Back to Journals » ImmunoTargets and Therapy » Volume 13
Thymic NK-Cells and Their Potential in Cancer Immunotherapy
Authors Forbes C ![]() , Nierkens S, Cornel AM
, Nierkens S, Cornel AM ![]()
Received 22 September 2023
Accepted for publication 20 January 2024
Published 25 March 2024 Volume 2024:13 Pages 183—194
DOI https://doi.org/10.2147/ITT.S441639
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Sarah Wheeler
Caitlyn Forbes,1 Stefan Nierkens,1,2,* Annelisa M Cornel1,2,*
1Princess Máxima Center for Pediatric Oncology, Utrecht University, Utrecht, the Netherlands; 2Center for Translational Immunology, University Medical Center Utrecht, Utrecht University, Utrecht, the Netherlands
*These authors contributed equally to this work
Correspondence: Stefan Nierkens, Utrecht University, Heidelberglaan 25, Utrecht, 3584 CS, the Netherlands, Tel +88 972 72 72, Fax +88 972 60 09, Email [email protected]
Abstract: Natural killer (NK)-cells are innate immune cells with potent anti-tumor capacity, capable of recognizing target cells without prior exposure. For this reason, NK-cells are recognized as a useful source of cell therapy. Although most NK-cells are derived from the bone marrow (BM), a separate developmental pathway in the thymus also exists, producing so-called thymic NK-cells. Unlike conventional NK-cells, thymic NK (tNK)-cells have a combined capacity for cytokine production and a natural ability to kill tumor cells in the presence of NK-cell receptor stimulatory ligands. Furthermore, tNK-cells are reported to express CD3 subunits intracellularly, without the presence of a rearranged T-cell receptor (TCR). This unique feature may enable harnessing of these cells with a TCR to combine NK- and T-cell effector properties in one cell type. The development, phenotype, and function of tNK-cells, and potential as a cell therapy is, however, poorly explored. In this review, we provide an overview of current literature on both murine and human tNK-cells in comparison to conventional BM-derived NK-cells, and discuss the potential applications of this cellular subset in the context of cancer immunotherapy.
Keywords: tumor immunogenicity, cancer immunotherapy, gene engineering, T-cell receptor
Introduction
Natural killer (NK) cells are large, granular CD3−CD56+ lymphocytes that function as a part of the innate immune system.1–3 Traditionally, NK-cells are thought to develop in the bone marrow (BM) and are subdivided into two classes: the “immature” CD56brightCD16− fraction, known for high cytokine production, and the “mature” CD56dimCD16+ NK-cell fraction, known for cytotoxicity.4–6 The activity of these cells is dictated based on a balance of engagement of activating and inhibitory receptors.5,6 Conventionally, NK-cells are known for their ability to target tumor cells without prior exposure, predominately through a mechanism referred to as “missing-self recognition”.3,5,7,8 Here, the absence of major histocompatibility complex class I (MHC-I) molecules on the surface of a target cell acts as a strong activating signal for NK-cells. Some tumors display plasticity of MHC-I expression in the presence of cytokines such as interferon (IFN)-γ, which is released by activated NK-cells:8 MHC-I upregulation can therefore be considered an escape mechanism to circumvent targeting by NK-cells. However, this at the same time increases the possibility for recognition by MHCI-antigen complex-specific cytotoxic effector cells.5
During the 1990s, evidence emerged suggesting the presence of an unconventional, thymus-derived NK-cell subclass in the CD56bright fraction.9–11 Interestingly, these cells share the cytotoxic capacity of the “mature” NK-cells with the cytokine-releasing capacity of the “immature” NK-cells.9–13 This intrinsic combination of effector mechanisms gives the cells a “dual functionality” and makes them an interesting alternative in the development of cellular therapy against cancer. Moreover, thymic NK (tNK)-cells express CD3 in their cytoplasm, harboring the potency of (forced) T-cell receptor (TCR) expression as a strategy to equip these innate cells with adaptive antigen-specific recognition receptors. Clinical use of tNK-cells could be complicated due to their low cell counts in peripheral blood (approximately 1–5%), although cord blood may present an interesting alternative source, with approximately 21–38% being identified as tNK-cells.14
Interestingly, De Smedt et al reported a method for the generation of human tNK-cells from CD34+ stem cells in co-culture with OP9-DL1 cells.14 These cells gradually upregulate CD56 expression, express high levels of intracellular CD3, produce high levels of cytokines (such as IFN-γ), and are highly cytolytic against the NK-sensitive cell line, K562.14 To support their generation, IL-15, FLT3L, SCF, and IL-7 are added to culture, and small levels of Notch signaling are reported to play an important role in supporting induction of the tNK phenotype. Similar culture techniques, albeit without Notch signaling, are already in use to generate conventional NK (cNK)-cells for therapeutic application, supporting the feasibility of this technique in a clinical setting.15–18
The field of tNK-cells remains in its infancy and has been explored by only a few. Murine models have predominately been utilized for research on these cells and translation to the human setting is not straightforward. Therefore, this review will give an overview of the phenotype, function, and developmental pathways of tNK-cells in both mice and humans, with reference to the differences between these and the better characterized cNK-cells. We will delve into how the unique features of tNK-cells make them an intriguing, novel source for cellular therapy in the treatment of tumors, in particular those exerting plasticity in MHC-I expression.
Phenotype of tNK-Cells
cNK-cells have been well characterized in both mice and humans over the years. There is no singular definitive marker for NK-cells: in mice, cNK-cells are defined as CD3−CD122+NK1.1+, whilst human cNK-cells are described as CD3−CD56+.1,9,19 Human cNK-cells can be further divided based on CD56 and CD16 expression, into the “immature” CD56brightCD16dim/- and “mature” CD56dimCD16+ subsets.1 Conventionally, the relationship between these two subsets is considered to be linear, although some studies argue there may be a higher degree of complexity than initially anticipated.2,11,20 Markers such as NKp46, NKG2D, CD49b, CD161, CD69, and CD27, amongst others, are commonly associated with cNK-cells.1,16,21,22 Further, receptors of the Ly49 family play important roles in cNK inhibition and activation.1 cNK and tNK-cells are reported to share expression of several markers including NK1.1 and DX5 (murine-specific), as well as CD122, NKp46, and NKG2D (murine and human); expression of further markers is also speculated.9,14,19,23,24 Despite some similarities, several key differences have been reported that give tNK-cells a distinct phenotype, of which the most notable distinguishing factors are intracellular expression of CD3 and rearranged TCR-γ transcripts.10,11,25 The former is predominately reported in human tNK-cells. Interestingly, few papers have reported these features in more recent years14,25,26 and one study contests the rearrangement of TCR-γ transcripts in mice.24 As later explored in the section “Developmental pathways of tNK-cells”, tNK-cells are hypothesised to develop from double negative (DN; CD4−CD8−) 1 and 2 thymocytes.12,27 The lack of a clear marker definition likely contributes to a lack of awareness of tNK-cells and scarce literature.
