory function, which impairs gas exchange, oxygen supply for tissue perfusion, and cellular reactions.4 Pneumonia arises in response to the invasion of microorganisms into the lower respiratory system and lung parenchyma. Intubation compromises the integrity of the oropharynx and trachea, potentially enabling gastric and oral secretions to enter the lower airways.5
In Intensive Care Units (ICUs), VAP is among the most common infectious processes among invasively intubated patients receiving mechanical ventilation. It has garnered significant research attention due to its high mortality rate, adverse clinical outcomes, and the substantial costs associated with prolonged hospitalization.6
In patients with VAP, various symptoms manifest, including secretions, leukocytosis, new or progressing infiltrates, declining oxygenation, systemic infection (fever, altered white blood cell count), purulent tracheobronchial secretions, changes in sputum characteristics, and the detection of a pathogenic agent.7 Studies have reported that VAP develops in approximately 10% to 40% of intubated patients on MV, with a hospital mortality rate ranging from 24% to 76%, depending on specific circumstances and pathogens.8,9
HAP is a significant yet often underestimated patient safety concern that can affect any hospitalized patient, not just those who are critically ill. Efforts to prevent HAP should be elevated to the same level of concern as the prevention of VAP. Its overall incidence is approximately 1.6%, although this figure may be underestimated as hospitals are not obligated to report cases of HAP as they are for VAP.10 Interest in HAP has grown due to its high occurrence rate, prolonged hospital stays, substantial cost increases, and elevated mortality rates among adults (ranging from 13.9% to 30%).10,11
Acinetobacter baumannii, the predominant pathogen in healthcare-associated infections (HAIs), frequently displays significant acquired resistance and adaptability to various environments, which facilitates its widespread dissemination. The prevalence of carbapenem-resistant A. baumannii (CRAB) has notably surged among critically ill patients in recent years. CRAB represents a significant pathogen associated with both mono and polybacterial VAP, posing substantial challenges for healthcare providers.12 VAP caused by CRAB predominantly affects patients with compromised physiological status.13 Disease severity is often linked to late-onset HAP/VAP, which is a critical factor associated with increased mortality rates.14,15 Ciginskiene et al reported a hospital mortality rate of 63.3% in patients with VAP caused by drug-resistant A. baumannii.16
A growing concern is the escalating multi-drug resistance displayed by this organism, which lacks a unanimous consensus on the optimal antibiotic treatment for VAP caused by CRAB.17 However, colistin remains the most frequently used primary agent in targeted treatment regimens despite concerns about its ability to achieve adequate lung exposure safely, the risk of renal toxicity, and suboptimal clinical response rates.18–20 During the COVID-19 pandemic, the excessive use of antibiotics, coupled with disruptions in infection control measures, exacerbated the nosocomial spread of CRAB.21 Severe infections resulting in crude mortality rates as high as 50% were notable, primarily due to patients’ underlying high-risk conditions, the severity of VAP, delays in targeted CRAB treatment, and the absence of scientifically validated effective antibiotics.22–24 In this article, we will explore essential aspects of this emerging pathogen, with a specific focus on pneumonia.
Taxonomy
The discovery of the Acinetobacter spp. Genus traces back to 1911 when microbiologist Martinus Beijerinck isolated a microorganism by enriching the soil with a medium containing calcium acetate. This microorganism was initially described as Micrococcus calcoaceticus.25 Later, in 1954, Brisou and Prévot, through transformation tests and motility assessments, renamed the genus Acinetobacter spp.26 Then, in 1968, Baumann et al, after conducting biochemical evaluations of the genus’s nutritional properties, suggested its inclusion in the Moraxellaceae family.27 Despite two species of Acinetobacter spp. Already being identified by that time, Bouvet and Grimont (1986) found the characterization unsatisfactory. They proposed a study to define genospecies (genomic groups) using the thermal stability of hybrids in the DNA-DNA hybridization technique as a criterion.28 This effort led to the discovery of 12 genospecies, some of which were formally named, including A. baumannii, A. calcoaceticus, A. haemolyticus, A. johnsonii, A. junii, and A. lwoffii.28,29 To date, 108 species have been described, with 83 having validated nomenclature, including synonyms.30
Notably, the A. calcoaceticus – A. baumannii complex (ACB complex) consists primarily of nonpathogenic environmental species, such as A. calcoaceticus (genomic species 1), along with five pathogenic species linked to human diseases: A. baumannii (genomic species 2), A. pittii (formerly genomic species 3), and A. nosocomialis (once genomic species 13 TU), A. seifertii and A. dijkshoorniae.31–33 Among these species, A. baumannii is the most clinically significant due to its complex reservoir of resistance genes resulting from its high genetic plasticity, which has a direct and detrimental influence on the clinical outcomes of patients with infections caused by this pathogen.34
Acinetobacter species are gram-negative, non-fermentative, non-spore-forming, strictly aerobic, immobile, catalase-positive, non-pigmented, oxidase-negative, and exhibit coccobacillary and pleomorphic forms. Occasionally, they may appear Gram-variable or even Gram-positive during the initial Gram stain, with their morphological characteristics changing based on the growth phase. During rapid growth, they may occur rod-shaped, transitioning to a coccobacillary form during the stationary phase.35 On common diagnostic media like sheep blood agar and tryptic soy agar at 37°C, Acinetobacter colonies are grayish-white, smooth, and mucoid. Its DNA G+C content varies from 39% to 47%.29 These microorganisms belong to the phylum Proteobacteria, class γ-proteobacteria, order Pseudomonadales, and the family Moraxellaceae.29,36 They are ubiquitous and can be isolated from clinical human samples, as well as various environments such as soil, water, food, and even wild animals.37
Identification
Identifying Acinetobacter at the species level remains a complex and challenging task due to the close genetic relatedness of species within the ACB complex, coupled with their similar phenotypic and biochemical properties.38 Protocols for differentiating Acinetobacter species have been established using phenotypic traits recorded at different growth temperatures, hemolysis assays, glucose-induced acidification, and carbon/energy sources,39 as well as through the utilization of commercial automated systems (such as API 20 NE, BD Phoenix, Microscan WalkAway, Vitek 2) and DNA-based tests like PCR (16S rRNA gene amplification). However, these approaches are labor-intensive, time-consuming, and often take several days.40 Consequently, there is a growing need for molecular or genomic methods to achieve accurate identification.38,41
The blaOXA-51-like gene stands as one reliable biomarker for identifying A. baumannii, which is located chromosomally and exhibits weak hydrolysis of carbapenems. This gene, while primarily associated with A. baumannii, has also been identified in non-baumannii species.28,29,42 To enhance accuracy, techniques such as 16S rRNA sequencing, DNA-DNA hybridization, and matrix-assisted laser desorption time-of-flight mass spectrometry (MALDI-TOF-MS) are steadily employed for species identification.43–45
Innovative rapid detection assays have been developed, including the MALDIxin test, which enables the precise and swift identification of colistin-resistant A. baumannii pathogens in under 15 minutes.46 Additionally, the multiple cross-displacement amplification (MCDA) technique coupled with lateral flow biosensors (LFB) has been created for the rapid and sensitive detection of A. baumannii. This method targets the pgaD gene, a sequence-specific and conserved marker for A. baumannii.47 The results are obtained quickly, with high specificity and sensitivity, visual interpretation, without the need for specialized equipment, and at a low cost. The results obtained have shown has shown that the detection capacity of this assay exceeds that of culture methods and conventional PCR, indicating its potential as a valuable tool for the swift identification of A. baumannii in hospital settings.48
Epidemiology
Acinetobacter sp are widely distributed in nature and can be found in various ecological niches across the globe, including animals, humans, and the environment.49 Despite limited research in this area, it has become evident that food can also serve as a significant reservoir for Acinetobacter spp., including multidrug-resistant (MDR) strains. These bacteria have been isolated from various food products, including but not limited to meat, vegetables, fruit, cheese, milk, fish, and drinking water.50–52 This highlights the critical role of food as a potential source for the spread of Acinetobacter spp between community and clinical environments. It underscores the importance of conducting investigations into the potential health risks associated with foodborne pathogens.51 Moreover, it’s worth noting that meals served in healthcare facilities may act as vehicles for pathogenic bacteria, especially among vulnerable individuals, and there have been documented outbreaks in such settings.53
Acinetobacter spp. has been found in various anatomical locations in humans and may constitute a part of the skin microbiota in at least 25% of healthy individuals, with even higher prevalence rates observed among patients and hospital staff.54 In hospitalized patients, particularly during ICU outbreaks associated with mechanical ventilation or other respiratory support, colonization by A. calcoaceticus species has been reported at high percentages, including in the hands of healthcare professionals.55 Recent studies demonstrated a decrease in the rate of colonization by MDR microorganisms in ICU patients following enhanced environmental cleaning and hand hygiene in a university hospital in China.56
HAIs caused by Acinetobacter spp. can affect multiple body sites, with a predominance of respiratory, urinary tract, and surgical wound infections, especially in immunosuppressed individuals or those receiving mechanical ventilation in ICUs.57,58 Reports have also indicated the presence of Acinetobacter spp. on sink and floor surfaces in healthcare facilities, emphasizing the challenges in eradicating them even with adequate disinfection measures.59 Of significant concern is the resistance of the ACB complex to multiple antimicrobials, which is directly associated with high rates of therapeutic failure and mortality, both in Brazilian ICUs and globally.60 In recent years, there has been an evolution in the resistance profile of Acinetobacter spp., particularly an increase in the minimum inhibitory concentration (MIC) to aminoglycosides, fluoroquinolones, and carbapenems, especially imipenem.60
Among the species within the ACB complex, A. baumannii is considered a global threat in healthcare settings and has been rapidly acquiring resistance.58 In early 2019, the World Health Organization (WHO) categorized MDR A. baumannii (MDRAB) as one of the most critical pathogens due to its resistance to multiple classes of antibiotics, particularly carbapenems and third-generation cephalosporins.61 The highest incidence of A. baumannii is reported in hospital environments, especially in adult and neonatal ICUs, burns units, neurosurgery units, surgical units, medical units, and oncological units.58 While the bacterium is often found in healthcare facilities, the initial infection can also be introduced by patients admitted from outside the facility.62 Colonized or infected patients serve as reservoirs for the infection, with direct contact and the hands of healthcare staff being the primary modes of transmission.63 Airborne transmission is also possible, with the microorganism being isolated at a distance of more than four meters from patients with airway colonization.64
Extensive resistance to carbapenem antibiotics is indicative of extensively drug-resistant (XDR) bacteria, and CRAB has posed significant challenges.65 In some regions, approximately 90% of clinical isolates of A. baumannii are resistant to carbapenems. A multinational study of ICUs revealed varying prevalence rates of A. baumannii infection in different regions, with particularly high rates in Asia.66 Globally, about 45% of A. baumannii isolates are MDR, with even higher rates in Latin America and the Middle East.67 Mortality rates due to A. baumannii infections have increased in many regions over the last decade, ranging from 30% to 75%.68
The first case of NP linked to CRAB was reported in Spain in 1998,69 marking the beginning of investigations into risk factors70,71 and the impact of empirical use of antibiotics.71,72 In recent years, specifically in 2019 (51.4%) and 2020 (53%), a strong connection was established between VAP due to A. baumannii caused by MDRAB and CRAB in ICU patients.73,74 This correlation was supported not only by clinical observations but also by molecular studies.
