Back to Journals » Nature and Science of Sleep » Volume 15
The Underlying Mechanisms of Sleep Deprivation Exacerbating Neuropathic Pain
Received 25 March 2023
Accepted for publication 25 July 2023
Published 28 July 2023 Volume 2023:15 Pages 579—591
DOI https://doi.org/10.2147/NSS.S414174
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sarah L Appleton
Manmin Zhu, Hao Huang
Department of Neurology, Affiliated Hospital of Zunyi Medical University, Guizhou, People’s Republic of China
Correspondence: Hao Huang, Department of Neurology, Affiliated Hospital of Zunyi Medical University, Guizhou, People’s Republic of China, Email [email protected]
Abstract: Pain disrupts sleep, and sleep deprivation or interference can alter pain perception in animals and humans, for example by increasing sensitivity to pain. However, the mechanism by which sleep affects neuropathic pain remains unclear. In this review, we discuss the available evidence from the epidemiologic, clinical, and human, as well as laboratory studies. In previous studies, we have found that sleep deprivation affects various injurious systems, including opioids, dopaminergic, immune, orexins, hypothalamic-pituitary-adrenal axis, and adenosine. At the same time, these systems play a crucial role in neuropathic pain regulation. In the complex interactions between these neurobiological systems, there may be potential regulatory pathways through which sleep deprivation amplifies neuropathic pain. Because of the impact sleep problems and neuropathic pain can have on the patients’ quality of life, studying the link between sleep and neuropathic pain is important for neuropathic pain prevention and public health.
Keywords: sleep, sleep deprivation, neuropathic pain, chronic pain, neurobiological mechanisms
Introduction
Pain is a physiological response designed to protect the organism from actual or potential tissue damage.1 In contrast to protective acute pain, chronic pain can lead to physical and emotional disturbances that can severely reduce a patient’s quality of life.2,3 Neuropathic pain is a persistent pain caused by damage to the central nervous system (CNS) or peripheral nervous system (PNS).1 Many diseases, such as trigeminal neuralgia, postherpetic neuralgia, diabetic neuropathy, stroke, and spinal cord injury, can cause neuropathic pain, which is often accompanied by sleep problems.4 Pain is a highly prevalent and costly public health problem worldwide and is one of the leading causes of disability worldwide.5,6 A good quality of sleep contributes to a better quality of life and health. Studies show that approximately one quarter of the US population experiences insomnia, that adults have experienced insomnia at least once, and that insomnia adversely affects disease prognosis and mortality risk.7 A growing body of epidemiological evidence suggests that sleep deprivation and disorders are risk factors for physical8,9 and mental health.10,11 It is widely accepted that the relationship between sleep deprivation and pain is strong.12 Pain disrupts sleep, and sleep deprivation or sleep disturbance increases pain or can lower pain thresholds, an association that has been demonstrated in many experimental models of sleep deprivation.13–16 However, we know very little about the underlying mechanisms. Therefore, an in-depth exploration of the mechanisms behind them would provide important guidance for the treatment of the disease.
Literature Search Methods
This narrative review searched for studies that discussed the specific effects of sleep deprivation on neuropathic pain and its underlying mechanisms. Original articles were identified through a systematic search strategy. MEDLINE (PubMed) was searched for eligible studies without specifying the study period. The review covers a wide range of areas, and the key search terms for each item are as follows: “sleep deprivation”, “neuropathic pain”, “opioid”, “dopamine”, “serotonin”, “5-hydroxytryptamine”, “immune system”, “inflammatory mediators”, “orexins”, “hypothalamic-pituitary-adrenal axis”, “adenosine”, “chronic pain”, “sleep stage”, “non-rapid eye movement sleep”, “rapid eye movement”, “sleep quality”, “sleep-wake states”, “pain sensitivity”, and “analgesics”.
Neuropathic Pain
Depending on the etiology, chronic pain can be classified as nociceptive pain, neuropathic pain and psychogenic pain.17 The injury or disease of the somatosensory nervous system causes neuropathic pain. It belongs to one of the chronic pain. It manifests mainly as abnormal pain (pain response to standard painless stimulus), hyperalgesia (increased response to pain stimulus), and spontaneous pain.1 Neuropathic pain can be divided into peripheral and central neuropathic pain based on the location of the lesion.1,3 Central neuropathic pain is caused by spinal or brain lesions, such as cerebrovascular disease affecting the central sensory system transmission pathway and spinal cord injury, syringomyelia, demyelination, and other illnesses caused by central neuropathic pain.1,3 Peripheral neuropathic pain is more common than central neuropathic pain, such as diabetes-related nerve pain, trigeminal neuralgia, and postherpetic neuralgia.1–3
Neuropathic pain affects about 10% of the global population, with women, the elderly, and rural residents mostly reporting pain, and it has significant adverse effects on their quality of life and daily functioning.18,19 Due to anatomical and biological changes, the perception of neuropathic pain is usually reduced in older people and therefore less data is reported on neuropathic pain. This is why this group of older people needs extra attention when it comes to neuropathic pain.20 Current treatments for neuropathic pain mainly rely on medications, such as opioids, antidepressants, and anticonvulsants.20 Although there has been tremendous progress in the research of analgesic drugs in recent years, the effect of traditional methods on neuropathic pain is not satisfactory. Therefore, it is essential to study the mechanism of neuropathic pain. For now, the research on the mechanism of pain at home and abroad is mainly based on animal models, and the research on the human body is relatively few.
Sleep
Under the physiological state, sleep can help people recover physical strength, improve metabolism, improve immunity, etc., and is a must for people to carry out normal life activities. According to an electroencephalogram, sleep is usually divided into two periods, namely non-rapid eye movement (NREM) and rapid eye movement (REM). The two periods alternate during night sleep, with NREM sleep usually most prominent at the beginning of sleep and REM sleep most prominent at the end.21 Slow wave activity (SWA) during NREM sleep is closely related to sleep homeostasis and is essential for maintaining physical and mental health as well as brain plasticity and optimal recovery. REM sleep is crucial to the quality of sleep. Changes in REM sleep will lead to sleep fragmentation to varying degrees and reduce sleep efficiency.22
Sleep is regulated through complex interactions between multiple brain regions and neurotransmitters. Many neurotransmitters such as the monoamines norepinephrine (NE), dopamine (DA), serotonin (5-HT) and the amino acids glutamic acid (GLU), gamma-aminobutyric acid (GABA) are involved in the sleep process.23 It was shown that activated glutamatergic neurons promote wakefulness by projecting to the nucleus accumbens and the lateral hypothalamus, and that GLU levels in parts of the mouse cortex were elevated during wakefulness and REM sleep, and decreased during non-REM sleep.24 DA, 5-HT and NE, as important excitatory neurotransmitters in the CNS, are involved in the formation of the upward reticular activation system and play an important role in the regulation of the waking state. During NREM sleep, a transient increase in DA in the basolateral amygdala (BLA) terminates NREM sleep and initiates REM sleep.25 5-HT plays an important role in the promotion and maintenance of sleep.26,27 NE increases the number of awakenings and reduces REM sleep.28,29
The quality of sleep is the basis of the quality of life. In general, the assessment of sleep quality can be divided into two aspects: subjective and objective assessments.30 Subjective assessment methods require the subjective perception of sleep and rely on various scales and questionnaires, such as the Pittsburgh Sleep Quality Index (PSQI), the Richard-Campbell Sleep Questionnaire (RCSQ), and the Verran and Snyder-Halpern Sleep Scale (VSH).31 Objective assessment methods use instruments to monitor sleep and produce objective data related to sleep quality, such as polysomnography (PSG).31 However, there may be differences between subjectively and objectively assessed sleep parameters and this must be taken into account when assessing sleep quality.
Sleep disorders are a common and often persistent health problem worldwide. According to the third edition of the International Classification of Sleep Disorders (ICSD), sleep disorders can be classified as: insomnia, sleep-related breathing disorders, central disorders of hypersomnolence, circadian rhythm sleep-wake disorders, parasomnias, sleep-related movement disorders, and other sleep disorders.32 Sleep disorders are one of the common manifestations of physical and neurological disorders, and when the primary disorders are not adjusted and treated in a timely manner, they can usually induce more serious psychological disorders, thus attracting the attention of more and more neurologists.