Most research into tNK-cells has been conducted in mice, pioneered by Vosshenrich et al, showing that murine tNK-cells can be defined as Lin−CD122+NK1.1+CD127+DX5+CD16dim/-CD11blowLy49low, with CD127 and GATA-3 as distinguishing markers.9,23,24 Based on this and other initial data, it was speculated that human tNK-cells feature homology with the CD56brightCD16dim/- NK-cell subset. This was supported by evidence that CD56bright cells in the human thymus express CD127 and feature functional similarity to tNK-cells.1,9 However, other studies challenge these findings, arguing that CD127 expression in CD56bright NK-cells is independent of the thymus, and does not correlate with intracellular CD3 expression.4,10,14,28,29 Interestingly, one study reports a clear difference between CD3−CD56+ cells in the thymus compared to the peripheral blood, identifying CD56dimCD16dim cells as human tNK-cells.30 However, these results have not been further explored in the literature. Further, in more recent years, updated innate lymphoid cell (ILC) nomenclature has complicated classification of certain innate subsets. Although not explored extensively in this review, readers can be directed to existing literature on the topic.23,31–35 In short, tNK-cells do not fit within any of the ILC groups, as they possess qualities that are attributed to more than one ILC subset. Further, the literature describes a divergence between ILC and cNK-cells during development, and where tNK-cells fall in these pathways remains a source of contention.23,31–35 Ultimately, despite some similarities with cNK-cells, tNK-cells have distinct features. Although some of these features are known, further research is warranted to define the phenotype of tNK-cells in detail, especially in the human setting in health and disease.
Similarities Between tNK-Cells and Other Immune Cell Subsets
tNK-cells bear resemblance to many different types of immune cells. Due to the aforementioned lack of a clear definition for these cells, they are often incorrectly grouped with other immune cell types in the literature. Therefore, it is important to draw clear distinctions between tNK-cells and similar cell types, such as natural killer T (NKT) cells, γδ T-cells, and mucosal-associated invariant T-cells (MAIT cells).
NKT-cells are conventionally described as T-cells that recognize lipid antigens presented by the CD1d molecule.36,37 There are two subsets of CD1d-restricted NKT-cells: the more commonly known type I invariant NKT (iNKT) cells, which express an invariant Vα24Jα18 chain on their surface in humans, and type II NKT-cells, which are more abundant and express diverse TCR-α and TCR-β chains.36,37 Both subsets, which arise in the thymus from double positive (DP) thymocytes or natural killer cell progenitor (NKP) cells, express markers commonly associated with NK-cells and can be activated either through their TCR or through NK-cell-like mechanisms.27,36 Their phenotypic similarities may confound identification of true tNK-cells;6,23,36 a recent publication even referred to these cells as ‘CD1d-restricted tNK-cells’.38 An alternative, albeit lesser-known subset of NKT-cells termed NKT-like cells also exist. These cells are CD3+CD56+, are not CD1d restricted, are more abundant in PB than their CD1d-restricted counterparts, secrete many cytokines, and mediate MHC-I restricted and unrestricted mechanisms of cytotoxicity.36,39,40 Despite many phenotypic and functional similarities, a key distinguishing feature is the presence of a TCR on the surface of the NKT subsets, marking them as distinct.39–42
γδ T-cells may also share resemblance to cNK or tNK-cells in their phenotype and function.43,44 Stewart et al refer to a γδ T-cell subset that express NK1.1 and CD127, alongside low TCR-γδ expression, as ‘NK-like γδ T-cells’. Indeed, the intracellular expression of CD3 in human tNK-cells, as well as rearrangements to TCR-γ and some TCR-δ genes in murine tNK-cells, suggest a relationship between γδ T-cells and tNK-cells.10,11,25,45 However, Ribeiro et al described different enzymatic requirements of the two subsets during development, and reported only low levels of TCR-γ rearrangements in murine tNK-cells.24 Ultimately, although the presence of TCR-γδ clearly distinguishes them from tNK-cells, the relationship between these two cell types remains unclear, and should be further investigated.
MAIT cells, which also develop in the thymus, are abundant in both mucosal and non-mucosal tissues in humans.46,47 These innate-like T-cells can be activated through TCR-dependent and -independent mechanisms, belonging to the T-cell subset that recognize antigens presented though MHC-related molecule 1 (MR1).46,47 Although innate-like in their function and despite expression of some similar markers such as NKG2D and NKp46, the presence of TCR chains – namely, Vα7.2-Jα33 combined with Vβ2 or Vβ13 chains – on the surface of MAIT cells distinguishes them from tNK-cells.46,47
Finally, it is relevant to note that, in studies investigating the development of tNK-cells from CD34+ progenitors in fetal thymic organ culture, tNK-cells never underwent full VDJ rearrangement and never expressed a TCR on their surface.14,48 Further research from Woo et al reports generation of iNKT-cells from CD34+ progenitor cells in a method resembling that used to generate tNK-cells.49 However, these cells clearly expressed invariant TCR chains in their cytoplasm and on their surface: these chains are not reported in the cultures generating tNK-cells, suggesting the two cell types follow alternative developmental pathways and are therefore separate entities. Thus, although tNK-cells share similar origins, functions, or markers with other cellular subsets, certain markers clearly identify them as a unique population. A summary of the key differences and similarities as discussed can be found in Table 1.
 |
Table 1 Summary of Key Phenotypic Differences Between tNK-Cells, cNK-Cells and Other Thymic Entities. Although tNK-cells bear some functional and phenotypic similarities to cNK-cells and to other thymic entities such as NKT-, γδ T- and MAIT Cells, there are key differences that mark tNK-cells as distinct. The key markers that distinguish these subsets, based on literature cited and described in this review, are reported in this table. |
Developmental Pathways of tNK-Cells
The developmental pathway and signaling requirements of tNK-cells are distinct from those of cNK-cells and differ depending on the tissue, as well as the species (ie mouse or human) of origin (Figure 1).
