Back to Journals » Cancer Management and Research » Volume 12
TOP2A Promotes Cell Migration, Invasion and Epithelial–Mesenchymal Transition in Cervical Cancer via Activating the PI3K/AKT Signaling
Authors Wang B, Shen Y, Zou Y, Qi Z, Huang G, Xia S, Gao R, Li F, Huang Z ![]()
Received 1 December 2019
Accepted for publication 22 April 2020
Published 21 May 2020 Volume 2020:12 Pages 3807—3814
DOI https://doi.org/10.2147/CMAR.S240577
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Eileen O'Reilly
Bi Wang,1,2,* Yaping Shen,3,* Yin Zou,4 Zhengjun Qi,4 Guijia Huang,4 Shan Xia,5 Rui Gao,6 Fenghu Li,5 Zhi Huang7
1Key Laboratory of Endemic and Ethnic Diseases, Ministry of Education, Guizhou Medical University, Guiyang, Guizhou, People’s Republic of China; 2Department of Paediatrics, Maternal and Child Health Hospital of Guiyang City, Guiyang, Guizhou, People’s Republic of China; 3Department of Interventional Radiology, The Affiliated Baiyun Hospital of Guizhou Medical University, Guiyang, Guizhou, People’s Republic of China; 4Department of Oncology, The Second Affiliated Hospital of Guizhou Medical University, Guiyang, Guizhou, People’s Republic of China; 5Department of Gynecologic Oncology, The Affiliated Cancer Hospital of Guizhou Medical University, Guiyang, Guizhou, People’s Republic of China; 6Guiyang Customs Guizhou International Travel Healthcare Center, Guiyang, Guizhou, People’s Republic of China; 7Department of Interventional Radiology, The Affiliated Hospital of Guizhou Medical University, Guiyang, Guizhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Fenghu Li
Department of Gynecologic Oncology, The Affiliated Cancer Hospital of Guizhou Medical University, Guiyang City, Guizhou Province, People’s Republic of China
Email [email protected]
Zhi Huang
Department of Interventional Radiology, The Affiliated Hospital of Guizhou Medical University, Guiyang City, Guizhou Province, People’s Republic of China
Email [email protected]
Background/Objective: Topoisomerases type IIA (TOP2A) was identified to present with a high-expression pattern in cervical cancer. However, TOP2A role in the progression of cervical cancer remains unknown. Here, we aimed to explore the effect and reveal the underlying mechanism of TOP2A in the migration, invasion and epithelial–mesenchymal transition (EMT) of cervical cancer.
Materials and Methods: The expression profiles of TOP2A in 20 paired cervical cancer tissues and the paracancerous normal tissues were detected by using Western blotting assay. Transwell chambers were used to test cell migration and invasion abilities. Cell morphology and the expressions of E-cadherin and N-cadherin were detected to assess cell EMT. LY294002 was used to inhibit the activation of PI3K/AKT signaling.
Results: Compared with the paracancerous normal tissues, TOP2A was overexpressed in 85% (17/20) cervical cancer tissues. Repression of TOP2A expression in SiHa cells significantly weakened cell migration and invasion abilities, reduced cell numbers in shuttle shape and increased E-cadherin expression while decreased E-cadherin expression. To the opposite, overexpression of TOP2A in Hela cells induced opposite results. In addition, the expression of p-AKT was increased when TOP2A was overexpressed in Hela cells, and p-AKT expression was decreased when TOP2A was silenced in SiHa cells. Moreover, suppression of the PI3K/AKT signaling with LY294002 treatment apparently rescued TOP2A-mediated promotions in cell migration, invasion and EMT in Hela cells.
Conclusion: This study reveals that TOP2A is abnormally overexpressed in cervical cancer tissues, and TOP2A overexpression leads to cell migration, invasion and EMT via activating PI3K/AKT signaling.
Keywords: topoisomerases type IIA, epithelial–mesenchymal transition, cell morphology, migration, PI3K/AKT signaling
Introduction
Cervical cancer ranks the second common cancer in developing countries and is the third most common reason for cancer-related deaths in women, with 570,000 new cases and 311,000 deaths per year worldwide.1 The persistent infection of human papillomavirus (HPV) is the major pathogenesis responsible for cervical cancer, and HPV16 and HPV18 accounts for about 73% of all the cervical cancer cases among more than100 HPV genotypes.2,3 Although improvements have been achieved in the diagnostic and therapeutical techniques, majority patients with cervical cancer showed metastasis present with high incidence in cervical cancer, which is one of the main reasons for the mortality in patients with cervical cancer.4 In consequence, it is necessary to explore the molecular mechanisms underlying the metastasis of cervical cancer.