Relationship Between Sleep and Neuropathic Pain
Sleep problems and pain problems were reported to be intertwined:33,34 Pain can lead to sleep disorders, sleep disorders can enhance the body’s sense of pain, can reduce the pain threshold and the psychological ability of pain management, and interfere with the pain inhibition effect of analgesic drugs, so it can cause pain and make pain persist, the correlation between the two has attracted people’s attention.35
The degree of neuropathic pain was significantly positively correlated with sleep quality.36 Therefore, sleep quality could be used as a predictor of pain prognosis. It has been confirmed that the slow-wave sleep of neuropathic pain model rats is reduced; namely, the sleep duration of the NREM phase is reduced.37 Sleep efficiency is affected by increased cortico-autonomic and somatosensory arousal during NREM in mice due to pain interference.38 In addition, Chloe et al found that chronic sleep deprivation rather than chronic sleep fragmentation gradually increased pain sensitivity in an animal sleep deprivation model.39 In the clinical study, Chen et al found that sleep deprivation can reduce the pain threshold of healthy people, resulting in hyperalgesia and oxidative stress.40 Thus, neuropathic pain can disrupt sleep in animal and clinical studies, and the loss of sleep quality can further alter pain sensitivity.
The Underlying Mechanisms of Sleep Deprivation Exacerbating Neuropathic Pain
Neuropathic pain and sleep disorders affect and interact with each other, so it is speculated that there is some common material basis or initiating factors for their mechanisms. In order to solve the problem of clinical neuropathic pain accompanied by sleep disorders, this paper focuses on the possible mechanism of sleep deprivation exacerbating neuropathic pain. Figure 1 summarizes the effects of sleep deprivation on neurobiological systems and mediators associated with neuropathic pain.
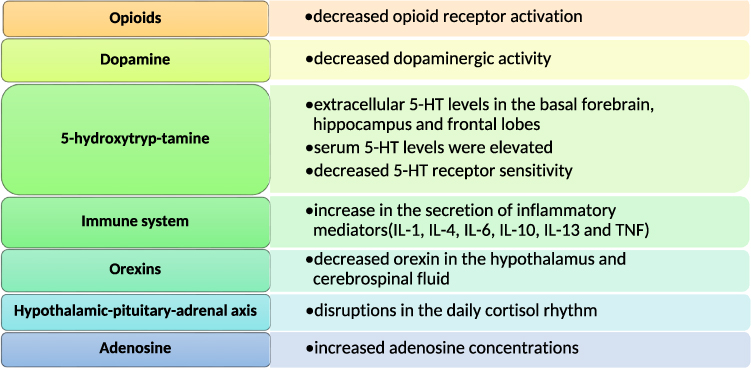 |
Figure 1 Neurobiological systems that may be affected by sleep deprivation. Abbreviations: 5-HT, 5-hydroxytryptamine; IL, interleukin; TNF, tumor necrosis factor. |
Opioids
Opioids are the most effective broad-spectrum analgesics available and are widely used in clinical practice for analgesic treatment.41 Opioids exert their pharmacological effects through three opioid receptors (µ, δ, and κ) that can be activated by endogenous opioid peptides, including enkephalins, dynorphins and endorphins, or by exogenous opioids, such as morphine.41 Among these receptors, the main analgesic effect of opioids is exerted by the µ-opioid receptor (MOR).42,43 Functional magnetic resonance imaging has shown that persistent pain induces the release of endogenous opioid peptides in various regions of the human brain, such as the cingulate gyrus, orbitofrontal, posterior parietal and insula cortices, and amygdala,44 and interacts with MOR.45 This highlights the importance of MOR in the endogenous regulation of pain in humans.
The role of opioids in modulating neuropathic pain has been extensively demonstrated in human and animal studies. During neuropathic pain, systemic and local treatment with opioids (naloxone) reversed the suppression of mechanical abnormal pain.46 Diabetic sensorimotor polyneuropathy (DSPN) is seen in approximately one-third of diabetic patients and significantly reduces their quality of life due to neuropathic pain and foot ulcers, and it has been investigated that painful DSPN may be as high as 25%.47 Pain can be substantially reduced with corresponding opioid use, with approximately 50% of individuals experiencing ≥50% pain reduction.47 In a mouse model of neuropathic pain induced by chronic constriction injury to the sciatic nerve, Celik et al found that activation of opioid receptors reduced pain after nerve injury in mice.48 The amygdala is thought to be the centre of integrated processing of negative emotional information and pain perception.49,50 In a study of post-perioperative pain in rats, it was found that reduced expression of MOR in the amygdala may be a key factor in chronic postoperative pain.51 In addition, hydrogen sulfide induced antioxidant and anti-apoptotic effects in the CNS and PNS were found to contribute to the analgesic effect of opioids during neuropathic pain in recent studies.46 This may provide new ideas for enhancing opioid treatment of neuropathic pain.
Sleep disruption and/or complete sleep deprivation can impair pain inhibition. Studies have found that sleep deprivation causes dysregulation of the endogenous opioid system52 and diminishes the analgesic effect of MOR agonists.53 As a result, the role played by opioids in the increase in pain-causing effects following sleep deprivation has attracted widespread attention. Randomized groups of 100 healthy adults were treated with morphine and placebo after two nights of undisturbed sleep and two nights of forced arousal sleep interruption. The results showed a 3-fold reduction in the analgesic effect of morphine after forced awakening sleep interruptions relative to undisturbed sleep.53 In a sleep deprivation rat model, sleep deprivation increased the duration of pain and significantly reduced the maximum morphine-induced anti-injury sensory effect.54 Sleep deprivation leads to reduced analgesia, increased pain and subsequent elevated opioid dose requirements, which in turn may further disrupt sleep. Sleep disruption may lead to a vicious dose-escalation cycle and thus an increasing abuse of opioids, which is also a topic of concern.
Dopamine
Dopamine (DA) has a wide range of effects on basic brain processes such as sleep, pain, addiction, emotional responses and motor control.55 There is growing evidence that dopamine is required for normal sleep-wake states. Drosophila mutants lacking functional dopamine transporters (DAT) exhibit prolonged wakefulness and shortened sleep.56 Similarly, Wisor et al found that mouse DAT mutants significantly increased the duration of arousal episodes during the second half of the active (nocturnal) phase of the circadian cycle.57 In addition, in clinical practice, impaired dopaminergic neurotransmission in the striatum of patients with Parkinson’s disease is often associated with severe sleep disturbances and daytime sleepiness, with low doses of dopamine agonists associated with insomnia and higher doses leading to excessive daytime sleepiness.58 Therefore, the use of moderate amounts of dopamine agonists in Parkinson’s patients, thus avoiding sleep disturbances, deserves further exploration by researchers.
DA regulates injury perception in various areas of the CNS, including the basal ganglia, thalamus, insular cortex, anterior cingulate cortex and periaqueductal gray (PAG).59,60 The PAG is a brain region involved in the regulation of injury perception, and MOR is densely expressed in the PAG.61–63 Given the central role of opioid receptors in mediating pain and their dense expression in the PAG, researchers investigated whether there is a link between them and dopamine in the regulation of pain and found that the ventral PAG/dorsal raphe dopamine neurons may play a key role in opioid anti-harm perception through the activation of downstream structures by dopamine and glutamate release.61 Diseases affected by DA dysfunction, such as Parkinson’s disease, addiction and depression, are often co-morbid with chronic pain symptoms. Patients with Parkinson’s with chronic pain often have a lower pain threshold compared to Parkinson’s without chronic pain.64,65
Studies have found that nociceptive hyperalgesia may occur following rapid eye movement (REM) sleep deprivation (SD) (REM-SD) by reducing the function of dopamine receptors in the post-synaptic dopaminergic system of the midbrain limbic but not the nigrostriatal dopaminergic system.66 This was validated in a rodent model of REM-SD, which increases pain by increasing adenosinergic activity and decreasing dopaminergic activity.67,68 Experimental administration of DA receptors agonists was shown to block the nociceptive hyperalgesic effects of sleep deprivation, further suggesting that sleep deprivation can increase pain by decreasing dopaminergic activity.68 These findings suggest that co-administration of dopaminergic drugs can improve the response to analgesics in patients with chronic pain associated with sleep disturbances.
5-Hydroxytryp-Tamine
5-hydroxytryptamine (5-HT) is currently acknowledged to act primarily to promote wakefulness and inhibit REM sleep (REMS).69,70 The relationship between sleep and 5-HT has been verified in both animal and human experiments. During sleep deprivation, the release of 5-HT is even higher than during the preceding waking period, as indicated by the findings in animals. After sleep deprivation in rats, extracellular 5-HT levels in the basal forebrain, hippocampus and frontal lobes all appear to be elevated.71,72 Zhao et al showed that sleep deprivation within 6–8 hours significantly increased the mRNA and protein levels of 5-HT 2A receptors in the frontal cortex of mice.73 In an acute sleep deprivation experiment on a healthy population, similar to animal experiments, serum 5-HT levels were elevated in subjects and metabolites of 5-HT were shown to have a possible role in antidepressants.74
5-HT is widely distributed in the peripheral and central nervous system. The most characteristic central system analgesic pathways are the 5-hydroxytryptaminergic-norepinephrinergic pathway and the opioidergic pathway.75 These pathways lead to the release of 5-HT, norepinephrine and endogenous opioids, they can inhibit the release of excitatory neurotransmitters such as glutamate and are activated in response to noxious stimuli, leading to a general decrease in pain sensitivity following exposure to acute painful stimuli.75 For example, intrathecal injection of 5-HT3 receptor antagonists significantly attenuated mechanical nociceptive hyperalgesia and thermal nociceptive hypersensitivity in 6-hydroxydopamine lesioned rats.76 The effectiveness of 5-hydroxytryptamine reuptake inhibitors in the treatment of various clinical pain conditions (eg fibromyalgia) further demonstrates the involvement of 5-HT receptors in pain regulation.77,78 In genetic studies, polymorphisms in monoamine metabolism and transport have been shown to correlate with the diagnosis or severity of fibromyalgia, such as serotonin 5-HT 2A receptor and 5-HT transporter.79
In that the 5-HT system is involved in sleep-wake and pain control, disruptions in this system may mediate the nociceptive hyperalgesic effects of sleep deprivation. Data from animal studies suggest that 5-HT does help promote pain-related behaviours in REM sleep deprived animals.80 Thus 5-HT may play a role in the mechanism by which sleep deprivation exacerbates pain.
