Back to Journals » Drug Design, Development and Therapy » Volume 15
The Volatile Oil of Zanthoxylum bungeanum Pericarp Improved the Hypothalamic-Pituitary-Adrenal Axis and Gut Microbiota to Attenuate Chronic Unpredictable Stress-Induced Anxiety Behavior in Rats
Authors Wei D, Zhao Y, Zhang M, Zhu L, Wang L, Yuan X, Wu C ![]()
Received 11 September 2020
Accepted for publication 30 December 2020
Published 22 February 2021 Volume 2021:15 Pages 769—786
DOI https://doi.org/10.2147/DDDT.S281575
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Jianbo Sun
Daneng Wei,1,* Yafei Zhao,2,* Mengmeng Zhang,1 Lei Zhu,1 Li Wang,1 Xing Yuan,3 Chunjie Wu1
1Pharmacy College, Chengdu University of Traditional Chinese Medicine, Chengdu, Sichuan 611137, People’s Republic of China; 2Acupuncture and Moxibustion College, Chengdu University of Traditional Chinese Medicine, Chengdu, Sichuan 610075, People’s Republic of China; 3School of Medical and Life Sciences, Chengdu University of Traditional Chinese Medicine, Chengdu, Sichuan 611137, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xing Yuan
School of Medical and Life Sciences, Chengdu University of Traditional Chinese Medicine, No. 1166 Liutai Avenue, Wenjiang District, Chengdu, Sichuan 611137, People’s Republic of China
Email [email protected]
Chunjie Wu
Pharmacy College, Chengdu University of Traditional Chinese Medicine, No. 1166 Liutai Avenue, Wenjiang District, Chengdu, Sichuan 611137, People’s Republic of China
Email [email protected]
Background: Anxiety disorders (ADs) are the most prevalent mental disorders worldwide. Stress-induced activation of the hypothalamic-pituitary-adrenal (HPA) axis and dysbiosis of gut microbiota seem to contribute to the onset of ADs. This study was designed to investigate the ameliorative effect of volatile oil of Zanthoxylum bungeanum (VOZB) on chronic unpredictable stress (CUS) induced anxiety behavior, as well as the altered HPA axis and gut microbiota.
Methods: Experimental rats were exposed to the CUS for 14 consecutive days. Meanwhile, VOZB was administered at doses of 50, 100 and 200 mg/kg/day for 14 days. The anxiety behavior was evaluated by elevated plus-maze (EPM) and open field (OF). The protein expressions and mRNA levels of corticotropin-releasing hormone (CRH) and glucocorticoid receptor (GR) in hypothalamus was determined, as well the hormone levels of HPA axis in serum. Furthermore, gut microbiota was detected by16S rRNA gene sequencing. The chemical constituents of VOZB were identified by GC-MS analysis.
Results: VOZB treatment (100 and 200 mg/kg/day) increased the ratio of open-arm entries and time in EPM test, as well as the central zone entries and time in OF test. Moreover, VOZB treatment reduced the protein expressions and mRNA levels of CRH, but elevated those of GR in hypothalamus. Similarly, the hormone levels of the HPA axis in serum were decreased by VOZB treatment. Besides, VOZB treatment restored the CUS-induced dysbiosis of gut microbiota, raising the Sobs and Chao indexes, inhibiting Lachnospiraceae, but facilitating Bacteroidales_S24-7_group, Lactobacillaceae, and Prevotellaceae. Additionally, Sobs and Chao indexes were negatively correlated to the serum corticosterone and CRH levels.
Conclusion: VOZB showed an ameliorative effect on CUS-induced anxiety behavior, potentially via inhibiting activation of the HPA axis and restoring the dysbiosis of gut microbiota, thus improving the stress-induced abnormality of the microbiota-gut-brain axis.
Keywords: Zanthoxylum bungeanum, anxiety disorders, stress, neuroendocrine, hypothalamic-pituitary-adrenal axis, gut microbiota
Introduction
Anxiety disorders (ADs), including generalized anxiety disorder, panic disorder, social anxiety disorder, among others, are the most prevalent mental disorders, seriously impairing the life quality of millions of individuals worldwide. The lifetime prevalence estimate of ADs was even higher than that of depression and occupied the first position in mental disorders according to the WHO World Mental Health Survey in 2009.1 The prevalence estimate of anxiety disorder varyies from 3.8% to 25% in the Western world.2 The increasing number of people with ADs implies raised medical costs, decreased work productivity, and impaired social relationships.3–5 Another finding from the Global Burden of Disease Study 2010 estimated that ADs contributed to 26.8 million disability-adjusted life years and substance abuse.6 Moreover, the persistence of ADs was considered as a risk factor for the development of other mental disorders. These comorbidities of ADs in turn brought new challenges for anti-anxiety treatment, resulting in low remission rates, poor prognosis, and increased suicide risk.7 Therefore, the exploration of potential pathogenesis and treatment of ADs has attracted increasing interest in the mental health field.
Compelling evidence from the clinic and laboratory indicated that not only the imbalance of central neurotransmitters, such as noradrenaline and serotonin, but also the abnormality of neuroendocrine hormones were involved in the occurrence and progression of ADs.8,9 The hypothalamic-pituitary-adrenal (HPA) axis, which is the core component of neuroendocrine networks, was reported to participate in the regulation of psychological and physical defense behaviors against the various stressors.10,11 Newly intensive efforts revealed that the hormone dysregulation of the HPA axis was responsible for the onset of ADs associated with chronic stress, including corticotropin-releasing hormone (CRH), adrenocorticotropic hormone (ACTH), and cortisol, known as corticosterone (CORT).12,13 Stress exposure beyond a threshold provoked the secretion of hypothalamic CRH, and then activated the HPA axis, thus triggering anxiety response. Restoring the balance of the HPA axis, in particular CRH, was an underlying way to improve the ADs, although the attempts were accompanied with a risk of failure.
Gut microbiota that inhabits the gastrointestinal tract of all mammals, plays an important role in a host’s biological processes, such as immunity, metabolism, digestion, and nutrition. Accumulating studies have revealed that gut microbiota also participate in the regulation of brain function and behavior through the gut-brain axis, which integrates immunological, neural, endocrine, and metabolic pathways. The alteration of gut microbiota was involved in a range of psychiatric diseases, including ADs. Germ-free (GF) mice exhibited less anxiety behavior than specific pathogen-free (SPF) mice,14–16 and SPF mice exposed to chronic restraint stress showed increased anxiety behavior compared with GF mice.17 Moreover, the transplantation of fecal microbiota derived from patients with depression induced anxiety and depressive behaviors in GF mice, rather than colonization with healthy microbiota derived from healthy individuals.18 The underlying mechanism whereby gut microbiota regulate the host’s mood and behavior mainly involves immune responses, vagus nerve, endocrine signaling, short chain fatty acids, and tryptophan metabolism.19,20 Recently, emerging studies suggested that the regulation of gut microbiota on host behaviors was associated with the HPA axis. In stress-related psychiatric disorders, including ADs, the disturbance of gut microbiota often contributed to the abnormality of HPA axis. For instance, GF mice showed higher levels of serum ACTH and CORT after acute stress compared with SPF mice.21,22 In addition, the levels of plasma ACTH and CORT were decreased by Bifidobacterium infantis, but increased by Escherichia coli, implying the important role of gut microbiota in HPA axis alterations.22,23 Reshaping gut microbiota has become a potential therapy for psychiatric disorders, especially the stress-related ADs.
Zanthoxylum bungeanum Maxim belongs to the Rutaceae family, and is widely distributed in East Asia. In China, Z. bungeanum pericarp is used as a popular spice in cooking because of its unique aroma and taste. Additionally, Z. bungeanum pericarp is also used as an important traditional Chinese herbal medicine to relieve pain, prevent itching, aid digestion, and stop diarrhea.24,25 Pharmacological investigations showed that Z. bungeanum shows various effects on the digestive system, nervous system, and circulatory system, mainly including anti-inflammatory, analgesic, gastrointestinal-promoting, anti-tumor, antibacterial, and neurotrophic effects.24 In recent years, emerging evidence suggested that Z. bungeanum extracts were beneficial for alleviating neurological and mental disorders, such as memory impairment and Alzheimer’s disease.26–28 Further identification of these neuroactive extracts found that unsaturated alkylamides and terpenoids (enriched in volatile oil) contributed to the neuropharmacological activities of Z. bungeanum. However, studies were rarely focused on evaluating the effect of Z. bungeanum extracts on stress-related ADs and their anti-anxiety mechanisms, although ADs bring about similar health damage to depression.
Therefore, we designed this study to investigate the ameliorative effect of volatile oil of Z. bungeanum (VOZB), known as the characteristic flavor compounds, on chronic unpredictable stress (CUS) induced anxiety behavior in rats. More importantly, we explored the changes in the HPA axis and gut microbiota to determine whether the regulatory effects of gut microbiota on the HPA axis were involved in the anxiolytic effect of VOZB.
Materials and Methods
Plant Materials and Extracts Preparation
Z. bungeanum pericarps were purchased from Chengdu Kangmei Pharmaceutical Co. Ltd (Chengdu, China) and identified by Professor Chun-Jie Wu (Chengdu University of Traditional Chinese Medicine, Chengdu, P.R. China). The voucher specimen was deposited at our laboratory (No. HJ201814). The dried pericarps of Z. bungeanum were powered and screened through a 65 mesh sieve. The powder was subjected to hydrodistillation for 5 h using a Clevenger type apparatus. The obtained volatile oils were dried using hydrous sodium sulfate, filtered, and then stored at 4°C. The extraction yield of VOZB was calculated as 5.12% (w/w). The density of VOZB was 0.8563 g/mL according to the exam.
Oral Toxicity Test
The oral toxicity test was conceptualized according to OECD guidelines for the Testing of Chemicals (Test No. 425, 2008). Female Sprague-Dawley rats were chosen for testing the toxicity of VOZB. The testing dosages were 55 mg/kg, 175 mg/kg, 550 mg/kg, and 2000 mg/kg according to the guidelines (doses were spaced by a factor of 0.5 on a log dose scale). The different doses of VOZB were uniformly suspended in 1% sodium carboxymethyl cellulose (CMC-Na) solution and orally administered to treated rats after fasting overnight under acute toxicity conditions. Then the mortality of the experimental rats was recorded within a 72 h monitoring period. Our results indicated that neither death nor any abnormal neurobehaviors could be observed with the doses ranging from 55 to 2000 mg/kg during 72 h. Therefore, the 50% lethal dose was greater than 2000 mg/kg.