Epithelial–mesenchymal transition (EMT) is a biological process involving the polarization of epithelial cells that enable cells to acquire mesenchymal cell phenotypes.5 Importantly, EMT confers cancer cell several properties, such as the enhanced migratory and invasive capacities and the increased resistance to apoptosis.6 EMT has been verified to play an important role in the metastasis and invasion of majority kinds of cancers, including cervical cancer,7,8 indicating a vital value of EMT possesses in cancer treatment.
Topoisomerases type IIA (TOP2A), as one of the two members of topoisomerase II (TOP2) family, with another is TOP2B, is expressed only in cycling cells and responsible for enzymatic uncoupling during the replication of DNA strands.9 TOP2A catalyzes double-strand DNA breaking and induces gene transcription during mitosis.10,11 Noticeably, TOP2A has been reported to be overexpressed in colon cancer tissues when compared with the adjacent normal tissues, and TOP2A depletion induced significant decreases in cell proliferation and invasion abilities and promoted cell apoptosis in both colon cancer HCT116 and SW480 cells.12 In cervical cancer, TOP2A was shown to overexpress during the transformation of cervical intraepithelial neoplasia 2 (CIN2)-CIN3 to cervical cancer.13,14 However, the role and mechanisms of TOP2A in the development and progression of cervical cancer still remain unknown.
The PI3K/AKT signaling is an important intracellular signaling pathway which can promote migration and induce EMT in many types of tumors including cervical.15 Wang et al16 reported that TOP2A was associated with the activation of PI3K/AKT and Wnt/β‑catenin signaling pathways in testicular seminoma. Therefore, we speculated that TOP2A might play a role in the migration, invasion and EMT of cervical cancer via regulating the activation of PI3K/AKT signaling.
In this study, we aimed to reveal the roles of TOP2A in the EMT, migration and invasion of cervical cancer, and to explore whether PI3K/AKT signaling was involved in. Our results showed that TOP2A was highly expressed in cervical cancer tissues, and TOP2A overexpression significantly facilitated cervical cancer cell migration, invasion and EMT, but these results were significantly repressed when the PI3K/AKT signaling was repressed by LY294002.
Materials and Methods
Tissue Samples and Ethic Statements
Twenty paired cervical cancer tissues and the paracancerous normal tissues were obtained from patients with cervical cancer (mean age 48.5±7.3 years old). All samples were collected between January 2015 and January 2018 from the Affiliated Cancer Hospital of Guizhou Medical University. The written informed consents were obtained from all patients before this study. The protocols involving human samples were conducted in accordance with the Declaration of Helsinki and were approved by the Ethic Committee of the Affiliated Cancer Hospital of Guizhou Medical University.
Cell Lines and Culture
Six human cervical cancer cell lines, including SiHa, CC-10A, CaSki, CAC-1, CaLo, and HeLa were acquired from the American Type Culture Collection (ATCC, Manassas, VA, USA). All of the cell lines were grown in Dulbecco’s modified Eagle’s medium (DMEM), supplemented with 10% fetal bovine serum (FBS) and 1% (v/v) penicillin and streptomycin, and maintained in a humidified atmosphere at 37°C with 5% CO2. All of the above reagents were obtained from Thermo Fisher Scientific (MA, USA).
Inhibition of PI3K/AKT Signaling
To repress the activation of PI3K/AKT signaling, the cervical cancer cells were treated with 25 μM LY294002 (MedChemExpress, Shanghai, China), a selective inhibitor of PI3K/AKT signaling for 24 hours.
Cell Transfection
To upregulate or downregulate TOP2A expression, the cervical cancer cells were instantaneously transfected with the small interfering RNAs (siRNAs, si-TOP2A, cat. no. SR322074, OriGene, Beijing, China) or the overexpressing plasmid of TOP2A (OE-TOP2A, cat. no. SC127832, OriGene). Cell transfection was carried out by using the Lipofectamine® 2000 reagent (Thermo Fisher Scientific) based on the manufacturer’s instructions, when cell confluence reached at 50–60%. In addition, the negative controls of si-TOP2A and OE-TOP2A vectors were also obtained from OriGene, which were named as si-NC and OE-NC.