Furthermore, an increased incidence of VAP attributed to MDRAB and CRAB was observed among patients admitted to ICUs. Molecular studies, as well as clinical data, continue to underscore this alarming trend.71,75,76 The increasing prevalence of VAP caused by MDRAB and CRAB emphasizes the critical need for focused attention on preventative strategies and alternative treatment approaches in the ICU setting.77–79
Research conducted over seven years at a tertiary center in Lebanon, focusing on MDRAB infections, revealed that the respiratory tract was the most frequent site of infection/colonization, with rates of 53.1% and 80.8%, respectively.80 In a study conducted at a university hospital, the rates of multidrug-resistant A. baumannii (MDRAB), XDR, and pan drug-resistant (PDR) strains recovered from VAP cases were 13.3%, 68.3%, and 18.3%, respectively. Within these cases, two independent risk factors for mortality were identified, namely, female sex and red blood cell transfusion.16 Furthermore, in southern Vietnam, an analysis conducted on 97 patients infected by A. baumannii through screening samples from the lower respiratory tract revealed their resistance to carbapenems and multi-drug resistance (MDR) at rates of 80% and 90%, respectively.81
A. baumannii may establish a subtle and persistent presence in patients who spend extended periods in the ICU, and this presence is closely related to the patient’s immunological status and the treatment of the primary disease. NP poses a significant threat to ventilator-dependent patients since A. baumannii can form biofilms externally in the endotracheal tube (ET), leading to excessive colonization in the lower respiratory tract. A. baumannii is capable of adhering to the respiratory tract or invasive channels within the biofilm. This fact facilitates the colonizing of the host. Therefore, it is challenging to eradicate these bacteria, and as a result, the presence of A. baumannii can persist in some patients for an extended period.82,83 According to a longitudinal prospective cohort study, the estimated risk of VAP was 1.5% per day during the first two weeks, decreasing to less than 0.5% per day after the 14th day of mechanical ventilation.84
Patients with NP, particularly those caused by MDRAB, have reported prevalence rates ranging from 40% to 95%,85–87 with associated mortality rates ranging from 45% to 85%.88 Various factors have been identified as predictors of mortality in these patients, which include unsuccessful empiric antibiotic therapy, septic shock, immunosuppression, septicemia, mechanical ventilation, long-term ICU stay, malignity, Pitt bacteremia and APACHE II score at the beginning of infection.89,90
In a study involving 164 respiratory ICU patients with A. baumannii lower respiratory tract infection, which included HAP and VAP cases, CRAB accounted for 84.8% (all MDRAB), and the mortality rate was 56.7%. The study identified several predictors strongly associated with mortality, such as the number of days spent in the ICU, the presence of CRAB infection, the duration of carbapenem use within 90 days of isolating A. baumannii, and the occurrence of septic shock.90 Furthermore, a systematic review and meta-analysis comprising 16 observational studies revealed that the mortality rate in patients infected with CRAB was twice as high as that in patients infected with carbapenem-susceptible A. baumannii.91 Additionally, another systematic review and meta-analysis, which aggregated data from 114 studies, indicated that the global prevalence of multi-drug resistance in HAP and VAP caused by A. baumannii was 79.9%.92 The highest prevalence was observed in Central America, Latin America, and the Caribbean (100% each), followed by Western Europe (91.4%), while East Asia had the lowest prevalence (64.6%). The collective mortality estimate, derived from a pooling of data from 27 studies, was found to be 42.6%.92 In a systematic review focusing on mortality predictors in a cohort of 1706 patients infected with CRAB, the reported mortality rate was 52.7%.93
A single-center, retrospective study conducted in four ICUs of a prominent Lithuanian university hospital aimed to compare the clinical features and 30-day mortality of monobacterial and polybacterial MDRAB VAP episodes. It was observed that the majority of MDRAB VAP episodes were monomicrobial (67.5%), and the study concluded that monobacterial MDRAB VAP exhibited different demographic and clinical characteristics compared to polybacterial infections and was associated with worse outcomes.94
HAP caused by A. baumannii is also a growing concern, mostly in tropical regions, identified by a serious course, a high level of bacteremia, and a remarkable mortality rate. People with a history of diabetes mellitus, alcoholism, smoking, and chronic lung disease are largely affected.95 HAP is prevalent in portions of Asia, Oceania, and Australia, including China, Taiwan, and Thailand.37
One of the most concerning manifestations of A. baumannii is the occurrence of epidemic and endemic MDR strains in hospital settings. The pattern of outbreaks may vary based on local factors and the specific strain involved, as some strains exhibit a greater propensity for epidemic dissemination than others. Epidemic strains are typically introduced into hospital environments through the admission of colonized patients.96 Since the late 1980s, A. baumannii’s clinical significance has increased dramatically due to the occurrence of outbreaks in hospitals across England, France, Germany, Italy, Spain, and Holland.29,97 Studies conducted during these outbreaks have described the coexistence of multiple clones within the same hospital, with one or two clones often predominating.98,99 The transfer of infected or colonized patients to other hospital units has been a contributing factor to inter-institutional and even international spread. In 2005, Schulte and collaborators demonstrated the clonal transmission of strains resistant to meropenem from Greece to Germany.100 Outbreaks of MDRAB infections have also been documented as complications of combat trauma in military patients during the Vietnam War and in the Middle East. Additionally, in 2004, an outbreak affected victims of a tsunami in Southeast Asia. However, the exact source of contamination remains undefined, with potential contributions from individual microbiota, the environment, or hospital units. In a study covering 12 hospital centers across three different European countries, a main clonal lineage of A. baumannii was related to VAP.101 This suggests the emergence of an XDR or PDR epidemic of A. baumannii, with resistance rates approaching 100% for carbapenems and 50% for colistin.84 Regarding HAP caused by A. baumannii, a Chinese study reported the presence of a rare sporadic clone (ST880) carrying the plasmid-encoded blaOXA-72 gene, conferring resistance to carbapenems.102
Virulence Factors and Pathogenesis
Virulence consists of the ability of the microorganism to cause disease through the production of molecules that help the microorganism in the invasion, colonization, and immunosuppression of the host.103 Among the virulence mechanisms studied, we can highlight the formation of biofilm, porins, outer membrane proteins (OMPs), outer membrane vesicles (OMVs), penicillin-binding proteins (PBPs), polysaccharide capsules, metal acquisition system, dysfunction proteins, phospholipases, lipopolysaccharides, and others.104
Studies related to genomic and phenotypic A. baumannii investigations have recognized important virulence factors responsible for its pathogenicity, demonstrating how the intrinsic and acquired mechanisms make this bacterium a major challenge for health care.105 Among the critical antigenic factors that enable immune evasion, we find the capsule polysaccharide and OmpA. Additionally, certain virulence factors promote the in vivo survival of pathogens by countering the host’s immune response, including lipopolysaccharides, phospholipase D, the iron acquisition system, outer membrane vesicles, and PBPs.106
Outer Membrane Proteins (Porins)
Porins are OMPs that are crucial in modulating cell permeability in A. baumannii.107 Among these porins, OmpA (initially identified as Omp38) stands out as the most abundant external porin found in the outer membrane of this bacterium. OmpA is the most broadly defined virulence factor in A. baumannii and exhibits in vitro model systems with many intriguing biological aspects.107,108 It possesses the capability to bind to host epithelial cells, leading to the release of proapoptotic molecules such as Cytochrome C, thereby inducing apoptosis.105 Through its interaction with fibronectin, this porin has a fundamental role in the adhesion and subsequent invasion of epithelial cells.109 Additionally, OmpA contributes to antibiotic resistance by expelling antimicrobials such as chloramphenicol, aztreonam, and nalidixic acid.105 Furthermore, it facilitates the bacteria’s persistence and promotes motility and biofilm formation.107
Another notable A. baumannii porin is the protein Omp34, previously known as Omp33-36. This porin functions as a water channel, and its expression is correlated with resistance to carbapenem antibiotics.109 Molecular research has shown that this protein is liberated inside immune and connective cells, which, by blocking autophagy, induces apoptosis, ultimately allowing for intracellular persistence with the subsequent development of cytotoxicity.110 Knockout strains of A. baumannii lacking functional Omp34 exhibit a faulty growth rate and notably lowered capacity for adhesion, invasion, and cytotoxicity, underscoring the pivotal role of Omp34 in the fitness and virulence of A. baumannii.109 CarO, also known as the carbapenem-sensitive porin, shares structural similarities with Omp34 and serves as an outer membrane channel protein with an 8-strand β-barrel structure. CarO lacks a continuous channel but mediates the influx of beta-lactam antibiotics, primarily imipenem, into A. baumannii. The CarO, like other outer membrane proteins, has the function of carbapenem resistance by facilitating antibiotic uptake.111 In clinical isolates of A. baumannii the elevated expression of CarO involveses delays pulmonary neutrophil infiltration by attenuating pro-inflammatory responses in the lungs and trachea. As a result, bacterial proliferation occurs, resulting in severe pneumonia.112
OmpW, identified in A. baumannii, bears a high resemblance to OmpW found in P. aeruginosa and E. coli. While the direct role of OmpW in A. baumannii remains unknown, in vitro-cultured colistin-resistant mutants of A. baumannii (ΔOmpW) exhibited decreased porin expression and showed less biofilm formation. However, reports have highlighted OmpW’s involvement in iron assimilation and its capacity to accommodate colistin molecules.113 Other porins, such as OprB, Omp25, OprC, and OprD, also play roles in A. baumannii physiology and virulence.114
Outer Membrane Vesicles
OmvS are spherical vesicles, typically measuring between 20–200 nm in diameter, discharged by the outer membranes of many Gram-negative bacteria, including A. baumannii.115 OMVs serve as delivery vehicles for various virulence factors into host cells and comprise lipopolysaccharides (LPS), periplasmic proteins, phospholipids, outer membrane components, and RNA or DNA.116 Numerous studies have demonstrated that A. baumannii strains secrete OMVs, carrying virulence factors like OmpA proteases, phospholipases, superoxide dismutase, and catalase. These OMVs also promote biofilm formation on abiotic surfaces.117 Interestingly, A. baumannii OMVs have been linked to the dissemination of antibiotic resistance, facilitating horizontal gene transfer of the OXA-24 carbapenemase gene.118 Given their notable part in A. baumannii virulence, OMVs from this bacterium have been investigated as potential acellular vaccines to enhance protective immunity.119
Cell Envelope Factors (Lipopolissacarídeo and Capsule)
Another crucial virulence factor is lipopolysaccharide (LPS), which is situated in the outer layer of the plasma membrane. It serves as a fundamental component of the cell surface in Gram-negative bacteria, acting as a permeability barrier that regulates the diffusion of substances into the cell. Additionally, LPS protects against environmental factors like water scarcity and antiseptics, as well as defense against phagocytosis.58,120 A key role of LPS is its contribution to resistance against normal human serum, providing a survival advantage in vivo and triggering pro-inflammatory responses.121
A. baumannii ‘s LPS consists of several components, including an O antigen, a carbohydrate core, and a lipid A moiety. It acts as a chemotactic agent, attracting inflammatory cells and inducing them to release cytotoxic substances.122
Moreover, pili, in conjunction with the antigenic polysaccharide of LPS, serve as the initial step in colonization, playing a pivotal role in promoting adhesion to host cells.123 The presence of a capsule on the bacterial surface represents another significant virulence determinant of A. baumannii. This capsule, comprised of densely packed, repetitive sugar units, forms a protective shield against environmental factors, immune system responses, and certain antimicrobial agents.124
Phospholipases
Phospholipases are crucial lipolytic enzymes involved in phospholipid metabolism and serve as important virulence factors in many Gram-negative bacteria. These enzymes exhibit lipolytic activity against phospholipids found in human cell membranes, which are the primary building blocks of biological membranes and a source of carbon and energy within the human host.125 Bacterial phospholipases generate products that aid the microorganism in evading phagocytic processes, thereby facilitating tissue colonization, infection progression, and suppression of the immune system.126
In the case of A. baumannii, three types of phospholipases are described: phospholipase A (PLA), phospholipase C (PLC), and phospholipase D (PLD), each contributing to different virulence mechanisms.127 PLCs are cytotoxic to epithelial cells, and their combined action with ethanol intensifies A. baumannii’s cytotoxicity. Specifically, 1,2-diacylglycerol, released by cellular phospholipase C, plays a role in modifying the biophysical properties of the membrane, including charge, fluidity, and permeability. It can also recruit cytosolic proteins, leading to spatial reorganization of signaling complexes that affect various cellular processes.127 PLDs, on the other hand, collectively promote serum resistance, epithelial cell invasion, and in vivo pathogenesis.128 When acting together with PLA and PLC, PLD contributes to cell invasion by hydrolyzing the fatty acids of the glycerol backbone, with subsequent PLC-mediated cleavage of the phosphorylated head group released from the glycerol backbone. PLD specifically cleaves its main group, generating instability in host cells and affecting cell signaling.128
All these phospholipases exhibit substrate specificity for phosphatidylcholine,127 a prominent component of eukaryotic membranes, constituting 50% of all phospholipids and increasing to 80% in pulmonary and tracheobronchial secretions.129 Experimental evidence suggests that phosphatidylcholine can serve as a nutrient source during pulmonary infections caused by pathogens like Pseudomonas aeruginosa and A. baumannii.130 The degradation of phospholipids by these enzymes compromises the stability of host cell membranes and directly interferes with cell signaling, resulting in alterations in the host’s immune response.127
Another identified virulence factor is the CpaA enzyme, characterized as a metallo-endopeptidase with similarities to adamalysin, specifically targeting glycans. CpaA interferes with the blood clotting cascade by inactivating factor XII, leading to an increase in clotting time, as observed in human and murine serum (PTTa).131 CpaA can cleave coagulation Factor V in human plasma, and analyses of Prothrombin Time (PT), fibrinogen, and TTPa have shown reduced coagulation in TTPa and fibrinogen but an increase in PT. Consequently, CpaA attenuates thrombus formation in intravascular sites, facilitating A. baumannii ‘s spread.132
Metal Acquisition System
The acquisition of essential micronutrients such as iron, zinc, and manganese are critical for A. baumannii and other bacterial species, both for host-pathogen interaction, reproduction, and pathogenesis.133 Despite iron’s abundant presence in both environmental and biological systems, the accessibility of biologically active ferric iron (Fe+3) remains relatively limited. This limitation arises from its reduced solubility under aerobic conditions and at neutral pH, as well as its propensity for chelation by low molecular weight compounds like heme or high-affinity iron-binding molecules such as lactoferrin and transferrin (iron-binding proteins).134 Hosts restrict the availability of these nutrients in their bloodstream to prevent bacterial propagation in the organism.135 In mammals, iron is usually bound to proteins such as transferrin, lactoferrin, and ferritin, thus limiting its serum availability.136 Nevertheless, to surmount this iron scarcity, the majority of aerobic bacteria synthesize high-affinity iron-chelating agents known as siderophores. These are low molecular weight compounds with a pronounced affinity for iron.137 Acinetobactin is the best-characterized siderophore in A. baumannii, occurring more frequently in MDRAB isolates.138 A. baumannii expresses several genes related to the iron acquisition system, such as bas A-J, which are responsible for the synthesis of these siderophores, barA/B which carry out the efflux and bauF accountable for the uptake of iron after its chelation.139 Hosts defend themselves against microorganisms through the sequestration of metals through nutritional immunity, where proteins called calprotectin are recruited to sites of infection with the function of restricting the access of bacteria to zinc and magnesium through a chelation process. Restriction of these compounds leads to inhibition of microbial growth and reduces virulence.140 A. baumannii has important mechanisms involved in establishing and progressing infections that are still poorly understood. Among these factors, there are protein secretion systems (SSP) named according to their type, 2 (T2SS) and 6 (T6SS).141 Most A. baumannii strains have T6SS. However, studies show that even with this factor, activation or deactivation occurs through interbacterial interactions or the loss of T6SS repressor plasmids.142 T2SS exhibits a structural resemblance to type IV pili systems and functions by facilitating the transport of proteins from the periplasmic space to the external environment.143
Biofilm/Quorum-Sensing
Among all virulence determinants, biofilm formation on biotic and abiotic surfaces has become an important feature of A. baumannii pathogenesis, making the MDR organism challenging in healthcare.144 Biofilms are complex three-dimensional structures composed of bacteria, which live in an extracellular matrix made of polysaccharides, proteins, cellular debris, and genetic material and are produced by microorganisms serving as protection against desiccation, oxidation, resistance to some antibiotics, radiation, and host defense cells.145
Biofilm formation begins differently between biotic and abiotic environments, where in biotic environments protein-protein bonds occur on the bacterial surface with the human cellular matrix. In abiotic locations, bacterial interaction with the surface depends on its hydrophobicity.146 The following steps are the same for both environments, which involve bacterial proliferation at the adhered site, followed by incorporation into an extracellular matrix, where maturation occurs where biofilm maturation occurs and, finally, its dispersion.120
A. baumannii possesses a chaperon/usher pilus system referred to as Csu pili, which is under the regulation of the BfmRS two-component system. This intricate network of molecules plays a pivotal role in influencing gene expression, enabling the formation of a protective capsule in response to antibiotic exposure and environmental stress. Furthermore, this same system regulates the formation of pili, an essential process for adhesion and biofilm formation for cell attachment.147 The presence of another regulatory system is also observed, GacSA, which can affect biofilm formation through the expression of Csu and thus intervene in the microorganism’s ability to produce biofilm.106 Another virulence mechanism is the protein associated with biofilm production, known as BapAb, which has also been described in S. aureus. This protein has the function of modulating the hydrophobicity of the cell surface, thus resulting in better adherence to epithelial cells.148 A significant factor contributing to A. baumannii’s biofilm formation is the production of the exopolysaccharide poly-β-1,6-N-acetylglucosamine (PNAG), a trait common among many Gram-negative species. PNAG is crucial for adhesion and aggregation processes.149
Studies have revealed that over 60% of hospital-acquired infections are attributed to bacteria capable of forming biofilms on medical devices.150 Among these devices, the ET presents an ideal environment for bacterial and fungal adhesion, supporting biofilm development on both its inner luminal surface and outer surface.151 The ET is recognized as an independent risk factor for pulmonary infections in intubated patients, elevating this risk by a factor of 6 to 10.152,153
In intubated patients, biofilm formation within the ET is an early and persistent occurrence, effectively serving as a reservoir for infectious microorganisms. Almost immediately after intubation, a mixed biofilm with the potential to harbor various microbial pathogens takes shape within the ET.154 Microorganisms can inhabit liquid mediums as solitary cells or form immobilized communities within biofilms. These microorganisms can enter the trachea through microaspiration or via biofilm development, particularly on the inner surface of the distal third of the ET.155,156
Transmission of these biofilms to the lower airways leads to serious complications such as pneumonia or sepsis.157 Different tracheostomy tube materials showed no change in susceptibility to biofilm formation.158 Prolonged tracheostomy duration had no significant correlation with biofilms, and they were formed as early as seven days.158,159 In a study that assessed the biofilm formation and antibiotic resistance of bacterial profile from ET of patients admitted to ICUs in southwest Iran, 22% (n = 121/546) of GNB isolated were A. baumannii with most exhibiting MDR (77%) and 74% (n=89) were biofilm producers.160 A prospective observational study carried out intending to know the most common organisms capable of forming a biofilm on the ET and their sensitivity to antibiotics showed that A. baumannii (45%) was the most common biofilm producer isolated exhibiting 35.7% MDR.161
The formation of biofilms on the surface of the ET is intricately tied to the pathogenesis of VAP. Fragments of biofilm become dislodged and are passively transported towards the lungs.162 Additionally, biofilm cells can become aerosolized and aspirated into the lungs due to gas flow during mechanical ventilation. In contrast, individual cells may be dislodged by fluids and migrate deeper into the lungs.154,162 A noteworthy study indicated that 70% of VAP patients harbored the same pathogens in both ET biofilms and lung tissues. This observation underscores the substantial and persistent role that biofilms play as a source of pathogenic bacteria.163 In a separate study aimed at understanding biofilm production by pathogens responsible for VAP and its connection to drug resistance, it was found that out of the 71 isolates examined, the A. calcoaceticus-baumannii complex was the second most frequently isolated organism, accounting for 16.9% of cases.164
Recently, a study compared the biofilm formation capacity, antimicrobial resistance patterns, and molecular typing based on Pulsed-Field Gel Electrophoresis (PFGE) in A. baumannii isolated from burn (23/50) and VAP patients (27/50). Overall, 74% of the strains were MDR and 26% were XDR. Regarding biofilm formation capacity, 52%, 36%, and 12% of the isolates were strong, moderate, and weak biofilm producers, and all the isolates harbored at least one biofilm-related gene.165
In A. baumannii, quorum sensing (QS) plays a crucial role in bacterial survival and pathogenicity. QS is recognized as a communication mechanism between bacterial communities to maintain population density. The QS system coordinates the behavior of individual bacteria in a population by mediating the synthesis, secretion, and binding of easily diffusible small hormone-like molecules known as autoinducers (AIs) that are released in proportion to cell density.166 These mechanisms are advantageous for the entire community, particularly at a higher cell density, eg, synthesis of virulence factors, formation of biofilms, and production of siderophores and proteases.167
Acinetobacter generates acyl homoserine lactones (AHLs) as signaling molecules for both inter- and intraspecies communication. The Quorum Sensing (QS) cycle in A. baumannii is orchestrated by the presence of the inducer AbaI and its cognate receptor AbaR. AbaI, which is encoded by the abaI gene, serves as a sensory protein responsible for producing AHL signaling molecules as autoinducers.168 In contrast, AbaR functions as a receptor protein, and upon binding to AHLs, it initiates a cascade of reactions. This binding event triggers a positive feedback loop, leading to the increased production of AHLs, ultimately regulating biofilm formation. It’s noteworthy that the deletion of the abaI synthase autoinducer gene in A. baumannii results in reduced biofilm formation and pathogenicity.169 Furthermore, the abaI/abaR QS system has been observed to exert influence on overgrowth characteristics, morphology, biofilm formation, resistance, motility, and virulence.170 Additionally, A. baumannii produces less-studied signaling molecules, such as diketopiperazines, 2-heptyl-3-hydroxy-4-quinolone, and retention factor 1, each of which contributes to the complex signaling network in the organism.171
Diagnosis
The diagnostic criteria for NP, including HAP and VAP, are intricate, making it challenging to differentiate between true infection and colonization. Various methodologies have been detailed in the literature for epidemiological surveillance and diagnostic purposes. However, more consensus has yet to emerge regarding the most appropriate set of criteria.172 Despite the ongoing debate, clinical symptoms, although sometimes nonspecific, serve as initial criteria for diagnosis. Radiological examinations and laboratory and microbiological criteria, such as bacteriological cultures, are also essential for defining the etiological agent.173 Clinical and radiological data, while valuable, exhibit limited specificity. The use of quantitative microbiological criteria can enhance specificity, but their accuracy has been called into question.174,175
It is crucial to note that VAP is considered a performance indicator in the USA and some other countries. According to a report by the National Healthcare Society Network (NHSC), the effective implementation of a multifaceted infection control program significantly reduced the incidence rate of VAP.176 The Center for Disease Control and Prevention (CDC) reported a 70% decrease in the incidence of VAP between 2006 and 2012. However, during the same period, VAP rates reported by the Medicare Patient Safety Monitoring System remained consistently high, affecting approximately 10% of ventilated patients.177 These findings highlight a notable disparity between the rates documented in a quality monitoring program and those mentioned in the patient assistance program.