Animals
Male Sprague-Dawley rats (200±10 g, 6–8 weeks) were purchased from the Sichuan Dashuo Experimental Animal Co. Ltd (Sichuan, China) and housed 2 per cage (34×26×19 cm) in controlled environmental conditions (temperature 23±2°C, humidity 55–65%, and 12/12 h light-dark cycle). Rats were allowed access to food and water ad libitum and habituated for 7 days in the laboratory before starting the chronic unpredictable stress (CUS) procedure. All experimental procedures were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and approved by the Animal Care and Use Committee of Chengdu University of Traditional Chinese Medicine.
Experimental Design
A total of 50 rats were randomly divided into five groups (n = 10), including the normal group, model group, and three treatment groups. The experimental protocol was designed according to the previous studies with some modifications,29–31 and the experimental schematic diagram is shown in Figure 1. Briefly, experimental rats in the model group and the three treatment groups were exposed to the CUS for 14 consecutive days. Meanwhile, different doses (50, 100, and 200 mg/kg/day, respectively) of VOZB were administered to rats in the three treatment groups by an intragastric gavage for 14 days, previously suspended in vehicle (1% CMC-Na). Rats in other groups were administered with the vehicle (1% CMC-Na, 10 mL/kg). Since there is no information on the dose of VOZB in the treatment of anxiety either in clinical or preclinical studies, the dose of VOZB in this study was determined by our preliminary experiment. The behavioral tests were conducted 2 h after the final drug administration. On the next day, all rats were anesthetized by 1% pentobarbital sodium (3 mL/kg). The inguinal skin was cut open to expose the femoral vein. Then blood samples were collected from the femoral vein and centrifuged for 10 min at 5000 rpm to prepare the serum samples. After blood sampling, the rats were killed by cervical dislocation and the brain samples were removed quickly from the skull. Some of the brain samples were immersed and fixed in a 4% paraformaldehyde (PFA) solution for immunohistochemical assay, and others were had the hypothalamus isolated for the next process. The serum samples and hypothalamus samples were stored at −80°C. In addition, the abdomen of rats was sterilized by iodophor after barbering on a super clean bench. The whole colon was removed and cut open lengthwise. The stools in colon were collected into sterile tubes and immediately frozen in liquid nitrogen for detection of gut microbiota.
 |
Figure 1 Schematic diagram illustrating the major steps of the experiment. The experimental rats were exposed to the CUS for 14 consecutive days. Meanwhile, different doses (50, 100, and 200 mg/kg/day, respectively) of VOZB were administered for 14 days. Abbreviations: CUS, chronic unpredictable stress; VOZB, volatile oil of Zanthoxylum bungeanum. |
Chronic Unpredictable Stress Procedure
The CUS procedure was modified referring to the published procedures.29–31 After 7 days of habituation, rats in model group and three VOZB treatment groups were housed individually and exposed to the following stressors for 14 days: wet bedding for 12 hours; 30-min restraint in a restraining device which was made of plexiglass and flexible nylon immobilization belts, thus restricting movement but allowing free respiration and air circulation; 12-h food and water deprivation (19:00–7:00); 10-min forced swim at 18°C in a cylindrical tank (60 cm height × 30 cm diameter) filled with fresh water to a 30 cm depth; reversing the light and dark cycle; 10-min tail pinch with a clothes-pin 1 cm from the base of the tail after being restrained in previous device; 15-min electric footshock (1.5mA, 30 s on and 150 s off). In order to maximize the unpredictability of stressors, all stressors were applied in a random order and at varying times, and the same stressors were not applied on two consecutive days. The CUS experimental schedule is outlined in Table 1. After daily CUS procedure, rats were placed in their home cages with fresh bedding and returned to the housing room until the next stress session. The normal group rats were handled daily for 2 min in the housing room, but not stressed.
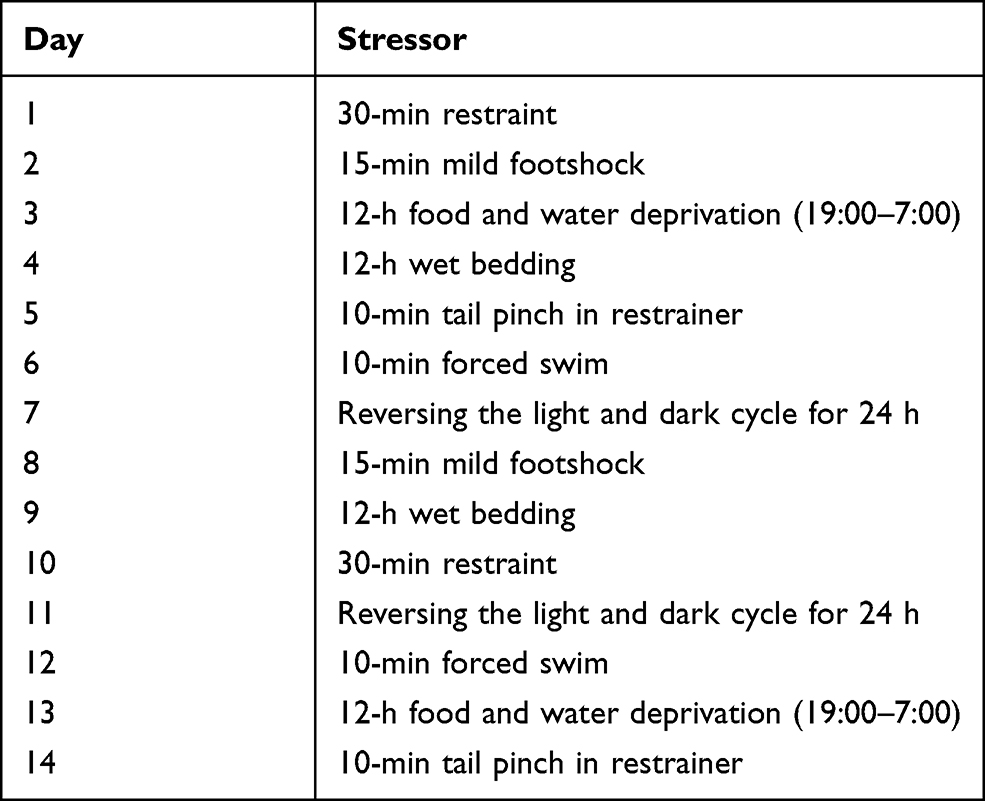 |
Table 1 Chronic Unpredictable Stress Experimental Schedule |
The Elevated Plus-Maze Test
The elevated plus-maze (EPM) test was carried out to evaluate the anxiety-like behavior after the final drug administration. Rats were transported to the well-ventilated testing room and allowed to acclimate for 30 min. The testing room was illuminated with a red lamp (10 and 25 lux) and kept quiet throughout the behavior test. The EPM (TECHMAN Co., Ltd, China) consisted of two opposite open-arms (50×10×1.5 cm), two opposite enclosed-arms (50×10×40 cm) and a center platform (10×10 cm) connecting four arms in the middle. The maze was elevated to a height of 60 cm above the floor and exposed to a fluorescent lamp suspended at height of 100 cm above the apparatus. At the beginning of the EPM test, the rat was placed in the center platform facing one of the open arms, and then allowed to explore freely for 5 min. The number of entries and time spent in each of the four arms was recorded by an automated video-tracking system (EthovisionXT9, NOLDUS). The proportion of entries and time spent in the open-arms relative to the total entries or time spent in all arms (open/open + closed) were calculated as indices of open-arm exploration. The fewer entries and less time spent in the open arms indicated a higher anxiety state.29,32 After each test, the apparatus was thoroughly cleaned with 30% ethanol solution and dried with paper towels to avoid olfactory trace of the previous animal.
The Open Field Test
The open field (OF) apparatus (TECHMAN Co., Ltd, China) was a cubic chamber (100×100×40 cm), consisting of four walls and a bottom. The bottom of the open field was divided into a central zone (40×40 cm) and a peripheral zone. Rats were placed individually at the center of the central zone and allowed to explore freely for 5 min. The locomotion of rats in OF was recorded by the automated video-tracking system (EthovisionXT9, NOLDUS) at the same time. The number of visits in the central zone was scored, as well as the time spent in the central zone. All four paws entering the central zone was considered as the criterion of center visiting. Between the interval of two tests, the OF was cleaned with 30% ethanol solution and dried with paper towels to eliminate the residual odor.
Immunohistochemistry
The immunohistochemical assay was carried out using the method described in previous studies.27 Briefly, the brain samples post-fixed with 4% PFA were coronally cut into 5 μm hypothalamus sections, which contain paraventricular nucleus. The brain sections were dewaxed and incubated in 3% hydrogen peroxide for 10 min, and then boiled in citrate buffer (pH 6.0) for 5 min. After blocking with goat serum for 30 min, the sections were incubated with anti-CRH antibody (dilution 1: 100, Abcam, Britain) and anti-glucocorticoid receptor (GR) antibody (dilution 1: 100, Bioss, China) overnight at 4°C. Subsequently, the secondary antibody was incubated at 37°C for 30 min, and specific labeling was visualized using a 3,3ʹ-diaminobenzidine (DAB) kit. Images were captured via a digital microscope (BA200 Digital, McAudi) and analyzed by Image-Pro Plus 6.0 software (Media Cybernetics, Silver Spring, USA). The CRH and GR expression in the hypothalamus were measured by the average optical density (AOD) to compare the potential differences between groups.
Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) Assay
Total RNA of hypothalamus samples was extracted using Trizol reagent (Invitrogen, USA) according to the manufacturer’s instructions. The ratio of optical density at 260 nm and 280 nm (OD 260/OD 280) was limited in the range of 1.8–2.1 to ensure the purity and quality of RNA. Then 2 μg of total RNA was reverse transcribed into cDNA using the TUREscript 1st Stand cDNA Synthesis Kit (Aidlab Biotechnologies, China). After adding the SYBR Green PCR Master Mix (TaKaRa Biotechnology, China), RT-qPCR was performed using the ABI StepOnePlus System (Applied Biosystems, USA) following the conditions: 95°C for 10 min, followed by 40 cycles of 95°C for 15 s, 60°C for 60 s, and 72°C for 60 s. The target mRNA primers were designed by Premier 5.0 and synthesized by Sangon Biotech (Shanghai, China). The primer sequences for GRfwd were as follows: 5ʹ-CACATCTCACCCGCACCGATTG-3ʹ, rev: 5ʹ-TTGGACAAACACGGATGCCTGAC-3ʹ; for CRHfwd: 5ʹ-TGCCAAGGGAGGA GAAGAGA GC-3ʹ, rev: 5ʹ-GACAGAGCCACCAGCAGCATG-3ʹ; for GAPDHfwd: 5ʹ-CTGGAGAAACCTGCCAAGTATG-3ʹ, rev: 5ʹ-GGTGGAAGAATGGGAGTTG CT-3ʹ; and GAPDH was used as an endogenous control. The gene expression levels of CRH and GR were calculated with the method of 2−ΔΔCT.