Western Blotting
With the help of RIPA lysis buffer (Solarbio, Beijing, China) and protease inhibitor cocktail (Roche Diagnostics, Indianapolis, IN), total proteins were extracted from cervical tissues and cells. Following protein concentrations being measured by using the Bradford assay (BCA, Thermo Fisher Scientific), same amount of protein samples (25 μg) from various groups were separated by the SDS-PAGE and blotted onto the nitrocellulose filter membranes (Millipore, MA, USA). Then, the membranes were sealed with 5% non-fat skim milk for 30 min at room temperature. After that, the membranes were probed with the primary antibodies at 4°C overnight and then incubated with the corresponding HRP-conjugated second antibodies (Santa Cruz Biotechnology, Shanghai, China) at room temperature for 1 hour. The primary antibodies used in this experiment included TOP2A (cat. no. ab52934; 1:1000 dilution), p-AKT (cat. no. ab38449; 1:1000 dilution), AKT (cat. no. ab8805; 1:2000 dilution), and GAPDH (cat. no. ab181602; 1:5000 dilution), all obtained from Abcam (Cambridge, MA, USA). After that, the protein levels in the bands were quantified by using an enhanced chemiluminescence (ECL) detection system (Millipore).
Transwell Chamber Assay
To evaluate the effect of TOP2A/AKT on the invasion and migration of cervical cancer cells, transwell chambers with/without basement membrane matrix (8-μm pore; Corning, Life Science, Lowell, MA, USA) were applied, respectively. In brief, the transfected cells (5×104/well) were suspended in serum-free medium and inoculated in the upper chambers, while the lower chambers were supplemented with 600 μL cell culture medium with 15% FBS. Then, the cells were maintained at 37 °C with 5% CO2 for 24 hour and 48 hours to detect cell migration and invasion abilities, severally. After that, cells in the lower of the upper chambers were stained with 1% crystal violet for 20 min at room temperature. Cell invasion and migration abilities were assessed by counting cell numbers with positive staining in six randomly selected regions.
Statistical Analysis
Statistical analyses were performed using the SPSS software (version 22.0). Data in this study were expressed as mean ± SD. Comparisons between 2 groups were executed by using the student’s t-test, including TOP2A expression levels between tumor tissues and normal tissues in Figures 1A and Supplementary Figure 1, and TOP2A expression in Table 1. And, comparisons between multiple groups were executed by using the one-way ANOVA followed by Tukey post hoc, including protein expression, cell invasion and cell migration abilities between 3 groups (Figures 2, 4 and 5). If a p value is less than 0.05, the differences between groups were considered statistically significant.
 |
Table 1 Relationship Between TOP2A Expression and Clinicopathological Features in 20 Cervical Cancer Cases (*p<0.05) |
 |
Figure 1 TOP2A expression was increased in cervical cancer tissues. (A) The expression of TOP2A protein in 4 paired cervical cancer tissues and the adjacent normal tissues was determined by using the Western blotting assay. (B) Western blotting analysis of the protein levels of TOP2A in six cervical cancer cell lines. (*p<0.05). Abbreviations: TOP2A, Topoisomerases type IIA; N, Normal tissue; T, Tumor tissue; GAPDH, glyceraldehyde-3-phosphate dehydrogenase. |
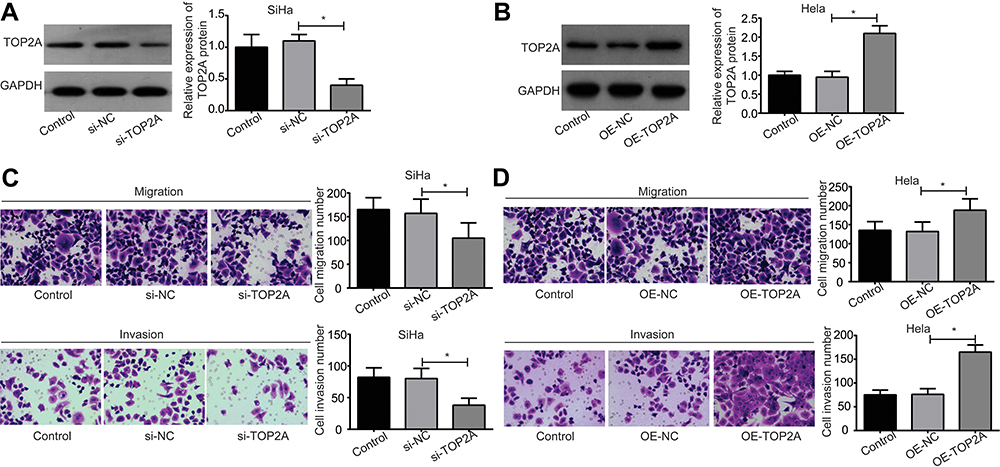 |