Several methods are available for diagnosing VAP (Table 1), including invasive and non-invasive techniques to obtain clinical specimens:
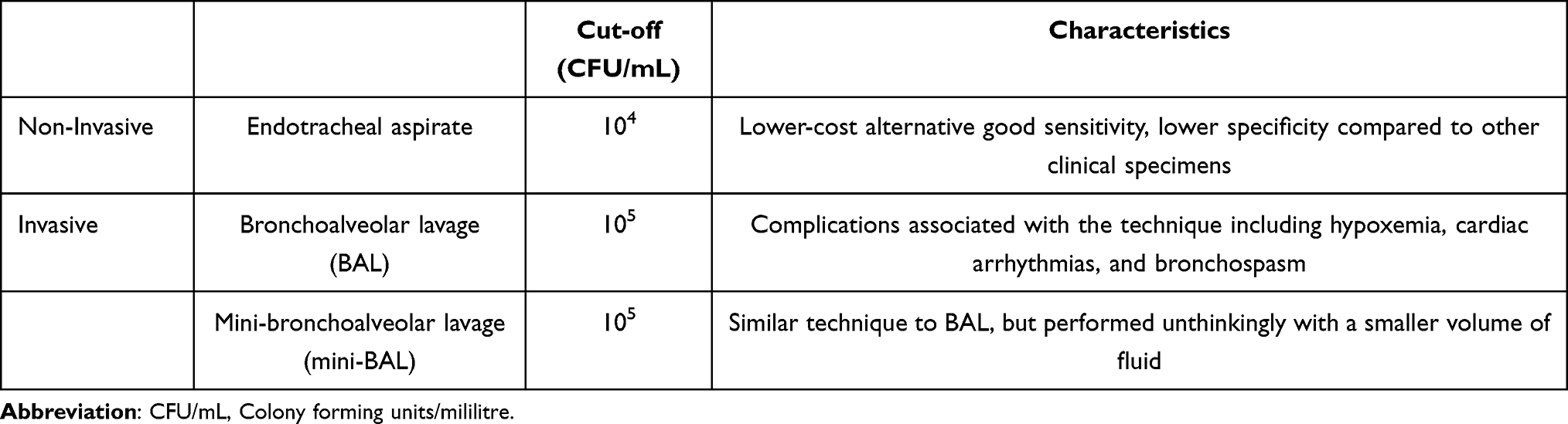 |
Table 1 Methods Available for Diagnosing Ventilator-Associated Pneumonia (VAP) |
Endotracheal aspirate: This is an easy, quick, and non-invasive technique, but it may yield high rates of false positives due to airway colonization.178 In quantitative cultures, a cut-off value of 104 Colony Forming Units (CFU)/mL is used.179 While the endotracheal aspirate offers a lower-cost alternative and good sensitivity, its specificity is lower compared to other clinical specimens. However, the analysis of the endotracheal aspirate using the quantitative technique with a cut-offpoint of 10^6 CFU/mL shows a good correlation with results obtained from minimally contaminated specimens.174
Bronchoalveolar lavage: This technique requires a bronchoscope to recover cellular and non-cellular components of the bronchioles and alveoli using a fluid vehicle, with a cut-off of 105 CFU/mL.179 Complications associated with the technique may include hypoxemia, cardiac arrhythmias, and bronchospasm.5
Mini-bronchoalveolar lavage (mini-BAL): This is a similar technique to bronchoalveolar lavage but performed unthinkingly with a smaller volume of fluid while maintaining the same cut-off value.179
The latter two methods are considered minimally contaminated by upper respiratory tract secretions but are more expensive and invasive. Bacterial growth in quantitative cultures with counts exceeding the mentioned cut-offpoints indicates, with higher specificity, that the isolated clinical specimen is associated with the etiology of VAP, while lower counts suggest colonization.179,180 It’s worth emphasizing that routine bacteriology techniques used to detect the etiological agent of VAP are time-consuming and less sensitive compared to molecular techniques.29 The optimization of molecular techniques for VAP diagnosis, especially for infections caused by A. baumannii, is imperative and could positively impact patients’ clinical outcomes.
Treatment
Current guidelines for the treatment of A. baumannii infections are greatly impacted by the rapid spread of resistance to commonly used antibiotics, severely limiting therapeutic options and posing a significant challenge.37 In the case of VAP patients suspected or confirmed to have the condition, immediate initiation of antibiotic therapy is crucial. Delaying treatment is associated with a significant increase in mortality rates and healthcare costs for survivors.13
The initial treatment of VAP in critically ill patients, particularly those in units with a high prevalence of A. baumannii infections or those previously colonized, relies on empirical choices. However, selecting the appropriate initial antibiotics is crucial, as an inadequate choice is linked to higher mortality rates.181 Additionally, the retrieval of multidrug-resistant (MDR) bacteria is strictly related to a growing risk of unsuitable therapy.182 Unfortunately, there is a lack of data on the optimal antimicrobial regimen for the treatment of VAP caused by MDR/XDR/PDR pathogens. Consequently, there needs to be a consensus on the strategy for treatment choice, which often relies on the doctor’s experience.183
Numerous risk factors contribute to VAP caused by A. baumannii. These include the duration of mechanical ventilation, reintubation, antibiotic therapy, perioperative blood product transfusion, enteral nutrition, supine head position, nasogastric tubes intra-hospital transport, lack of subglottic secretion drainage, paralyzing agents use, continuous sedation, and constant changes in the ventilator circuit. Host-related risk factors encompass medical history, underlying diseases, male gender, extreme age, acute underlying diseases, previous central nervous system disorders, acute renal failure, immunocompromised status, emergent surgery, neurosurgery, acute respiratory distress syndrome, thoracic and cardiac surgery, burns, intra-aortic support, reintervention, ulcer disease and extracorporeal membrane oxygenation (ECMO).184–186
The propensity of A. baumannii to develop antibiotic resistance complicates the selection of empiric antimicrobial regimens that can be applied consistently across different healthcare facilities.187 Few randomized clinical trials have focused on defining optimal therapies for pneumonia caused by these difficult-to-treat resistant strains, resulting in limited therapeutic strategies against A. baumannii.188
For susceptible A. baumannii isolates, initial therapy typically involves carbapenems like imipenem-cilastatin, meropenem, or doripenem (except ertapenem), depending on local susceptibility patterns.189 Imipenem, historically, has been regarded as the gold standard for treating VAP caused by A. baumannii.190 Clinical cure rates with imipenem range from 57% to 83%.191 It is important to highlight that susceptibility tests are necessary before clinical use, as isolates susceptible to imipenem may be resistant to meropenem and vice versa.190
In settings with high local resistance to carbapenems, empirical combined therapy is recommended to ensure broad coverage. This approach also guides the reevaluation of initial therapy based on microbiological results and clinical response variables such as fever and oxygenation support.189 To enhance patient outcomes and reduce the risk of resistance, a common strategy involves the use of combination therapy until drug susceptibility test results become available, aiming to broaden antimicrobial coverage. However, the results from human trials are limited, and the clinical data supporting this approach still need to be more conclusive.192 As an alternative to carbapenem-sparing monotherapy, high doses of sulbactam can be employed for the treatment of A. baumannii pneumonia, whether the strains are susceptible or resistant to carbapenems.193 Sulbactam, an antibiotic acting as a β-lactamase inhibitor, directly exhibits antimicrobial activity against A. baumannii through its affinity for the bacterium’s PBPs.194
While some studies have shown clinical and bacteriological efficacy, other evidence is contradictory, suggesting that sulbactam-based therapies may not be superior to non-sulbactam regimens (including colistin, cephalosporins, and tigecycline) for A. baumannii treatment.27,195,196
Other agents, such as minocycline and tigecycline, have shown clinical success in treating VAP caused by A. baumannii, particularly in cases of multi-drug resistance.197 However, the efficacy and safety of these antibiotics can vary, with some studies indicating inferior outcomes compared to other agents.198,199 Cefiderocol, a novel β-lactam with activity against CRAB, has shown promise, although its role in treating pulmonary infections caused by this pathogen remains controversial due to limited evidence.200,201
The global rise in carbapenem resistance among A. baumannii strains has significantly narrowed the antibiotic arsenal.37 polymyxin B and colistin (polymyxin E) are increasingly used to treat bacteremia, A. baumannii meningitis, and VAP.190 Although effective in suppressing A. baumannii growth in vitro, these antibiotics have limitations such as a narrow therapeutic window, bacteriostatic action, variable pharmacokinetics/pharmacodynamics, high rates of neurotoxicity, and an increased risk of nephrotoxicity, especially with loading doses.202 Additionally, they exhibit poor pulmonary penetration, which limits their utility.189,203 There is also a need for more consensus on whether colistin should be used alone or in combination therapy. However, combining colistin with other antibiotics like carbapenems due to synergism can lead to better results.204
Inhaled aerosolized antibiotics with colistin and tobramycin have been explored as treatment options for VAP, particularly in cases of difficult-to-treat pathogens. Inhaled antibiotics allow for higher local concentrations and can reduce the need for toxic intravenous antibiotics. However, there is limited and conflicting data on their efficacy and adverse effects, with some studies indicating potential benefits in terms of reduced mortality and hospital stay. In contrast, others suggest respiratory complications and no significant impact on mortality.205–208
Finally, the treatment of A. baumannii infections is a complex challenge due to antibiotic resistance, especially in the situation of VAP (Table 2). While some antibiotics, such as colistin and sulbactam, show promise in treating these infections, the choice of therapy is often based on local susceptibility patterns and clinical judgment. Further research and clinical trials are needed to establish definitive treatment guidelines for A. baumannii infections, especially those caused by multidrug-resistant strains.