Enzyme-Linked Immunosorbent Assay (ELISA)
The levels of ACTH and CORT in the serum mentioned above were detected by commercial ELISA kits following the corresponding manufacturer’s protocol. All of the reagents were prepared at room temperature and the thawed serum samples were centrifuged again before the assay.
Microbial Analysis
The microbial analysis was performed according to previous studies.33,34 Briefly, bacterial DNA was extracted from fecal samples using the QIAamp DNA Stool Mini Kit (QIAGEN, Germany) following the manufacturer’s protocols. DNA integrity was assessed by electrophoresis on a 1% agarose gel containing 0.5 mg/mL ethidium bromide. Then, the V3-V4 region of the 16S rRNA gene was amplified by PCR using the universal bacterial primers (338F: 5ʹ-ACTCCTACGGGAGGCAGCAG-3ʹ and 806R: 5ʹ-GGACTACHVGGGTWTCTAAT-3ʹ). Further, amplicon sequencing was performed on an Illumina MiSeq platform (PE300). Raw reads were demultiplexed and filtered by QIIME (version 1.9.1) following the read length and quality, and the filtered reads were dereplicated to unique sequences. Operational units (OTUs) were clustered with 97% similarity cutoff using UPARSE (version 7.1) and chimeric sequences were identified and removed using UCHIME. Taxonomy of each OTU representative sequence was analyzed by Ribosomal Database Project Classifier (version 2.11) against the silva 16S rRNA database. The diversity index and composition of the gut microbiota were determined by bioinformatic analysis.
Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
The GC-MS analysis was performed according to our previous study.35 Chromatography was performed on a Shimadzu GC-MS-TQ8040 gas chromatograph with a HP-1510 gas chromatography headspace heater and a mass-selective detector with electron impact ionization. Samples were added to a headspace vial and incubated for 30 min at a temperature of 50°C in the headspace heater. Subsequently, the fiber was inserted in the headspace vial and extracted for 10 min at 50°C. After extraction, the SPME fiber was exposed to the hot GC injection port at 230°C for 3 min for desorption of aroma components. For the GC-MS analysis, the analytes were separated using an HP-5MS capillary column (30 m × 0.25 mm, 0.25 μm). The temperature program used for analysis was as follows: the initial temperature was 50°C for 2 min, which was increased to 200°C at 4°C/min; 200°C was maintained for 5 min; then the temperature was increased to 240°C at 15°C/min and maintained for 5 min. Helium (99.999%) was the carrier gas maintained at a flow rate of 1 mL/min. The split rate was 36:1 and inlet volume was 1.0 μL. The electron impact ionization conditions were ion energy 70eV and the mass range scanned was 50–550 a.m.u in the full-scan acquisition mode. Compounds were identified using the NIST Mass Spectral Search Program (National Institute of Standards and Technology, Washington, DC, USA). The relative amounts of individual components of the fatty oils were expressed as percentages of the peak area relative to the total peak area.
Statistical Analysis
Data are presented as the mean ± standard deviation (SD) and analyzed using SPSS software version 19.0 (IBM Corporation, USA). The Shapiro–Wilk test was used to test the normal distribution of observed indicators. The indicators obtained from the EPM test, OF test, and α diversity analysis of gut microbiota, as well as the hormone levels showed normal distribution. Therefore, statistical difference between the groups was estimated by one‑way analysis of variance (ANOVA) followed by Fisher’s LSD test.27 Moreover, the β diversity of gut microbiota was analyzed by principal coordinates analysis (PCoA) and nonmetric multidimensional scaling analysis (NMDS). The relative abundance of gut microbiota which was not in accordance with normal distribution was compared using Kruskal–Wallis test (a nonparametric test). Besides, the correlation coefficient of α diversity indexes of gut microbiota and serum hormone levels was estimated by Pearson correlation test. p < 0.05 was considered statistically significant.
Results
The VOZB Alleviated the CUS Induced Anxiety Behavior in Rats
EPM and OF tests were used to evaluate the effect of VOZB on the CUS induced anxiety behaviors in rats. In the EPM test, one-way ANOVA revealed significant differences between groups for the ratio of open-arm entries (ROE) (F(4,45)= 20.025; p = 0.0000) and the ratio of open-arm time (ROT) (F(4,45)= 13.876; p = 0.0000). Particularly, the ROE in the CUS group was significantly fewer than that of the normal group (p = 0.0000, Figure 2A), as well as the ROT (p = 0.0000, Figure 2B). However, the ROE were elevated by the treatment with low-dose (50 mg/kg/day), moderate-dose (100 mg/kg/day), and high-dose (200 mg/kg/day) of VOZB in a dose-dependent manner vs the model group (p = 0.0003, 0.0000, and 0.0000, respectively, Figure 2A), the elevation of ROT was also observed in the moderate-dose group and high-dose group vs the model group (p = 0.0000 and 0.0000, respectively, Figure 2B). In the OF test, one-way ANOVA revealed significant differences between groups for the entries in the central zone (F(4,45)= 7.013; p = 0.0002). The entries in the central zone of the CUS group were significantly fewer than those of the normal group (p = 0.0002, Figure 2C), as well as the time spent in the central zone (p = 0.0137, Figure 2D). The entries in the central zone were significantly increased by the treatment with the moderate-dose and high-dose of VOZB (p = 0.0115 and 0.0002, respectively, Figure 2C and D). Collectively, these behavioral results indicated that the VOZB treatment alleviated the CUS-induced anxiety behavior in rats. The exposure of CUS decreased the ROE and ROT of model rats in the EPM test, as well the central zone entries and cumulative time in OF test. These behavioral test results were consistent with those proposed in other literature,29,36 indicating that the modified CUS procedures successfully and reliably induced anxiety behavior in rats. However, the VOZB significantly alleviated the CUS-induced anxiety behavior in the EPM and OF tests.
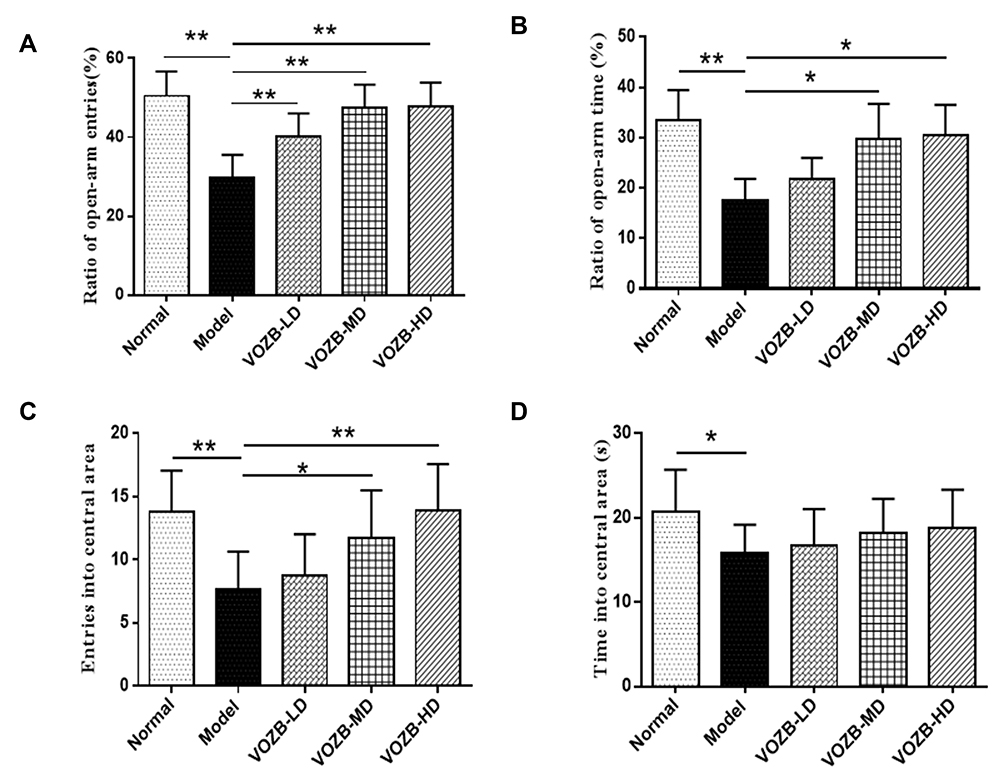 |
Figure 2 The VOZB alleviated the CUS-induced anxiety behavior in the elevated plus maze (EPM) and open field (OF) tests. Ratio of open-arm entries (A) and ratio of open-arm time (B) in the EPM test were evaluated, as well as the entries into central area (C) and accumulative time spent in central area (D) in the OF test. Data obtained from the EPM and OF test was in accordance with normal distribution and analyzed by one-way ANOVA followed by Fisher’s LSD test. Data were expressed as the mean ± SD (n = 10), *p<0.05 and **p<0.01 vs the model group. Abbreviations: VOZB-LD, the low dose of VOZB; VOZB-MD, the moderate dose of VOZB; VOZB-HD, the high dose of VOZB. |
The VOZB Regulated the Protein Expressions and mRNA Levels of CRH in the Hypothalamus
Hypothalamic CRH is considered to be a vital neuroendocrine factor involved in the stress response and emotion regulation. Therefore, we investigated the CRH expressions of hypothalamus by immunohistochemistry and RT-qPCR. The immunohistochemical assays showed that CRH appearing yellow or brownish yellow staining, was mainly expressed in the paraventricular nucleus cells and enriched in the cytoplasmic fractions (Figure 3A). The number of hypothalamic CRH immuno-positive cells in the CUS group was increased visibly compared with that in the normal group. However, the treatment with different doses of VOZB decreased the hypothalamic CRH immuno-positive cells of the CUS induced rats. Further, one-way ANOVA revealed significant differences between groups for the AOD of hypothalamic CRH expression (F(4,20)= 3.831; p = 0.0146). The CUS markedly raised the AOD of CRH expressions in the hypothalamus (p = 0.0111, Figure 3A), while treatments with the moderate-dose and high-dose VOZB successfully reversed these raising effects induced by CUS (p = 0.0163 and 0.0147, respectively, Figure 3A). Additionally, the results of the immunohistochemistry were confirmed by subsequent RT-qPCR. One-way ANOVA revealed significant differences between groups for the hypothalamic CRH mRNA levels (F(4, 20)= 70.438; p = 0.0000). The levels of CRH mRNA in hypothalamus were significantly elevated by CUS (p = 0.0000), whereas the VOZB inhibited the elevation of CRH mRNA levels at the doses of 100 and 200 mg/kg/day (p = 0.0001 and 0.0000, respectively, Figure 4A). These results suggested that the VOZB reduced the expression of CRH and its mRNA in the hypothalamus, thus relieving the CUS induced anxiety behavior in rats.