Figure 2 Evaluation of the effect of TOP2A on the migration and invasion of cervical cancer cells. (A) The expression of TOP2A protein was detected by using the Western blotting assay after SiHa cells were transfected with the si-NC or si-TOP2A. (B) The expression of TOP2A protein was detected by using the Western blotting assay after Hela cells were transfected with the OE-NC or OE-TOP2A. (C) The migration and invasion abilities of SiHa cells were determined by using the transwell chambers after the cells were transfected with si-NC or si-TOP2A. (D) The effect of TOP2A overexpression on the migration and invasion of Hela cells were measured by using the transwell chambers. (*p<0.05). Abbreviations: TOP2A, Topoisomerases type IIA; si, small interfering RNA; NC, negative control; OE, overexpressing; GAPDH, glyceraldehyde-3-phosphate dehydrogenase. |
Results
TOP2A is Overexpressed in Cervical Cancer Tissues
To reveal the role of TOP2A plays in the development and progression of cervical cancer, we first compared the expression patterns of TOP2A in 20 cases of cervical cancer tissues and the paired adjacent normal tissues. Compared with the normal tissues, TOP2A protein level was significantly increased in 85% (17/20) of cervical cancer tissues (Figures 1 and Supplementary Figure 1), and 4 representative images selected from the 17 paired cancer tissues with TOP2A high expression were shown in Figure 1A. In addition, TOP2A expression was significantly increased in cervical cancer tissues with advanced TNM stage, poor differentiation, large tumor size and lymph node metastasis (Table 1). Moreover, we randomly selected six cervical cancer cell lines (SiHa, CC-10A, CaSki, CAC-1, CaLo, and HeLa) and detected the expression levels of TOP2A in these cell lines. The results demonstrated that TOP2A protein showed different expression patterns of TOP2A in SiHa, CC-10A, CaSki, CAC-1, CaLo, and HeLa cells, with Hela the lowest and SiHa the highest (Figure 1B). Therefore, we chose Hela and SiHa cell lines for the next experiments.
TOP2A Promotes the Migration and Invasion of Cervical Cancer Cells
Then, both of the loss-of-function assays in SiHa cells and the gain-of-function assays in Hela cells were performed to investigate the effect of TOP2A on the migration and invasion of cervical cancer cells. The expression of TOP2A protein was significantly decreased when SiHa cells were transfected with the si-TOP2A as compared with the si-NC group, and si-NC transfection induced no obvious change in the expression of TOP2A compared to control group (Figure 2A). However, OE-TOP2A transfection leaded to an obvious increase in the protein level of TOP2A in Hela cells as compared with the OE-NC group (Figure 2B). Knockdown of TOP2A significantly weakened SiHa cell migration and invasion abilities (Figure 2C), and overexpression of TOP2A in Hela cells significantly enhanced cell migration and invasion abilities (Figure 2D). These results demonstrate that TOP2A facilitates cell migration and invasion in cervical cancer.
TOP2A Induces EMT and PI3K/AKT Activation in Cervical Cancer Cells
To reveal the underlying mechanism of TOP2A in promoting cervical cancer migration and invasion, we also assessed the effects of TOP2A on EMT process and the activation of PI3K/AKT signaling. The results showed that the cell morphology of SiHa was transformed from shuttle shape to roundness when TOP2A was downregulated (Figure 3), whereas the morphology of Hela cells was transformed to shuttle shape when TOP2A was overexpressed (Figure 3). In addition, the expression levels of N-cadherin and p-AKT were significantly decreased, and E-cadherin expression was increased when SiHa cells were transfected with si-TOP2A (Figure 4). To the contrary, the expression levels of N-cadherin and p-AKT were increased and E-cadherin expression was decreased when Hela cells were transfected with OE-TOP2A (Figure 4). These results illustrate that TOP2A induces EMT and PI3K/AKT activation in cervical cancer cells.