Immune System
Modern society has many health-related problems following acute and chronic sleep deprivation, primarily involving the immune system.31 The immune system functions in the face of infection, tissue damage, or stress, relying primarily on inflammatory mediators’ secretion (eg, cytokines). With acute elevations of inflammatory proteins, this response is adaptive; however, chronic elevations can lead to health problems, including pain and sleep disturbances, endocrine disruption, and mood changes.81–84 It is a well-established fact that sleep and immunity are linked in both directions. Activation of the immune system can alter sleep, which in turn affects our body’s immune system. The most extensively studied roles of inflammatory mediators in sleep regulation are interleukin-1 (IL-1) and tumor necrosis factor (TNF).85–87 Levels of IL-1 and TNF in the brain are associated with sleep. For instance, their levels increase after sleep deprivation;88–91 injection of IL-1 or TNF enhances non-rapid eye movement sleep (NREMS), and inhibition of IL-1 or TNF inhibits spontaneous sleep and sleep rebound that occurs after sleep deprivation.92 Inflammatory mediators not only have an effect on NREMS, but REM sleep deprivation can also induce an inflammatory response.93 In addition to IL-1 and TNF, which regulate sleep, other anti-inflammatory cytokines, such as IL-4, IL-10, and IL-13, may act indirectly by regulating IL-1 and TNF.94 As mentioned above, sleep and the immune system are closely related.
It goes without saying that the immune system or inflammatory response is central to the body’s pain. There is evidence that pro-inflammatory cytokines (PICs) such as IL-1, IL-6 and TNF induce central sensitization and nociceptive hypersensitivity via different and overlapping synaptic mechanisms (increased excitatory synaptic transmission or decreased inhibitory synaptic transmission) in superficial dorsal horn neurons.95 Firstly, it is induced mainly in glial cells (microglia and astrocytes) following inflammation and nerve injury and promotes pain through neuro-glial cell interactions, contributing for example to the development and maintenance of pain promotion in diseases such as arthritis and multiple sclerosis.96,97 Secondly, intrathecal injection of IL-1 (5–5000 pg) or TNF (5–5000 pg) enhanced the dorsal horn neuronal response, with the effects of IL-1 being stronger than those of TNF. Intrathecal administration of IL-1 also led to the development of mechanically abnormal pain and nociceptive hyperalgesia, whereas TNF did not.98 Nevertheless, in other studies, it was found that TNF could likewise lead to the development of mechanically abnormal pain.99,100 Besides, the significant reduction in neuropathic pain after injection of antibodies related to PIC is further evidence of a role for inflammatory factors in mediating the mechanism of neuropathic pain.101
Sleep disorders, pain, and inflammatory processes are frequently observed in a variety of medical diseases and conditions, such as fibromyalgia, rheumatoid diseases, and surgery. In a human experimental setting, chronic sleep deprivation leads to elevated IL-6 plasma levels and is associated with increased pain sensation and overall physical discomfort.102 In animal experiments, IL-1, IL-6, and TNF mediated microglia activation and promoted CNS inflammation and neuronal apoptosis, lowering pain thresholds and thereby exacerbating pain behavior in sleep-deprived rats.103,104 In summary, sleep deprivation can lead to an inflammatory response that increases sensitivity to pain.
Orexins
Orexins are hypothalamic neuropeptides, divided into Orexin A and Orexin B, which act in conjunction with Orexin-1 and Orexin-2 receptors.105 Orexin neurons are present only in the lateral hypothalamic region, but project throughout the central nervous system, activating orexin neurons, monoaminergic and cholinergic neurons in the hypothalamus/brainstem region to maintain long, consolidated periods of wakefulness.106 In addition to this, orexin is associated with a variety of functions, including feeding and energy regulation, neuroendocrine regulation, and regulation of pain.107–109
Early on, the discovery that orexin deficiency could cause episodic sleeping sickness in several mammals, including mice, rats, dogs, and humans, caught the attention of scientists.110–112 Researchers have been shown to improve sleep in insomnia patients by inhibiting dual orexin receptors, findings that reinforce the importance of the orexin system in regulating sleep/wakefulness. Increased expression of orexin neurons was observed in the hypothalamus of sleep-deprived rats.113 It has also been reported that the number of orexin neurons is significantly reduced in patients with episodic sleeping sickness and that the level of orexin peptide in the cerebrospinal fluid is reduced to undetectable levels.114 This further supports the involvement of orexins in maintaining wakefulness. Hence, in clinical practice, orexin receptors are the target of pharmacological treatment for episodic sleeping sickness and insomnia, such as the dual orexin receptor antagonists Almorexant, Lemborexant, etc. The wakefulness-promoting effects of orexin have been validated in several multicenter, double-blind, randomized, placebo-controlled studies115–118 and have recently been approved by the FDA for the treatment of adults who have insomnia.119
Orexins are also associated with brain regions that regulate pain;120 for example, orexins in the ventral tegmental area are associated with injurious responses.121 Exogenously administered orexins have been shown to have analgesic effects in animal models. Orexins A and B, delivered by intracerebroventricular and intrathecal injection, enhance anti-injury perception in mice.122 Orexin-A exerts analgesic effects in a chemotherapy-induced neuropathic pain mouse model. In a mouse model of oxaliplatin-induced neuropathic pain, Mobarakeh et al found that abnormal mechanical pain was reversed by orexin-A and was more effective than duloxetine, whose analgesic effects were mediated by orexin type 1 receptor.122 These data suggest that mechanical nociceptive sensitization can potentially treat pain by activating the orexin A-orexin 1 receptor pathway. Orexin 1 receptor agonists may have a potential therapeutic role in the treatment of pain.
Collectively, the mechanisms involved in the involvement of the orexin system in pain and sleep-wake control, which may mediate the nociceptive hyperalgesic effects of sleep deprivation, still deserve further investigation and validation.
Hypothalamic-Pituitary-Adrenal Axis
The hypothalamic-pituitary-adrenal (HPA) axis includes interactions between the hypothalamus, pituitary, and adrenal glands, whose activation leads to the release of cortisol hormones,123 and regulates circulating levels of cortisol hormones, the primary neuroendocrine system providing rapid response and defense against stress in mammals.124
Circadian rhythm control of the HPA axis is critical for the regulation of hormones involved in the stress response, such as cortisol hormones. The daily rhythm of cortisol secretion is relatively stable and is mainly influenced by the biological clock. In a large number of human studies, sleep onset exerts an inhibitory effect on cortisol secretion.125 Abrupt changes in sleep duration can lead to severe disruptions in the daily cortisol rhythm, while sleep deprivation and/or decreased sleep quality appear to lead to a functionally important activation of the HPA axis.126,127 Because of the close relationship between HPA and circadian rhythms, disruption of circadian rhythms can have important effects on HPA. Animal experiments have shown that disruption of the circadian system through Per2 gene deletion dysregulates the HPA stress axis and is subsequently also associated with increased depression-like behavior and deficits in startle response.128
The HPA axis plays a pathogenic role in the pathogenesis of autoimmune/inflammatory diseases such as rheumatoid arthritis and rheumatic polymyalgia, where cortisol concentrations increase throughout the day and show diurnal variation.129 Rheumatic polymyalgia usually shows a good clinical response to glucocorticoid therapy,130 reduces pain, and has a decrease in all serum indicators of inflammation compared to pre-treatment.131 Plasma cortisol levels are significantly and consistently elevated after nerve injury, and the glucocorticoid receptors (GR) agonist dexamethasone treatment in a rat model of chronic constriction nerve injury-induced pain leads to increased neuropathic pain behavior, and GR antagonists such as RU38486 may be used to protect against the central effects of GR activation.132 Activation of glucocorticoid receptors can also lead to enhanced expression of glutamate receptor mRNA along with increased mechanical pain sensitivity.133
In summary, these studies in chronic pain and insomnia patients and animals observed more cortisol secretion and an over response of the HPA axis which may mediate the association between sleep deprivation and pain sensitivity.