 |
Figure 1 Proposed Model of tNK-cell Development. This model proposes the process of human tNK-cell development, based on existing murine and human research. Common lymphoid progenitors (CLP) seed the thymus in the form of NKPs or ETPs, the latter of which will form DN thymocytes. DN thymocytes will progress through typical stages of T-cell development, forming DN1 and DN2 thymocytes. These cells act as TNKP, thus can follow a pathway of NK or T-cell lineage commitment. NKP are also speculated to migrate from the BM (red arrow) and immediately follow the (thymic) NK-cell fate. The steps in tNK-cell development are unique to that seen in the BM, also involving different cytokines and resulting in the upregulation of different markers. Figure created with BioRender.com. Abbreviations: CLP, common lymphoid progenitor; cNK, conventional NK-cell; DN, double negative; ETP, early thymic progenitor; HSC, hematopoietic stem cell; iCD3, intracellular CD3; iNK, immature natural killer cell; NKP, natural killer progenitor; NKT, natural killer T-cell; rNKP, refined natural killer progenitor; TNKP, (bipotent) T and natural killer cell progenitor. |
An early model for generation of tNK-cells in mice was proposed by Di Santo and Vosshenrich in 2006, stating that early progenitor cells developed in the thymus, but differentiation and maturation occurred in peripheral lymphoid organs.19 Since this model, the possibility of murine tNK-cells developing from double negative (DN; CD4−CD8−) 1 and 2 thymocytes, which arise from early thymic progenitors (ETPs), has been explored.12,27 An alternative hypothesis describes the development of tNK-cells from a common, or bipotent, T- and NK-cell progenitor (TNKP) within the thymus. Interestingly, the concept of a bipotent TNKP arose from human studies, based on the expression of intracellular CD3 and TCR chain transcripts in the cytoplasm of NK-cells in the thymus,10,11,45,50 and has now also been identified and characterized in mice.51,52
Triple negative (TN; CD3−CD4−CD8−) thymocytes were first identified as having T- and NK-cell potential, although more recent work described TNKPs as being DN2 thymocytes, which display early stages of TCR gene rearrangement.25,26,45,53 Although the exact subset is unknown, it is agreed that DN thymocytes in mice also act as TNKPs.12,13,25–27,52 In the human thymus, TNKP have been described as CD34+CD7+CD1a−; murine equivalents can be defined as Lin−CD117+NK1.1+.21,54 Matching this definition, a publication by Hidalgo et al describes a pathway of human tNK-cell development involving CD34+CD1a−BMPRIA+ intrathymic precursor cells.55 They liken tNK-cell development to that seen in lymph nodes and tonsils, whereby CD34+BMPRIA+ cells may be the thymic equivalent of CD34+CD117+CD94− pre-NK-cells described in more conventional pathways of NK-cell development.55 Interestingly, these CD34+BMPRIA+ cells are also able to differentiate into T-cells, but predominately form NK-cells, even in fetal thymic organ cultures.55 These findings are supported in the literature, with multiple other reports of the generation of NK- or T-cells from human CD34+CD1a− or murine CD34+Lin− progenitor cells in fetal thymic organ culture.1,11,14,48,53
Some studies suggest additional alternative sources for tNK-cells besides a TNKP. For example, cells matching the phenotype and function of tNK-cells are produced in Rag−/− mice,1 as well as in the absence of Notch signaling.21 However, De Smedt et al report that early Notch signaling in progenitor cells is sufficient to skew the cell towards the T or NK lineage, even if the Notch signaling is not maintained at later stages of tNK development. Thus, it is possible that the signal that determines commitment to the tNK-cell fate may occur earlier than predicted, before a progenitor cell matches the conventionally accepted definition of TNKP.
Throughout the literature, it is clear that different signals are required in the development of murine and human tNK-cells. For example, the transcription factor GATA-3 and cytokine IL-7 are critical for murine tNK-cell development, yet neither are required for developing human tNK-cells.4,9,14,28 This is also reflected in the aforementioned phenotype of the cells, whereby expression of the IL-7 receptor, CD127, does not act as a definitive marker of human tNK-cells. Despite these differences, some parallels can be drawn. Low levels of Notch signaling support intracellular CD3 expression in humans, and potentially play a role in murine tNK-cell development.14,19,28,48,52 The Notch target gene DTX1 is also reported to play a role in tNK-development in humans.28 For example, NFIL3 is regarded as essential in the murine setting.23,52,55 It is hypothesized to also be important in the development of human tNK-cells, although it remains uncertain at what time point it may be required.52,55 IL-15 has been reported as a critical component for the development of all NK-cells,9 and tNK-cells are no exception. This cytokine plays roles in the proliferation, differentiation, and survival of precursor cells committed to the NK lineage,52,56 with a reported synergistic role with BMP signaling.55 IL-15 can also prime NK-cells and thus increase their sensitivity to tumors in vivo and in vitro.57 Other factors such as ID2, SCF, and FLT3L are also implicated in the development of tNK-cells in either setting.14,19,23,52,55,56 Finally, the presence of IL-2 has been reported as important in the human setting,58 although not all evidence supports this statement.53 Thus, although the precise details of tNK-cell development remain controversial, it is clear that these cells develop distinctly from their BM-derived cNK counterparts.
Role of tNK-Cells
The first function ascribed to tNK-cells was the targeting of proliferating thymocytes and selection and maintenance of T-cells in the thymus.12,21,59–61 Studies also suggest that they egress from the thymus and migrate to lymph nodes, BM, and liver.9,12,25 The presence of tNK-cells in peripheral tissues may allude to their potential migration and retention in the tumor microenvironment (TME). In fact, the source and in-depth identification of tumor-infiltrating NK-cells remains unknown,12 thus some may have thymic origins.
Murine tNK-cells have an immature phenotype and the potential to produce high levels of cytokines including IFN-γ, granulocyte-macrophage colony-stimulating factor (GM-CSF), and TNF-α.9 Although this spurred the hypothesis that human tNK-cells belong to the conventional immature subset, tNK-cells have also been described to retain their cytotoxic capacity – a feature typically associated with maturity.9 Multiple studies report that tNK-cells can effectively lyse YAC-1 cells, a murine thymoma cell line used to assess NK cytotoxic capacity,1 albeit at lower levels than cNK-cells or spleen-isolated NK-cells.9,13,19 Similar results are observed with other cell lines.25 Further, an early study describes a correlation between CD56 acquisition on human thymocyte precursors and an increase in cytolytic capacity against NK-sensitive tumor cell line K562.45 In the human setting, De Smedt et al describe production of CD56 expressing thymus-derived NK-cells, with a strong cytolytic and cytokine-producing capacity.14 It should be noted that the influence of variables such as age, gender, and disease status on the functionality of tNK-cells is not reported in the literature, thus is not explored in this review. Overall, the evidence therefore suggests that both murine and human tNK-cells are important cytokine producers, whilst also retaining cytolytic capacity against tumor cells.