 |
Figure 3 Assessment of TOP2A roles in cervical cell EMT. The cell morphology of Hela and SiHa cells in different groups was recorded using the inverted microscope. (Scale bar=100 μm). Abbreviations: TOP2A, Topoisomerases type IIA; si, small interfering RNA; NC, negative control; OE, overexpressing; EMT, epithelial–mesenchymal transition. |
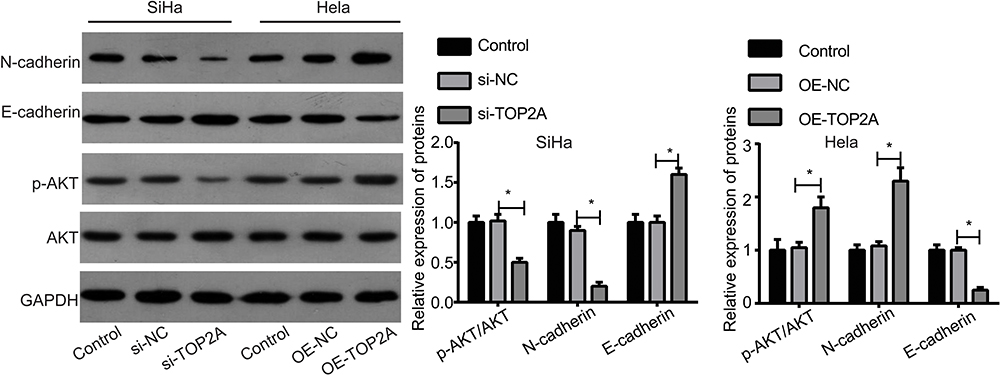 |
Figure 4 Assessment of the effects of TOP2A on the expressions of E-cadherin, N-cadherin, p-AKT and AKT. The protein levels of p-AKT, AKT, N-cadherin and E-cadherin were tested by Western blotting in different groups of Hela and SiHa cell lines. (*p<0.05). Abbreviations: TOP2A, Topoisomerases type IIA; si, small interfering RNA; NC, negative control; OE, overexpressing; AKT, serine/threonine kinase; p-AKT, phosphorylated-serine/threonine kinase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase. |
TOP2A Promotes Cervical Cancer Cell Migration, Invasion and EMT via Activating the PI3K/AKT Signaling
Finically, we explored the effect of PI3K/AKT pathway on TOP2A-mediated cervical cancer cell migration, invasion and EMT. LY294002, an inhibitor of PI3K/AKT signaling was recruited to repress the activation of PI3K/AKT signaling in Hela cells. Compared with the OE-TOP2A group, the expression of p-AKT was significantly reduced when Hela cells were treated with LY294002 (Figure 5A). In addition, LY294002 administration dramatically abolished the morphology change of Hela cells induced by TOP2A overexpression (Figure 5B). Moreover, LY294002 treatment obviously rescued the promotions of cell migration and invasion caused by TOP2A overexpression in Hela cells (Figure 5C). These results show that TOP2A promotes cervical cancer cell migration, invasion and EMT via activating the PI3K/AKT signaling.
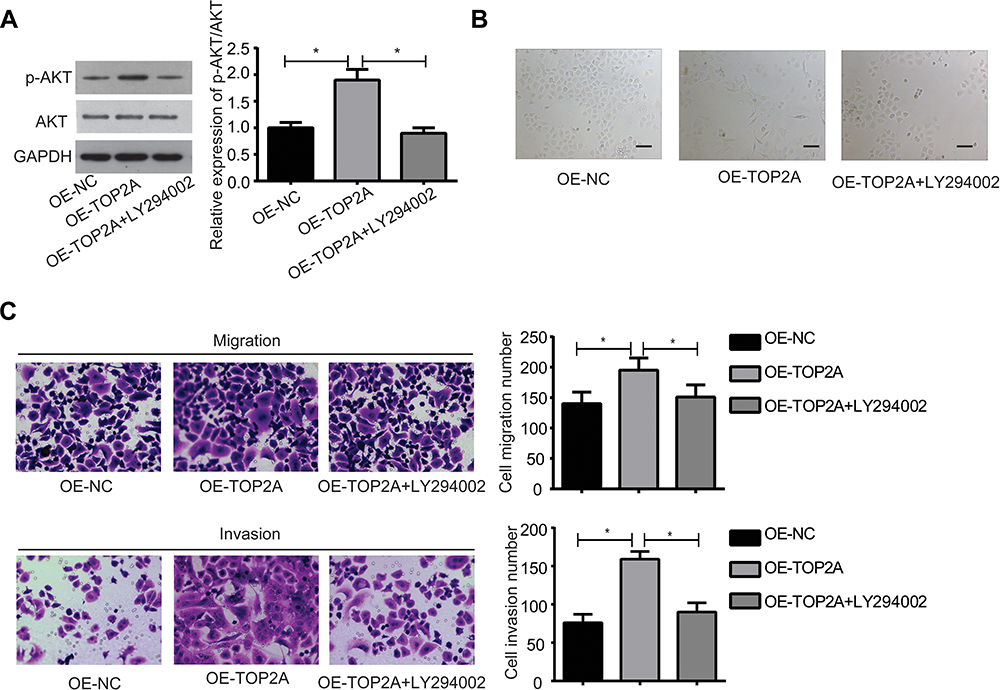 |
Figure 5 Inhibition of the PI3K/AKT signaling rescued TOP2A roles in the promotions of cell EMT, migration and invasion in Hela cells. Hela cells were divided into the OE-NC, OE-TOP2A and OE-TOP2A+LY294002 groups and were submitted to the following experiments. (A) Western blotting assay was used to detect the expressions of p-AKT and AKT proteins. (B) Cell morphology was recorded using the inverted microscope (Scale bar=100 μm). (C) Cell migration and invasion were assessed by using the transwell chambers. (*p<0.05). Abbreviations: TOP2A, Topoisomerases type IIA; NC, negative control; OE, overexpressing; p-AKT, phosphorylated-serine/threonine kinase; AKT, serine/threonine kinase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; EMT, epithelial–mesenchymal transition; PI3K, phosphatidylinositol 3-kinase. |
Discussion
It’s well known that the malignant progression of cervical cancer is closely associated with the enhanced motility and invasiveness of cancer cells.17–19 In this study, we investigated the effects of TOP2A on the migration, invasion and EMT of cervical cancer cells. Our results demonstrated that upregulation of TOP2A significantly enhanced the invasion, migration and EMT of cervical cancer cells via facilitating the activation of PI3K/AKT signaling.