Adenosine
Adenosine is a pleiotropic bioactive substance with powerful neuromodulatory properties. Due to its ability to easily cross the blood-brain barrier, it can act as a signaling molecule between the periphery and the brain. It acts through four (A1, A2A, A2B, and A3) adenosine receptors (ARs). These receptors are expressed on almost all cell types within the CNS.134 Although adenosine and its receptors represent clear targets for the pharmacological treatment of various diseases and pathologies, including pain, few drugs acting on the adenosinergic system have entered the market so far. The reason behind this discrepancy may be partly due to the universal distribution of AR in almost every cell and tissue, making it difficult to avoid unwanted side effects.
Adenosine plays an important role in sleep regulation and sleep promotion. Adenosine concentrations in the basal forebrain and cortical regions increase during SD, and this increase induces sleep. This suggests that adenosine may play a role in the local regulation of sleep homeostasis.135 In addition, it has been shown that sleep deprivation increases extracellular adenosine and A1 receptor (A1R) mRNA levels in basal forebrain cholinergic areas regions involved in sleep homeostasis.136 Thus, it appears that the basal forebrain, which appears to be central to the regulation/execution of SD or recovery from sleep after prolonged wakefulness.137
In recent years, extensive progress has been made in elucidating the role of adenosine in pain modulation. Most of the anti-injury sensory effects of adenosine depend on A1R activation.138 In a mouse model of acute postoperative pain, nociceptive hyperalgesia occurs through the involvement of reduced peripheral and central A1Rs.139 In another report, to elucidate the effect of adenosine on pain signaling, intrathecal adenosine injections were tested in two types of neuropathic pain (spinal cord compression and chronic constriction of the sciatic nerve) and in postoperative pain (plantar incision). The results showed that intrathecal adenosine injection suppressed nociceptive hyperalgesia in both neuropathic pain models but not in the postoperative pain model. However, A1R mRNA and protein expression were reduced in this model.140 This suggests that adenosine is functional for inhibiting pain signaling in neuropathic pain but less effective in postoperative pain due to the diminution of adenosine A1 receptors.
Recent studies have identified a possible mediating role for adenosine in linking sleep deprivation to increased pain sensitivity. Caffeine and modafinil are two adenosine antagonists that immediately normalize pain sensitivity in sleep-deprived animals.39 In a rat model of postoperative pain, systemic administration of caffeine at the onset of sleep deprivation prevented sleep deprivation-induced increases in postoperative hypersensitivity pain responses.141 In addition, microinjection of the A2A receptor antagonist ZM241385 into the preoptic nucleus blocked the previous sleep deprivation-induced increase in the level and duration of surgical pain and eliminated sleep deprivation-induced thermal hyperalgesia in a group of non-surgical (ie, no surgical incision) rats.141 Through the above experimental studies, we can find that one way in which sleep deprivation enhances pain is by possibly enabling an increase in adenosine activity.
Discussion
Recent clinical and experimental studies have identified an integral role for sleep deprivation in the mechanisms of neuropathic pain, mainly in the regulation of analgesic receptors, neurotransmitters, the neuroendocrine system, and the inflammatory response. In sleep deprivation-related experiments it has been found that there is typically a decrease in analgesic receptor such as opioid receptor activity, with MOR activity changes predominating, followed by downregulation of pain thresholds, increased pain sensitivity and prolonged pain; therefore, sleep disorders may lead to increased doses of opioid analgesics, forming a vicious cycle that leads to increasing opioid abuse, and clinicians need to be alerted. Neurotransmitter and neuroendocrine regulation play an important role in the pathogenesis of sleep-related neuropathic pain. REM-SD increases pain by decreasing dopaminergic activity, while in contrast to dopaminergic, 5-HT and adenosine activity is increased and can have widely increased levels of central and peripheral expression. The researchers found that the expression and peripheral levels of orexin neurons were significantly reduced in sleep disorders compared to normal, and that orexin injected intracerebroventricularly and intrathecally enhanced anti-damage perception in mice, so the orexin system may mediate the nociceptive hyperalgesia effect of sleep deprivation, but further investigation and validation are needed due to the paucity of related studies. In both human and animal studies, sleep deprivation typically induces an inflammatory response, and increased release of pro-inflammatory factors can promote inflammation in the central nervous system and apoptosis of neurons, such as IL-6, TNF, thereby lowering pain thresholds. The HPA axis is closely linked to the inflammatory response, and pro-inflammatory factors activate the HPA axis, leading to increased cortisol production, which in turn inhibits the production of pro-inflammatory cytokines. Thus, cortisol and synthetic glucocorticoids may indirectly regulate pain by altering the secretion of pro-inflammatory factors. We still know very little about the interaction between sleep and pain, and the mechanisms by which they interact are one of the hot topics of research worldwide. Therefore, an in-depth exploration of the mechanisms of interaction between sleep deprivation and neuropathic pain and the implementation of individualized treatment are important clinical guidelines for preventing and treating the disease and its prognosis.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This paper was supported by grants from the National Natural Science Foundation of China (No. 81760247), Graduate Workstation of Neurology, Zunyi Medical College (No. GZZ2017004), the Scientific Research Foundation of Doctor in the Affiliated Hospital of Zunyi Medical University (grant no. (2016)14), the Guizhou epilepsy basic and clinical research scientific and technological innovation talent team project (No: CXTD[2022]013), the Collaborative Innovation Center of Chinese Ministry of Education (No: 2020-39), the Guizhou provincial “hundred” level innovative talents funds (No: GCC-2022-038-1), the Guizhou Provincial Science and Technology Foundation (No: ZK2022-656), and the Zunyi City Science and Technology Foundation (No: 2019-71 and 2021-30), and Guizhou Provincial Science and Technology Foundation (No. (2019)1350).
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Costigan M, Scholz J, Woolf CJ. Neuropathic pain: a maladaptive response of the nervous system to damage. Annu Rev Neurosci. 2009;32:1–32. doi:10.1146/annurev.neuro.051508.135531
2. Breivik H, Collett B, Ventafridda V, Cohen R, Gallacher D. Survey of chronic pain in Europe: prevalence, impact on daily life, and treatment. Eur J Pain. 2006;10(4):287–333. doi:10.1016/j.ejpain.2005.06.009
3. Cohen SP, Vase L, Hooten WM. Chronic pain: an update on burden, best practices, and new advances. Lancet. 2021;397(10289):2082–2097. doi:10.1016/S0140-6736(21)00393-7
4. Afolalu EF, Ramlee F, Tang NKY. Effects of sleep changes on pain-related health outcomes in the general population: a systematic review of longitudinal studies with exploratory meta-analysis. Sleep Med Rev. 2018;39:82–97. doi:10.1016/j.smrv.2017.08.001
5. Nissen A, Hynek KA, Scales D, Hilden PK, Straiton M. Chronic pain, mental health and functional impairment in adult refugees from Syria resettled in Norway: a cross-sectional study. BMC Psychiatry. 2022;22(1):571. doi:10.1186/s12888-022-04200-x
6. Blyth FM, Noguchi N. Chronic musculoskeletal pain and its impact on older people. Best Pract Res Clin Rheumatol. 2017;31(2):160–168. doi:10.1016/j.berh.2017.10.004
7. Ohayon MM. Epidemiology of insomnia: what we know and what we still need to learn. Sleep Med Rev. 2002;6(2):97–111. doi:10.1053/smrv.2002.0186
8. Shin DS, Jeong BY. Older female farmers and modeling of occupational hazards, wellbeing, and sleep-related problems on musculoskeletal pains. Int J Environ Res Public Health. 2022;19(12):7274. doi:10.3390/ijerph19127274
9. Alghadir AH, Khan M, Alshehri MM, Alqahtani AS, Aldaihan M. In hypertensive individuals, sleep time and sleep efficiency did not affect the number of angina episodes: a cross-sectional study. Sci Rep. 2022;12(1):16290. doi:10.1038/s41598-022-20255-y
10. Jolly T, Vadukapuram R, Trivedi C, et al. Risk of suicide in patients with major depressive disorder and comorbid chronic pain disorder: an insight from national inpatient sample data. Pain Physician. 2022;25(6):419–425.