 |
Table 2 Initial Teraphy to A. baumannii Pneumonia |
The widespread prevalence of antimicrobial resistance in A. baumannii, especially against last-resort antibiotics, has sounded the alarm in recent years. Therefore, various studies have been conducted to provide new treatment strategies.209 A potential alternative management for treating infections with MDR A. baumannii is bacteriophages, the most widely encountered organisms on the planet.210 Bacteriophage therapy (using viruses to treat bacterial infections) is a promising antibacterial approach, which treats and prevents infectious disease by lysis of bacterial cells.211 Another approach is the use of antimicrobial peptides (AMPs), components of the innate immune system of eukaryotic and prokaryotic organisms that have broad-spectrum and low resistance, which has attracted the attention of researchers.212 Silver nanoparticles have also demonstrated antimicrobial activity against various microbes, including A. baumannii, likely caused by their diverse bactericidal mechanisms.213 These nanoparticles coupled with plant extract have also been intensively studied.214
Prevention
The guidelines provided by the Infectious Disease Society of America (IDSA) and the American Thoracic Society (ATS) offer a comprehensive overview of proven methods for preventing VAP and HAP. To effectively reduce the occurrence of these disorders, the primary approach involves minimizing exposure to previously identified intervention-related risk factors and host-related risk factors. Whenever feasible, intubation should be avoided, and alternative procedures, such as non-invasive ventilation with sedation and positive pressure, should be employed. Additionally, implementing weaning protocols is essential to reduce or shorten the duration of mechanical ventilation. However, it’s worth noting that the timing of tracheotomy does not significantly impact the incidence of VAP.215 A range of preventive measures should be considered for patients at risk of VAP. These encompass risk-reduction strategies in ventilation (such as avoiding intubation, minimizing sedation, and enhancing physical fitness), follow-up measures (including education, performance measurement with feedback, improving the overall safety culture in healthcare settings, and public reporting), and preventive measures (such as changing the ventilation circuit only when visibly soiled or malfunctioning, selective oropharyngeal decontamination (SOD) or selective digestive decontamination (SDD), using endotracheal tubes with subglottic drainage of secretions, regular mouthwash with chlorhexidine for hygiene, and prophylactic probiotics).216
The optimal combination of essential preventive measures remains uncertain, and it is unlikely that any single preventive strategy can eliminate VAP.217 Utilizing a few key care interventions derived from evidence-based guidelines tends to yield more favorable outcomes when implemented consistently as a collective package rather than in isolation. It’s worth noting that while recent studies have shown significant success in reducing VAP rates with specific precautions, meta-analyses have revealed that most preventive measures have failed to demonstrate sustained effectiveness.218,219 In fact, there has been no significant improvement over the past decade.177
However, Selective Digestive Decontamination (SDD) and Selective Oropharyngeal Decontamination (SOD) have emerged as the only preventive measures that have a positive impact on mortality.220 On the other hand, oral hygiene with chlorhexidine has not demonstrated a significant effect on VAP rates, the duration of mechanical ventilation, or the length of stay in the Intensive Care Unit (ICU).221
Currently, a clinical trial titled “RGNOSIS: Ecological Effects of Decolonization Strategies in Intensive Care” is underway in six European countries. This trial encompasses various interventions, including a control group, oral hygiene with chlorhexidine, SDD, and SOD. The researchers intend to evaluate the effects of these treatments on the incidence of antibiotic-resistant bacteria and bacterial infections in the ICU. The study also seeks to determine the most effective treatment. The control group will receive standard treatment, while the treatment groups will receive 1% chlorhexidine oral gel (CHX), an antibiotic mouth paste containing three different antibiotics (SOD), and an antibiotic mouth paste and suspension for the stomach and intestines containing three other antibiotics (SDD). All treatments will be administered four times a day to eliminate harmful bacteria in the digestive tract (SDD) and the mouth (CHX, SOD, SDD).222
Some studies reported a need for more adherence by doctors and nurses in complying with published guidelines for VAP prevention.223,224 An online cross-sectional quantitative survey was conducted in two Australian ICUs to explore nurses’ knowledge of VAP and self-reported adherence to evidence-based guidelines for the prevention of ventilator-associated events.225 According to a qualitative descriptive study on the opinion of health professionals about the barriers to the implementation of evidence-based practices in the prevention of events associated with mechanical ventilation, it was possible to address previously reported deficits in nurses’ knowledge and adherence to practice based on mechanical ventilation evidence in intensive care. The results suggested the need for a well-established policy to sustain the practice. The barriers faced by nurses and physicians in preventing events associated with mechanical ventilation need to be addressed to optimize the quality of patient care in ICUs.226
It’s essential to keep in mind that to encourage individuals to adopt specific behaviors and enhance adherence, we must address predisposing factors, which include knowledge, perceptions, and beliefs. By addressing these factors, we can facilitate access to new processes or technologies and consistently reinforce correct behaviors.
Non-compliance with these measures is likely more closely tied to behavioral, structural, and organizational aspects than to the nursing workload. Offering specific education about ventilator-associated events can be instrumental in raising awareness and promoting adherence to guidelines.227,228
Pneumonia and COVID-19
Infections caused by A. baumannii are pivotal in influencing hospital outcomes during the novel Coronavirus Disease 2019 (COVID-19) pandemic, exerting a significant impact on global public health. Overall, almost 25% of COVID-19 patients necessitated intensive care, leading to their hospitalization in the intensive care unit (ICU) because of acute respiratory distress syndrome (ARDS).229 Recent data indicates that as many as 80% of COVID-19 patients admitted to the ICU required respiratory support via mechanical ventilation due to severe respiratory failure induced by the virus in more critical cases. This additional factor increases the risk of developing VAP(VAP).230 Moreover, new challenges have arisen in managing the occurrence of VAPs, as the clinical and radiological criteria traditionally employed for diagnosing VAP do not apply to critically ill patients with pneumonia stemming from SARS-CoV-2 infection. Since these patients present a clinical picture that overlaps with that caused by VAPs, with fever, hypoxemia, intense leukocytosis, and extensive bilateral radiological infiltrate, the microbiological criterion was the only one to support the diagnosis of VAP in patients with COVID-19.231 As per a conducted meta-analysis, it was found that during their stay in intensive care Unit (ICU), as many as 48% of ventilated COVID-19 patients developed VAP(VAP). Several other studies corroborate these findings, with an average of around 50% of ICU-admitted patients developing VAP.232 The incidence of VAP ranged from 36% to 85%, while ICU mortality rates varied between 29% and 43%.230,232,233
Septic shock and severe acute respiratory syndrome (ARDS) are the primary factors associated with mortality in VAP. However, specific characteristics and risk factors of VAP in COVID-19 patients have yet to be established.234 Notably, experiencing multiple episodes of VAP in the context of COVID-19 prolongs ICU stays and increases the risk of colonization and infection by CRAB by 7.5 and 5.5 times, respectively.235,236 The literature describes a higher incidence and recurrence rate of VAP during COVID-19 than in the pre-COVID-19 period, with reports demonstrating a high rate of VAP in patients with COVID-19 when compared to those observed in non-COVID-19 patients.237–239 However, we observed a decrease in the incidence of VAP in critically ill non-COVID-19 patients during the pandemic compared to before the pandemic, especially concerning A. baumannii.240 Although studies are heterogeneous and sample sizes are limited, the median time to onset of VAP in patients with COVID-19 does not appear to differ from that reported in VAP in non-COVID-19 patients.229,241 Dysbiotic oral microbiota also has the potential to colonize the lower respiratory tract causing VAP and a recent study revealed substantial qualitative and quantitative dysbiosis with a high frequency of potentially pathogenic species, including A. baumannii.242
Further compounding the impact of the pandemic are bacterial coinfections and secondary infections triggered by SARS-CoV-2, which often lurk as hidden threats amidst the backdrop of COVID-19. These infections, exemplified by those attributed to A. baumannii, have intensified the severity and risks associated with SARS-CoV-2 infection.243 They play a significant role in the morbidity and mortality of COVID-19 patients. Individuals with COVID-19 who develop coinfections or secondary infections involving A. baumannii tend to experience more adverse outcomes, including prolonged hospital and ICU stays, as well as higher mortality rates. It’s vital to emphasize that these outcomes are primarily influenced by bacterial infections rather than viral ones.244
Given the challenges of diagnosing coinfections with VAP in intubated COVID-19 patients, a significant portion receive broad-spectrum antimicrobials preventatively, even in the absence of evidence of bacterial coinfection.245 This approach, while aiming to be proactive, elevates the risk of selecting multidrug-resistant microorganisms, particularly in patients expected to require prolonged mechanical ventilation. Notably, implementing the cleaning and disinfection of mechanical ventilators for SARS-CoV-2 patients using enzymatic detergents and isopropyl alcohol has demonstrated positive effects by reducing post-disinfection microbial contamination rates.246
The challenges faced by healthcare professionals have been further compounded by the emergence of outbreaks involving multidrug-resistant (MDR) and XDR A. baumannii.247–249 Approximately 50% of hospitalized COVID-19 patients’ deaths are attributed to secondary bacterial infections, with MDR A. baumannii frequently identified as a causative agent, associated with a two-fold increase in COVID-19-related mortality.250
It is well-established that there is a connection between increased antibiotic use and the rise of antimicrobial resistance in ICUs. During the pandemic, in-hospital antimicrobial usage has surged due to concerns about bacterial and fungal coinfections alongside COVID-19. In the context of VAP with COVID-19, the term “early antibiotic therapy” is introduced, but treatments often commence before patients deteriorate and require endotracheal intubation and mechanical ventilation.251
It’s important to note that antibiotic administration has been identified as a risk factor for the onset of VAP, particularly because a substantial proportion of patients with COVID-19 (73%) and those without (88%) receive high levels of antibiotic therapy before the development of VAP. Additionally, COVID-19 patients exhibit a higher rate of multidrug-resistant bacterial colonization upon ICU admission (31% vs 16%) but do not demonstrate a greater risk of acquiring multidrug-resistant bacteria during their ICU stay. Consequently, antibiotic treatment in COVID-19 patients should be reserved for cases with well-documented bacterial infections to minimize the risk of VAP caused by multidrug-resistant bacteria.16
Conclusion
When the acquisition of resistance traits became significant, A. baumannii emerged as a troublesome nosocomial pathogen, significantly complicating the treatment process. Its genetic flexibility has enabled this bacterium to evolve rapidly, thereby elevating it to one of the gravest threats facing hospitalized patients. Currently, multidrug-resistant and pan-drug-resistant A. baumannii strains have become pervasive, causing a depletion in available treatment options. The emergence of CRAB strains further compounds these challenges.
Several virulence mechanisms are associated with multidrug-resistant A. baumannii strains, particularly the development of biofilms on endotracheal surfaces, which are closely linked to the pathogenesis of VAP. This association, especially for patients admitted to Intensive Care Units, is alarming and represents a significant global medical issue. As such, the timely diagnosis of A. baumannii infections is of utmost priority to mitigate harm to patients, and the administration of appropriate and timely antimicrobial therapy is essential for VAP treatment.
Given the lack of effective therapeutic agents, the optimal treatment for A. baumannii pneumonia remains undefined, with polymyxins, either alone or in combination with other agents, being empirically recommended. VAP in patients with COVID-19 presents a unique challenge for clinicians, given the high incidence of septic shock and mortality associated with this infection. The prevalence of antibiotic resistance among pathogens isolated from mechanically ventilated COVID-19 patients in the ICU underscores the importance of preventing coinfections caused by this pathogen. This situation calls for a standardized approach to antibiotic administration in COVID-19 patients to ensure treatment success.
From a microbiological perspective, the potential association of A. baumannii with bloodstream and polymicrobial infections should be carefully considered when prescribing antibiotic treatment, whether empirical or targeted, to minimize the risk of treatment failure and VAP recurrence.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, and interpretation, or all these areas; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by the Carlos Chagas Filho Foundation for Research Support of the State of Rio de Janeiro/FAPERJ (#110.198-13) and the Brazilian Council for Scientific Research (CNPq, #467.488/2014-2 and 301744/2019-0). Funding was also provided by FAPERJ (#210.003/2018) through the National Institutes of Science and Technology Program (INCT) to Carlos M. Morel (INCT-IDPN).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Kalil AC, Metersky ML, Klompas M, et al. Management of adults with hospital-acquired and ventilator-associated pneumonia: 2016 Clinical Practice Guidelines by the Infectious Diseases Society of America and the American Thoracic Society. Clin Infect Dis. 2016;63(5):e61–e111. doi:10.1093/cid/ciw353
2. Munro S, Baker D. Reducing missed oral care opportunities to prevent non-ventilator associated hospital acquired pneumonia at the Department of Veterans Affairs. Appl Nurs Res. 2018;44:48–53. doi:10.1016/j.apnr.2018.09.004
3. Hickey SM, Giwa AO. Mechanical Ventilation. Treasure Island (FL): StatPearls Publishing; 2023.
4. Antalová N, Klučka J, Říhová M, Poláčková S, Pokorná A, Štourač P. VAPprevention in pediatric patients: narrative review. Children. 2022;9(10):1540. doi:10.3390/children9101540
5. Charles MP, Kali A, Easow JM, et al. Ventilator-associated pneumonia. Australas Med J. 2014;7(8):334–344. doi:10.4066/AMJ.2014.2105
6. Papazian L, Klompas M, Luyt CE. VAPin adults: a narrative review. Intensive Care Med. 2020;46(5):888–906. doi:10.1007/s00134-020-05980-0
7. American Thoracic Society; Infectious Diseases Society of America. Guidelines for the management of adults with hospital-acquired, ventilator-associated, and healthcare-associated pneumonia. Am J Respir Crit Care Med. 2005;171(4):388–416. doi:10.1164/rccm.200405-644ST
8. Seguin P, Laviolle B, Dahyot-Fizelier C, et al. Effect of oropharyngeal povidone-iodine preventive oral care on VAP in severely brain-injured or cerebral hemorrhage patients: a multicenter, randomized controlled trial. Crit Care Med. 2014;42(1):1–8. doi:10.1097/CCM.0b013e3182a2770f
9. Herkel T, Uvizl R, Doubravska L, et al. Epidemiology of hospital-acquired pneumonia: results of a Central European multicenter, prospective, observational study compared with data from the European region. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2016;160(3):448–455. doi:10.5507/bp.2016.014
10. Giuliano KK, Baker D, Quinn B. The epidemiology of nonventilator hospital-acquired pneumonia in the United States. Am J Infect Control. 2018;46(3):322–327. doi:10.1016/j.ajic.2017.09.005
11. Magill SS, Edwards JR, Bamberg W, et al. Multistate point-prevalence survey of health care-associated infections. N Engl J Med. 2014;370(13):1198–1208. doi:10.1056/NEJMoa1306801
12. Cai B, Echols R, Magee G, et al. Prevalence of carbapenem-resistant gram-negative infections in the United States predominated by Acinetobacter baumannii and Pseudomonas aeruginosa. Open Forum Infect Dis. 2017;4(3):ofx176. doi:10.1093/ofid/ofx176
13. Zilberberg MD, Nathanson BH, Sulham K, Fan W, Shorr AF. Multidrug resistance, inappropriate empiric therapy, and hospital mortality in Acinetobacter baumannii pneumonia and sepsis. Crit Care. 2016;20(1):221. doi:10.1186/s13054-016-1392-4
14. Timsit JF, Esaied W, Neuville M, Bouadma L, Mourvllier B. Update on ventilator-associated pneumonia. F1000Res. 2017;6:2061. doi:10.12688/f1000research.12222.1
15. Arayasukawat P, So-Ngern A, Reechaipichitkul W, Chumpangern W, Arunsurat I, Ratanawatkul P. Microorganisms, and clinical outcomes of early- and late-onset VAPat Srinagarind Hospital, a tertiary center in Northeastern Thailand. BMC Pulm Med. 2021;21(1):47. doi:10.1186/s12890-021-01415-8
16. Čiginskienė A, Dambrauskienė A, Rello J, Adukauskienė D. VAPdue to drug-resistant Acinetobacter baumannii: risk factors and mortality relation with resistance profiles, and independent predictors of in-hospital mortality. medicina. 2019;55(2):49. doi:10.3390/medicina55020
17. Tamma PD, Aitken SL, Bonomo RA, Mathers AJ, van Duin D, Clancy CJ. Infectious diseases society of America guidance on the treatment of AmpC β-lactamase-producing Enterobacterales, carbapenem-resistant Acinetobacter baumannii, and Stenotrophomonas maltophilia infections. Clin Infect Dis. 2022;74(12):2089–2114. doi:10.1093/cid/ciab1013
18. Drwiega EN, Rodvold KA. Penetration of Antibacterial Agents into Pulmonary Epithelial Lining Fluid: an Update. Clin Pharmacokinet. 2022;61(1):17–46. doi:10.1007/s40262-021-01061-7
19. Eljaaly K, Bidell MR, Gandhi RG, et al. Colistin nephrotoxicity: meta-analysis of randomized controlled trials. Open Forum Infect Dis. 2021;8(2):ofab026. doi:10.1093/ofid/ofab026
20. Aydemir H, Akduman D, Piskin N, et al. Colistin vs. the combination of colistin and rifampicin for the treatment of carbapenem-resistant Acinetobacter baumannii ventilator-associated pneumonia. Epidemiol Infect. 2013;141(6):1214–1222. doi:10.1017/S095026881200194X
21. Segala FV, Bavaro DF, Di Gennaro F, et al. Impact of SARS-CoV-2 Epidemic on antimicrobial resistance: a literature review. Viruses. 2021;13(11):2110. doi:10.3390/v13112110
22. Vincent JL, Sakr Y, Singer M, et al. Prevalence and outcomes of infection among patients in intensive care units in 2017. JAMA. 2020;323(15):1478–1487. doi:10.1001/jama.2020.2717
23. Tacconelli E, Carrara E, Savoldi A, et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 2018;18(3):318–327. doi:10.1016/S1473-3099(17)30753-3
24. Zilberberg MD, Nathanson BH, Sulham K, Fan W, Shorr AF. A novel algorithm to analyze epidemiology and outcomes of carbapenem resistance among patients with hospital-acquired and ventilator-associated pneumonia: a retrospective cohort study. Chest. 2019;155(6):1119–1130. doi:10.1016/j.chest.2018.12.024
25. Beijerinck M. Pigmenten als oxydatieproducten gevormd door bacterien. Versl Koninklijke Akad Wetensch Amsterdam. 1911;19:1092–1103.