TOP2A, as a member of TOP2 family, is essential for DNA replication, chromosome condensation, and the separation of sister chromatids during mitosis, as well as plays an important role in the transcriptional initiation.20 Up to now, TOP2A was shown to be implicated in the progression of many kinds of cancers. A most well-known cancer associated with TOP2A is breast cancer, in which TOP2A is usually simultaneously deleted or overexpressed with ERBB2 (erb-b2 receptor tyrosine kinase 2), thus the two genes are usually co-tested to guide the usage of Herceptin, an anticancer agent.21,22 With the help of bioinformatic analysis and immunohistochemistry (IHC) technology, Ma at al.23 revealed that TOP2A was overexpressed in non‑small cell lung cancer (NSCLC) comparing to normal lung tissues, which associated with the worse overall survival in NSCLC patients. Song et al24 demonstrated that TOP2A expression was dramatically increased following human cytomegalovirus (HCMV) infection in glioma cells, and miR-144-3p upregulation significantly reduced TOP2A expression to inhibit cell viability and invasion of HCMV-positive glioma cells. Labbe et al25 found that the high expression of TOP2A and EZH2 in prostate cancer patients closely linked to aggressive disease progression. In addition, they also clarified that TOP2A and EZH2 acted as key driving oncogenes in prostate cancer, which was concluded from a fact of sensitivity to combination-targeted therapy.25 In cervical cancer, TOP2A expression was not only identified to be increased, but also associated with HPV infection,13,14,26 indicating that TOP2A might exert a role in the progression of cervical cancer. To further elucidate the specific role of TOP2A in cervical cancer progression, we first investigated its expression pattern in cervical cancer tissues and cells. The results showed that TOP2A expression was significantly upregulated in cervical cancer tissues, especially in cervical cancer tissues with advanced TNM stage, poor differentiation, large tumor size and lymph node metastasis, as compared with the normal tissues. In addition, we observed that TOP2A was differently expressed in various cervical cancer cell lines, including SiHa, CC-10A, CaSki, CAC-1, CaLo, and HeLa, among which SiHa showed the highest expression and Hela showed the lowest expression. Moreover, we clarified that TOP2A overexpression in Hela cells markedly enhanced cell migration and invasion abilities, while knockdown of TOP2A in SiHa cells caused the opposite results.
EMT is of importance to induce cancer cell migration and invasion.8 In this process, epithelial cells transform into mesenchymal cells with decreased expression of epithelial cells markers, such as E-cadherin, and increased expression of mesenchymal cell markers, such as N-cadherin.27 To further reveal the role and mechanism underlying TOP2A in promoting the migration of cervical cancer, we also explored TOP2A function in the EMT of cervical cancer cells. Consistent with cell migration and invasion, TOP2A upregulation dramatically accelerated the EMT process in cervical cancer cells with a decreased expression of E-cadherin and an increased expression of N-cadherin, as well as induced cell morphology to shuttle shape.