11. Lazaridou A, Paschali M, Zgierska AE, Garland EL, Edwards RR. Exploring the relationship between endogenous pain modulation, pain intensity, and depression in patients using opioids for chronic low back pain. Clin J Pain. 2022;38(10):595–600. doi:10.1097/AJP.0000000000001063
12. Krause AJ, Prather AA, Wager TD, Lindquist MA, Walker MP. The pain of sleep loss: a brain characterization in humans. J Neurosci. 2019;39(12):2291–2300. doi:10.1523/JNEUROSCI.2408-18.2018
13. Chang JR, Fu SN, Li X, et al. The differential effects of sleep deprivation on pain perception in individuals with or without chronic pain: a systematic review and meta-analysis. Sleep Med Rev. 2022;66:101695. doi:10.1016/j.smrv.2022.101695
14. Staffe AT, Bech MW, Clemmensen SLK, Nielsen HT, Larsen DB, Petersen KK. Total sleep deprivation increases pain sensitivity, impairs conditioned pain modulation and facilitates temporal summation of pain in healthy participants. PLoS One. 2019;14(12):e0225849. doi:10.1371/journal.pone.0225849
15. Schrimpf M, Liegl G, Boeckle M, Leitner A, Geisler P, Pieh C. The effect of sleep deprivation on pain perception in healthy subjects: a meta-analysis. Sleep Med. 2015;16(11):1313–1320. doi:10.1016/j.sleep.2015.07.022
16. Li Q, Zhu ZY, Lu J, et al. Sleep deprivation of rats increases postsurgical expression and activity of L-type calcium channel in the dorsal root ganglion and slows recovery from postsurgical pain. Acta Neuropathol Commun. 2019;7(1):217. doi:10.1186/s40478-019-0868-2
17. Stump PR, Dalben Gda S. Mechanisms and clinical management of pain. Braz Oral Res. 2012;26(Suppl 1):115–119. doi:10.1590/S1806-83242012000700017
18. Pricope CV, Tamba BI, Stanciu GD, et al. The roles of imaging biomarkers in the management of chronic neuropathic pain. Int J Mol Sci. 2022;23(21):13038. doi:10.3390/ijms232113038
19. Zimmer Z, Fraser K, Grol-Prokopczyk H, Zajacova A. A global study of pain prevalence across 52 countries: examining the role of country-level contextual factors. Pain. 2022;163(9):1740–1750. doi:10.1097/j.pain.0000000000002557
20. Giovannini S, Coraci D, Brau F, et al. Neuropathic pain in the elderly. Diagnostics. 2021;11(4):613. doi:10.3390/diagnostics11040613
21. Simon KC, Nadel L, Payne JD. The functions of sleep: a cognitive neuroscience perspective. Proc Natl Acad Sci U S A. 2022;119(44):e2201795119. doi:10.1073/pnas.2201795119
22. Barbato G. REM sleep: an unknown indicator of sleep quality. Int J Environ Res Public Health. 2021;18(24):12976. doi:10.3390/ijerph182412976
23. Helfrich-Förster C. Sleep in insects. Annu Rev Entomol. 2018;63(1):69–86. doi:10.1146/annurev-ento-020117-043201
24. Weigend S, Holst SC, Treyer V, et al. Dynamic changes in cerebral and peripheral markers of glutamatergic signaling across the human sleep-wake cycle. Sleep. 2019;42(11). doi:10.1093/sleep/zsz161
25. Hasegawa E, Miyasaka A, Sakurai K, Cherasse Y, Li Y, Sakurai T. Rapid eye movement sleep is initiated by basolateral amygdala dopamine signaling in mice. Science. 2022;375(6584):994–1000. doi:10.1126/science.abl6618
26. Oikonomou G, Altermatt M, Zhang RW, et al. The serotonergic raphe promote sleep in zebrafish and mice. Neuron. 2019;103(4):686–701.e8. doi:10.1016/j.neuron.2019.05.038
27. Cespuglio R. Serotonin: its place today in sleep preparation, triggering or maintenance. Sleep Med. 2018;49:31–39. doi:10.1016/j.sleep.2018.05.034
28. Kjaerby C, Andersen M, Hauglund N, et al. Memory-enhancing properties of sleep depend on the oscillatory amplitude of norepinephrine. Nat Neurosci. 2022;25(8):1059–1070. doi:10.1038/s41593-022-01102-9
29. Mitchell HA, Weinshenker D. Good night and good luck: norepinephrine in sleep pharmacology. Biochem Pharmacol. 2010;79(6):801–809. doi:10.1016/j.bcp.2009.10.004
30. McCarter SJ, Hagen PT, St Louis EK, et al. Physiological markers of sleep quality: a scoping review. Sleep Med Rev. 2022;64:101657. doi:10.1016/j.smrv.2022.101657
31. Besedovsky L, Lange T, Haack M. The sleep-immune crosstalk in health and disease. Physiol Rev. 2019;99(3):1325–1380. doi:10.1152/physrev.00010.2018
32. Sateia MJ. International classification of sleep disorders-third edition: highlights and modifications. Chest. 2014;146(5):1387–1394. doi:10.1378/chest.14-0970
33. Morphy H, Dunn KM, Lewis M, Boardman HF, Croft PR. Epidemiology of insomnia: a longitudinal study in a UK population. Sleep. 2007;30(3):274–280.
34. Jansson-Fröjmark M, Boersma K. Bidirectionality between pain and insomnia symptoms: a prospective study. Br J Health Psychol. 2012;17(2):420–431. doi:10.1111/j.2044-8287.2011.02045.x
35. Lautenbacher S, Kundermann B, Krieg JC. Sleep deprivation and pain perception. Sleep Med Rev. 2006;10(5):357–369. doi:10.1016/j.smrv.2005.08.001
36. Habib G, Khazin F, Artul S, Aloisi AM. The effect of medical cannabis on pain level and quality of sleep among rheumatology clinic outpatients. Pain Res Manag. 2021;2021:1756588. doi:10.1155/2021/1756588
37. Leys LJ, Chu KL, Xu J, et al. Disturbances in slow-wave sleep are induced by models of bilateral inflammation, neuropathic, and postoperative pain, but not osteoarthritic pain in rats. Pain. 2013;154(7):1092–1102. doi:10.1016/j.pain.2013.03.019
38. Cardis R, Lecci S, Fernandez LM, et al. Cortico-autonomic local arousals and heightened somatosensory arousability during NREMS of mice in neuropathic pain. Elife. 2021;10. doi:10.7554/eLife.65835
39. Alexandre C, Latremoliere A, Ferreira A, et al. Decreased alertness due to sleep loss increases pain sensitivity in mice. Nat Med. 2017;23(6):768–774. doi:10.1038/nm.4329
40. Chen S, Xie Y, Li Y, et al. Sleep deprivation and recovery sleep affect healthy male resident’s pain sensitivity and oxidative stress markers: the medial prefrontal cortex may play a role in sleep deprivation model. Front Mol Neurosci. 2022;15:937468. doi:10.3389/fnmol.2022.937468
41. Nosek K, Leppert W, Puchała Ł, Łoń K. Efficacy and safety of topical morphine: a narrative review. Pharmaceutics. 2022;14(7):1499. doi:10.3390/pharmaceutics14071499
42. Sanchez GA, Jutkiewicz EM, Ingram S, Smrcka AV. Coincident regulation of PLCβ signaling by Gq-coupled and μ-opioid receptors opposes opioid-mediated antinociception. Mol Pharmacol. 2022;102(6):269–279. doi:10.1124/molpharm.122.000541
43. Roeckel LA, Utard V, Reiss D, et al. Morphine-induced hyperalgesia involves mu opioid receptors and the metabolite morphine-3-glucuronide. Sci Rep. 2017;7(1):10406. doi:10.1038/s41598-017-11120-4
44. Leppä M, Korvenoja A, Carlson S, et al. Acute opioid effects on human brain as revealed by functional magnetic resonance imaging. Neuroimage. 2006;31(2):661–669. doi:10.1016/j.neuroimage.2005.12.019
45. Higginbotham JA, Markovic T, Massaly N, Morón JA. Endogenous opioid systems alterations in pain and opioid use disorder. Front Syst Neurosci. 2022;16:1014768. doi:10.3389/fnsys.2022.1014768
46. Bai X, Batallé G, Balboni G, Pol O. Hydrogen sulfide increases the analgesic effects of µ- and Δ-opioid receptors during neuropathic pain: pathways implicated. Antioxidants. 2022;11(7):1321. doi:10.3390/antiox11071321
47. Ziegler D, Papanas N, Schnell O, et al. Current concepts in the management of diabetic polyneuropathy. J Diabetes Investig. 2021;12(4):464–475. doi:10.1111/jdi.13401
48. Celik M, Labuz D, Henning K, et al. Leukocyte opioid receptors mediate analgesia via Ca(2+)-regulated release of opioid peptides. Brain Behav Immun. 2016;57:227–242. doi:10.1016/j.bbi.2016.04.018
49. Robertson RV, Crawford LS, Meylakh N, et al. Regional hypothalamic, amygdala, and midbrain periaqueductal gray matter recruitment during acute pain in awake humans: a 7-Tesla functional magnetic resonance imaging study. Neuroimage. 2022;259:119408. doi:10.1016/j.neuroimage.2022.119408
50. Presto P, Neugebauer V. Sex differences in CGRP regulation and function in the amygdala in a rat model of neuropathic pain. Front Mol Neurosci. 2022;15:928587. doi:10.3389/fnmol.2022.928587