tNK-Cells as a Novel Source for Cell Therapy
In recent years, NK-cell-based immune therapies have increased in popularity, as the innate features of NK-cells offer a solution to some of the limitations of T-cell therapies. Adoptive transfer of NK-cells is a commonly used approach in a broad range of cancer types with varying degrees of therapeutic success.5,6 NK-cell-engaging methods, such as antibody-based approaches, further extend the capacity of missing-self recognition with antibody-dependent cellular cytotoxicity.3,5,6 Other approaches involve genetic modification of NK-cells, for example in production of chimeric antigen receptor (CAR)-NK-cells.6,62 For a more in-depth overview of current efforts in NK-cell therapies, we refer interested readers to a comprehensive review on the topic.63 Within each of these approaches, the benefit of the dual characteristics of tNK-cells makes them interesting candidates for cellular immunotherapy (Figure 2A), although they are not explicitly employed as of yet.
 |
Figure 2 The Potential of tNK-cells in Anti-Cancer Immunotherapy. (A) tNK-cells combine the cytokine-releasing potential of immature, CD56brightCD16− conventional NK-cells with the cytotoxic potential of mature, CD56dimCD16+ conventional NK-cells, making them a unique cell source for NK-cell based therapies. (B) Evasion of MHC-I plasticity-mediated immune escape via TCR introduction in tNK-cells. T-cell cytotoxicity is evaded by low MHC-I expression, while tumor cells are subject to missing-self recognition by tNK-cells. Missing-self recognition results in IFN-γ secretion by NK-cells, triggering MHC-I expression on tumor cells, thereby evading missing-self mediated cytotoxicity by NK-cells. iCD3 expression in the tNK-cells provides a unique window to combine missing-self- and antigen-dependent cytotoxicity, thereby circumventing MHC-I plasticity-mediated immune escape. Figure created with BioRender.com. Abbreviations: cNK, conventional natural killer cell; iCD3, intracellular CD3; IFN, interferon; KIR, killer immunoglobulin-like receptor; MHC-I, major histocompatibility complex I; NCR, natural cytotoxicity receptor; TCR, T-cell receptor; tNK, thymic natural killer cell. |
Downregulation or absence of MHC-I is a commonly employed mechanism of immune escape displayed by a range of tumor types, but as earlier described, this change can be reversible: for example, in adult tumors such as head and neck squamous cell carcinoma and bladder cancer.8 Furthermore, in embryonal cell-derived pediatric tumors such as neuroblastoma (NBL), malignant rhabdoid tumors (MRT), and medulloblastoma (MBL), no/low MHC-I expression is an intrinsic feature3,5,6,8,64,65 but is highly plastic.64–68 In NBL tumors, the expression of MHC-I can be induced by the presence of active NK-cells through a contact-dependent process reliant on interferon (IFN)-γ production. Although this makes them less likely to be recognized by NK-cells, the tumor cells become more susceptible to targeting by cytotoxic T lymphocytes.64,66 The combination of NK-mediated missing-self cytotoxicity with TCR-driven antigen-specificity would therefore be an ideal therapeutic approach. This concept is already being explored in NK-cells,69 in which NK-cells are engineered to express a tumor-specific recombinant TCR as well as subunits of the CD3 complex. Utilization of tNK-cells for such an approach would circumvent the need to co-introduce CD3 subunits, making them an interesting alternative cell source for such a therapy approach (Figure 2B).
tNK-cells may be a particularly suitable NK-cell population for cellular immunotherapy, as they are considered potent cytokine producers and their transfer into the TME may thus be highly beneficial. For example, factors produced by tNK-cells, such as TNF-α and IFN-γ, are reported to overcome immune escape in MBL and NBL by inducing MHC-I expression.8,65 The innate dual functionality of tNK-cells may induce concordant lysis and additional recruitment of other immune cells. They further provide a unique opportunity for TCR transfer without the additional need to incorporate the machinery required for TCR functionality, as tNK-cells already express CD3 and rearranged TCR chains in the cytoplasm.
As a clear phenotype signature to isolate tNK-cells from peripheral- or cord blood is currently lacking, it is difficult to study the potential of tNK-cells as a cellular immunotherapy source. However, methods for the artificial generation of tNK-cells are described in the literature. The availability of a method to generate human tNK-cells from CD34+ progenitor cells, as described by De Smedt et al,14 provides the tools to study their potential and engineering. Similarly, Vargas et al describe a method for the generation of tNK-cells from murine thymocytes, also utilizing an OP9-based system.13 However, unlike De Smedt et al, they do not introduce Notch into culture, and these cells are not reported to express intracellular CD3 or TCR transcripts.13 Although it should be investigated whether this is a result of absence of Notch or a difference between murine and human tNK-cells, this protocol provides additional evidence for the feasibility of artificial generation of tNK-cells. In addition, OP9-DL cells are used in the generation of CAR-NK cells.70 Similar culture methods have been used to produce cNK-cells for application in clinical trials.15,71–73 Thus, it is possible to develop this protocol to suit good manufacturing practice (GMP) techniques for future therapeutic application. In particular, this approach has potential for clinical application in patients for which CD34+ progenitor cells are available, eg in patients with relapsed/refractory leukemias or neuroblastoma that receive a hematopoietic cell transplantation using harvested CD34+ cells. Finally, other stem cells, namely hematopoietic stem cells (HSCs) and induced pluripotent stem cells (iPSCs), are a common source for therapeutic cell products, including iNKT-, NK-, and T-cells.74 There are various existing protocols describing the generation of NK- and T-cells from these progenitors, many of which are covered by Li et al.74 To our knowledge, there is currently no protocol for the generation of tNK-cells from HSCs or iPSCs, however, these may offer an interesting alternative source for tNK-cells to explore. Overall, tNK-cells present an interesting alternative to current NK-based cell therapies, especially for pediatric tumors with inducible MHC-I expression. There is at least one existing approach for the artificial production of these cells, as described by De Smedt et al, enabling investigation into characteristics of the tNK-cell population. Furthermore, this approach has potential to be developed into a GMP-compliant protocol to enable therapeutic application.