 |
Figure 3 The VOZB regulated the protein expressions of CRH and GR in the hypothalamus. Representative photomicrographs and AOD values of CRH (A) and GR (B) in the paraventricular nucleus were shown respectively. The red arrows were used to mark the CRH positive cells. Data obtained from the EPM and OF test was in accordance with normal distribution and analyzed by one‑way ANOVA followed by Fisher’s LSD test. The AOD values of CRH and GR expressions in the hypothalamus were in accordance with normal distribution and analyzed by one‑way ANOVA followed by Fisher’s LSD test. Data were expressed as mean ± SD (n = 5), *p<0.05 vs the model group. Abbreviations: CRH, corticotropin-releasing hormone; GR, glucocorticoid receptor; VOZB-LD, the low dose of VOZB; VOZB-MD, the moderate dose of VOZB; VOZB-HD, the high dose of VOZB. |
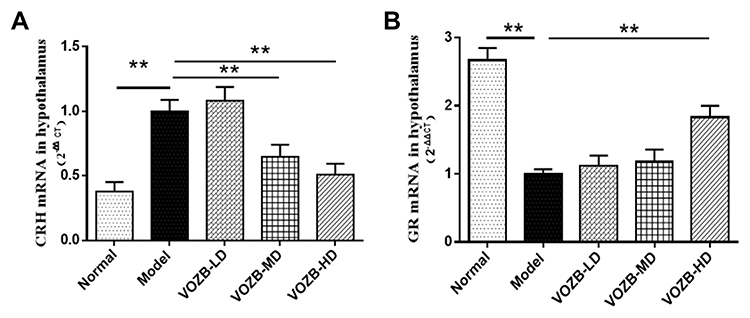 |
Figure 4 The VOZB improved the mRNA levels of CRH and GR in the hypothalamus. Levels of CRH mRNA (A) and GR mRNA (B) were respectively determined by RT-qPCR. The levels of CRH mRNA and GR mRNA in the hypothalamus were in accordance with normal distribution and analyzed by one‑way ANOVA followed by Fisher’s LSD test. Data were expressed as mean ± SD (n = 5), **p<0.01 vs the model group. Abbreviations: CRH, corticotropin-releasing hormone; GR, glucocorticoid receptor; VOZB-LD, the low dose of VOZB; VOZB-MD, the moderate dose of VOZB; VOZB-HD, the high dose of VOZB. |
The VOZB Elevated the Protein Expressions and mRNA Levels of GR in the Hypothalamus
Hypothalamic GR mediates the negative feedback effect of corticosterone on the CRH secretion, thus regulating the activity of the HPA axis. In this study, we also assessed the GR expression of hypothalamus by immunohistochemistry and RT-qPCR. The GR immuno-positive cells in the paraventricular nucleus showed a visible reduction in the CUS group compared with the normal group (Figure 3B). However, the treatment with VOZB partly increased the CUS-induced GR immuno-positive cells in the hypothalamus (Figure 3B). Moreover, one-way ANOVA revealed significant differences between groups for the AOD of hypothalamic GR expression (F(4,20) = 2.829; p = 0.0460). The AOD of hypothalamic GR expressions was markedly decreased in the CUS group rats compared with that in the normal group rats (p = 0.0143, Figure 3B), while the high-dose of VOZB partly reversed the CUS-induced reduction of GR expression (p = 0.056, Figure 3B). Similarly, one-way ANOVA revealed significant differences between groups for the hypothalamic GR mRNA levels (F(4, 20)= 101.634; p = 0.0000). The levels of hypothalamic GR mRNA were also significantly reduced in the CUS group (p = 0.0000, Figure 4B), whereas the VOZB attenuated the CUS-induced reduction of GR mRNA levels, especially at the dose of 200 mg/kg/day (p = 0.0000, Figure 4B). These findings suggested that elevating the protein expressions and mRNA levels of GR in the hypothalamus might be closely related to the effect of VOZB on improving the CUS-induced anxiety behavior in rats.
The VOZB Regulated the Levels of CRH, ACTH and CORT in Serum
Hormones secreted from the HPA axis in serum were also investigated by ELISA, including CRH, ACTH, and CORT. One-way ANOVA revealed significant differences between groups for CRH (F(4, 45)= 4.048; p = 0.0069), ACTH (F(4, 45) = 14.866; p = 0.0000), and CORT levels (F(4, 45) = 9.732; p = 0.0000) in serum. Consistent with the result of hypothalamic CRH expression, the CRH levels in serum were significantly raised in the model group compared with the normal group (p = 0.0021). However, the VOZB at the doses of 200 mg/kg/day reversed the CUS induced elevation of CRH levels (p = 0.0088, Figure 5A). Similarly, the levels of ACTH and CORT in serum were raised in the CUS-induced rats compared with the normal group rats (p = 0.0000 and 0.0000, respectively, Figure 5B and C), but the VOZB reduced the levels of ACTH at the doses of 100 and 200 mg/kg/day (p = 0.0000 and 0.0000, respectively, Figure 5B), as well as the CORT levels (p = 0.0183 and 0.0006, respectively, Figure 5C). Interestingly, the VOZB treatment showed a dose-dependent effect on the CUS-induced ACTH imbalance. The above results suggested that the VOZB reversed the imbalance of HPA-related hormones to improve the CUS-induced anxiety behavior in rats.
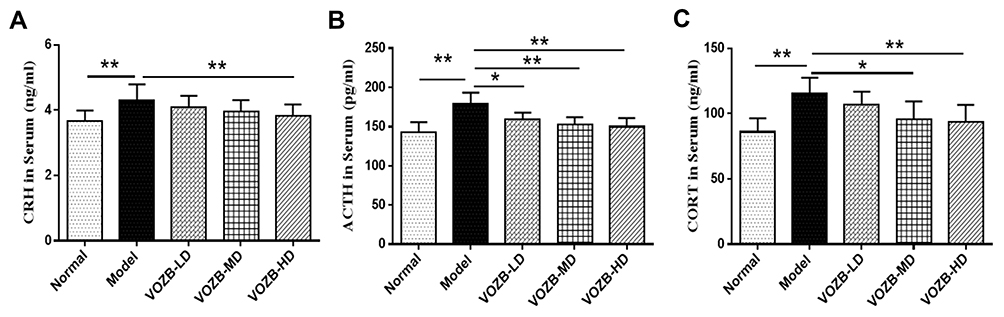 |
Figure 5 The VOZB regulated the levels of CRH, ACTH and CORT in serum. Levels of CRH (A), ACTH (B) and CORT (C) were respectively determined by ELISA. The levels of hormones in serum were in accordance with normal distribution and analyzed by one‑way ANOVA followed by Fisher’s LSD test. Data were expressed as mean ± SD (n = 10), *p<0.05 and **p<0.01 vs the model group. Abbreviations: CRH, corticotropin-releasing hormone; ACTH, adrenocorticotropic hormone; CORT, corticosterone; VOZB-LD, the low dose of VOZB; VOZB-MD, the moderate dose of VOZB; VOZB-HD, the high dose of VOZB |
The VOZB Restored the CUS-Induced Dysbiosis of Gut Microbiota
Gut microbiota has been confirmed to be involved in the regulation of host’s mood and behavior through the gut-brain axis. In this study, we detected the microbiota changes by 16S rRNA gene sequencing. The α diversity analysis of gut microbiota included the Sobs, Chao, Shannon and Simpson indexes. One-way ANOVA revealed significant differences between groups for the Sobs (F(2, 21)= 4.665; p= 0.0211) and Chao indexes (F(2, 21)= 5.310; p= 0.0136), but not Shannon and Simpson indexes. Sobs and Chao indexes reflecting community richness were obviously decreased in the model group compared with those in the normal group (p = 0.0116 and 0.0058, respectively, Figure 6A and B), but the VOZB at dose of 200 mg/kg/day significantly reversed the CUS-induced reduction of Sobs and Chao index (p = 0.0203 and 0.0211, respectively, Figure 6A and B). However, Shannon and Simpson indexes reflecting community diversity showed little change between groups (Figure 6C and D). Moreover, the PCoA showed that the cumulative percentages of PC1 and PC2 accounted for 64.41% of total variations. Most of the samples were clustered according to groups and distributed differently, indicating that the β diversity of gut microbiota was different between groups (R = 0.2361, p = 0.0020, Figure 6E). The VOZB treatment samples showed less distance to the normal samples than that to the model samples. The NMDS showed a more clear distribution difference between groups (R2 = 0.2819, p = 0.0010, Figure 6F).