51. Zhu Y, Sun M, Liu P, Shao W, Xiong M, Xu B. Perioperative stress prolong post-surgical pain via miR-339-5p targeting oprm1 in the amygdala. Korean J Pain. 2022;35(4):423–432. doi:10.3344/kjp.2022.35.4.423
52. Skinner GO, Damasceno F, Gomes A, de Almeida OM. Increased pain perception and attenuated opioid antinociception in paradoxical sleep-deprived rats are associated with reduced tyrosine hydroxylase staining in the periaqueductal gray matter and are reversed by L-dopa. Pharmacol Biochem Behav. 2011;99(1):94–99. doi:10.1016/j.pbb.2011.04.009
53. Smith MT, Mun CJ, Remeniuk B, et al. Experimental sleep disruption attenuates morphine analgesia: findings from a randomized trial and implications for the opioid abuse epidemic. Sci Rep. 2020;10(1):20121. doi:10.1038/s41598-020-76934-1
54. Tomim DH, Pontarolla FM, Bertolini JF, et al. The pronociceptive effect of paradoxical sleep deprivation in rats: evidence for a role of descending pain modulation mechanisms. Mol Neurobiol. 2016;53(3):1706–1717. doi:10.1007/s12035-014-9059-0
55. Holst SC, Bersagliere A, Bachmann V, Berger W, Achermann P, Landolt HP. Dopaminergic role in regulating neurophysiological markers of sleep homeostasis in humans. J Neurosci. 2014;34(2):566–573. doi:10.1523/JNEUROSCI.4128-13.2014
56. Ueno T, Tomita J, Tanimoto H, et al. Identification of a dopamine pathway that regulates sleep and arousal in Drosophila. Nat Neurosci. 2012;15(11):1516–1523. doi:10.1038/nn.3238
57. Wisor JP, Nishino S, Sora I, Uhl GH, Mignot E, Edgar DM. Dopaminergic role in stimulant-induced wakefulness. J Neurosci. 2001;21(5):1787–1794. doi:10.1523/JNEUROSCI.21-05-01787.2001
58. Moderie C, Carrier J, Dang-Vu TT. Les troubles du sommeil chez les patients atteints d'un trouble neurocognitif [Sleep disorders in patients with a neurocognitive disorder]. Encephale. 2022;48(3):325–334. French. doi:10.1016/j.encep.2021.08.014
59. Li C, Liu S, Lu X, Tao F. Role of descending dopaminergic pathways in pain modulation. Curr Neuropharmacol. 2019;17(12):1176–1182. doi:10.2174/1570159X17666190430102531
60. Edwards S, Callicoatte CN, Barattini AE, et al. Pramipexole treatment attenuates mechanical hypersensitivity in male rats experiencing chronic inflammatory pain. Neuropharmacology. 2022;208:108976. doi:10.1016/j.neuropharm.2022.108976
61. Li C, Sugam JA, Lowery-Gionta EG, et al. Mu opioid receptor modulation of dopamine neurons in the periaqueductal gray/dorsal raphe: a role in regulation of pain. Neuropsychopharmacology. 2016;41(8):2122–2132. doi:10.1038/npp.2016.12
62. Taylor NE, Pei J, Zhang J, et al. The Role of glutamatergic and dopaminergic neurons in the periaqueductal gray/dorsal raphe: separating analgesia and anxiety. eNeuro. 2019;6(1):ENEURO.0018–18.2019. doi:10.1523/ENEURO.0018-18.2019
63. Lau BK, Winters BL, Vaughan CW. Opioid presynaptic disinhibition of the midbrain periaqueductal grey descending analgesic pathway. Br J Pharmacol. 2020;177(10):2320–2332. doi:10.1111/bph.14982
64. Fu YT, Mao CJ, Ma LJ, et al. Pain correlates with sleep disturbances in parkinson’s disease patients. Pain Pract. 2018;18(1):29–37. doi:10.1111/papr.12578
65. Thompson T, Gallop K, Correll CU, et al. Pain perception in Parkinson’s disease: a systematic review and meta-analysis of experimental studies. Ageing Res Rev. 2017;35:74–86. doi:10.1016/j.arr.2017.01.005
66. Asakura W, Matsumoto K, Ohta H, Watanabe H. REM sleep deprivation decreases apomorphine-induced stimulation of locomotor activity but not stereotyped behavior in mice. Gen Pharmacol. 1992;23(3):337–341. doi:10.1016/0306-3623(92)90092-X
67. Vanini G. Nucleus accumbens: a novel forebrain mechanism underlying the increase in pain sensitivity caused by rapid eye movement sleep deprivation. Pain. 2018;159(1):5–6. doi:10.1097/j.pain.0000000000001073
68. Sardi NF, Tobaldini G, Morais RN, Fischer L. Nucleus accumbens mediates the pronociceptive effect of sleep deprivation: the role of adenosine A2A and dopamine D2 receptors. Pain. 2018;159(1):75–84. doi:10.1097/j.pain.0000000000001066
69. Kato T, Mitsukura Y, Yoshida K, Mimura M, Takata N, Tanaka KF. Oscillatory population-level activity of dorsal raphe serotonergic neurons is inscribed in sleep structure. J Neurosci. 2022;42(38):7244–7255. doi:10.1523/JNEUROSCI.2288-21.2022
70. Vaseghi S, Arjmandi-Rad S, Eskandari M, Ebrahimnejad M, Kholghi G, Zarrindast MR. Modulating role of serotonergic signaling in sleep and memory. Pharmacol Rep. 2022;74(1):1–26. doi:10.1007/s43440-021-00339-8
71. Zant JC, Leenaars CH, Kostin A, Van Someren EJ, Porkka-Heiskanen T. Increases in extracellular serotonin and dopamine metabolite levels in the basal forebrain during sleep deprivation. Brain Res. 2011;1399:40–48. doi:10.1016/j.brainres.2011.05.008
72. Azizi H, Hwang J, Suen V, et al. Sleep deprivation induces changes in 5-HT actions and 5-HT(1A) receptor expression in the rat hippocampus. Neurosci Lett. 2017;655:151–155. doi:10.1016/j.neulet.2017.06.053
73. Zhao X, Ozols AB, Meyers KT, et al. Acute sleep deprivation upregulates serotonin 2A receptors in the frontal cortex of mice via the immediate early gene Egr3. Mol Psychiatry. 2022;27(3):1599–1610. doi:10.1038/s41380-021-01390-w
74. Ye H, Ji M, Wang C, et al. Integrated functional neuroimaging, monoamine neurotransmitters, and behavioral score on depressive tendency in intensive care unit medical staffs induced by sleep deprivation after night shift work. Front Psychiatry. 2022;13:848709. doi:10.3389/fpsyt.2022.848709
75. Lee YC, Nassikas NJ, Clauw DJ. The role of the central nervous system in the generation and maintenance of chronic pain in rheumatoid arthritis, osteoarthritis and fibromyalgia. Arthritis Res Ther. 2011;13(2):211. doi:10.1186/ar3306
76. Li CJ, Zhang LG, Liu LB, et al. Inhibition of spinal 5-HT3 receptor and spinal dorsal horn neuronal excitability alleviates hyperalgesia in a rat model of parkinson’s disease. Mol Neurobiol. 2022;59(12):7253–7264. doi:10.1007/s12035-022-03034-8
77. Chuck AJ, Swannell AJ, House AO, Pownall R. The effects of dothiepin on subjects with rheumatoid arthritis and depression. Rheumatology. 2000;39(12):1425–1427. doi:10.1093/rheumatology/39.12.1425
78. Kim W, Chung Y, Choi S, Min BI, Kim SK. Duloxetine protects against oxaliplatin-induced neuropathic pain and spinal neuron hyperexcitability in rodents. Int J Mol Sci. 2017;18(12). doi:10.3390/ijms18122626
79. Siracusa R, Paola RD, Cuzzocrea S, Impellizzeri D. Fibromyalgia: pathogenesis, mechanisms, diagnosis and treatment options update. Int J Mol Sci. 2021;22(8):3891. doi:10.3390/ijms22083891
80. Wei H, Ma A, Wang YX, Pertovaara A. Role of spinal 5-HT receptors in cutaneous hypersensitivity induced by REM sleep deprivation. Pharmacol Res. 2008;57(6):469–475. doi:10.1016/j.phrs.2008.05.007
81. Thompson KI, Chau M, Lorenzetti MS, Hill LD, Fins AI, Tartar JL. Acute sleep deprivation disrupts emotion, cognition, inflammation, and cortisol in young healthy adults. Front Behav Neurosci. 2022;16:945661. doi:10.3389/fnbeh.2022.945661
82. Hunt CA, Smith MT, Mun CJ, Irwin MR, Finan PH. Trait positive affect buffers the association between experimental sleep disruption and inflammation. Psychoneuroendocrinology. 2021;129:105240. doi:10.1016/j.psyneuen.2021.105240
83. Irwin MR, Olmstead R, Kruse J, Breen EC, Haque R. Association of interleukin-8 and risk of incident and recurrent depression in long-term breast cancer survivors. Brain Behav Immun. 2022;105:131–138. doi:10.1016/j.bbi.2022.07.003
84. Raff H, Phillips J, Simpson P, Weisman SJ, Hainsworth KR. Interaction of chronic pain, obesity and time of day on cortisol in female human adolescents. Stress. 2022;25(1):331–336. doi:10.1080/10253890.2022.2142778
85. Fahmawi A, Khalifeh M, Alzoubi KH, Rababa’h AM. The effects of acute and chronic sleep deprivation on the immune profile in the rat. Curr Mol Pharmacol. 2022.
