Back to Journals » Drug Design, Development and Therapy » Volume 17
The Protective Effect of Selenium Nanoparticles in Osteoarthritis: In vitro and in vivo Studies
Authors Li Y, Zhu S, Luo J, Tong Y, Zheng Y, Ji L, He Z, Jing Q, Huang J, Zhang Y, Bi Q
Received 4 February 2023
Accepted for publication 21 April 2023
Published 22 May 2023 Volume 2023:17 Pages 1515—1529
DOI https://doi.org/10.2147/DDDT.S407122
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Yong Li,1,2,* Senbo Zhu,2,* Junchao Luo,2 Yu Tong,2 Yixuan Zheng,1 Lichen Ji,2 Zeju He,2 Qiangan Jing,1 Jiaqing Huang,2 Yinjun Zhang,1 Qing Bi2
1College of Biotechnology and Bioengineering, Zhejiang University of Technology, Hangzhou, Zhejiang, 310014, People’s Republic of China; 2Laboratory Medicine Center, Department of Laboratory Medicine, Zhejiang Provincial People’s Hospital, Affiliated People’s Hospital, Hangzhou Medical College, Hangzhou, Zhejiang, 310014, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yinjun Zhang, College of Biotechnology and Bioengineering, Zhejiang University of Technology, Hangzhou, Zhejiang, 310014, People’s Republic of China, Tel +86-13735494556, Email [email protected] Qing Bi, Laboratory Medicine Center, Department of Laboratory Medicine, Zhejiang Provincial People’s Hospital, Affiliated People’s Hospital, Hangzhou Medical College, Hangzhou, Zhejiang, 310014, People’s Republic of China, Tel +86-13588302991, Email [email protected]
Introduction: Osteoarthritis (OA) is a common chronic joint disease characterized by articular cartilage degeneration. OA usually manifests as joint pain, limited mobility, and joint effusion. Currently, the primary OA treatment is non-steroidal anti-inflammatory drugs (NSAIDs). Although they can alleviate the disease’s clinical symptoms and signs, the drugs have some side effects. Selenium nanoparticles (SeNPs) may be an alternative to relieve OA symptoms.
Materials and Results: We confirmed the anti-inflammatory effect of selenium nanoparticles (SeNPs) in vitro and in vivo experiments for OA disease in this study. In vitro experiments, we found that SeNPs could significantly reduce the expression of nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), the major inflammatory factors, and had significant anti-inflammatory and anti-arthritic effects. SeNPs can inhibit reactive oxygen species (ROS) production and increased glutathione peroxidase (GPx) activity in interleukin-1beta (IL-1β)-stimulated cells. Additionally, SeNPs down-regulated matrix metalloproteinase-13 (MMP-13) and thrombospondin motifs 5 (ADAMTS-5) expressions, while up-regulated type II collagen (COL-2) and aggrecan (ACAN) expressions stimulated by IL-1β. The findings also indicated that SeNPs may exert their effects through suppressing the NF-κB p65 and p38/MAPK pathways. In vivo experiments, the prevention of OA development brought on by SeNPs was demonstrated using a DMM model.
Discussion: Our results suggest that SeNPs may be a potential anti-inflammatory agent for treating OA.
Keywords: osteoarthritis, NF-κB p65 signaling pathway, p38/MAPK signaling pathway, selenium nanoparticles
Introduction
Osteoarthritis (OA) is a prevalent chronic degenerative joint disease that is more common in middle-aged and elderly people.1 Its primary characteristics are subchondral sclerosis, joint swelling, synovitis, and gradual loss of articular cartilage.2 The clinical manifestations of OA are slow joint pain, tenderness, stiffness, joint swelling, limited mobility, and joint deformity, which seriously hinders work and affects human life quality.3,4 Inflammation and catabolism are closely related to the occurrence and development of OA.5 The potential pathogenesis of OA is still elusive.6 Articular cartilage degradation is primarily associated with the protein overproduction that breaks down cartilage, including matrix metalloproteinases (MMPs), a disintegrin and metalloproteinase with thrombospondin motifs (ADAMTS), and other catabolic enzymes.7 Inflammatory factors, such as IL-1β, play a vital role in bone and joint damage and inflammation.8 The production of ADAMTS and MMPs can be increased by IL-1β, stimulating extracellular matrix (ECM) degradation, and decreasing aggrecan (ACAN) and type II collagen (COL-2) synthesis.9,10
Meanwhile, IL-1β can induce the release of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), which can create nitric oxide (NO) and prostaglandin E2 (PGE2).11 IL-1β can induce inflammatory cytokines interleukin (IL)-6 and NO expressions with excessive reactive oxygen species (ROS) production.12 ROS or free radicals cause oxidative stress, the primary pathophysiology of OA.13 Increasing evidence suggests that neutralizing IL-1β-induced inflammatory mediators may help to treat OA.14 Patients with advanced OA are treated with total knee replacement, whereas patients with mild disease are mostly treated with nonsteroidal anti-inflammatory drugs (NSAIDs).15 Although these treatments can improve a patient’s range of motion and clinical symptoms, they do not slow or reverse the degenerative process, and long-term side effects are significant.16,17 This has prompted researchers to investigate alternate solutions.