 |
Figure 6 The VOZB affected the diversity of gut microbiota. The α diversity analysis of gut microbiota includes Sobs index (A), Chao index (B), Shannon index (C) and Simpson index (D). Sobs and Chao are community richness indexes, Shannon and Simpson were community diversity indexes. The β diversity of gut microbiota were also evaluated by PCoA (E) and NMDS (F). The distance between the different colored samples represents the similarity of gut microbiota, and a closer distance indicates higher similarity. The α diversity indexes (Sobs, Chao, Shannon, and Simpson) were in accordance with normal distribution and analyzed by one‑way ANOVA followed by Fisher’s LSD test. PCoA and NMDS were conducted according to weighted UniFrac distance metrics. All data were analyzed at genus level and the values were expressed as mean ± SD (n = 8), *p<0.05 and **p<0.01 vs the model group. Abbreviations: VOZB-HD, the high dose of VOZB; PCoA, principal coordinates analysis; NMDS, nonmetric multidimensional scaling. |
Taxonomy analysis of gut microbiota obtained 623 OTUs in total, including 582 in the normal group, 501 in the model group, and 584 in the VOZB group. Of these, 115 OTUs in the normal group and 111 OTUs in the VOZB group were different from those in the model group (Figure 7A). At the phylum level, Firmicutes, Bacteroidetes, Proteobacteria, and Deferribacteres were the dominant communities in all samples (Figure 7B and Supplementary Table 1). The proportion of Bacteroidetes, Proteobacteria, and Actinobacteria was decreased after the CUS exposure, whereas Firmicutes and Deferribacteres were significantly increased (p = 0.0357 and 0.0117, respectively, Figure 7B). The VOZB treatment at dose of 200 mg/kg/day partly improved the CUS-induced dysbiosis, decreasing Firmicutes and Deferribacteres, but increasing Bacteroidetes (Figure 7B and C). Further, the main OTUs identified at the family level are shown in Figure 7D. The results showed that Lachnospiraceae, Bacteroidales_S24-7_group, Lactobacillaceae, Bacteroidaceae, Ruminococcaceae, and Porphyromonadaceae were the dominant communities at the family level (Figure 7D and Supplementary Table 2). After the CUS exposure, the relative abundance of Lachnospiraceae and Ruminococcaceae were obviously increased (p = 0.0157 and 0.0118, respectively, Figure 7E), while the relative abundance of Bacteroidales_S24-7_group, Lactobacillaceae, and Prevotellaceae were significantly decreased (p = 0.0008, 0.0087, and 0.0167, respectively, Figure 7E). The VOZB treatment showed some benefit on restoring the CUS-induced dysbiosis, reducing Lachnospiraceae, but raising Bacteroidales_S24-7_group, Lactobacillaceae, and Prevotellaceae. Notably, the proportions of Lactobacillaceae and Prevotellaceae in the VOZB group were obviously more than those in the model group (p = 0.0063 and 0.0274, respectively, Figure 7E).
 |
Figure 7 The VOZB treatment restored the dysbiosis of gut microbiota. (A) The change of OTUs were identified. (B) The composition profiles of gut microbiota were analyzed at phylum level. (C) The dominant phyla were compared between the different treatment groups. (D) The composition profiles of gut microbiota were further analyzed at family level. (E) The dominant genera were also compared between the different treatment groups. (F) The correlation of α diversity indexes (Sobs and Chao) and the HPA axis hormones in serum were analyzed by Pearson correlation test. The relative abundance of dominant communities at the phylum and family level was not in accordance with normal distribution and compared using Kruskal–Wallis test (a nonparametric test). The relative abundance of gut microbiota on different taxonomic levels was expressed as mean ± SD (n = 8), *p<0.05 and **p<0.01 vs the model group. Abbreviations: VOZB-HD, the high dose of VOZB; OTUs, operational taxonomic units; HPA axis, hypothalamic-pituitary-adrenal axis. |
Additionally, Sobs and Chao indexes were negatively correlated to the serum CORT level (r = −0.62, p = 0.0012 and r = −0.63, p = 0.0009, respectively) and CRH level (r = −0.57, p = 0.0034 and r = −0.50, p = 0.0131, respectively), but not correlated with the serum ACTH level (Figure 7F).
Composition Analysis of the Volatile Oil of Z. bungeanum
The chemical compositions of VOZB were separated and identified by GC-MS analysis. In this study, we detected 30 compounds (peaks) from the total ion chromatograms of the VOZB within 31 min (Figure 8). These compounds were subsequently identified based on the mass spectrometric data interpretation reported in the literature, and their relative contents were also calculated by the peak area normalization method. The 30 identified compounds were α-Thujene (peak 1, 0.38%), α-Pinene (peak 2, 3.03%), Sabinene (peak 3, 10.27%), β-Pinene (peak 4, 0.38%), β-Myrcene (peak 5, 10.56%), α-Phellandrene (peak 6, 2.34%), α-Terpinene (peak 7, 0.13%), Limonene (peak 8, 28.88%), β-Phellandrene (peak 9, 12.91%), trans-β-Ocimene (peak 10, 4.54%), Ocimene (peak 11, 2.35%), Cineole (peak 12, 6.64%), γ-Terpinene (peak 13, 0.35%), Sabinene hydrate (peak 14, 0.22%), Terpinolene (peak 15, 0.46%), Linalool (peak 16, 1.00%), trans-Sabinene hydrate (peak 17, 0.12%), 3-Carene (peak 18, 8.91%), α-Terpineol (peak 19, 0.32%), Linalyl acetate (peak 20, 0.58%), 4-Terpinenyl acetate (peak 21, 0.51%), Piperitone (peak 22, 0.24%), β-Elemene (peak 23, 0.18%), Terpinyl acetate (peak 24, 1.60%), Caryophyllene (peak 25, 1.09%), α-Humulene (peak 26, 0.15%), Germacrene D (peak 27, 0.98%), Spathulenol (peak 28, 0.17%), γ-Cadinene (peak 29, 0.26%), and δ-Cadinene (peak 30, 0.45%), respectively (Figure 6 and Table 2). Of these 30 compounds, 13 were with a relative content more than 1% (Table 2), including Limonene, β-Phellandrene, myrcene, Sabinene, 3-Carene, Cineole, trans-β-Ocimene, α-Pinene, Ocimene, α-Phellandrene, Terpinyl acetate, Caryophyllene, and Linalool. From the present results, the VOZB mainly consisted of alkenes, such as Limonene, β-Phellandrene, Myrcene, Sabinene, and 3-Carene.
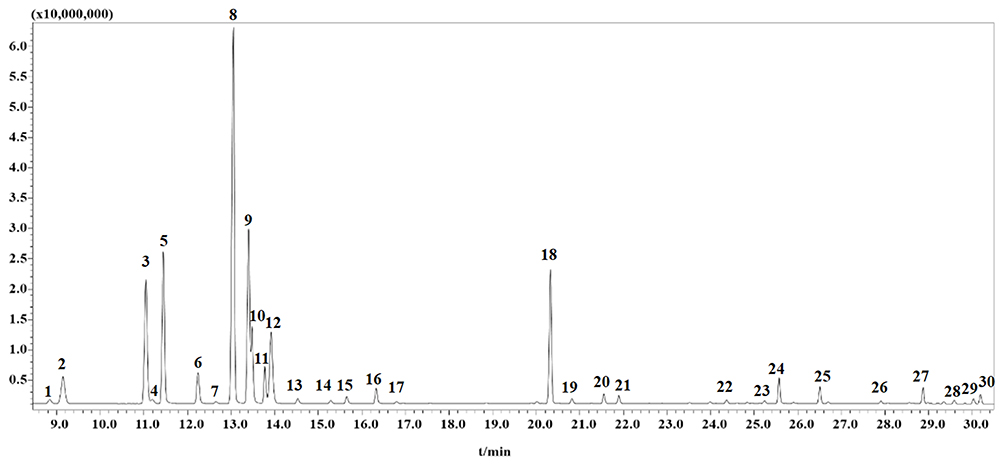 |
Figure 8 Composition analysis of the VOZB. A total of 30 peaks were detected by GC‑MS. Abbreviations: VOZB, the volatile oil of Zanthoxylum bungeanum; GC-MS, gas chromatography-mass spectrometry. |
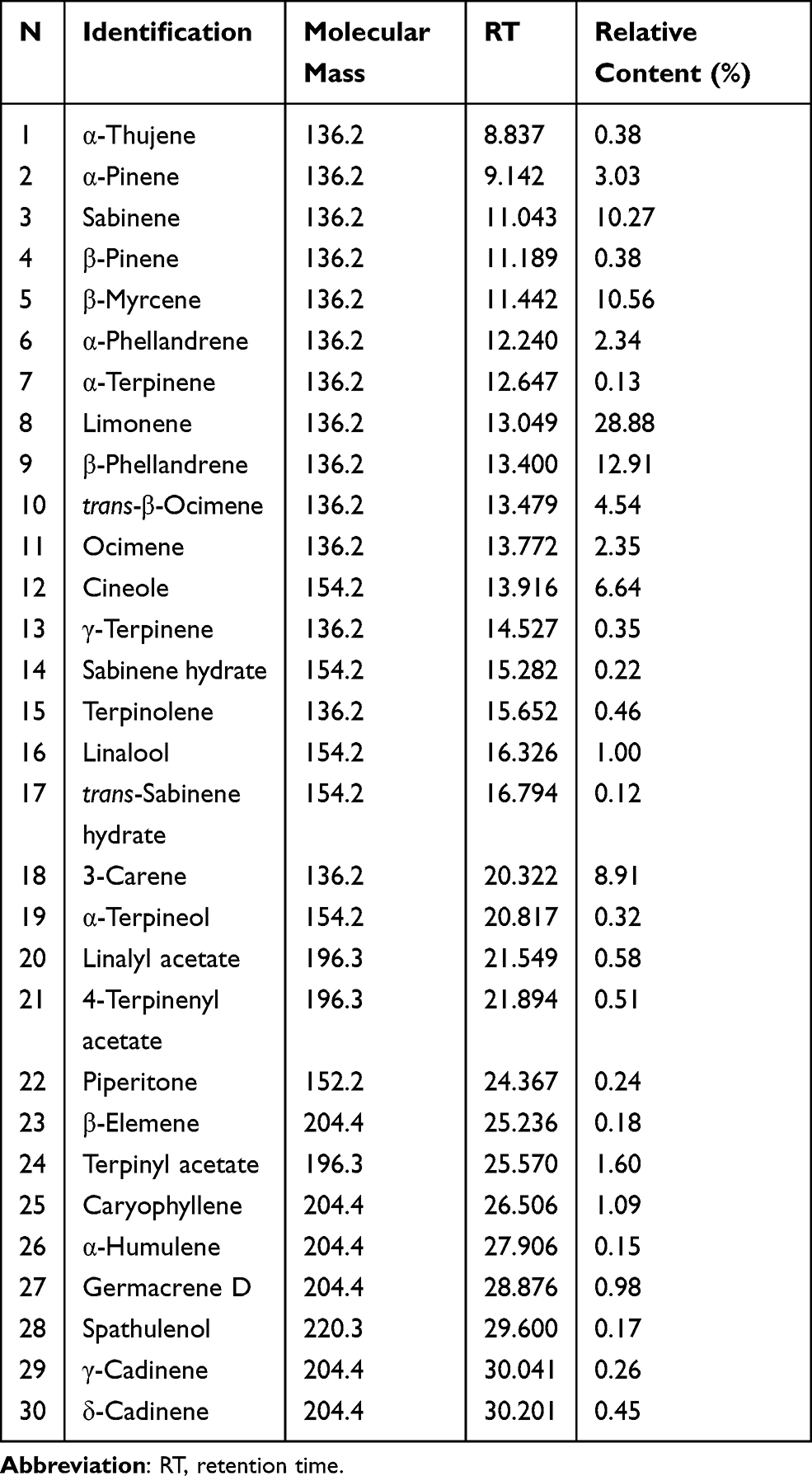 |
Table 2 Composition Analysis of the Volatile Oil of Z. bungeanum |
Discussion
Anxiety disorders, common debilitating and disabling psychiatric conditions, are characterized by excessive and persistent worry, fear, and avoidant response, differing from transient anxiety and fear in daily life.37,38 The etiology of ADs is conceptualized as the interaction of psychosocial factors (e.g., early childhood adversity and stressful life events) and genetic vulnerability.37,39 Among these risk factors associated with ADs, stressful life events seem to run through the lifetime of individuals and have an enormous impact on their behavior. Chronic experiences of life stress, such as disrupted interpersonal relationships, family and marriage conflicts, financial difficulties, job problems and trauma, promote the onset of ADs.40 Importantly, ADs, as well as depression, are associated with a history of early life stress.40–42 According to the above findings of etiology, we applied the CUS to imitate the psychosocial stress suffered by patients with ADs in the present study. The exposure of CUS decreased the ROE and ROT of model rats in EPM test, as well the central zone entries in OF test. These behavioral test results were consistent with those proposed in other literature,29 indicating that the modified CUS procedures successfully and reliably induced anxiety behavior in rats. However, the VOZB significantly alleviated the CUS-induced anxiety behavior in the EPM and OF tests.