26. Brisou J, Prevot AR. Etudes de systématique bactérienne. Ann Inst Pasteur. 1954;86(6):722–728.
27. Baumann P, Doudoroff M, Stanier RY. A study of the Moraxella group. II. Oxidative-negative species (genus Acinetobacter). J Bacteriol. 1968;95(5):1520–1541. doi:10.1128/jb.95.5.1520-1541
28. Bouvet PJ, Grimont PA. Taxonomy of the genus Acinetobacter with the recognition of Acinetobacter baumannii sp. nov., Acinetobacter haemolyticus sp. nov., Acinetobacter johnsonii sp. nov., and Acinetobacter junii sp. nov., and emended description of Acinetobacter calcoaceticus and Acinetobacter lwoffii. Int J Syst Bacteriol. 1986;36:228–240.
29. Peleg AY, Seifert H, Paterson DL. Acinetobacter baumannii: emergence of a successful pathogen. Clin Microbiol Rev. 2008;21(3):538–582. doi:10.1128/CMR.00058-07
30. LPSN.dsmz.de; 2023. Available from: https://www.bacterio.net/genus/acinetobacter.
31. Gerner-Smidt P. Ribotyping of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex. J Clin Microbiol. 1992;30(10):2680–2685. doi:10.1128/jcm.30.10.2680-2685.1992
32. Nemec A, Krizova L, Maixnerova M, Sedo O, Brisse S, Higgins PG. Acinetobacter seifertii sp. nov., a member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex isolated from human clinical specimens. Int J Syst Evol Microbiol. 2015;65(Pt3):934–942. doi:10.1099/ijs.0.000043
33. Osgaya C, Marí-Almirall M, Van Assche A, et al. Acinetobacter dijkshoorniae sp. nov., a member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex mainly recovered from clinical samples in different countries. Int J Syst Evol Microbiol. 2016;66(10):4105–4111. doi:10.1099/ijsem.0.001318
34. Ramirez MS, Bonomo RA, Tolmasky ME. Carbapenemases: transforming Acinetobacter baumannii into a yet more dangerous menace. Biomolecules. 2020;10(5):720. doi:10.3390/biom10050720
35. Visca P, Seifert H, Towner KJ. Acinetobacter infection-an emerging threat to human health. IUBMB Life. 2011;63(12):1048–1054. doi:10.1002/iub.534
36. Rossau R, Van Landschoot A, Gillis M, De Ley J. Taxonomy of Moraxellaceae fam. nov., a New bacterial family to accommodate the genera Moraxella, Acinetobacter, and Psychrobacter and related organisms. Int J Syst Evolut Microbiol. 1991;41:310–319. doi:10.1099/00207713-41-2-310
37. Wong D, Nielsen TB, Bonomo RA, Pantapalangkoor P, Luna B, Spellberg B. Clinical and pathophysiological overview of Acinetobacter Infections: a century of challenges. Clin Microbiol Rev. 2017;30(1):409–447. doi:10.1128/CMR.00058-16
38. Sarshar M, Behzadi P, Scribano D, Palamara AT, Ambrosi C. Acinetobacter baumannii: an ancient commensal with weapons of a pathogen. Pathogens. 2021;10(4):387. doi:10.3390/pathogens1004
39. Vijayakumar S, Biswas I, Veeraraghavan B. Accurate identification of clinically important Acinetobacter spp.: an update. Future Sci OA. 2019;5(6):FSO395. doi:10.2144/fsoa-2018-0127
40. Li Y, Yang X, Zhao W. Emerging microtechnologies and automated systems for rapid bacterial identification and antibiotic susceptibility testing. SLAS Technol. 2017;22(6):585–608. doi:10.1177/2472630317727519
41. Evans BA, Amyes SG. OXA β-lactamases. Clin Microbiol Rev. 2014;27(2):241–263. doi:10.1128/CMR.00117-13
42. Lee YT, Turton JF, Chen TL, et al. First identification of blaOXA-51-like in non-baumannii Acinetobacter spp. J Chemother. 2009;21(5):514–520. doi:10.1179/joc.2009.21.5.514
43. Tjernberg I, Ursing J. Clinical strains of Acinetobacter classified by DNA-DNA hybridization. APMIS. 1989;97(7):595–605. doi:10.1111/j.1699-0463.1989.tb00449.x
44. Misbah S, Hassan H, Yusof MY, Hanifah YA, AbuBakar S. Genomic species identification of Acinetobacter of clinical isolates by 16S rDNA sequencing. Singapore Med J. 2005;46(9):461–464.
45. Šedo O, Nemec A, Křížová L, Kačalová M, Zdráhal Z. Improvement of MALDI-TOF MS profiling for the differentiation of species within the Acinetobacter calcoaceticus-Acinetobacter baumannii complex. Syst Appl Microbiol. 2013;36(8):572–578. doi:10.1016/j.syapm.2013.08.001
46. Dortet L, Potron A, Bonnin RA, et al. Rapid detection of colistin resistance in Acinetobacter baumannii using MALDI-TOF-based lipidomics on intact bacteria. Sci Rep. 2018;8(1):16910. doi:10.1038/s41598-018-35041-y
47. Wang Q, Zhou Y, Li S, et al. Real-time fluorescence loop mediated isothermal amplification for the detection of Acinetobacter baumannii. PLoS One. 2013;8(7):e66406. doi:10.1371/journal.pone.0066406
48. Cheng X, Yang J, Wang M, et al. Visual and rapid detection of Acinetobacter baumannii by a multiple cross displacement amplification combined with nanoparticles-based biosensor assay. AMB Express. 2019;9(1):30. doi:10.1186/s13568-019-0754-0
49. Doughari HJ, Ndakidemi PA, Human IS, Benade S. The ecology, biology and pathogenesis of Acinetobacter spp.: an overview. Microbes Environ. 2011;26(2):101–112. doi:10.1264/jsme2me10179
50. Amorim AM, Nascimento JD. Acinetobacter: an underrated foodborne pathogen? J Infect Dev Ctries. 2017;11(2):111–114. doi:10.3855/jidc.8418
51. Carvalheira A, Silva J, Teixeira P. Acinetobacter spp. in food and drinking water - A review. Food Microbiol. 2021;95:103675. doi:10.1016/j.fm.2020.103675
52. Carvalheira A, Casquete R, Silva J, Teixeira P. Prevalence and antimicrobial susceptibility of Acinetobacter spp. isolated from meat. Int J Food Microbiol. 2017;243:58–63. doi:10.1016/j.ijfoodmicro.2016.12.001
53. Lund BM. Provision of microbiologically safe food for vulnerable people in hospitals, care homes and in the community. Food Control. 2019;97:535–547. doi:10.1016/j.foodcont
54. Rodríguez-Baño J, Cisneros JM, Fernández-Cuenca F, et al. Clinical features and epidemiology of Acinetobacter baumannii colonization and infection in Spanish hospitals. Infect Control Hosp Epidemiol. 2004;25(10):819–824. doi:10.1086/502302
55. Bergogne-Bérézin E, Towner KJ. Acinetobacter spp. as nosocomial pathogens: microbiological, clinical, and epidemiological features. Clin Microbiol Rev. 1996;9(2):148–165. doi:10.1128/CMR.9.2.148
56. Li Y, Ge H, Zhou H, et al. Impact of environmental cleaning on the colonization and infection rates of multidrug-resistant Acinetobacter baumannii in patients within the intensive care unit in a tertiary hospital. Antimicrob Resist Infect Control. 2021;10(1):4. doi:10.1186/s13756-020-00870-y
57. Biderman P, Bugaevsky Y, Ben-Zvi H, Bishara J. Multidrug-resistant Acinetobacter baumannii infections in lung transplant patients in the cardiothoracic intensive care unit. Clin Transplant. 2015;29(9):756–762. doi:10.1111/ctr.12575
58. Harding CM, Hennon SW, Feldman MF. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat Rev Microbiol. 2018;16(2):91–102. doi:10.1038/nrmicro.2017.148
59. Alouane T, Uwingabiye J, Lemnouer A, et al. First whole-genome sequences of two multidrug-resistant Acinetobacter baumannii Strains Isolated from a Moroccan Hospital Floor. Genome Announc. 2017;5(18):e00298–17. doi:10.1128/genomeA.00298-17
60. Lupo A, Haenni M, Madec JY. Antimicrobial resistance in Acinetobacter spp. And Pseudomonas spp. Microbiol Spectr. 2018;6:3. doi:10.1128/microbiolspec.ARBA-0007-2017
61. Krol V, Hamid NS, Cunha BA. Neurosurgically related nosocomial Acinetobacter baumannii meningitis: report of two cases and literature review. J Hosp Infect. 2009;71(2):176–180. doi:10.1016/j.jhin.2008.09.018
62. Peacock SJ, Parkhill J, Brown NM. Changing the paradigm for hospital outbreak detection by leading with genomic surveillance of nosocomial pathogens. Microbiology. 2018;164(10):1213–1219. doi:10.1099/mic.0.000700
63. Joly-Guillou M-L, Brun-Buisson C. Epidemiology of Acinetobacter spp.: surveillance and management of outbreaks. In: Acinetobacter. CRC Press; 2020:28,71–100. doi:10.1201/9781003069263-4
64. Antunes LC, Visca P, Towner KJ. Acinetobacter baumannii: evolution of a global pathogen. Pathog Dis. 2014;71(3):292–301. doi:10.1111/2049-632X.12125
65. Ma C, McClean S. Mapping Global Prevalence of Acinetobacter baumannii and Recent Vaccine Development to Tackle It. Vaccines. 2021;9(6):570. doi:10.3390/vaccines9060570
66. Gautam D, Dolma KG, Khandelwal B, et al. Acinetobacter baumannii: an overview of emerging multidrug-resistant pathogen. Med J Malaysia. 2022;77(3):357–370.