Selenium is vital to human health due to its anti-inflammatory, potent antioxidant, and antiviral properties.18,19 Selenium provides several health advantages and is crucial for thyroid hormone metabolism, male reproduction, the immune system, anti-inflammatory processes, and removing free radicals.20–22 According to studies, a selenium deficiency can cause growth retardation, bone and cartilage metabolism dysfunction, and joint anomalies.23 Selenium can lessen oxidative stress-induced cartilage damage by enhancing chondrocytes’ antioxidant activity, removing ROS, and reducing the negative effects of ROS on cartilage homeostasis.24 Selenium in the form of selenomethionine inhibits the pro-inflammatory genes activated by IL-1β, thereby lowering the inflammatory response.25 Numerous research has demonstrated the anti-inflammatory properties of selenium.26,27 Selenium bioavailability and toxicity are the primary factors restricting its application.28 Selenium nanoparticles (SeNPs) have attracted attention due to their unique physical and chemical properties.29 Nanotechnology has shown its potential to treat OA in drug delivery, biological scaffolds, genetic engineering, and lubricants.30 Nanoscale materials exhibit new properties different from isolated atoms and blocky materials.31 The smaller the nanoparticle, the greater its activity.32 With the decrease of size, the ratio of surface to volume increases sharply, which enhances the interaction between nanomaterials and biological systems and improves their functions.33 In addition to pain relief, nanoparticle therapy is also expected to alleviate cartilage deterioration and even promote regeneration.34 Furthermore, the nanoparticles have excellent bone conductivity, biocompatibility, and biological functions that promote cell adhesion, proliferation, and osteogenic differentiation.35 Previous studies have demonstrated that highly dispersed selenium (called selenium nanoparticles) has similar biological activities and effects to sodium selenite and selenomethionine, yet with lower toxicity and acceptable bioavailability for inducing selenoproteins.36
This study utilizes chondrocytes and a mouse OA model to assess the anti-inflammatory effects of SeNPs generated by the microbial reduction approach. We assessed the anti-inflammatory effect of SeNPs on OA development using a mouse experiment model in vivo. We verified the protective effect of SeNPs on chondrocyte damage caused by IL-1β in this study. We also discuss the mechanism of inhibiting related pathways to delay OA development.
Materials and Methods
Materials and Reagents
Recombinant mouse IL-1β was purchased from Novoprotein Scientific Inc. (Shanghai, China). Collagenase II, 0.25% trypsin, Penicillin–streptomycin solution (100 x), SYBR Green master mix, and cDNA Synthesis reagent were purchased from Yi Sheng Biotechnology (Shanghai) Co., LTD. Medium (DMEM)/F12 was purchased from Shanghai Xiaopeng Biological Technology Co., LTD. Australia Origin Fetal Bovine Serum was purchased from Hangzhou Ouyuan Biotechnology Co., LTD. Phosphate-buffered saline (PBS) was purchased from Sangon Biotech (Shanghai) Co., Ltd. RNA-Quick Purification kit was purchased from Shanghai Yi Shan Biological Technology Co., LTD. Reactive Oxygen Species (ROS) Assay Kit, Phosphatase inhibitor mixture A, Hoechst 33342 were purchased from Beyotime Institute of Biotechnology. Reduced glutathione (GSH) assay kit was purchased from Nanjing Jiancheng Bioengineering Institute. PageRuler™ Prestained Protein Ladder and Pierce™ BCA were purchased from Thermo Fisher Scientific. EDTA Decalcification solution was purchased from SenBeiJia Biological Technology. A 4% universal tissue fixative was purchased from Wuhan Xavier Biotechnology. A 20 × TBST Buffer was purchased from Solarbio (Beijing, China). ELISA kits were purchased from Wuhan yunclone Technology. RIPA Lysis Buffer and PMSF were purchased from Hangzhou Verde Biotechnology.