Early treatments are beneficial for individuals with ADs to prevent the impairment of daily functioning and reverse the tendency of relapse.37 However, long-term pharmacotherapy (e.g., selective serotonin-reuptake inhibitors, serotonin-noradrenaline-reuptake inhibitors) is often accompanied by some intolerable adverse effects, particularly nausea and dizziness, reducing the treatment rate and compliance of patients with ADs.39 Thus, these patients with limited treatment options turn their attention to complementary and alternative medicine.43 In traditional Chinese medicine, herbal medicines have been widely applied in the treatment of various diseases for thousands of years, including mental disorders.27,44 In addition, plant‑derived compounds/extracts are regarded as the primary drug sources, providing over 50% of the available drugs.45,46 Previous studies revealed that Z. bungeanum compounds/extracts (e.g., hydroxy-α-sanshool, gx-50, and polyphenol extracts) showed notable neural activities to alleviate memory impairment and Alzheimer’s disease,26–28 but the effects of Z. bungeanum on ADs were unknown. Our present study indicated that VOZB was effective in alleviating the CUS-induced anxiety behavior in rats. Subsequently, a total of 30 compounds with relatively enrichment were identified as the potential curative compounds of VOZB. Most of them are classified into the terpenoids and reported in phytochemical studies of Z. bungeanum. However, the chemical ingredients and contents of terpenoids in present study were not completely consistent with other phytochemical studies, because of the difference in genes, species, geographic regions, growth conditions, and extraction methods.47
The pathogenesis of ADs has become a major topic in psychiatry, both for prevention and treatment.40 Nowadays, it is broadly accepted that imbalance of neuroendocrine homeostasis resulting from psychosocial stress plays a prominent role in the occurrence of mental disorders, such as ADs and depression.41,48 Of the neuroendocrine axes, the HPA axis plays a fundamental role in response to external and internal stimuli, including psychological and social stressors.41,49 The activation of the HPA axis by stress experiences implies a cascade of events starting with the secretion of CRH in the hypothalamus, which is then transported into the pituitary via the portal circulation and stimulates the synthesis and release of ACTH. ACTH further promotes the synthesis and release of the glucocorticoid hormone cortisol (also known as corticosterone) by the adrenal cortex, which has a significant role in maintaining homoeostasis in response to stress.50 The elevated glucocorticoid hormone ultimately reduces the CRH secretion of the hypothalamus through negative feedback regulation. Increasing studies suggest that stressful life events and abnormalities of HPA axis seem to be involved in the onset of ADs.41 Early life stress might trigger permanent changes in the HPA axis and predispose individuals to develop ADs in adulthood.51 For instance, hyperactivity of the HPA axis was also described in patients with generalized anxiety disorder, who often reported a history of early life stress.9 Additionally, psychosocial stressors provoked hyper-responsiveness of the adrenal cortex in social phobics, and altered the function of the glucocorticoids receptor.40 Consequently, these consistent literatures indicate that increased activity of the HPA axis and reduced inhibitory feedback contribute to the pathogenesis of ADs, and similar results were obtained in our present study. The CUS significantly elevated the expression of CRH, then persistently stimulated the HPA axis, leading ultimately to the increase of CORT level, but reducing the expression of GR. However, VOZB showed an inhibitory regulation on the hyperactivity of the HPA axis and reversed the alternation of GR, thus alleviating the CUS-induced anxiety behavior in rats.
Gut microbiota, the most complex community of microorganisms in the human body, has been proposed to participate in the regulation of host’s brain function and behaviors via the gut-brain axis,52 which integrates immune, neuroendocrine, and neural pathways. The alteration of gut microbiota has been increasingly recognized to affect the brain function and behavior, including neuroendocrine responses to stress.52,53 Growing preclinical studies revealed that the disturbance of gut microbiota contributes to stress-related psychiatric disorders, such as ADs. The transplantation of fecal microbiota from patients with depression or irritable bowel syndrome induced anxiety-like behavior in microbiota-depleted rats.54,55 The mild anxiety-like behavior was also reported in ampicillin-induced dysbiosis rats.56 Besides, maternal separation induced anxiety-like behavior and behavioral despair were only observed in the presence of gut microbiota but not in GF mice.57 Interestingly, stress-sensitive strains of rodents, such as BALB/c mice and F344 rats, showed increased anxiety behavior under GF conditions.58,59 In our present study, the CUS-induced anxiety rats showed appreciable alterations of gut microbiota. At phylum level, Firmicutes and Deferribacteres were increased after the CUS exposure. At family level, Lachnospiraceae and Ruminococcaceae were increased while Bacteroidales_S24-7_group, Lactobacillaceae, and Prevotellaceae were decreased after the CUS exposure. These results indicated that the CUS exposure, similar to the abnormal fecal microbiota transplantation and antibiotic treatment, triggered the microbial dysbiosis, which contributes to the host’s anxiety behavior. In recent years, targeting gut microbiota has become a potential therapeutic strategy for mental disorders, including fecal microbiota transplantation and probiotics. Supplementing certain strains of Bifidobacterium and Lactobacillus, such as B. pseudocatenulatum, B. longum, and L. rhamnosus, is beneficial for improving stress-induced anxiety behavior in rodents.60–62 Moreover, supplements of L. plantarum, L. helveticus, and B. longum also attenuated increases of subjective anxiety in volunteers.63–65 Increasing evidence indicates that VOZB and its main compounds could improve the disruption of gut microbiota. The treatment with VOZB increased the levels of the intestinal commensal flora including Lactobacillus and Bifidobacteria, whereas decreased Escherichia coli in dextran sulfate sodium-induced colitis mice.66 Another study showed that VOZB had an excellent inhibitory effect on E. coli both in vitro and in vivo, improving the enteritis.67 Besides, limonene, as an important component of VOZB, was revealed to elevate the relative abundance of Lactobacillus in cecum and colon of mice, as well as linalool.68 Similarly, cineole promoted the members of the phylum Firmicutes, including segmented filamentous bacteria in Citrobacter rodentium-induced colitis.69 Our present study found that the VOZB treatment effectively restored the compositions of gut microbiota in the CUS-induced anxiety rats, particularly raising Lactobacillaceae and Prevotellaceae. These findings implied that the ameliorative effect of VOZB on the CUS-induced anxiety was involved in restoring the dysbiosis of gut microbiota.
Although the precise mechanisms by which the gut microbiota affects host’s behavior are not yet absolutely delineated, the interaction of gut microbiota and neuroendocrine system has been regarded to be involved in the regulation of host’s mood and behavior. Considered to be the pivotal neuroendocrine system, the HPA axis ensures an adequate response to psychological and physical stressors. While assessment of HPA axis activity in GF or dysbiosis conditions has yielded inconsistent results, most studies supported that hormones of the HPA axis are of pivotal importance for the regulatory effect of gut microbiota on the host’s mood and behaviors. In some stress-related psychiatric disorders, such as ADs and depression, the disturbed gut microbiota apparently contributes to abnormality of the HPA axis. GF mice showed elevations of corticocerebral CRH, plasma ACTH, and plasma corticosterone, and reduction of corticocerebral GR, in response to restraint stress.16,17,70 Colonization with fecal microbiota of SPF mice or B. infantis in neonatal stage attenuated the increased responsiveness of the HPA axis in GF mice, whereas colonization with E. coli exacerbated the HPA response to restraint stress.22 Similarly, the corticosterone level of GF mice were exaggerated in response to novel environment stress.21 Moreover, antibiotic treatment was demonstrated to raise the plasma (or serum) corticosterone levels in mice.71,72 In addition, restoration of gut microbiota was available for reversing the activation of the HPA axis induced by stress exposure. Probiotics, mainly including Bifidobacterium, Lactobacillus and their formulations, have been proved to attenuate the activation of the HPA axis in response to various stressors, such as maternal separation,60,73 environmental stress,74 and partial restraint stress.75 Consistent with the above laboratory results, supplement of probiotics was beneficial for preventing the elevation of cortisol levels in volunteers with anxiety.63,64,76 These findings provided a clue that alteration of the HPA axis induced by stress exposure has a possible involvement of the microbiota. Our present study found that activation of the HPA axis was paralleled by altered gut microbiota in the CUS-induced anxiety rats, but the VOZB treatment was able to alleviate this abnormal interaction of gut microbiota and the HPA axis. Herein, we speculated that the ameliorative effect of VOZB stress-related anxiety potentially depended on restoring interactions of gut microbiota and the HPA axis.