The hyper-activation of PI3K/AKT signaling can significantly accelerate cancer cell proliferation, migration and EMT and inhibit cell apoptosis in majority kinds of cancers, including cervical cancer.28,29 A work by Wang et al16 in testicular seminoma using bioinformatics analysis demonstrated that TOP2A expression was associated with the activation of PI3K/AKT and Wnt/β‑catenin signaling pathways. Here, we found that TOP2A overexpression significantly promoted the activation of PI3K/AKT signal with increased expression of p-AKT in cervical cancer Hela cells, suggesting that PI3K/AKT signal might be involved in TOP2A-mediated cervical cancer progression. To this end, LY294002 was applied to suppress the activation of PI3K/AKT signaling in Hela cells. We observed that suppression of PI3K/AKT signaling apparently whittled TOP2A-mediated enhancements in cell migration, invasion and EMT.
In conclusion, our findings demonstrate that TOP2A is overexpressed in cervical cancer tissues, which then obviously promotes cancer cell migration, invasion and EMT via activating the PI3K/AKT signaling. This study might provide an efficient target for the treatment and prevention of cervical cancer migration.
Disclosure
The authors declare that they have no competing interests.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. PMID: 30207593. doi:10.3322/caac.21492
2. Stoler MH, Schiffman M. Interobserver reproducibility of cervical cytologic and histologic interpretations: realistic estimates from the ASCUS-LSIL Triage Study. JAMA. 2001;285(11):1500–1505. PMID: 11255427. doi:10.1001/jama.285.11.1500
3. Bardin A, Vaccarella S, Clifford GM, et al. Human papillomavirus infection in women with and without cervical cancer in Warsaw, Poland. Eur J Cancer. 2008;44(4):557–564. PMID: 18191395. doi:10.1016/j.ejca.2007.12.001
4. Zhang X, Wang Y, Cao Y, Zhao H. Increased CCL19 expression is associated with progression in cervical cancer. Oncotarget. 2017;8(43):73817–73825. PMID: 29088748. doi:10.18632/oncotarget.17982
5. Samatov TR, Tonevitsky AG, Schumacher U. Epithelial-mesenchymal transition: focus on metastatic cascade, alternative splicing, non-coding RNAs and modulating compounds. Mol Cancer. 2013;12(1):107. PMID: 24053443. doi:10.1186/1476-4598-12-107
6. Kalluri R, Neilson EG. Epithelial-mesenchymal transition and its implications for fibrosis. J Clin Invest. 2003;112(12):1776–1784. PMID: 14679171. doi:10.1172/JCI20530
7. Rout-Pitt N, Farrow N, Parsons D, Donnelley M. Epithelial mesenchymal transition (EMT): a universal process in lung diseases with implications for cystic fibrosis pathophysiology. Respir Res. 2018;19(1):136. PMID: 30021582. doi:10.1186/s12931-018-0834-8
8. Qureshi R, Arora H, Rizvi MA. EMT in cervical cancer: its role in tumour progression and response to therapy. Cancer Lett. 2015;356(2):321–331. PMID: 25281477. doi:10.1016/j.canlet.2014.09.021
9. Morimoto S, Tsuda M, Bunch H, Sasanuma H, Austin C, Takeda S. Type II DNA topoisomerases cause spontaneous double-strand breaks in genomic DNA. Genes. 2019;10(11):868. doi:10.3390/genes10110868 PMID: 31671674.