Discussion
With this review, we have summarized what is known about tNK-cells, an interesting but poorly recognized subtype of NK-cells that develop in the thymus. We have compared the phenotype (summarized in Table 2) and developmental pathway of tNK-cells with that of cNK and hypothesized their additive potential to contribute to cell-based anti-tumor therapies. Furthermore, we described the possibility of these cells to provide a powerful, multi-pronged approach in the treatment of pediatric MHC-I inducible tumors, such as NBL, MRT, or MBL. However, many questions remain regarding the true identity and function of tNK-cells, especially in humans.
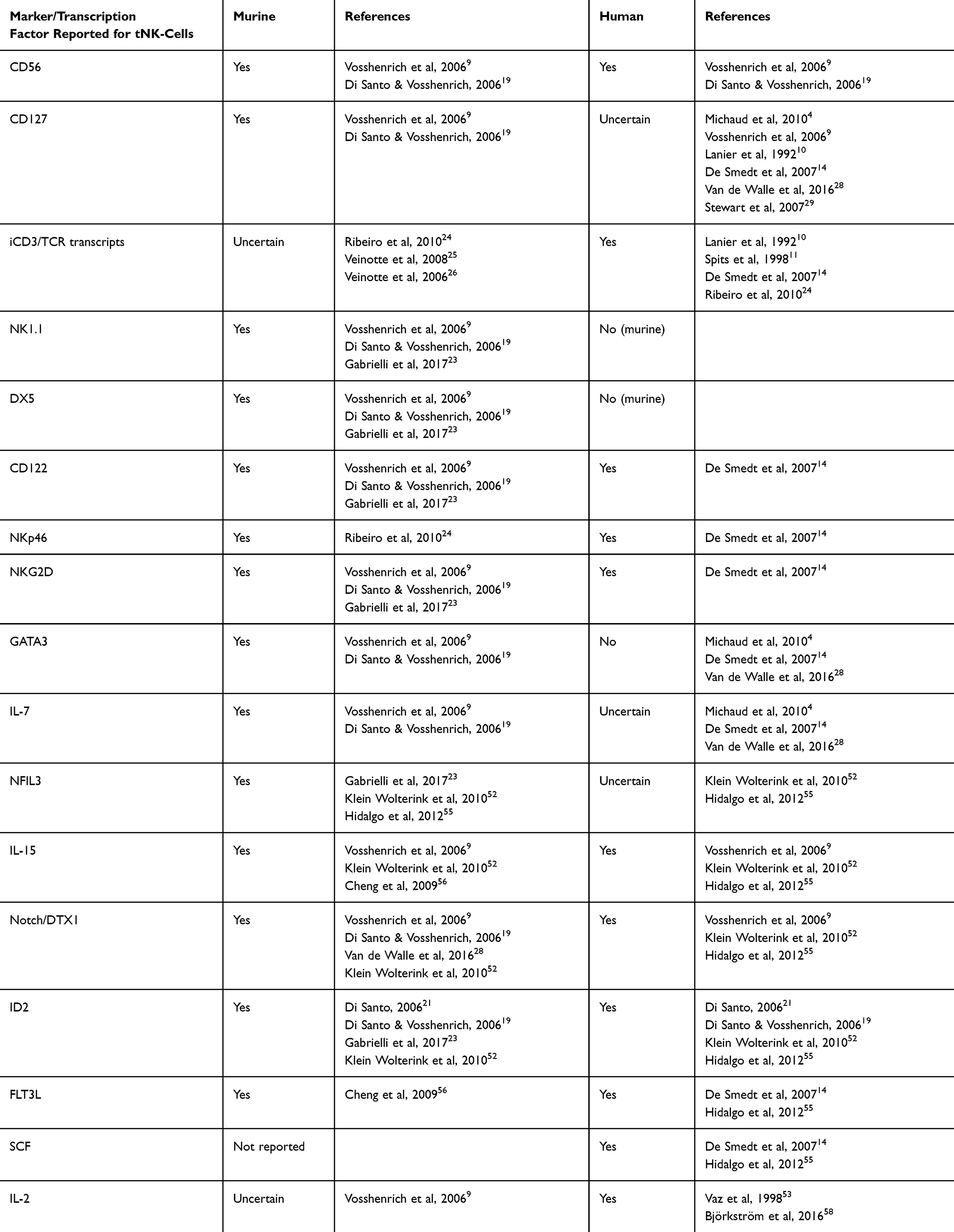 |
Table 2 A Summary of the Phenotype and Transcription Factor Requirements for Murine and Human tNK-Cells, as Reported in the Literature. Murine and human tNK-cells have varying phenotypes and transcription factor requirements during development. This table summarizes the most commonly described phenotypic markers and transcription factors in tNK-cells and whether these have been described in the murine or human setting/s. When opinions in the literature are mixed, the presence of these markers/factors is described as “uncertain”. |
To date, most studies on tNK-cells have been conducted in murine models, and translation to the human setting is not always obvious. Furthermore, there have been little to no updates in the research on tNK-cells since the earlier work of Vosshenrich et al, with most recent papers continuing to cite this work in their description of both murine and human tNK-cells. A lack of clear markers for tNK-cells means it is challenging to distinguish true tNK-cells, and complicates finding them in the periphery. Thus, further investigation of these cells will be key in uncovering their potential in (cancer) immunotherapy.
Future work should first focus on clarifying the phenotype and function of tNK-cells. Single cell RNA sequencing, or transcriptomics, could be utilized to identify the tNK-cell cluster first in thymic tissues, and later in peripheral compartments: De Smedt et al report a relatively higher percentage of tNK-cells in cord blood,14 thus this may offer an interesting source for investigation. Extensive flow panels including markers such as CD56, CD16, GATA3, CD127, CD1a, CD7, BMPRIA, and intracellular stains for CD3 and TCR chains (including γ and δ chains) can validate the expected tNK phenotype (based on literature, as summarized in Table 2), and can be used to compare naturally occurring tNK-cells with in vitro generated tNK-cells. Finally, surface proteomics can be implemented to investigate expression of further, potentially discriminating, markers on the surface of tNK-cells. After identification of tNK markers, these techniques can be used to study the presence of these cells within peripheral compartments, as well as their migratory cues. Being able to identify tNK-cells in the periphery and understand their function in these compartments would also enable better determination of their potential presence and role within the TME. In a therapeutic context, this further gives insight into how to target and activate these cells, and how to utilize their dual functionality. It would also be interesting to investigate their persistence and distribution at different ages (ie infants, young children, adults). Although it is clear that tNK-cells are distinct from other similar subsets such as cNK-cells NKT/iNKT-cells, γδ T-cells, and MAIT cells – made clear through distinct expression/absence of various surface markers – the relationship between some of these subsets and tNK-cells is not yet understood, and should be explored in more detail. Finally, the ability to generate these cells in culture is an important step, as it will support both further research and, in the future, clinical application. De Smedt et al have described the generation of cells matching the description of human tNK-cells, and this method holds promise for future clinical application and development into a GMP-compliant protocol.