The present study involved some limitations that should be considered. First, we did not use fecal microbiota transplantation to further validate that the anxiolytic effect of VOZB is dependent on improving the abnormal interaction of gut microbiota and HPA axis, thus, we will further verify the present results in future studies. Moreover, we only delineated the changes in gut microbiota composition in this study, the effects of these apparently changed communities on CUS-induced anxiety require further validation. In addition, microbial metabolites, such as tryptophan metabolites, may also affect activation of the HPA axis. Thus, we will combine the genomics and metabolomics to further reveal the underlying mechanisms by which microorganisms regulate the activity of the HPA axis.
Conclusion
VOZB, the representative compounds of Z. bungeanum, has ameliorative effects on CUS-induced anxiety behavior. This anxiolytic effect is potentially involved in inhibiting activation of the HPA axis and restoring the dysbiosis of gut microbiota, thus improving the stress-induced abnormality of the microbiota-gut-brain axis and brain homeostasis. This work provides a new insight into the pharmacological role of Z. bungeanum in stress-related ADs, and may help us reap the full benefits of this plant.
Abbreviations
ADs, anxiety disorders; VOZB, volatile oil of Zanthoxylum bungeanum; HPA axis, hypothalamic-pituitary-adrenal axis; CUS, chronic unpredictable stress; EPM, elevated plus-maze; OF, open field; CRH, corticotropin-releasing hormone; GR, glucocorticoid receptor; ACTH, adrenocorticotropic hormone; CORT, corticosterone; GC-MS, Gas Chromatography-Mass Spectrometry; GF, Germ-free; SPF, specific pathogen-free; CMC-Na, sodium carboxymethyl cellulose; AOD, average optical density; ANOVA, analysis of variance; ROE, ratio of open-arm entries; ROT, ratio of open-arm time; OTUs, operational taxonomic units; PCoA, principal coordinates analysis; NMDS, nonmetric multidimensional scaling.
Data Sharing Statement
The data used to support the findings of this study are available from Chunjie Wu upon request.
Acknowledgments
This work was supported by the “Xinglin Scholars” Scientific Research Promotion Program of Chengdu University of Traditional Chinese Medicine (No. 030041167).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Kessler RC, Aguilar-Gaxiola S, Alonso J, et al. The global burden of mental disorders: an update from the WHO World Mental Health (WMH) surveys. Epidemiol Psichiatr Soc. 2009;18(1):23–33. doi:10.1017/s1121189x00001421
2. Remes O, Brayne C, van der Linde R, Lafortune L. A systematic review of reviews on the prevalence of anxiety disorders in adult populations. Brain Behav. 2016;6(7):e00497. doi:10.1002/brb3.497
3. Chisholm D, Sweeny K, Sheehan P, et al. Scaling-up treatment of depression and anxiety: a global return on investment analysis. Lancet Psychiatry. 2016;3(5):415–424.
4. Kessler RC, Petukhova M, Sampson NA, Zaslavsky AM, Wittchen HU. Twelve-month and lifetime prevalence and lifetime morbid risk of anxiety and mood disorders in the United States. Int J Methods Psychiatr Res. 2012;21(3):169–184. doi:10.1002/mpr.1359
5. Wittchen HU, Jacobi F, Rehm J, et al. The size and burden of mental disorders and other disorders of the brain in Europe 2010. Eur Neuropsychopharmacol. 2011;21(9):655–679. doi:10.1016/j.euroneuro.2011.07.018
6. Whiteford HA, Degenhardt L, Rehm J, et al. Global burden of disease attributable to mental and substance use disorders: findings from the Global Burden of Disease Study 2010. Lancet. 2013;382(9904):1575–1586. doi:10.1016/S0140-6736(13)61611-6
7. Stein DJ, Scott KM, de Jonge P, Kessler RC. Epidemiology of anxiety disorders: from surveys to nosology and back. Dialogues Clin Neurosci. 2017;19(2):127–136.
8. Cosci F, Mansueto G. Biological and clinical markers to differentiate the type of anxiety disorders. Adv Exp Med Biol. 2020;1191:197–218. doi:10.1007/978-981-32-9705-0_13
9. Maron E, Nutt D. Biological markers of generalized anxiety disorder. Dialogues Clin Neurosci. 2017;19(2):147–158.
10. Bartlett AA, Singh R, Hunter RG. Anxiety and epigenetics. Adv Exp Med Biol. 2017;978:145–166. doi:10.1007/978-3-319-53889-1_8
11. Faravelli C, Lo Sauro C, Godini L, et al. Childhood stressful events, HPA axis and anxiety disorders. World J Psychiatry. 2012;2(1):13–25. doi:10.5498/wjp.v2.i1.13
12. Dieleman GC, Huizink AC, Tulen JH, et al. Alterations in HPA-axis and autonomic nervous system functioning in childhood anxiety disorders point to a chronic stress hypothesis. Psychoneuroendocrinology. 2015;51:135–150. doi:10.1016/j.psyneuen.2014.09.002
13. Frankiensztajn LM, Elliott E, Koren O. The microbiota and the hypothalamus-pituitary-adrenocortical (HPA) axis, implications for anxiety and stress disorders. Curr Opin Neurobiol. 2020;62:76–82. doi:10.1016/j.conb.2019.12.003
14. Neufeld KM, Kang N, Bienenstock J, Foster JA. Reduced anxiety-like behavior and central neurochemical change in germ-free mice. Neurogastroenterol Motil. 2011;23(3):255–264. doi:10.1111/j.1365-2982.2010.01620.x
15. Zeng L, Zeng B, Wang H, et al. Microbiota modulates behavior and protein kinase C mediated cAMP response element-binding protein signaling. Sci Rep. 2016;6:29998. doi:10.1038/srep29998
16. Luo Y, Zeng B, Zeng L, et al. Gut microbiota regulates mouse behaviors through glucocorticoid receptor pathway genes in the hippocampus. Transl Psychiatry. 2018;8(1):187. doi:10.1038/s41398-018-0240-5
17. Huo R, Zeng B, Zeng L, et al. Microbiota modulate anxiety-like behavior and endocrine abnormalities in hypothalamic-pituitary-adrenal axis. Front Cell Infect Microbiol. 2017;7:489. doi:10.3389/fcimb.2017.00489
18. Zheng P, Zeng B, Zhou C, et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol Psychiatry. 2016;21(6):786–796. doi:10.1038/mp.2016.44
19. Bauer KC, Huus KE, Finlay BB. Microbes and the mind: emerging hallmarks of the gut microbiota-brain axis. Cell Microbiol. 2016;18(5):632–644. doi:10.1111/cmi.12585
20. Sherwin E, Sandhu KV, Dinan TG, Cryan JF. May the force be with you: the light and dark sides of the microbiota-gut-brain axis in neuropsychiatry. CNS Drugs. 2016;30(11):1019–1041. doi:10.1007/s40263-016-0370-3
21. Clarke G, Grenham S, Scully P, et al. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol Psychiatry. 2013;18(6):666–673. doi:10.1038/mp.2012.77
22. Sudo N, Chida Y, Aiba Y, et al. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol. 2004;558(Pt 1):263–275. doi:10.1113/jphysiol.2004.063388
23. Barouei J, Moussavi M, Hodgson DM. Effect of maternal probiotic intervention on HPA axis, immunity and gut microbiota in a rat model of irritable bowel syndrome. PLoS One. 2012;7(10):e46051. doi:10.1371/journal.pone.0046051
24. Zhang M, Wang J, Zhu L, et al. Zanthoxylum bungeanum maxim. (Rutaceae): a systematic review of its traditional uses, botany, phytochemistry, pharmacology, pharmacokinetics, and toxicology. Int J Mol Sci. 2017;18(10):2172. doi:10.3390/ijms18102172
25. Chen X, Wei Z, Zhu L, et al. Efficient approach for the extraction and identification of red pigment from zanthoxylum bungeanum maxim and its antioxidant activity. Molecules. 2018;23(5):1109. doi:10.3390/molecules23051109
26. Nakamura T, Komai N, Isogami I, et al. Memory and learning-enhancing effect of Daikenchuto, a traditional Japanese herbal medicine, in mice. J Nat Med. 2006;60:64–67. doi:10.1007/s11418-005-0012-4
27. Zhang M, Xie M, Wei D, et al. Hydroxy-α-sanshool isolated from Zanthoxylum bungeanum attenuates learning and memory impairments in scopolamine-treated mice. Food Funct. 2019;10(11):7315–7324. doi:10.1039/c9fo00045c
28. Tang M, Wang Z, Zhou Y, et al. A novel drug candidate for Alzheimer’s disease treatment: gx-50 derived from Zanthoxylum bungeanum. J Alzheimers Dis. 2013;34(1):203–213. doi:10.3233/JAD-121831
29. Bondi CO, Rodriguez G, Gould GG, Frazer A, Morilak DA. Chronic unpredictable stress induces a cognitive deficit and anxiety-like behavior in rats that is prevented by chronic antidepressant drug treatment. Neuropsychopharmacology. 2008;33(2):320–331. doi:10.1038/sj.npp.1301410
30. Das SK, Barhwal K, Hota SK, Thakur MK, Srivastava RB. Disrupting monotony during social isolation stress prevents early development of anxiety and depression like traits in male rats. BMC Neurosci. 2015;16:2. doi:10.1186/s12868-015-0141-y
31. Doron R, Lotan D, Versano Z, et al. Escitalopram or novel herbal mixture treatments during or following exposure to stress reduce anxiety-like behavior through corticosterone and BDNF modifications. PLoS One. 2014;9(4):e91455. doi:10.1371/journal.pone.0091455
32. Sidor MM, Rilett K, Foster JA. Validation of an automated system for measuring anxiety-related behaviours in the elevated plus maze. J Neurosci Methods. 2010;188(1):7–13. doi:10.1016/j.jneumeth.2010.01.021
33. Wei D, Xie L, Zhuang Z, et al. Gut microbiota: a new strategy to study the mechanism of electroacupuncture and moxibustion in treating ulcerative colitis. Evid Based Complement Alternat Med. 2019;2019:9730176. doi:10.1155/2019/9730176
34. Qi Q, Liu YN, Jin XM, et al. Moxibustion treatment modulates the gut microbiota and immune function in a dextran sulphate sodium-induced colitis rat model. World J Gastroenterol. 2018;24(28):3130–3144. doi:10.3748/wjg.v24.i28.3130
35. Zhu L, Wang L, Chen X, et al. Comparative studies on flavor substances of leaves and pericarps of zanthoxylum bungeanum maxim. at different harvest periods. Trop J Pharm Res. 2018;18(2):279–286. doi:10.4314/tjpr.v18i2.9
36. Gawali NB, Bulani VD, Gursahani MS, Deshpande PS, Kothavade PS, Juvekar AR. Agmatine attenuates chronic unpredictable mild stress-induced anxiety, depression-like behaviours and cognitive impairment by modulating nitrergic signalling pathway. Brain Res. 2017;1663:66–77. doi:10.1016/j.brainres.2017.03.004