86. Dykstra-Aiello C, Koh KMS, Nguyen J, Xue M, Roy S, Krueger JM. A wake-like state in vitro induced by transmembrane TNF/soluble TNF receptor reverse signaling. Brain Behav Immun. 2021;94:245–258. doi:10.1016/j.bbi.2021.01.036
87. Oles V, Koh KMS, Dykstra-Aiello CJ, et al. Sleep- and time of day-linked RNA transcript expression in wild-type and IL1 receptor accessory protein-null mice. J Appl Physiol. 2020;128(6):1506–1522. doi:10.1152/japplphysiol.00839.2019
88. Chennaoui M, Gomez-Merino D, Drogou C, et al. Effects of exercise on brain and peripheral inflammatory biomarkers induced by total sleep deprivation in rats. J Inflamm. 2015;12(1):56. doi:10.1186/s12950-015-0102-3
89. Zielinski MR, Krueger JM. Sleep and innate immunity. Front Biosci. 2011;3(2):632–642.
90. Smith C, Trageser KJ, Wu H, et al. Anxiolytic effects of NLRP3 inflammasome inhibition in a model of chronic sleep deprivation. Transl Psychiatry. 2021;11(1):52. doi:10.1038/s41398-020-01189-3
91. Aghelan Z, Karima S, Khazaie H, et al. Interleukin-1α and tumor necrosis factor α as an inducer for reactive-oxygen-species-mediated NOD-like receptor protein 1/NOD-like receptor protein 3 inflammasome activation in mononuclear blood cells from individuals with chronic insomnia disorder. Eur J Neurol. 2022;29(12):3647–3657. doi:10.1111/ene.15540
92. Krueger JM, Clinton JM, Winters BD, et al. Involvement of cytokines in slow wave sleep. Prog Brain Res. 2011;193:39–47.
93. Hou J, Shen Q, Wan X, Zhao B, Wu Y, Xia Z. REM sleep deprivation-induced circadian clock gene abnormalities participate in hippocampal-dependent memory impairment by enhancing inflammation in rats undergoing sevoflurane inhalation. Behav Brain Res. 2019;364:167–176. doi:10.1016/j.bbr.2019.01.038
94. Krueger JM, Obál FJ, Fang J, Kubota T, Taishi P. The role of cytokines in physiological sleep regulation. Ann N Y Acad Sci. 2001;933:211–221. doi:10.1111/j.1749-6632.2001.tb05826.x
95. Kawasaki Y, Zhang L, Cheng JK, Ji RR. Cytokine mechanisms of central sensitization: distinct and overlapping role of interleukin-1beta, interleukin-6, and tumor necrosis factor-alpha in regulating synaptic and neuronal activity in the superficial spinal cord. J Neurosci. 2008;28(20):5189–5194. doi:10.1523/JNEUROSCI.3338-07.2008
96. Cao H, Zhang YQ. Spinal glial activation contributes to pathological pain states. Neurosci Biobehav Rev. 2008;32(5):972–983. doi:10.1016/j.neubiorev.2008.03.009
97. Moalem G, Tracey DJ. Immune and inflammatory mechanisms in neuropathic pain. Brain Res Rev. 2006;51(2):240–264. doi:10.1016/j.brainresrev.2005.11.004
98. Reeve AJ, Patel S, Fox A, Walker K, Urban L. Intrathecally administered endotoxin or cytokines produce allodynia, hyperalgesia and changes in spinal cord neuronal responses to nociceptive stimuli in the rat. Eur J Pain. 2000;4(3):247–257. doi:10.1053/eujp.2000.0177
99. Ma D, Wang X, Liu X, et al. Macrophage infiltration initiates RIP3/MLKL-dependent necroptosis in paclitaxel-induced neuropathic pain. Mediators Inflamm. 2022;2022:1567210. doi:10.1155/2022/1567210
100. Rakhshandeh H, Pourbagher-Shahri AM, Hasanpour M, Iranshahi M, Forouzanfar F. Effects of Capparis Spinosa extract on the neuropathic pain induced by chronic constriction injury in rats. Metab Brain Dis. 2022;37(8):2839–2852. doi:10.1007/s11011-022-01094-2
101. Ishikawa T, Miyagi M, Kamoda H, et al. Differences between tumor necrosis factor-α receptors types 1 and 2 in the modulation of spinal glial cell activation and mechanical allodynia in a rat sciatic nerve injury model. Spine. 2013;38(1):11–16. doi:10.1097/BRS.0b013e3182610fa9
102. Haack M, Sanchez E, Mullington JM. Elevated inflammatory markers in response to prolonged sleep restriction are associated with increased pain experience in healthy volunteers. Sleep. 2007;30(9):1145–1152. doi:10.1093/sleep/30.9.1145
103. Hirotsu C, Pedroni MN, Berro LF, Tufik S, Andersen ML. Nicotine and sleep deprivation: impact on pain sensitivity and immune modulation in rats. Sci Rep. 2018;8(1):13837. doi:10.1038/s41598-018-32276-7
104. Dai Y, Liu S, Chen J, Liu L, Zhou C, Zuo Y. Microglial responses and pain behaviors are exacerbated by chronic sleep deprivation in rats with chronic pain via neuroinflammatory pathways. Neuroscience. 2022;503:83–94. doi:10.1016/j.neuroscience.2022.09.004
105. Soya S, Sakurai T. Evolution of Orexin Neuropeptide system: structure and function. Front Neurosci. 2020;14:691. doi:10.3389/fnins.2020.00691
106. Inutsuka A, Yamanaka A. The physiological role of orexin/hypocretin neurons in the regulation of sleep/wakefulness and neuroendocrine functions. Front Endocrinol. 2013;4:18. doi:10.3389/fendo.2013.00018
107. Xiang X, Chen Y, Li KX, et al. Neuroanatomical basis for the orexinergic modulation of anesthesia arousal and pain control. Front Cell Neurosci. 2022;16:891631. doi:10.3389/fncel.2022.891631
108. McAlpine CS, Kiss MG, Rattik S, et al. Sleep modulates haematopoiesis and protects against atherosclerosis. Nature. 2019;566(7744):383–387. doi:10.1038/s41586-019-0948-2
109. Gao XB, Horvath TL. From molecule to behavior: hypocretin/orexin revisited from a sex-dependent perspective. Endocr Rev. 2022;43(4):743–760. doi:10.1210/endrev/bnab042
110. Chemelli RM, Willie JT, Sinton CM, et al. Narcolepsy in orexin knockout mice: molecular genetics of sleep regulation. Cell. 1999;98(4):437–451. doi:10.1016/S0092-8674(00)81973-X
111. Beuckmann CT, Sinton CM, Williams SC, et al. Expression of a poly-glutamine-ataxin-3 transgene in orexin neurons induces narcolepsy-cataplexy in the rat. J Neurosci. 2004;24(18):4469–4477. doi:10.1523/JNEUROSCI.5560-03.2004
112. Sutcliffe JG, de Lecea L. The hypocretins: excitatory neuromodulatory peptides for multiple homeostatic systems, including sleep and feeding. J Neurosci Res. 2000;62(2):161–168. doi:10.1002/1097-4547(20001015)62:2<161::AID-JNR1>3.0.CO;2-1
113. Xu A, Sakurai E, Kuramasu A, et al. Roles of hypothalamic subgroup histamine and orexin neurons on behavioral responses to sleep deprivation induced by the treadmill method in adolescent rats. J Pharmacol Sci. 2010;114(4):444–453. doi:10.1254/jphs.10177FP
114. Alakuijala A, Sarkanen T, Partinen M. Hypocretin-1 levels associate with fragmented sleep in patients with narcolepsy type 1. Sleep. 2016;39(5):1047–1050. doi:10.5665/sleep.5750
115. Murphy P, Moline M, Mayleben D, et al. Lemborexant, A Dual Orexin Receptor Antagonist (DORA) for the treatment of insomnia disorder: results from a Bayesian, Adaptive, Randomized, Double-Blind, Placebo-Controlled Study. J Clin Sleep Med. 2017;13(11):1289–1299. doi:10.5664/jcsm.6800
116. Hoever P, Dorffner G, Beneš H, et al. Orexin receptor antagonism, a new sleep-enabling paradigm: a proof-of-concept clinical trial. Clin Pharmacol Ther. 2012;91(6):975–985. doi:10.1038/clpt.2011.370