Conclusion
The unique, intrinsic dual nature of tNK-cells – namely their combined cytokine-production and cytotoxic capacity – makes them an interesting novel option to explore in cancer cell therapy, especially in pediatric MHC-I inducible tumors. Unlike cNK, tNK-cells produce cytokines that are able to upregulate MHC-I on the tumor cell surface, whilst also retaining an ability to target these cells in the absence of MHC-I through missing-self recognition. Further, the intracellular expression of CD3 by these cells may prove advantageous in a therapeutic context. Additional arming of the cell through a tumor specific TCR would extend their functionality, enabling targeting of tumor cells regardless of MHC-I expression status, thus making them a powerful form of cell therapy. Overall, the unique characteristics of tNK-cells distinguish them from other cell types currently exploited in treatment of tumors, and provide a strong basis that can be utilized to develop a robust, multi-pronged cell therapy for use against MHC-I inducible pediatric tumors.
Abbreviations
BM, bone marrow; CAR, chimeric antigen receptor; CLP, common lymphoid progenitor; cNK, conventional natural killer cell; DN, double negative; DP, double positive; ETP, early thymic progenitor; GMP, good manufacturing practice; HSC, hematopoietic stem cell; iCD3, intracellular CD3; IFN, interferon; ILC, innate lymphoid cell; iNK, immature natural killer cell; iNKT, invariant natural killer T-cell; iPSC, induced pluripotent stem cell; KIR, killer immunoglobulin-like receptor; MAIT, mucosal-associated invariant T-cell; MBL, medulloblastoma; MHC-I, major histocompatibility complex class I; MRT, malignant rhabdoid tumors; MR1, MHC-related molecule 1; NBL, neuroblastoma; NCR, natural cytotoxicity receptor; NK, natural killer; NKP, natural killer cell progenitor; NKT, natural killer T-cell; rNKP, refined natural killer cell progenitor; SCF, stem cell factor; TCR, T-cell receptor; TME, tumor microenvironment; tNK, thymic natural killer cell; TNF, tumor necrosis factor; TNKP, (bipotent) T and natural killer cell progenitor.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Villa Joep Foundation (IWOV-Actief.51391.180034).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Huntington ND, Vosshenrich CAJ, Di Santo JP. Developmental pathways that generate natural-killer-cell diversity in mice and humans. Nat Rev Immunol. 2007;7(9):703–714. doi:10.1038/nri2154
2. Amand M, Iserentant G, Poli A, et al. Human CD56dimCD16dimCells as an individualized natural killer cell subset. Front Immunol. 2017;8:699. doi:10.3389/fimmu.2017.00699
3. Abel AM, Yang C, Thakar MS, et al. Natural killer cells: development, maturation, and clinical utilization. Front Immunol. 2018;9:1869. doi:10.3389/fimmu.2018.01869
4. Michaud A, Dardari R, Charrier E, et al. IL-7 enhances survival of human CD56bright NK cells. J Immunother. 2010;33(4):382–390. doi:10.1097/CJI.0b013e3181cd872d
5. Wu SY, Fu T, Jiang YZ, et al. Natural killer cells in cancer biology and therapy. Mol Cancer. 2020;19(1):120. doi:10.1186/s12943-020-01238-x
6. Molgora M, Cortez VS, Colonna M. Killing the invaders: NK cell impact in tumors and anti-tumor therapy. Cancers. 2021;13(4):595. doi:10.3390/cancers13040595
7. Raulet DH. Missing self recognition and self tolerance of natural killer (NK) cells. Semin Immunol. 2006;18(3):145–150. doi:10.1016/j.smim.2006.03.003
8. Cornel AM, Mimpen IL, Nierkens S. MHC class I downregulation in cancer: underlying mechanisms and potential targets for cancer immunotherapy. Cancers. 2020;12(7):1760. doi:10.3390/cancers12071760
9. Vosshenrich CAJ, García-Ojeda ME, Samson-Villéger SI, et al. A thymic pathway of mouse natural killer cell development characterized by expression of GATA-3 and CD127. Nat Immunol. 2006;7(11):1217–1224. doi:10.1038/ni1395
10. Lanier LL, Spits H, Phillips JH. The developmental relationship between NK cells and T cells. Immunol Today. 1992;13(10):392–395. doi:10.1016/0167-5699(92)90087-N
11. Spits H, Blom B, Jaleco AC, et al. Early stages in the development of human T, natural killer and thymic dendritic cells. Immunol Rev. 1998;165:75–86. doi:10.1111/j.1600-065X.1998.tb01231.x
12. Hashemi E, Malarkannan S. Tissue-Resident NK cells: development, maturation, and clinical relevance. Cancers. 2020;12(6):1553. doi:10.3390/cancers12061553
13. Vargas CL, Poursine-Laurent J, Yang L, et al. Development of thymic NK cells from double negative 1 thymocyte precursors. Blood. 2011;118(13):3570–3578. doi:10.1182/blood-2011-06-359679
14. De Smedt M, Taghon T, Van De Walle I, et al. Notch signaling induces cytoplasmic CD3ε expression in human differentiating NK cells. Blood. 2007;110(7):2696–2703. doi:10.1182/blood-2007-03-082206
15. Alves PCM, De Angelo Andrade LAL, Petta CA, et al. Ex vivo Expansion of CD56 + NK and NKT-like lymphocytes from peripheral blood mononuclear cells of patients with ovarian neoplasia. Scand J Immunol. 2011;74(3):244–252. doi:10.1111/j.1365-3083.2011.02576.x
16. Bi J, Wang X. Molecular regulation of NK cell maturation. Front Immunol. 2020;11:1945. doi:10.3389/fimmu.2020.01945
17. Yu H, Fehniger TA, Fuchshuber P, et al. Flt3 ligand promotes the generation of a distinct CD34+ human natural killer cell progenitor that responds to interleukin-15. Blood. 1998;92(10):3647–3657. doi:10.1182/blood.V92.10.3647
18. Grzywacz B, Kataria N, Sikora M, et al. Coordinated acquisition of inhibitory and activating receptors and functional properties by developing human natural killer cells. Blood. 2006;108(12):3824–3833. doi:10.1182/blood-2006-04-020198