10. Wu KZ, Wang G-N, Fitzgerald J, et al. DDK dependent regulation of TOP2A at centromeres revealed by a chemical genetics approach. Nucleic Acids Res. 2016;44(18):8786–8798. PMID: 27407105. doi:10.1093/nar/gkw626
11. Terashima M, Ichikawa W, Ochiai A, et al. TOP2A, GGH, and PECAM1 are associated with hematogenous, lymph node, and peritoneal recurrence in stage II/III gastric cancer patients enrolled in the ACTS-GC study. Oncotarget. 2017;8(34):57574–57582. PMID: 28915696. doi:10.18632/oncotarget.15895
12. Zhang R, Xu J, Zhao J, Bai JH. Proliferation and invasion of colon cancer cells are suppressed by knockdown of TOP2A. J Cell Biochem. 2018;119(9):7256–7263. PMID: 29761838. doi:10.1002/jcb.26916
13. Branca M, Giorgi C, Ciotti M, et al. Over-expression of topoisomerase IIalpha is related to the grade of cervical intraepithelial neoplasia (CIN) and high-risk human papillomavirus (HPV), but does not predict prognosis in cervical cancer or HPV clearance after cone treatment. Int J Gynecol Pathol. 2006;25(4):383–392. PMID: 16990717. doi:10.1097/01.pgp.0000209573.54457.32
14. Brown CA, Bogers J, Sahebali S, Depuydt CE, De Prins F, Malinowski DP. Role of protein biomarkers in the detection of high-grade disease in cervical cancer screening programs. J Oncol. 2012;2012:289315. PMID: 22481919. doi:10.1155/2012/289315
15. Dai B, Yu R, Fan M, Yang T, Wang B, Zhang Y. HMQTF2 suppresses migration of the human cervical cancer HeLa cells by reversing EMT via the PI3K/Akt signaling pathway. Oncol Rep. 2019;42(4):1451–1458. PMID: 31364732. doi:10.3892/or.2019.7245
16. Wang K, Chen Y, Zhao Z, Feng M, Zhang S. Identification of potential core genes and miRNAs in testicular seminoma via bioinformatics analysis. Mol Med Rep. 2019;20(5):4013–4022. PMID: 31545448. doi:10.3892/mmr.2019.10684
17. Li X, Zhou Q, Tao L, Yu C. MicroRNA-106a promotes cell migration and invasion by targeting tissue inhibitor of matrix metalloproteinase 2 in cervical cancer. Oncol Rep. 2017;38(3):1774–1782. PMID: 28731196. doi:10.3892/or.2017.5832
18. Ye H, Zhang Y, Geng L, Li Z. Cdc42 expression in cervical cancer and its effects on cervical tumor invasion and migration. Int J Oncol. 2015;46(2):757–763. PMID: 25394485. doi:10.3892/ijo.2014.2748
19. Wang C, Gu W, Zhang Y, Ji Y, Wen Y, Xu X. Nicotine promotes cervical carcinoma cell line HeLa migration and invasion by activating PI3k/Akt/NF-kappaB pathway in vitro. Exp Toxicol Pathol. 2017;69(6):402–407. PMID: 28385482. doi:10.1016/j.etp.2017.03.006
20. Madabhushi R. The roles of DNA topoisomerase IIbeta in transcription. Int J Mol Sci. 2018;19(7):1917. doi:10.3390/ijms19071917 PMID: 29966298.
21. Chen JR, Chien HP, Chen KS, et al. Amplification of HER2 and TOP2A and deletion of TOP2A genes in a series of Taiwanese breast cancer. Medicine. 2017;96(2):e5582. PMID: 28079792. doi:10.1097/MD.0000000000005582
22. Huijsmans CJ, van den Brule AJ, Rigter H, et al. Allelic imbalance at the HER2/TOP2A locus in breast cancer. Diagn Pathol. 2015;10:56. PMID: 26022247. doi:10.1186/s13000-015-0289-x
23. Ma W, Wang B, Zhang Y, et al. Prognostic significance of TOP2A in non-small cell lung cancer revealed by bioinformatic analysis. Cancer Cell Int. 2019;19:239. PMID: 31528121. doi:10.1186/s12935-019-0956-1
24. Song J, Ma Q, Hu M, Qian D, Wang B, He N. The inhibition of miR-144-3p on cell proliferation and metastasis by targeting TOP2A in HCMV-positive glioblastoma cells. Molecules. 2018;23(12):3259. doi:10.3390/molecules23123259 PMID: 30544723.
25. Labbe DP, Sweeney CJ, Brown M, et al. TOP2A and EZH2 provide early detection of an aggressive prostate cancer subgroup. Clin Cancer Res. 2017;23(22):7072–7083. PMID: 28899973. doi:10.1158/1078-0432.CCR-17-0413
26. Peres AL, Paz ESKM, de Araujo RF, et al. Immunocytochemical study of TOP2A and Ki-67 in cervical smears from women under routine gynecological care. J Biomed Sci. 2016;23(1):42. PMID: 27175798. doi:10.1186/s12929-016-0258-z
27. Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–196. PMID: 24556840. doi:10.1038/nrm3758
28. Yan SP, Chu DX, Qiu HF, et al. LncRNA LINC01305 silencing inhibits cell epithelial-mesenchymal transition in cervical cancer by inhibiting TNXB-mediated PI3K/Akt signalling pathway. J Cell Mol Med. 2019;23(4):2656–2666. PMID: 30697971. doi:10.1111/jcmm.14161
29. Huang L, Huang Z, Fan Y, et al. FOXC1 promotes proliferation and epithelial-mesenchymal transition in cervical carcinoma through the PI3K-AKT signal pathway. Am J Transl Res. 2017;9(3):1297–1306. PMID: 28386355.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.