117. Roth T, Black J, Cluydts R, et al. Dual orexin receptor antagonist, almorexant, in elderly patients with primary insomnia: a Randomized, Controlled Study. Sleep. 2017;40(2). doi:10.1093/sleep/zsw034
118. Black J, Pillar G, Hedner J, et al. Efficacy and safety of almorexant in adult chronic insomnia: a randomized placebo-controlled trial with an active reference. Sleep Med. 2017;36:86–94. doi:10.1016/j.sleep.2017.05.009
119. Robinson CL, Supra R, Downs E, et al. Daridorexant for the treatment of insomnia. Health Psychol Res. 2022;10(3):37400. doi:10.52965/001c.37400
120. Ghalebandi S, Zareie F, Askari K, Yuzugulen J, Haghparast A. Intra-CA1 injection of orexin receptors antagonism attenuates the stress-induced analgesia in a rat acute pain model. Behav Brain Res. 2022;423:113785. doi:10.1016/j.bbr.2022.113785
121. Shakerinava P, Sayarnezhad A, Karimi-Haghighi S, Mesgar S, Haghparast A. Antagonism of the orexin receptors in the ventral tegmental area diminished the stress-induced analgesia in persistent inflammatory pain. Neuropeptides. 2022;96:102291. doi:10.1016/j.npep.2022.102291
122. Mobarakeh JI, Takahashi K, Sakurada S, et al. Enhanced antinociception by intracerebroventricularly and intrathecally-administered orexin A and B (hypocretin-1 and −2) in mice. Peptides. 2005;26(5):767–777. doi:10.1016/j.peptides.2005.01.001
123. den Boon FS, Sarabdjitsingh RA. Circadian and ultradian patterns of HPA-axis activity in rodents: significance for brain functionality. Best Pract Res Clin Endocrinol Metab. 2017;31(5):445–457. doi:10.1016/j.beem.2017.09.001
124. Spiga F, Walker JJ, Terry JR, Lightman SL. HPA axis-rhythms. Compr Physiol. 2014;4(3):1273–1298.
125. Balbo M, Leproult R, Van Cauter E. Impact of sleep and its disturbances on hypothalamo-pituitary-adrenal axis activity. Int J Endocrinol. 2010;2010:759234. doi:10.1155/2010/759234
126. Nollet M, Wisden W, Franks NP. Sleep deprivation and stress: a reciprocal relationship. Interface Focus. 2020;10(3):20190092. doi:10.1098/rsfs.2019.0092
127. Sgoifo A, Buwalda B, Roos M, Costoli T, Merati G, Meerlo P. Effects of sleep deprivation on cardiac autonomic and pituitary-adrenocortical stress reactivity in rats. Psychoneuroendocrinology. 2006;31(2):197–208. doi:10.1016/j.psyneuen.2005.06.009
128. Russell AL, Miller L, Yi H, Keil R, Handa RJ, Wu TJ. Knockout of the circadian gene, Per2, disrupts corticosterone secretion and results in depressive-like behaviors and deficits in startle responses. BMC Neurosci. 2021;22(1):5. doi:10.1186/s12868-020-00607-y
129. Galbo H, Kall L. Circadian variations in clinical symptoms and concentrations of inflammatory cytokines, melatonin, and cortisol in polymyalgia rheumatica before and during prednisolone treatment: a controlled, observational, clinical experimental study. Arthritis Res Ther. 2016;18(1):174. doi:10.1186/s13075-016-1072-4
130. Rosenberg H, Halman S, Yadav K. Polymyalgia rheumatica. CMAJ. 2021;193(46):E1770. doi:10.1503/cmaj.210541
131. Kreiner F, Langberg H, Galbo H. Increased muscle interstitial levels of inflammatory cytokines in polymyalgia rheumatica. Arthritis Rheum. 2010;62(12):3768–3775. doi:10.1002/art.27728
132. Wang S, Lim G, Zeng Q, et al. Expression of central glucocorticoid receptors after peripheral nerve injury contributes to neuropathic pain behaviors in rats. J Neurosci. 2004;24(39):8595–8605. doi:10.1523/JNEUROSCI.3058-04.2004
133. Le Coz GM, Anton F, Hanesch U. Glucocorticoid-mediated enhancement of glutamatergic transmission may outweigh anti-inflammatory effects under conditions of neuropathic pain. PLoS One. 2014;9(3):e91393. doi:10.1371/journal.pone.0091393
134. Chiu GS, Freund GG. Modulation of neuroimmunity by adenosine and its receptors: metabolism to mental illness. Metabolism. 2014;63(12):1491–1498. doi:10.1016/j.metabol.2014.09.003
135. Gvilia I, Suntsova N, Kostin A, et al. The role of adenosine in the maturation of sleep homeostasis in rats. J Neurophysiol. 2017;117(1):327–335. doi:10.1152/jn.00675.2016
136. Basheer R, Bauer A, Elmenhorst D, Ramesh V, McCarley RW. Sleep deprivation upregulates A1 adenosine receptors in the rat basal forebrain. Neuroreport. 2007;18(18):1895–1899. doi:10.1097/WNR.0b013e3282f262f6
137. Blanco-Centurion C, Xu M, Murillo-Rodriguez E, et al. Adenosine and sleep homeostasis in the Basal forebrain. J Neurosci. 2006;26(31):8092–8100. doi:10.1523/JNEUROSCI.2181-06.2006
138. Hao JW, Qiao WL, Li Q, et al. A1 adenosine receptor activation inhibits P2X3 receptor-mediated ATP currents in rat dorsal root ganglion neurons. Mol Neurobiol. 2022;59(11):7025–7035. doi:10.1007/s12035-022-03019-7
139. Martins DF, Mazzardo-Martins L, Cidral-Filho FJ, Stramosk J, Santos AR. Ankle joint mobilization affects postoperative pain through peripheral and central adenosine A1 receptors. Phys Ther. 2013;93(3):401–412. doi:10.2522/ptj.20120226
140. Yamaoka G, Horiuchi H, Morino T, Miura H, Ogata T. Different analgesic effects of adenosine between postoperative and neuropathic pain. J Orthop Sci. 2013;18(1):130–136. doi:10.1007/s00776-012-0302-0
141. Hambrecht-Wiedbusch VS, Gabel M, Liu LJ, Imperial JP, Colmenero AV, Vanini G. Preemptive caffeine administration blocks the increase in postoperative pain caused by previous sleep loss in the rat: a potential role for preoptic adenosine A2A receptors in sleep-pain interactions. Sleep. 2017;40(9). doi:10.1093/sleep/zsx116
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Intravenous Lidocaine for Treatment of Chronic Pain: A Retrospective Cohort Study
Horvat S, Staffhorst B, Cobben JHMG
Journal of Pain Research 2022, 15:3459-3467
Published Date: 28 October 2022
Tapentadol: A Review of Experimental Pharmacology Studies, Clinical Trials, and Recent Findings
Alshehri FS
Drug Design, Development and Therapy 2023, 17:851-861
Published Date: 21 March 2023
A Randomized, Double-Blind, Placebo-Controlled Trial to Evaluate the Therapeutic Effect of Magnesium-L-Threonate Supplementation for Persistent Pain After Breast Cancer Surgery
Ni Y, Deng F, Yu S, Zhang J, Zhang X, Huang D, Zhou H
Breast Cancer: Targets and Therapy 2023, 15:495-504
Published Date: 25 July 2023
Association Between Chronic Pain, Coping Strategies, and Sleep Quality in Rural Chinese Older Adults: A Cross-Sectional Study
Tao X, Cheng C, Bai J
Clinical Interventions in Aging 2025, 20:1005-1014
Published Date: 12 July 2025
Review of Physiology, Pathology, and Pharmacology of Voltage-Gated Sodium Channels in Regards to the Potential Role of NaV 1.8 Selective Inhibitor, Suzetrigine, for Chronic Pain Management
Lam CM, Robinson CL, Aiyer R, Mata R, Xie K, Slitzky M, Latif H, Braun EB, Schatman ME, Gilligan C, Soin A, Latif U
Journal of Pain Research 2026, 19:577002
Published Date: 2 February 2026