67. Colquhoun JM, Rather PN. Insights Into Mechanisms of Biofilm Formation in Acinetobacter baumannii and Implications for Uropathogenesis. Front Cell Infect Microbiol. 2020;10:253. doi:10.3389/fcimb.2020.00253
68. Garnacho-Montero J, Gutiérrez-Pizarraya A, Díaz-Martín A, et al. Acinetobacter baumannii in critically ill patients: molecular epidemiology, clinical features and predictors of mortality. Enferm Infecc Microbiol Clin. 2016;34(9):551–558. doi:10.1016/j.eimc.2015.11.018
69. López-Hernández S, Alarcón T, López-Brea M. Carbapenem resistance mediated by beta-lactamases in clinical isolates of Acinetobacter baumannii in Spain. Eur J Clin Microbiol Infect Dis. 1998;17(4):282–285. doi:10.1007/BF01699988
70. Husni RN, Goldstein LS, Arroliga AC, et al. Risk factors for an outbreak of multi-drug-resistant Acinetobacter nosocomial pneumonia among intubated patients. Chest. 1999;115(5):1378–1382. doi:10.1378/chest.115.5.1378
71. Garnacho-Montero J, Ortiz-Leyba C, Fernández-Hinojosa E, et al. Acinetobacter baumannii ventilator-associated pneumonia: epidemiological and clinical findings. Intensive Care Med. 2005;31(5):649–655. doi:10.1007/s00134-005-2598-0
72. Iregui M, Ward S, Sherman G, Fraser VJ, Kollef MH. Clinical importance of delays in the initiation of appropriate antibiotic treatment for ventilator-associated pneumonia. Chest. 2002;122(1):262–268. doi:10.1378/chest.122.1.262
73. Sader HS, Castanheira M, Mendes RE, Flamm RK. Frequency and antimicrobial susceptibility of Gram-negative bacteria isolated from patients with pneumonia hospitalized in ICUs of US medical centres (2015-17). J Antimicrob Chemother. 2018;73(11):3053–3059. doi:10.1093/jac/dky279
74. Saleem M, Syed Khaja AS, Hossain A, et al. Molecular characterization and antibiogram of Acinetobacter baumannii clinical isolates recovered from the patients with ventilator-associated pneumonia. Healthcare. 2022;10(11):2210. doi:10.3390/healthcare10112210
75. Royer S, Faria AL, Seki LM, et al. Spread of multidrug-resistant Acinetobacter baumannii and Pseudomonas aeruginosa clones in patients with VAPin an adult intensive care unit at a university hospital. Braz J Infect Dis. 2015;19(4):350–357. doi:10.1016/j.bjid.2015.03.009
76. Koulenti D, Lisboa T, Brun-Buisson C, et al. Spectrum of practice in the diagnosis of nosocomial pneumonia in patients requiring mechanical ventilation in European intensive care units. Crit Care Med. 2009;37(8):2360–2368. doi:10.1097/CCM.0b013e3181a037ac
77. Bassetti M, Righi E, Vena A, Graziano E, Russo A, Peghin M. Risk stratification and treatment of ICU-acquired pneumonia caused by multidrug- resistant/extensively drug-resistant/pandrug-resistant bacteria. Curr Opin Crit Care. 2018;24(5):385–393. doi:10.1097/MCC.0000000000000534
78. Behera B, Mahapatra A, Kunjan Pillai JS, et al. Infection-Related ventilator-associated complication and possible VAPamong mechanically ventilated patients of adult medical and surgical intensive care units. J Lab Physicians. 2022;15(1):45–47. doi:10.1055/s-0042-1750076
79. He Q, Wang W, Zhu S, et al. The epidemiology and clinical outcomes of ventilator-associated events among 20,769 mechanically ventilated patients at intensive care units: an observational study. Crit Care. 2021;25(1):44. doi:10.1186/s13054-021-03484-x
80. Kanafani ZA, Zahreddine N, Tayyar R, et al. Multi-drug resistant Acinetobacter species: a seven-year experience from a tertiary care center in Lebanon. Antimicrob Resist Infect Control. 2018;7:9. doi:10.1186/s13756-017-0297-6
81. Hoang Quoc C. Carbapenemase genes and multidrug resistance of Acinetobacter baumannii: a cross sectional study of patients with pneumonia in southern Vietnam. Antibiotics. 2019;8(3):148. doi:10.3390/antibiotics8030148
82. Gedefie A, Demsis W, Ashagrie M, et al. Acinetobacter baumannii Biofilm formation and its role in disease pathogenesis: a review. Infect Drug Resist. 2021;14:3711–3719. doi:10.2147/IDR.S332051
83. Howard A, O’Donoghue M, Feeney A, Sleator RD. Acinetobacter baumannii: an emerging opportunistic pathogen. Virulence. 2012;3(3):243–250. doi:10.4161/viru.19700
84. Bouadma L, Sonneville R, Garrouste-Orgeas M, et al. Ventilator-associated events: prevalence, outcome, and relationship with ventilator-associated pneumonia. Crit Care Med. 2015;43(9):1798–1806. doi:10.1097/CCM.0000000000001091
85. Nowak J, Zander E, Stefanik D, et al. High incidence of pandrug-resistant Acinetobacter baumannii isolates collected from patients with VAPin Greece, Italy and Spain as part of the MagicBullet clinical trial. J Antimicrob Chemother. 2017;72(12):3277–3282. doi:10.1093/jac/dkx322
86. Almomani BA, McCullough A, Gharaibeh R, Samrah S. Incidence, and predictors of 14-day mortality in multidrug-resistant Acinetobacter baumannii in ventilator-associated pneumonia. J Infect Dev Ctries. 2015;9(12):1323–1330. doi:10.3855/jidc.6812
87. Ziółkowski G, Pawłowska I, Krawczyk L, Wojkowska-Mach J. Antibiotic consumption versus the prevalence of multidrug-resistant Acinetobacter baumannii and Clostridium difficile infections at an ICU from 2014-2015. J Infect Public Health. 2018;11(5):626–630. doi:10.1016/j.jiph.2018.02.003
88. Özgür ES, Horasan ES, Karaca K, Ersöz G, Naycı Atış S, Kaya A. VAPdue to extensive drug-resistant Acinetobacter baumannii: risk factors, clinical features, and outcomes. Am J Infect Control. 2014;42(2):206–208. doi:10.1016/j.ajic.2013.09.003
89. Ju M, Hou D, Chen S, et al. Risk factors for mortality in ICU patients with Acinetobacter baumannii ventilator-associated pneumonia: impact of bacterial cytotoxicity. J Thorac Dis. 2018;10(5):2608–2617. doi:10.21037/jtd.2018.04.86
90. Pei Y, Huang Y, Pan X, et al. Nomogram for predicting 90-day mortality in patients with Acinetobacter baumannii-caused hospital-acquired and VAPin the respiratory intensive care unit. J Int Med Res. 2023;51(3):3000605231161481. doi:10.1177/03000605231161481
91. Lemos EV, de la Hoz FP, Einarson TR, et al. Carbapenem resistance and mortality in patients with Acinetobacter baumannii infection: systematic review and meta-analysis. Clin Microbiol Infect. 2014;20(5):416–423. doi:10.1111/1469-0691.12363
92. Mohd Sazlly Lim S. The global prevalence of multidrug-resistance among Acinetobacter baumannii causing hospital-acquired and VAPand its associated mortality: a systematic review and meta-analysis. J Infect. 2019;79(6):593–600. doi:10.1016/j.jinf.2019.09.012
93. Du X, Xu X, Yao J, et al. Predictors of mortality in patients infected with carbapenem-resistant Acinetobacter baumannii: a systematic review and meta-analysis. Am J Infect Control. 2019;47(9):1140–1145. doi:10.1016/j.ajic.2019.03.003
94. Adukauskiene D, Ciginskiene A, Adukauskaite A, Koulenti D, Rello J. Clinical features and outcomes of monobacterial and polybacterial episodes of VAPdue to multidrug-resistant Acinetobacter baumannii. Antibiotics. 2022;11(7):892. doi:10.3390/antibiotics11070892
95. Dexter C, Murray GL, Paulsen IT, Peleg AY. Community-acquired Acinetobacter baumannii: clinical characteristics, epidemiology and pathogenesis. Expert Rev Anti Infect Ther. 2015;13(5):567–573. doi:10.1586/14787210.2015.1025055
96. Dijkshoorn L, Nemec A, Seifert H. An increasing threat in hospitals: multidrug-resistant Acinetobacter baumannii. Nat Rev Microbiol. 2007;5(12):939–951. doi:10.1038/nrmicro1789
97. Villegas MV, Hartstein AI. Acinetobacter outbreaks, 1977-2000. Infect Control Hosp Epidemiol. 2003;24(4):284–295. doi:10.1086/502205
98. Monterrubio-Villar J, González-Velasco C, Valdezate-Ramos S, Córdoba-López A, Villalón-Panzano P, Saéz-Nieto JA. Outbreak of multiresistant Acinetobacter baumannii in a polyvalent intensive care unit: clinical, epidemiological analysis and PFGE-printing evolution. Eur J Clin Microbiol Infect Dis. 2009;28(10):1281–1284. doi:10.1007/s10096-009-0777-6
99. Araújo lima AV, da Silva SM. Occurrence and diversity of intra- and interhospital drug-resistant and biofilm-forming Acinetobacter baumannii and Pseudomonas aeruginosa. Microb Drug Resist. 2020;26(7):802–814. doi:10.1089/mdr.2019.0214
100. Schulte B, Goerke C, Weyrich P, et al. Clonal spread of meropenem-resistant Acinetobacter baumannii strains in hospitals in the Mediterranean region and transmission to South-west Germany. J Hosp Infect. 2005;61(4):356–357. doi:10.1016/j.jhin.2005.05.009
101. Meumann EM, Anstey NM, Currie BJ, et al. Genomic epidemiology of severe community-onset Acinetobacter baumannii infection. Microb Genom. 2019;5(3):e000258. doi:10.1099/mgen.0.000258
102. Jia H, Sun Q, Ruan Z, Xie X. Characterization of a small plasmid carrying the carbapenem resistance gene blaOXA-72 from community-acquired Acinetobacter baumannii sequence type 880 in China. Infect Drug Resist. 2019;12:1545–1553. doi:10.2147/IDR.S202803
103. Sharma AK, Dhasmana N, Dubey N, et al. Bacterial virulence factors: secreted for survival. Indian J Microbiol. 2017;57(1):1–10. doi:10.1007/s12088-016-0625-1
104. Kumar S, Anwer R, Azzi A. Virulence potential and treatment options of multidrug-resistant (MDR) Acinetobacter baumannii. Microorganisms. 2021;9(10):2104. doi:10.3390/microorganisms9102104
105. Lee CR, Lee JH, Park M, et al. Biology of Acinetobacter baumannii: pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front Cell Infect Microbiol. 2017;13(7):55. doi:10.3389/fcimb.2017.00055
106. Ayoub Moubareck C, Hammoudi Halat D. Insights into Acinetobacter baumannii: a review of microbiological, virulence, and resistance traits in a threatening nosocomial pathogen. Antibiotics. 2020;9(3):119. doi:10.3390/antibiotics9030119
107. McConnell MJ, Actis L, Pachón J. Acinetobacter baumannii: human infections, factors contributing to pathogenesis and animal models. FEMS Microbiol Rev. 2013;37(2):130–155. doi:10.1111/j.1574-6976.2012.00344.x
108. Smith SG, Mahon V, Lambert MA, Fagan RP. A molecular Swiss army knife: ompA structure, function and expression. FEMS Microbiol Lett. 2007;273(1):1–11. doi:10.1111/j.1574-6968.2007.00778.x
109. Smani Y, Dominguez-Herrera J, Pachón J. Association of the outer membrane protein Omp33 with fitness and virulence of Acinetobacter baumannii. J Infect Dis. 2013;208(10):1561–1570. doi:10.1093/infdis/jit386
110. Rumbo C, Tomás M, Fernández Moreira E, et al. The Acinetobacter baumannii Omp33-36 porin is a virulence factor that induces apoptosis and modulates autophagy in human cells. Infect Immun. 2014;82(11):4666–4680. doi:10.1128/IAI.02034-14
111. Zhu LJ, Chen XY, Hou PF. Mutation of CarO participates in drug resistance in imipenem-resistant Acinetobacter baumannii. J Clin Lab Anal. 2019;33(8):e22976. doi:10.1002/jcla.22976
112. Sato Y, Unno Y, Kawakami S, Ubagai T, Ono Y. Virulence characteristics of Acinetobacter baumannii clinical isolates vary with the expression levels of omps. J Med Microbiol. 2017;66(2):203–212. doi:10.1099/jmm.0.000394
113. Gil-Marqués ML, Pachón J, Smani Y. iTRAQ-Based quantitative proteomic analysis of Acinetobacter baumannii under hypoxia and normoxia reveals the role of OmpW as a virulence factor. Microbiol Spectr. 2022;10(2):e0232821. doi:10.1128/spectrum.02328-21
114. Uppalapati SR, Sett A, Pathania R. The Outer membrane proteins OmpA, CarO, and OprD of Acinetobacter baumannii confer a two-pronged defense in facilitating its success as a potent human pathogen. Front Microbiol. 2020;11:589234. doi:10.3389/fmicb.2020.589234
115. Kulp A, Kuehn MJ. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu Rev Microbiol. 2010;64:163–184. doi:10.1146/annurev.micro.091208.073413
116. Jun SH, Lee JH, Kim BR, et al. Acinetobacter baumannii outer membrane vesicles elicit a potent innate immune response via membrane proteins. PLoS One. 2013;8(8):e71751. doi:10.1371/journal.pone.0071751
117. Nho JS, Jun SH, Oh MH, et al. Acinetobacter nosocomialis secretes outer membrane vesicles that induce epithelial cell death and host inflammatory responses. Microb Pathog. 2015;81:39–45. doi:10.1016/j.micpath.2015.03.012
118. Rumbo C, Fernández-Moreira E, Merino M, et al. Horizontal transfer of the OXA-24 carbapenemase gene via outer membrane vesicles: a new mechanism of dissemination of carbapenem resistance genes in Acinetobacter baumannii. Antimicrob Agents Chemother. 2011;55(7):3084–3090. doi:10.1128/AAC.00929-10
119. Huang W, Yao Y, Long Q, et al. Immunization against multidrug-resistant Acinetobacter baumannii effectively protects mice in both pneumonia and sepsis models. PLoS One. 2014;9(6):e100727. doi:10.1371/journal.pone.0100727
120. Dehbanipour R, Ghalavand Z. Acinetobacter baumannii: pathogenesis, virulence factors, novel therapeutic options and mechanisms of resistance to antimicrobial agents with emphasis on tigecycline. J Clin Pharm Ther. 2022;47(11):1875–1884. doi:10.1111/jcpt.13787
121. Knapp S, Wieland CW, Florquin S, et al. Differential roles of CD14 and toll-like receptors 4 and 2 in murine Acinetobacter pneumonia. Am J Respir Crit Care Med. 2006;173(1):122–129. doi:10.1164/rccm.200505-730OC
122. Rossi E, Longo F, Barbagallo M, et al. Glucose availability enhances lipopolysaccharide production and immunogenicity in the opportunistic pathogen Acinetobacter baumannii. Future Microbiol. 2016;11(3):335–349. doi:10.2217/fmb.15.153
123. Haseley SR, Pantophlet R, Brade L, Holst O, Brade H. Structural and serological characterisation of the O-antigenic polysaccharide of the lipopolysaccharide from Acinetobacter junii strain 65. Eur J Biochem. 1997;245(2):477–481. doi:10.1111/j.1432-1033.1997.t01-1-00477.x
124. Geisinger E, Isberg RR. Antibiotic modulation of capsular exopolysaccharide and virulence in Acinetobacter baumannii. PLoS Pathog. 2015;11(2):e1004691. doi:10.1371/journal.ppat.1004691
125. van der Meer-Janssen YP, van Galen J, Batenburg JJ, Helms JB. Lipids in host-pathogen interactions: pathogens exploit the complexity of the host cell lipidome. Prog Lipid Res. 2010;49(1):1–26. doi:10.1016/j.plipres.2009.07.003
126. Camarena L, Bruno V, Euskirchen G, Poggio S, Snyder M. Molecular mechanisms of ethanol-induced pathogenesis revealed by RNA-sequencing. PLoS Pathog. 2010;6(4):e1000834. doi:10.1371/journal.ppat.1000834
127. Flores-Díaz M, Monturiol-Gross L, Naylor C, Alape-Girón A, Flieger A. Bacterial sphingomyelinases and phospholipases as virulence factors. Microbiol Mol Biol Rev. 2016;80(3):597–628. doi:10.1128/MMBR.00082-15
128. Stahl J, Bergmann H, Göttig S, Ebersberger I, Averhoff B. Acinetobacter baumannii virulence is mediated by the concerted action of three phospholipases D. PLoS One. 2015;10(9):e0138360. doi:10.1371/journal.pone.0138360
129. Tomaras AP, Dorsey CW, Edelmann RE, Actis LA. Attachment to and biofilm formation on abiotic surfaces by Acinetobacter baumannii: involvement of a novel chaperone-usher pili assembly system. Microbiology. 2003;149(Pt 12):3473–3484. doi:10.1099/mic.0.26541-0
130. Özarslan TO, Sirmatel F, Karabörk SO, Düzcü SE, Astarci HM. Acinetobacter baumannii pneumonia increases surfactant proteins SP-A, SP-B, and SP-D levels, while decreasing SP-C level in bronchoalveolar lavage in rats. Microbes Infect. 2023;25:105064. doi:10.1016/j.micinf.2022.105064
131. Waack U, Warnock M, Yee A, et al. cpaa is a glycan-specific adamalysin-like protease secreted by Acinetobacter baumannii that inactivates coagulation factor XII. mBio. 2018;9(6):e01606–18. doi:10.1128/mBio.01606-18
132. Tilley D, Law R, Warren S, Samis JA, Kumar A. CpaA a novel protease from Acinetobacter baumannii clinical isolates deregulates blood coagulation. FEMS Microbiol Lett. 2014;356(1):53–61. doi:10.1111/1574-6968.12496
133. Law SKK, Tan HS. The role of quorum sensing, biofilm formation, and iron acquisition as key virulence mechanisms in Acinetobacter baumannii and the corresponding anti-virulence strategies. Microbiol Res. 2022;260:127032. doi:10.1016/j.micres.2022.127032
134. Antunes LC, Imperi F, Towner KJ, Visca P. Genome-assisted identification of putative iron-utilization genes in Acinetobacter baumannii and their distribution among a genotypically diverse collection of clinical isolates. Res Microbiol. 2011;162(3):279–284. doi:10.1016/j.resmic.2010.10.010
135. Miethke M, Marahiel MA. Siderophore-based iron acquisition and pathogen control. Microbiol Mol Biol Rev. 2007;71(3):413–451. doi:10.1128/MMBR.00012-07
136. Ferreira D, Seca AML. Targeting human pathogenic bacteria by siderophores: a proteomics review. J Proteomics. 2016;145:153–166. doi:10.1016/j.jprot.2016.04.006
137. Kim M, Kim DY, Song WY, et al. distinctive roles of two acinetobactin isomers in challenging host nutritional immunity. mBio. 2021;12(5):e0224821. doi:10.1128/mBio.02248-21
138. Penwell WF, Arivett BA, Actis LA. The Acinetobacter baumannii entA gene located outside the acinetobactin cluster is critical for siderophore production, iron acquisition and virulence. PLoS One. 2012;7(5):e36493. doi:10.1371/journal.pone.0036493
139. Conde-Pérez K, Vázquez-Ucha JC, Álvarez-Fraga L, et al. In-depth analysis of the role of the acinetobactin cluster in the virulence of Acinetobacter baumannii. Front Microbiol. 2021;12:752070. doi:10.3389/fmicb.2021.752070
140. Nairn BL, Lonergan ZR, Wang J, et al. The Response of Acinetobacter baumannii to Zinc Starvation. Cell Host Microbe. 2016;19(6):826–836. doi:10.1016/j.chom.2016.05.007
141. Repizo GD. Prevalence of Acinetobacter baumannii strains expressing the Type 6 secretion system in patients with bacteremia. Virulence. 2017;8(7):1099–1101. doi:10.1080/21505594.2017.1346768
142. Weber BS, Hennon SW, Wright MS, et al. Genetic dissection of the type vi secretion system in Acinetobacter and identification of a novel peptidoglycan hydrolase, TagX, required for its biogenesis. mBio. 2016;7(5):e01253–16. doi:10.1128/mBio.01253-16
143. Johnson TL, Waack U, Smith S, Mobley H, Sandkvist M. Acinetobacter baumannii Is dependent on the type ii secretion system and its substrate lipa for lipid utilization and in vivo fitness. J Bacteriol. 2015;198(4):711–719. doi:10.1128/JB.00622-15
144. Russo TA, Manohar A, Beanan JM, et al. The response regulator bfmr is a potential drug target for Acinetobacter baumannii. mSphere. 2016;1(3):e00082–16. doi:10.1128/mSphere.00082-16
145. Yang CH, Su PW, Moi SH, Chuang LY. Biofilm formation in Acinetobacter baumannii: genotype-phenotype correlation. Molecules. 2019;24(10):1849. doi:10.3390/molecules24101849
146. Flemming HC, van Hullebusch ED, Neu TR, et al. The biofilm matrix: multitasking in a shared space. Nat Rev Microbiol. 2023;21(2):70–86. doi:10.1038/s41579-022-00791-0
147. Greene C, Wu J, Rickard AH, Xi C. Evaluation of the ability of Acinetobacter baumannii to form biofilms on six different biomedical relevant surfaces. Lett Appl Microbiol. 2016t;63(4):233–239. doi:10.1111/lam.12627
148. Loehfelm TW, Luke NR, Campagnari AA. Identification and characterization of an Acinetobacter baumannii biofilm-associated protein. J Bacteriol. 2008;190(3):1036–1044. doi:10.1128/JB.01416-07
149. Choi AH, Slamti L, Avci FY, Pier GB, Maira-Litrán T. The pgaABCD locus of Acinetobacter baumannii encodes the production of poly-beta-1-6-N-acetylglucosamine, which is critical for biofilm formation. J Bacteriol. 2009;191(19):5953–5963. doi:10.1128/JB.00647-09
150. Treter J, Macedo AJ. Catheters: a suitable surface for biofilm formation. Sci Against Microbial Pathogens. 2011;2(3):835–842.