19. Di Santo JP, Vosshenrich CAJ. Bone marrow versus thymic pathways of natural killer cell development. Immunol Rev. 2006;214:35–46. doi:10.1111/j.1600-065X.2006.00461.x
20. Freud AG, Yu J, Caligiuri MA. Human natural killer cell development in secondary lymphoid tissues. Semin Immunol. 2014;26(2):132–137. doi:10.1016/j.smim.2014.02.008
21. Di Santo JP. Natural killer cell developmental pathways: a question of balance. Annu Rev Immunol. 2006;24:257–286. doi:10.1146/annurev.immunol.24.021605.090700
22. Valero-Pacheco N, Beaulieu AM. Transcriptional regulation of mouse tissue-resident natural killer cell development. Front Immunol. 2020;11:309. doi:10.3389/fimmu.2020.00309
23. Gabrielli S, Sun M, Bell A, et al. Murine thymic NK cells are distinct from ILC1s and have unique transcription factor requirements. Eur J Immunol. 2017;47(5):800–805. doi:10.1002/eji.201646871
24. Ribeiro VSG, Hasan M, Wilson A, et al. Cutting edge: thymic NK cells develop independently from T cell precursors. J Immunol. 2010;185(9):4993–4997. doi:10.4049/jimmunol.1002273
25. Veinotte LL, Halim TYF, Takei F. Unique subset of natural killer cells develops from progenitors in lymph node. Blood. 2008;111(8):4201–4208. doi:10.1182/blood-2007-04-087577
26. Veinotte LL, Greenwood CP, Mohammadi N, et al. Expression of rearranged TCRγ genes in natural killer cells suggests a minor thymus-dependent pathway of lineage commitment. Blood. 2006;107(7):2673–2679. doi:10.1182/blood-2005-07-2797
27. Nozad Charoudeh H, Tang Y, Cheng M, et al. Identification of an NK/T cell-restricted progenitor in adult bone marrow contributing to bone marrow- and thymic-dependent NK cells. Blood. 2010;116(2):183–192. doi:10.1182/blood-2009-10-247130
28. Van De Walle I, Dolens AC, Durinck K, et al. GATA3 induces human T-cell commitment by restraining Notch activity and repressing NK-cell fate. Nat Commun. 2016;7:11171. doi:10.1038/ncomms11171
29. Stewart CA, Walzer T, Robbins SH, et al. Germ-line and rearranged Tcrd transcription distinguish bona fide NK cells and NK-like γδ T cells. Eur J Immunol. 2007;37(6):1442–1452. doi:10.1002/eji.200737354
30. Gerstner S, Köhler W, Heidkamp G, et al. Specific phenotype and function of CD56-expressing innate immune cell subsets in human thymus. J Leukoc Biol. 2016;100(6):1297–1310. doi:10.1189/jlb.1A0116-038R
31. Krabbendam L, Bernink JH, Spits H. Innate lymphoid cells: from helper to killer. Curr Opin Immunol. 2021;68:28–33. doi:10.1016/j.coi.2020.08.007
32. Seillet C, Brossay L, Vivier E. Natural killers or ILC1s? That is the question. Curr Opin Immunol. 2021;68:48–53. doi:10.1016/j.coi.2020.08.009
33. Shin SB, McNagny KM. ILC-you in the thymus: a fresh look at innate lymphoid cell development. Front Immunol. 2021;12:681110. doi:10.3389/fimmu.2021.681110
34. Spits H, Cupedo T. Innate lymphoid cells: emerging insights in development, lineage relationships, and function. Annu Rev Immunol. 2012;30:647–675. doi:10.1146/annurev-immunol-020711-075053
35. Vivier E, Artis D, Colonna M, et al. Innate Lymphoid Cells: 10 Years On. Cell. 2018;174(5):1054–1066. doi:10.1016/j.cell.2018.07.017
36. Krijgsman D, Hokland M, Kuppen PJK. The role of natural killer T cells in cancer-A phenotypical and functional approach. Front Immunol. 2018;9:367. doi:10.3389/fimmu.2018.00367
37. Dhodapkar MV, Kumar V. Type II Natural Killer T (NKT) cells and their emerging role in health and disease. J Immunol. 2017;198(3):1015–1021. doi:10.4049/jimmunol.1601399
38. Albano F, Vecchio E, Renna M, et al. Insights into thymus development and viral thymic infections. Viruses. 2019;11(9):836. doi:10.3390/v11090836
39. Kaszubowska L, Foerster J, Kwiatkowski P, et al. NKT-like cells reveal higher than T lymphocytes expression of cellular protective proteins HSP70 and SOD2 and comparably increased expression of SIRT1 in the oldest seniors. Folia Histochem Cytobiol. 2018;56(4):231–240. doi:10.5603/FHC.a2018.0025
40. Almeida JS, Couceiro P, López-Sejas N, et al. NKT-like (CD3+CD56+) cells in chronic myeloid leukemia patients treated with tyrosine kinase inhibitors. Front Immunol. 2019;10:2493. doi:10.3389/fimmu.2019.02493
41. Yu J, Mitsui T, Wei M, et al. NKp46 identifies an NKT cell subset susceptible to leukemic transformation in mouse and human. J Clin Invest. 2011;121(4):1456–1470. doi:10.1172/JCI43242
42. Seillet C, Huntington ND, Gangatirkar P, et al. Differential requirement for Nfil3 during NK cell development. J Immunol. 2014;192(6):2667–2676. doi:10.4049/jimmunol.1302605
43. Pistoia V, Tumino N, Vacca P, et al. Human γδ T-cells: from surface receptors to the therapy of high-risk leukemias. Front Immunol. 2018;9:984. doi:10.3389/fimmu.2018.00984
44. Parker ME, Ciofani M. Regulation of γδ T cell effector diversification in the thymus. Front Immunol. 2020;11:42. doi:10.3389/fimmu.2020.00042
45. Sánchez MJ, Spits H, Lanier LL, et al. Human natural killer cell committed thymocytes and their relation to the T cell lineage. J Exp Med. 1993;178(6):1857–1866. doi:10.1084/jem.178.6.1857
46. Li YR, Zhou K, Wilson M, et al. Mucosal-associated invariant T cells for cancer immunotherapy. Mol Ther. 2023;31(3):631–646. doi:10.1016/j.ymthe.2022.11.019
47. Godfrey DI, Koay HF, McCluskey J, et al. The biology and functional importance of MAIT cells. Nat Immunol. 2019;20(9):1110–1128. doi:10.1038/s41590-019-0444-8