37. Craske MG, Stein MB. Anxiety. Lancet. 2016;388(10063):3048–3059. doi:10.1016/S0140-6736(16)30381-6
38. Craske MG, Stein MB, Eley TC, et al. Anxiety disorders. Nat Rev Dis Primers. 2017;3:17024. doi:10.1038/nrdp.2017.24
39. Bandelow B, Michaelis S, Wedekind D. Treatment of anxiety disorders. Dialogues Clin Neurosci. 2017;19(2):93–107.
40. Faravelli C, Lo Sauro C, Lelli L, et al. The role of life events and HPA axis in anxiety disorders: a review. Curr Pharm Des. 2012;18(35):5663–5674. doi:10.2174/138161212803530907
41. Juruena MF, Eror F, Cleare AJ, Young AH. The role of early life stress in HPA axis and anxiety. Adv Exp Med Biol. 2020;1191:141–153. doi:10.1007/978-981-32-9705-0_9
42. Chocyk A, Majcher-Maślanka I, Dudys D, Przyborowska A, Wędzony K. Impact of early-life stress on the medial prefrontal cortex functions - a search for the pathomechanisms of anxiety and mood disorders. Pharmacol Rep. 2013;65(6):1462–1470. doi:10.1016/s1734-1140(13)71506-8
43. Trkulja V, Barić H. Current research on complementary and alternative medicine (CAM) in the treatment of anxiety disorders: an evidence-based review. Adv Exp Med Biol. 2020;1191:415–449. doi:10.1007/978-981-32-9705-0_22
44. Zhang Y, Long Y, Yu S, et al. Natural volatile oils derived from herbal medicines: a promising therapy way for treating depressive disorder. Pharmacol Res. 2020;11. doi:10.1016/j.phrs.2020.105376
45. Newman DJ, Cragg GM. Natural products as sources of new drugs from 1981 to 2014. J Nat Prod. 2016;79(3):629–661. doi:10.1021/acs.jnatprod.5b01055
46. Zhang Q, Liu J, Li R, et al. A network pharmacology approach to investigate the anticancer mechanism and potential active ingredients of Rheum palmatum L. against lung cancer via induction of apoptosis. Front Pharmacol. 2020;11:528308. doi:10.3389/fphar.2020.528308
47. Xing-dong L, Hua-li X. Antifungal activity of the essential oil of Zanthoxylum bungeanum and its major constituent on Fusarium sulphureum and dry rot of potato tubers. Phytoparasitica. 2014;42(4):509–517. doi:10.1007/s12600-014-0388-3
48. Zorn JV, Schür RR, Boks MP, Kahn RS, Joëls M, Vinkers CH. Cortisol stress reactivity across psychiatric disorders: a systematic review and meta-analysis. Psychoneuroendocrinology. 2017;77:25–36. doi:10.1016/j.psyneuen.2016.11.036
49. Gold PW. The organization of the stress system and its dysregulation in depressive illness. Mol Psychiatry. 2015;20(1):32–47. doi:10.1038/mp.2014.163
50. Juruena MF, Agustini B, Cleare AJ, Young AH. A translational approach to clinical practice via stress-responsive glucocorticoid receptor signaling. Stem Cell Investig. 2017;4:13. doi:10.21037/sci.2017.02.01
51. Maniam J, Antoniadis C, Morris MJ. Early-life stress, HPA axis adaptation, and mechanisms contributing to later health outcomes. Front Endocrinol (Lausanne). 2014;5:73. doi:10.3389/fendo.2014.00073
52. Mohajeri MH, La Fata G, Steinert RE, Weber P. Relationship between the gut microbiome and brain function. Nutr Rev. 2018;76(7):481–496. doi:10.1093/nutrit/nuy009
53. O’Mahony SM, Clarke G, Dinan TG, Cryan JF. Early-life adversity and brain development: is the microbiome a missing piece of the puzzle? Neuroscience. 2017;342:37–54. doi:10.1016/j.neuroscience.2015.09.068
54. Kelly JR, Borre Y, O’ Brien C, et al. Transferring the blues: depression-associated gut microbiota induces neurobehavioural changes in the rat. J Psychiatr Res. 2016;82:109–118. doi:10.1016/j.jpsychires.2016.07.019
55. De Palma G, Lynch MD, Lu J, et al. Transplantation of fecal microbiota from patients with irritable bowel syndrome alters gut function and behavior in recipient mice. Sci Transl Med. 2017;9(379):eaaf6397. doi:10.1126/scitranslmed.aaf6397
56. Wang T, Hu X, Liang S, et al. Lactobacillus fermentum NS9 restores the antibiotic induced physiological and psychological abnormalities in rats. Benef Microbes. 2015;6(5):707–717. doi:10.3920/BM2014.0177
57. De Palma G, Blennerhassett P, Lu J, et al. Microbiota and host determinants of behavioural phenotype in maternally separated mice. Nat Commun. 2015;6:7735. doi:10.1038/ncomms8735
58. Nishino R, Mikami K, Takahashi H, et al. Commensal microbiota modulate murine behaviors in a strictly contamination-free environment confirmed by culture-based methods. Neurogastroenterol Motil. 2013;25(6):521–528. doi:10.1111/nmo.12110
59. Crumeyrolle-Arias M, Jaglin M, Bruneau A, et al. Absence of the gut microbiota enhances anxiety-like behavior and neuroendocrine response to acute stress in rats. Psychoneuroendocrinology. 2014;42:207–217. doi:10.1016/j.psyneuen.2014.01.014
60. Moya-Pérez A, Perez-Villalba A, Benítez-Páez A, Campillo I, Sanz Y. Bifidobacterium CECT 7765 modulates early stress-induced immune, neuroendocrine and behavioral alterations in mice. Brain Behav Immun. 2017;65:43–56. doi:10.1016/j.bbi.2017.05.011
61. Savignac HM, Kiely B, Dinan TG, Cryan JF. Bifidobacteria exert strain-specific effects on stress-related behavior and physiology in BALB/c mice. Neurogastroenterol Motil. 2014;26(11):1615–1627. doi:10.1111/nmo.12427
62. Barrera-Bugueño C, Realini O, Escobar-Luna J, et al. Anxiogenic effects of a Lactobacillus, inulin and the synbiotic on healthy juvenile rats. Neuroscience. 2017;359:18–29. doi:10.1016/j.neuroscience.2017.06.064
63. Andersson H, Tullberg C, Ahrné S, et al. Oral administration of lactobacillus plantarum 299v reduces cortisol levels in human saliva during examination induced stress: a randomized, double-blind controlled trial. Int J Microbiol. 2016;2016:8469018. doi:10.1155/2016/8469018
64. Allen AP, Hutch W, Borre YE, et al. Bifidobacterium longum 1714 as a translational psychobiotic: modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl Psychiatry. 2016;6(11):e939. doi:10.1038/tp.2016.191
65. Yang B, Wei J, Ju P, Chen J. Effects of regulating intestinal microbiota on anxiety symptoms: a systematic review. Gen Psychiatr. 2019;32(2):e100056. doi:10.1136/gpsych-2019-100056
66. Zhang Z, Shen P, Liu J, et al. In vivo study of the efficacy of the essential oil of zanthoxylum bungeanum pericarp in dextran sulfate sodium-induced murine experimental colitis. J Agric Food Chem. 2017;65(16):3311–3319. doi:10.1021/acs.jafc.7b01323
67. Hong L, Jing W, Qing W, et al. Inhibitory effect of Zanthoxylum bungeanum essential oil (ZBEO) on Escherichia coli and intestinal dysfunction. Food Funct. 2017;8(4):1569–1576. doi:10.1039/c6fo01739h
68. Wang L, Zhang Y, Fan G, Ren JN, Zhang LL, Pan SY. Effects of orange essential oil on intestinal microflora in mice. J Sci Food Agric. 2019;99(8):4019–4028. doi:10.1002/jsfa.9629
69. Baker J, Brown K, Rajendiran E, et al. Medicinal lavender modulates the enteric microbiota to protect against Citrobacter rodentium-induced colitis. Am J Physiol Gastrointest Liver Physiol. 2012;303(7):G825–G836. doi:10.1152/ajpgi.00327.2011
70. Cohen SM, Tsien RW, Goff DC, Halassa MM. The impact of NMDA receptor hypofunction on GABAergic neurons in the pathophysiology of schizophrenia. Schizophr Res. 2015;167(1–3):98–107. doi:10.1016/j.schres.2014.12.026
71. Fröhlich EE, Farzi A, Mayerhofer R, et al. Cognitive impairment by antibiotic-induced gut dysbiosis: analysis of gut microbiota-brain communication. Brain Behav Immun. 2016;56:140–155. doi:10.1016/j.bbi.2016.02.020
72. Scheer S, Medina TS, Murison A, et al. Early-life antibiotic treatment enhances the pathogenicity of CD4+ T cells during intestinal inflammation. J Leukoc Biol. 2017;101(4):893–900. doi:10.1189/jlb.3MA0716-334RR
73. Gareau MG, Jury J, MacQueen G, Sherman PM, Perdue MH. Probiotic treatment of rat pups normalises corticosterone release and ameliorates colonic dysfunction induced by maternal separation. Gut. 2007;56(11):1522–1528. doi:10.1136/gut.2006.117176
74. Bravo JA, Forsythe P, Chew MV, et al. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci U S A. 2011;108(38):16050–16055. doi:10.1073/pnas.1102999108
75. Ait-Belgnaoui A, Durand H, Cartier C, et al. Prevention of gut leakiness by a probiotic treatment leads to attenuated HPA response to an acute psychological stress in rats. Psychoneuroendocrinology. 2012;37(11):1885–1895. doi:10.1016/j.psyneuen.2012.03.024
76. Messaoudi M, Lalonde R, Violle N, et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br J Nutr. 2011;105(5):755–764. doi:10.1017/S0007114510004319
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.