151. Li H, Song C, Liu D, Ai Q, Yu J. Molecular analysis of biofilms on the surface of neonatal endotracheal tubes based on 16S rRNA PCR-DGGE and species-specific PCR. Int J Clin Exp Med. 2015;8(7):11075–11084.
152. Bardes JM, Waters C, Motlagh H, Wilson A. The Prevalence of oral flora in the biofilm microbiota of the endotracheal tube. Am Surg. 2016;82(5):403–406.
153. Ferreira Tde O, Koto RY. Microbial investigation of biofilms recovered from endotracheal tubes using sonication in intensive care unit pediatric patients. Braz J Infect Dis. 2016t;20(5):468–475. doi:10.1016/j.bjid.2016.07.003
154. Vandecandelaere I, Matthijs N, Nelis HJ, Depuydt P, Coenye T. The presence of antibiotic-resistant nosocomial pathogens in endotracheal tube biofilms and corresponding surveillance cultures. Pathog Dis. 2013;69(2):142–148. doi:10.1111/2049-632X.12100
155. Perkins SD, Woeltje KF, Angenent LT. Endotracheal tube biofilm inoculation of oral flora and subsequent colonization of opportunistic pathogens. Int J Med Microbiol. 2010;300(7):503–511. doi:10.1016/j.ijmm.2010.02.005
156. Coppadoro A, Bellani G. Non-Pharmacological Interventions to Prevent Ventilator-Associated Pneumonia: a Literature Review. Respir Care. 2019;64(12):1586–1595. doi:10.4187/respcare.07127
157. Sottile FD, Marrie TJ, Prough DS, et al. Nosocomial pulmonary infection: possible etiologic significance of bacterial adhesion to endotracheal tubes. Crit Care Med. 1986;14(4):265–270.
158. Jarrett WA, Ribes J, Manaligod JM. (2002) Biofilm formation on tracheostomy tubes. Ear Nose Throat J. 2002;81(9):659–661. doi:10.1177/014556130208100915
159. M’hamedi I, Hassaine H, Bellifa S, Lachachi M, Terki IK, Djeribi R. Biofilm formation by Acinetobacter baumannii isolated from medical devices at the intensive care unit of the University Hospital of Tlemcen (Algeria). African J Microbio Res. 2014;8(3):270–276. doi:10.5897/AJMR2013.6288
160. Dargahi Z, Hamad AA, Sheikh AF, et al. The biofilm formation and antibiotic resistance of bacterial profile from endotracheal tube of patients admitted to intensive care unit in southwest of Iran. PLoS One. 2022;17(11):e0277329. doi:10.1371/journal.pone.0277329
161. Raveendra N, Rathnakara SH, Haswani N, Subramaniam V. Bacterial biofilms on tracheostomy tubes. Indian J Otolaryngol Head Neck Surg. 2022;74:4995–4999. doi:10.1007/s12070-021-02598-6
162. Diaconu O, Siriopol I, Poloșanu LI, Grigoraș I. Endotracheal tube biofilm and its impact on the pathogenesis of ventilator-associated pneumonia. J Crit Care Med. 2018;4(2):50–55. doi:10.2478/jccm-2018-0011
163. Cairns S, Thomas JG, Hooper SJ, et al. Molecular analysis of microbial communities in endotracheal tube biofilms. PLoS One. 2011;6(3):e14759. doi:10.1371/journal.pone.0014759
164. Baidya S, Sharma S, Mishra SK, Kattel HP, Parajuli K, Sherchand JB. Biofilm formation by pathogens causing VAPat intensive care units in a tertiary care hospital: an armor for refuge. Biomed Res Int. 2021;2021:8817700. doi:10.1155/2021/8817700
165. Khoshnood S, Sadeghifard N, Mahdian N, et al. Antimicrobial resistance and biofilm formation capacity among Acinetobacter baumannii strains isolated from patients with burns and ventilator-associated pneumonia. J Clin Lab Anal. 2023;37(1):e24814. doi:10.1002/jcla.24814
166. Whiteley M, Diggle SP, Greenberg EP. Progress in and promise of bacterial quorum sensing research. Nature. 2017;551(7680):313–320. doi:10.1038/nature24624
167. Rémy B, Mion S, Plener L, Elias M, Chabrière E, Daudé D. Interference in bacterial quorum sensing: a biopharmaceutical perspective. Front Pharmacol. 2018;9:203. doi:10.3389/fphar.2018.00203
168. Saipriya K, Swathi CH, Ratnakar KS, Sritharan V. Quorum-sensing system in Acinetobacter baumannii: a potential target for new drug development. J Appl Microbiol. 2020;128(1):15–27. doi:10.1111/jam.14330
169. Xiong L, Yi F, Yu Q, et al. Transcriptomic analysis reveals the regulatory role of quorum sensing in the Acinetobacter baumannii ATCC 19606 via RNA-seq. BMC Microbiol. 2022;22(1):198. doi:10.1186/s12866-022-02612-z
170. Sun X, Ni Z, Tang J, Ding Y, Wang X, Li F. The abaI/abaR quorum sensing system effects on pathogenicity in Acinetobacter baumannii. Front Microbiol. 2021;12:679241. doi:10.3389/fmicb.2021.679241
171. Abraham WR. Going beyond the Control of Quorum-Sensing to Combat Biofilm Infections. Antibiotics. 2016;5(1):3. doi:10.3390/antibiotics5010003
172. Yoshimura J, Kinoshita T, Yamakawa K, et al. Impact of Gram stain results on initial treatment selection in patients with ventilator-associated pneumonia: a retrospective analysis of two treatment algorithms. Crit Care. 2017;21(1):156. doi:10.1186/s13054-017-1747-5
173. Khan R, Al-Dorzi HM, Tamim HM, et al. The impact of onset time on the isolated pathogens and outcomes in ventilator associated pneumonia. J Infect Public Health. 2016;9(2):161–171. doi:10.1016/j.jiph.2015.09.002
174. Craven DE, Hudcova J, Lei Y. Diagnosis of ventilator-associated respiratory infections (VARI): microbiologic clues for tracheobronchitis (VAT) and pneumonia (VAP). Clin Chest Med. 2011;32(3):547–557. doi:10.1016/j.ccm.2011.06.001
175. Nair GB, Niederman MS. Ventilator-associated pneumonia: present understanding and ongoing debates. Intensive Care Med. 2015;41(1):34–48. doi:10.1007/s00134-014-3564-5
176. Dudeck MA, Horan TC, Peterson KD, et al. National Healthcare Safety Network (NHSN) Report, data summary for 2010, device-associated module. Am J Infect Control. 2011;39(10):798–816. doi:10.1016/j.ajic.2011.10.001
177. Metersky ML, Wang Y, Klompas M, Eckenrode S, Bakullari A, Eldridge N. trend in VAPrates between 2005 and 2013. JAMA. 2016;316(22):2427–2429. doi:10.1001/jama.2016.16226
178. Kalanuria AA, Ziai W, Mirski M. VAPin the ICU. Crit Care. 2014;18(2):208. doi:10.1186/cc13775
179. ANVISA. Agência Nacional de Vigilância Sanitária. 2013. Critérios diagnósticos de infecção relacionada à assistência à saúde.4-150.
180. Prates CG, Martins AF, Superti SV, et al. Risk factors for 30-day mortality in patients with carbapenem-resistant Acinetobacter baumannii during an outbreak in an intensive care unit. Epidemiol Infect. 2011;139(3):411–418. doi:10.1017/S0950268810001238
181. Zahar JR, Timsit JF, Garrouste-Orgeas M, et al. Outcomes in severe sepsis and patients with septic shock: pathogen species and infection sites are not associated with mortality. Crit Care Med. 2011;39(8):1886–1895. doi:10.1097/CCM.0b013e31821b827c
182. Tabah A, Koulenti D, Laupland K, et al. Characteristics and determinants of outcome of hospital-acquired bloodstream infections in intensive care units: the EUROBACT International Cohort Study. Intensive Care Med. 2012;38(12):1930–1945. doi:10.1007/s00134-012-2695-9
183. Sarda C, Fazal F, Rello J. Management of VAP(VAP) caused by resistant gram-negative bacteria: which is the best strategy to treat? Expert Rev Respir Med. 2019;13(8):787–798. doi:10.1080/17476348.2019.1632195
184. Reignier J, Darmon M, Sonneville R, et al. Impact of early nutrition and feeding route on outcomes of mechanically ventilated patients with shock: a post hoc marginal structural model study. Intensive Care Med. 2015;41(5):875–886. doi:10.1007/s00134-015-3730-4
185. Fitch ZW, Whitman GJ. Incidence, risk, and prevention of VAPin adult cardiac surgical patients: a systematic review. J Card Surg. 2014;29(2):196–203. doi:10.1111/jocs.12260
186. Schwebel C. Safety of intrahospital transport in ventilated critically ill patients: a multicenter cohort study. Crit Care Med. 2013;41(8):1919–1928. doi:10.1097/CCM.0b013e31828a3bbd
187. Bornstain C, Azoulay E, De Lassence A, et al. Sedation, sucralfate, and antibiotic use are potential means for protection against early-onset ventilator-associated pneumonia. Clin Infect Dis. 2004;38(10):1401–1408. doi:10.1086/386321
188. Kadri SS, Adjemian J, Lai YL, et al. Difficult-to-treat resistance in gram-negative bacteremia at 173 us hospitals: retrospective cohort analysis of prevalence, predictors, and outcome of resistance to all first-line agents. Clin Infect Dis. 2018;67(12):1803–1814. doi:10.1093/cid/ciy378
189. Vazquez Guillamet C, Kollef MH. Acinetobacter pneumonia: improving outcomes with early identification and appropriate therapy. Clin Infect Dis. 2018;67(9):1455–1462. doi:10.1093/cid/ciy375
190. Garnacho-Montero J, Ortiz-Leyba C, Jiménez-Jiménez FJ, et al. Treatment of multidrug-resistant Acinetobacter baumannii VAP(VAP) with intravenous colistin: a comparison with imipenem-susceptible VAP. Clin Infect Dis. 2003;36(9):1111–1118. doi:10.1086/374337
191. Superti SV, Martins Dde S, Caierão J, Soares Fda S, Prochnow T, Zavascki AP. Indications of carbapenem resistance evolution through heteroresistance as an intermediate stage in Acinetobacter baumannii after carbapenem administration. Rev Inst Med Trop Sao Paulo. 2009;51(2):111–113. doi:10.1590/s0036-46652009000200010
192. Fishbain J, Peleg AY. Treatment of Acinetobacter infections. Clin Infect Dis. 2010;51(1):79–84. doi:10.1086/653120
193. Levin AS, Levy CE, Manrique AE, Medeiros EA, Costa SF. Severe nosocomial infections with imipenem-resistant Acinetobacter baumannii treated with ampicillin/sulbactam. Int J Antimicrob Agents. 2003;21(1):58–62. doi:10.1016/s0924-8579(02)00276-5
194. Wood GC, Hanes SD, Croce MA, Fabian TC, Boucher BA. Comparison of ampicillin-sulbactam and imipenem-cilastatin for the treatment of Acinetobacter ventilator-associated pneumonia. Clin Infect Dis. 2002;34(11):1425–1430. doi:10.1086/340055
195. Chu H, Zhao L, Wang M, Liu Y, Gui T, Zhang J. Sulbactam-based therapy for Acinetobacter baumannii infection: a systematic review and meta-analysis. Braz J Infect Dis. 2013;17(4):389–394. doi:10.1016/j.bjid.2012.10.029
196. Penwell WF, Shapiro AB, Giacobbe RA, et al. Molecular mechanisms of sulbactam antibacterial activity and resistance determinants in Acinetobacter baumannii. Antimicrob Agents Chemother. 2015;59(3):1680–1689. doi:10.1128/AAC.04808-14
197. Ritchie DJ, Garavaglia-Wilson A. A review of intravenous minocycline for treatment of multidrug-resistant Acinetobacter infections. Clin Infect Dis. 2014;59 Suppl 6:S374–80. doi:10.1093/cid/ciu613
198. Lee YT, Tsao SM, Hsueh PR. Clinical outcomes of tigecycline alone or in combination with other antimicrobial agents for the treatment of patients with healthcare-associated multidrug-resistant Acinetobacter baumannii infections. Eur J Clin Microbiol Infect Dis. 2013;32(9):1211–1220. doi:10.1007/s10096-013-1870-4
199. Freire AT, Melnyk V, Kim MJ, et al. Comparison of tigecycline with imipenem/cilastatin for the treatment of hospital-acquired pneumonia. Diagn Microbiol Infect Dis. 2010;68(2):140–151. doi:10.1016/j.diagmicrobio.2010.05.012
200. Rando E, Cutuli SL, Sangiorgi F, et al. Cefiderocol-containing regimens for the treatment of carbapenem-resistant A. baumannii ventilator-associated pneumonia: a propensity-weighted cohort study. JAC Antimicrob Resist. 2023;5(4):dlad085. doi:10.1093/jacamr/dlad085
201. Corcione S, De Benedetto I, Pinna SM, et al. Cefiderocol use in Gram negative infections with limited therapeutic options: is combination therapy the key? J Infect Public Health. 2022;15(9):975–979. doi:10.1016/j.jiph.2022.07.006
202. Alp E, Eren E, Elay G, Cevahir F, Esmaoğlu A, Rello J. Efficacy of loading dose of colistin in Acinetobacter baumannii ventilator-associated pneumonia. Infez Med. 2017;25(4):311–319.