48. De Smedt M, Hoebeke I, Reynvoet K, et al. Different thresholds of Notch signaling bias human precursor cells toward B-, NK-, monocytic/dendritic-, or T-cell lineage in thymus microenvironment. Blood. 2005;106(10):3498–3506. doi:10.1182/blood-2005-02-0496
49. Woo SY, Jung YJ, Ryu KH, et al. In vitro differentiation of natural killer T cells from human cord blood CD34 + cells. Br J Haematol. 2003;121(1):148–156. doi:10.1046/j.1365-2141.2003.04230.x
50. Ritz J, Campen TJ, Schmidt RE, et al. Analysis of T-cell receptor gene rearrangement and expression in human natural killer clones. Science. 1985;228(4707):1540–1543. doi:10.1126/science.2409597
51. Rodewald HR, Moingeon P, Lucich JL, et al. A population of early fetal thymocytes expressing FcγRII III contains precursors of T lymphocytes and natural killer cells. Cell. 1992;69(1):139–150. doi:10.1016/0092-8674(92)90125-V
52. Klein Wolterink RGJ, García-Ojeda ME, Vosshenrich CAJ, et al. The intrathymic crossroads of T and NK cell differentiation. Immunol Rev. 2010;238(1):126–137. doi:10.1111/j.1600-065X.2010.00960.x
53. Vaz F, Srour EF, Almeida-Porada G, et al. Human thymic stroma supports human natural killer (NK) cell development from immature progenitors. Cell Immunol. 1998;186(2):133–139. doi:10.1006/cimm.1998.1303
54. Michie AM, Carlyle JR, Schmitt TM, et al. Clonal characterization of a bipotent T Cell and NK cell progenitor in the mouse fetal thymus. J Immunol. 2000;164(4):1730–1733. doi:10.4049/jimmunol.164.4.1730
55. Hidalgo L, Martínez VG, Valencia J, et al. Expression of BMPRIA on human thymic NK cell precursors: role of BMP signaling in intrathymic NK cell development. Blood. 2012;119(8):1861–1871. doi:10.1182/blood-2011-07-370650
56. Cheng M, Nozad Charoudeh H, Brodin P, et al. Distinct and overlapping patterns of cytokine regulation of thymic and bone marrow-derived NK cell development. J Immunol. 2009;182(3):1460–1468. doi:10.4049/jimmunol.182.3.1460
57. Wagner JA, Rosario M, Romee R, et al. CD56bright NK cells exhibit potent antitumor responses following IL-15 priming. J Clin Invest. 2017;127(11):4042–4058. doi:10.1172/JCI90387
58. Björkström NK, Ljunggren HG, Michaëlsson J. Emerging insights into natural killer cells in human peripheral tissues. Nat Rev Immunol. 2016;16(5):310–320. doi:10.1038/nri.2016.34
59. Hansson M, Kärre K, Kiessling R, et al. Natural NK-cell targets in the mouse thymus: characteristics of the sensitive cell population. J Immunol. 1979;123(2):765–771. doi:10.4049/jimmunol.123.2.765
60. Garni-Wagner BA, Witte PL, Tutt MM, et al. Natural killer cells in the thymus. Studies in mice with severe combined immune deficiency. J Immunol. 1990;144(3):796–803. doi:10.4049/jimmunol.144.3.796
61. Schott E, Bonasio R, Ploegh HL. Elimination in vivo of developing T cells by natural killer cells. J Exp Med. 2003;198(8):1213–1224. doi:10.1084/jem.20030918
62. McNerney KO, Karageorgos SA, Hogarty MD, et al. Enhancing neuroblastoma immunotherapies by engaging iNKT and NK cells. Front Immunol. 2020;11:873. doi:10.3389/fimmu.2020.00873
63. Chu J, Gao F, Yan M, et al. Natural killer cells: a promising immunotherapy for cancer. J Transl Med. 2022;20(1):240. doi:10.1186/s12967-022-03437-0
64. Spel L, Schiepers A, Boes M. NFκB and MHC-1 Interplay in Neuroblastoma and Immunotherapy. Trends Cancer. 2018;4(11):715–717. doi:10.1016/j.trecan.2018.09.006
65. Pasqualini C, Kozaki T, Bruschi M, et al. Modeling the interaction between the microenvironment and tumor cells in brain tumors. Neuron. 2020;108(6):1025–1044. doi:10.1016/j.neuron.2020.09.018
66. Spel L, Boelens JJ, Van Der Steen DM, et al. Natural killer cells facilitate PRAME-specific T-cell reactivity against neuroblastoma. Oncotarget. 2015;6(34):35770–35781. doi:10.18632/oncotarget.5657
67. Custers L, Khabirova E, Coorens THH, et al. Somatic mutations and single-cell transcriptomes reveal the root of malignant rhabdoid tumours. Nat Commun. 2021;12(1):1407. doi:10.1038/s41467-021-21675-6
68. Yarmarkovich M, Maris JM. When cold is hot: immune checkpoint inhibition therapy for rhabdoid tumors. Cancer Cell. 2019;36(6):575–576. doi:10.1016/j.ccell.2019.11.006
69. Karahan ZS, Aras M, Sütlü T. TCR-NK cells: a novel source for adoptive immunotherapy of cancer. Turk J Hematol. 2023;40(1):1–10. doi:10.4274/tjh.galenos.2023.2022.0534
70. Lowe E, Truscott LC, De Oliveira SN. In vitro generation of human NK cells expressing chimeric antigen receptor through differentiation of gene- modified hematopoietic stem cells. Methods Mol Biol. 2016;1441:241–251.
71. Dolstra H, Roeven MWH, Spanholtz J, et al. Successful transfer of umbilical cord blood CD34+ hematopoietic stem and progenitor-derived NK cells in older acute myeloid leukemia patients. Clin Cancer Res. 2017;23(15):4107–4118. doi:10.1158/1078-0432.CCR-16-2981
72. Lehmann D, Spanholtz J, Osl M, et al. Ex vivo generated natural killer cells acquire typical natural killer receptors and display a cytotoxic gene expression profile similar to peripheral blood natural killer cells. Stem Cells Dev. 2012;21(16):2926–2938. doi:10.1089/scd.2011.0659
73. Spanholtz J, Preijers F, Tordoir M, et al. Clinical-grade generation of active NK cells from cord blood hematopoietic progenitor cells for immunotherapy using a closed-system culture process. PLoS One. 2011;6(6):e20740. doi:10.1371/journal.pone.0020740
74. Li YR, Dunn ZS, Yu Y, et al. Advancing cell-based cancer immunotherapy through stem cell engineering. Cell Stem Cell. 2023;30(5):592–610. doi:10.1016/j.stem.2023.02.009
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.