203. Wang J, Niu H, Wang R, Cai Y. Safety and efficacy of colistin alone or in combination in adults with Acinetobacter baumannii infection: a systematic review and meta-analysis. Int J Antimicrob Agents. 2019;53(4):383–400. doi:10.1016/j.ijantimicag.2018.10.020
204. Khawcharoenporn T, Pruetpongpun N, Tiamsak P, Rutchanawech S, Mundy LM, Apisarnthanarak A. Colistin-based treatment for extensively drug-resistant Acinetobacter baumannii pneumonia. Int J Antimicrob Agents. 2014;43(4):378–382. doi:10.1016/j.ijantimicag.2014.01.016
205. Biagi M, Butler D, Tan X, Qasmieh S, Wenzler E. A breath of fresh air in the fog of antimicrobial resistance: inhaled polymyxins for gram-negative pneumonia. Antibiotics. 2019;8(1):27. doi:10.3390/antibiotics8010027
206. Moghaddam O, Niakan Lahiji M, Talebi-Taher M, Mahmoodiyeh B. effect of inhaled colistin on the treatment of VAPdue to multi-drug resistant Acinetobacter. Tanaffos. 2019;18(1):66–73.
207. Valachis A, Samonis G, Kofteridis DP. The role of aerosolized colistin in the treatment of ventilator-associated pneumonia: a systematic review and metaanalysis. Crit Care Med. 2015;43(3):527–533. doi:10.1097/CCM.0000000000000771
208. Solé-Lleonart C, Rouby JJ, Blot S, et al. Nebulization of Antiinfective Agents in Invasively Mechanically Ventilated Adults: a Systematic Review and Meta-analysis. Anesthesiology. 2017;126(5):890–908. doi:10.1097/ALN.0000000000001570
209. Kisil OV, Efimenko TA, Gabrielyan NI, et al. Development of antimicrobial therapy methods to overcome the antibiotic resistance of Acinetobacter baumannii. Acta Naturae. 2020;12(3):34–45. doi:10.32607/actanaturae.10955
210. Comeau AM, Hatfull GF, Krisch HM, et al. Exploring the prokaryotic virosphere. Res Microbiol. 2008;159(5):306–313. doi:10.1016/j.resmic.2008.05.001
211. Bagińska N, Pichlak A, Górski A, et al. Specific and selective bacteriophages in the fight against multidrug-resistant Acinetobacter baumannii. Virol Sin. 2019;34(4):347–357. doi:10.1007/s12250-019-00125-0
212. Jung CJ, Liao YD, Hsu CC, et al. Identification of potential therapeutic antimicrobial peptides against Acinetobacter baumannii in a mouse model of pneumonia. Sci Rep. 2021;11(1):7318. doi:10.1038/s41598-021-86844-5
213. Losasso C, Belluco S, Cibin V, et al. Antibacterial activity of silver nanoparticles: sensitivity of different Salmonella serovars. Front Microbiol. 2014;26(5):227. doi:10.3389/fmicb.2014.00227
214. Intorasoot A, Chornchoem P, Sookkhee S, et al. Bactericidal activity of herbal volatile oil extracts against multidrug-resistant Acinetobacter baumannii. J Intercult Ethnopharmacol. 2017;6(2):218–222. doi:10.5455/jice.20170411091159
215. Meng L, Wang C, Li J, Zhang J. Early vs late tracheostomy in critically ill patients: a systematic review and meta-analysis. Clin Respir J. 2016;10(6):684–692. doi:10.1111/crj.12286
216. Klompas M, Branson R, Eichenwald EC, et al. Strategies to prevent VAPin acute care hospitals: 2014 update. Infect Control Hosp Epidemiol. 2014;35:S133–54. doi:10.1017/s0899823x00193894
217. Rello J, Lode H, Cornaglia G, Masterton R, VAP Care Bundle Contributors. A European care bundle for prevention of ventilator-associated pneumonia. Intensive Care Med. 2010;36(5):773–780. doi:10.1007/s00134-010-1841-5
218. Speck K, Rawat N, Weiner NC, Tujuba HG, Farley D, Berenholtz S. A systematic approach for developing a VAPprevention bundle. Am J Infect Control. 2016;44(6):652–656. doi:10.1016/j.ajic.2015.12.020
219. Roquilly A, Marret E, Abraham E, Asehnoune K. Pneumonia prevention to decrease mortality in intensive care unit: a systematic review and meta-analysis. Clin Infect Dis. 2015;60(1):64–75. doi:10.1093/cid/ciu740
220. de Smet AM, Kluytmans JA, Cooper BS, et al. Decontamination of the digestive tract and oropharynx in ICU patients. N Engl J Med. 2009;360(1):20–31. doi:10.1056/NEJMoa0800394
221. Klompas M. Oropharyngeal Decontamination with Antiseptics to Prevent Ventilator-Associated Pneumonia: rethinking the Benefits of Chlorhexidine. Semin Respir Crit Care Med. 2017;38(3):381–390. doi:10.1055/s-0037-1602584
222. LPSN.dsmz.de; 2023. Available from:https://classic.clinicaltrials.gov/ct2/show/.
223. Aloush SM. Does educating nurses with VAPprevention guidelines improve their compliance? Am J Infect Control. 2017;45(9):969–973. doi:10.1016/j.ajic.2017.04.009
224. Jansson M, Ala-Kokko T, Ylipalosaari P, Syrjälä H, Kyngäs H. Critical care nurses’ knowledge of, adherence to and barriers towards evidence-based guidelines for the prevention of ventilator-associated pneumonia--a survey study. Intensive Crit Care Nurs. 2013;29(4):216–227. doi:10.1016/j.iccn.2013.02.006
225. Madhuvu A, Endacott R, Plummer V, Morphet J. Nurses’ knowledge, experience and self-reported adherence to evidence-based guidelines for prevention of ventilator-associated events: a national online survey. Intensive Crit Care Nurs. 2020;59:102827. doi:10.1016/j.iccn.2020.102827
226. Madhuvu A, Endacott R, Plummer V, Morphet J. Healthcare professional views on barriers to implementation of evidence-based practice in prevention of ventilator-associated events: a qualitative descriptive study. Intensive Crit Care Nurs. 2022;68:103133. doi:10.1016/j.iccn.2021.103133
227. Jam R, Mesquida J, Hernández Ó, et al. Nursing workload and compliance with non-pharmacological measures to prevent ventilator-associated pneumonia: a multicentre study. Nurs Crit Care. 2018;23(6):291–298. doi:10.1111/nicc.12380
228. Bouadma L, Mourvillier B, Deiler V, et al. Changes in knowledge, beliefs, and perceptions throughout a multifaceted behavioral program aimed at preventing ventilator-associated pneumonia. Intensive Care Med. 2010;36(8):1341–1347. doi:10.1007/s00134-010-1890-9
229. Wiersinga WJ, Rhodes A, Cheng AC, Peacock SJ, Prescott HC. Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19): a Review. JAMA. 2020;324(8):782–793. doi:10.1001/jama.2020.12839
230. Maes M, Higginson E, Pereira-Dias J, et al. VAPin critically ill patients with COVID-19. Crit Care. 2021;25(1):25. doi:10.1186/s13054-021-03460-5
231. François B, Laterre PF, Luyt CE, Chastre J. The challenge of VAPdiagnosis in COVID-19 patients. Crit Care. 2020;24(1):289. doi:10.1186/s13054-020-03013-2
232. Jain S, Khanna P, Sarkar S. Comparative evaluation of VAPin critically ill COVID- 19 and patients infected with other Corona viruses: a systematic review and meta-analysis. Monaldi Arch Chest Dis. 2021;92:2. doi:10.4081/monaldi.2021.1610
233. Ippolito M, Misseri G, Catalisano G, et al. VAPin patients with covid-19: a systematic review and meta-analysis. Antibiotics. 2021;10(5):545. doi:10.3390/antibiotics10050545
234. Gamberini L, Tonetti T, Spadaro S, et al. Factors influencing liberation from mechanical ventilation in coronavirus disease 2019: multicenter observational study in fifteen Italian ICUs. J Intensive Care. 2020;8:80. doi:10.1186/s40560-020-00499-4
235. Nseir S, Martin-Loeches I, Povoa P, et al. Relationship between VAPand mortality in COVID-19 patients: a planned ancillary analysis of the coVAPid cohort. Crit Care. 2021;25(1):177. doi:10.1186/s13054-021-03588-4
236. Meawed TE, Ahmed SM, Mowafy SMS, Samir GM, Anis RH. Bacterial and fungal ventilator associated pneumonia in critically ill COVID-19 patients during the second wave. J Infect Public Health. 2021;14(10):1375–1380. doi:10.1016/j.jiph.2021.08.003
237. Pascale R, Bussini L, Gaibani P, et al. Carbapenem-resistant bacteria in an intensive care unit during the coronavirus disease 2019 (COVID-19) pandemic: a multicenter before-and-after cross-sectional study. Infect Control Hosp Epidemiol. 2022;43(4):461–466. doi:10.1017/ice.2021.144
238. Gottesman T, Fedorowsky R, Yerushalmi R, Lellouche J, Nutman A. An outbreak of carbapenem-resistant Acinetobacter baumannii in a COVID-19 dedicated hospital. Infect Prev Pract. 2021;3(1):100113. doi:10.1016/j.infpip.2021.100113
239. Shinohara DR. Outbreak of endemic carbapenem-resistant Acinetobacter baumannii in a coronavirus disease 2019 (COVID-19)-specific intensive care unit. Infect Control Hosp Epidemiol. 2022;43(6):815–817. doi:10.1017/ice.2021.98
240. Rouyer M, Strazzulla A, Youbong T, et al. VAPin COVID-19 Patients: a Retrospective Cohort Study. Antibiotics. 2021;10(8):988. doi:10.3390/antibiotics10080988
241. Zand F, Vakili H, Asmarian N, et al. Unintended impact of COVID-19 pandemic on the rate of catheter related nosocomial infections and incidence of multiple drug resistance pathogens in three intensive care units not allocated to COVID-19 patients in a large teaching hospital. BMC Infect Dis. 2023;23(1):11. doi:10.1186/s12879-022-07962-7
242. Giacobbe DR, Battaglini D, Enrile EM, et al. Incidence and prognosis of VAPin critically ill patients with COVID-19: a multicenter study. J Clin Med. 2021;10(4):555. doi:10.3390/jcm10040555
243. Gregorczyk-Maga I, Pałka A, Fiema M, et al. Correction: impact of tooth brushing on oral bacteriota and health care-associated infections among ventilated COVID-19 patients: an intervention study. Antimicrob Resist Infect Control. 2023;12(1):51. doi:10.1186/s13756-023-01256-6
244. Puzniak L, Bauer KA, Yu KC, et al. Effect of inadequate empiric antibacterial therapy on hospital outcomes in SARS-CoV-2-positive and -negative us patients with a positive bacterial culture: a multicenter evaluation from march to November 2020. Open Forum Infect Dis. 2021;8(6):ofab232. doi:10.1093/ofid/ofab232
245. Jeffreys S, Chambers JP, Yu JJ, Hung CY, Forsthuber T, Arulanandam BP. Insights into Acinetobacter baumannii protective immunity. Front Immunol. 2022;13:1070424. doi:10.3389/fimmu.2022.1070424
246. Cox MJ, Loman N, Bogaert D, O’Grady J. Co-infections: potentially lethal and unexplored in COVID-19. Lancet Microbe. 2020;1(1):e11. doi:10.1016/S2666-5247(20)30009-4
247. Cureño-Díaz MA, Durán-Manuel EM, Cruz-Cruz C, et al. Impact of the modification of a cleaning and disinfection method of mechanical ventilators of COVID-19 patients and ventilator-associated pneumonia: one year of experience. Am J Infect Control. 2021;49(12):1474–1480. doi:10.1016/j.ajic.2021.09.012
248. Perez S, Innes GK, Walters MS, et al. Increase in hospital-acquired carbapenem-resistant Acinetobacter baumannii infection and colonization in an acute care hospital during a surge in COVID-19 admissions - New Jersey, February-July 2020. MMWR Morb Mortal Wkly Rep. 2020;69(48):1827–1831. doi:10.15585/mmwr.mm6948e1
249. Vijay S, Bansal N, Rao BK, et al. Secondary infections in hospitalized COVID-19 patients: Indian experience. Infect Drug Resist. 2021;14:1893–1903. doi:10.2147/IDR.S299774
250. Loyola-Cruz MÁ, Durán-Manuel EM, Cruz-Cruz C, et al. ESKAPE bacteria characterization reveals the presence of Acinetobacter baumannii and Pseudomonas aeruginosa outbreaks in COVID-19/VAP patients. Am J Infect Control. 2023;51(7):729–737. doi:10.1016/j.ajic.2022.08.012
251. Ukuhor HO. The interrelationships between antimicrobial resistance, COVID-19, past, and future pandemics. J Infect Public Health. 2021;14(1):53–60. doi:10.1016/j.jiph.2020.10.018
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Old Age is an Independent Risk Factor for Pneumonia Development in Patients with SARS-CoV-2 Omicron Variant Infection and a History of Inactivated Vaccine Injection
Tong X, Huang Z, Zhang X, Si G, Lu H, Zhang W, Xue Y, Xie W
Infection and Drug Resistance 2022, 15:5567-5573
Published Date: 21 September 2022
Platelet-to-White Blood Cell Ratio as a Predictor of Mortality in Patients with Severe COVID-19 Pneumonia: A Retrospective Cohort Study
Thungthienthong M, Vattanavanit V
Infection and Drug Resistance 2023, 16:445-455
Published Date: 24 January 2023
Value of Laboratory Indicators in Predicting Pneumonia in Symptomatic COVID-19 Patients Infected with the SARS-CoV-2 Omicron Variant
Zhu K, Ma S, Chen H, Xie J, Huang D, Fu C, Ma G, Huang Y
Infection and Drug Resistance 2023, 16:1159-1170
Published Date: 28 February 2023
Impact of Enhanced in-Hospital Infection Prevention During the COVID-19 Pandemic on Postoperative Pneumonia in Older Surgical Patients
Ju JW, You J, Hong H, Kang CK, Kim WH, Lee HJ
International Journal of General Medicine 2023, 16:1943-1951
Published Date: 23 May 2023
A Case of Severe Rhabdomyolysis, Acute Myocardial Damage and Multi-Organ Dysfunction Syndrome in a Patient with Novel Coronavirus Pneumonia
Yuan S, Huang Y, Xie P, Li P
Open Access Emergency Medicine 2024, 16:19-28
Published Date: 1 February 2024