Preparation and Characterization of SeNPs
SeNPs were synthesized by microbial reduction using Na2SeO3 as the starting precursor. Saccharomyces cerevisiae was inoculated into YPD medium (yeast extract 1%, peptone 2%, glucose 2%) for activation, and then incubated in a shaker at 30 °C for 12 h, from which 10% was inoculated into a shaker containing 30 mL medium with 100 g/L mother liquor. The shaker was incubated for 12 h, and a certain amount of sodium selenite was added until the final concentration of the medium was 2 g/L. Distilled water was added and the resulting precipitate was mixed evenly. The medium was incubated at 30 ℃ for 48 h, and then autolysis at 40 °C for 24 h. After ultrasonic crushing, SeNPs were separated by centrifugation. Particle size and zeta potential were measured by dynamic light scattering (Malvern, United States). The morphology of SeNPs particles was observed by transmission electron microscopy (Hitachi, Japan). The particle composition was determined by Energy Dispersion X-ray spectroscopy (Shimadzu, Japan).
Isolation and Culture of Primary Chondrocytes
This study was approved by the Animal Ethical and Welfare Committee of Zhejiang Provincial People’s Hospital (Hangzhou, China). All procedures associated with the care of animals were performed according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee. Wild-type embryonic young mice born 1–2 days ago had their cartilage tissue removed from the knee joint, cut into small pieces, thoroughly washed with sterilized PBS, centrifuged at 1000 rpm for 3 minutes three times, discarded the supernatant, added to 12-well culture plates, and digested with type II collagenase at a concentration of 2 mg/mL for four hours before single cells were collected by short-term centrifugation. In DMEM/F12 with 10% fetal bovine serum and 1% antibiotic (penicillin/streptomycin) at 37 °C and 5% CO2, chondrocytes were cultivated in a 100 mm petri dish. To prevent phenotypic alterations, F1 cells were subcultured when the cell density reached 80% and employed in the experiment.
Cell Viability Assay
CCK-8 reagent was used to test the impact of SeNPs on primary chondrocyte viability following the manufacturer’s recommendations. Chondrocytes were inoculated in 96-well plates at a cell density of 5 × 103 for each well. After 12 h, chondrocytes were treated with different concentrations of SeNPs (10 nM, 20 nM, 50 nM, 100 nM, 200 nM, 400 nM, 800 nM, 1200 nM, 1600 nM, 2000 nM) for 24 h or 48 h. Change to medium containing 10% CCK‐8 reagent and incubate away from light, then measure the absorbance at 450 nm with a microplate reader (Synergy LX, BioTek, United States).
Glutathione Peroxidase Activity
Glutathione peroxidase (GPx) activity in cell lysates was measured according to the instructions of the kit. The principle of determination is that dithiodinitrobenzoic acid reacts with sulfhydryl compounds to produce a yellow compound, which can be quantified by colorimetric measurement.
Measurement of IL-6, NO, Reactive Oxygen Species
Intracellular ROS production was determined using 2, 7-dichlorodi-hydrofluorescein diacetate (DCFH-DA) fluorescence intensity changes in chondrocytes treated with IL-1β and different concentrations of SeNPs. The cells were incubated in a serum-free medium containing 10 μm DCFH-DA at 37 °C for 30 min. The maximum excitation wavelength was 480 nm, and the maximum emission wavelength was 525 nm. The ROS generation was measured by flow cytometry (BD Biosciences, New Jersey, United States) according to the fluorescence signal intensity. The concentration of IL-6 in the cell culture supernatant was determined using an ELISA kit and the level of nitric oxide (NO) in the cell culture supernatant was determined using the Griess Reaction kit according to the instructions.
RNA Extraction and Real-Time Quantitative PCR
Total RNA was extracted from mouse chondrocytes using the RNA-Quick Purification kit according to the manufacturer’s instructions and then reverse transcribed into cDNA using cDNA Synthesis reagent, which was diluted 1:10 with nuclease-free water. Real-time quantitative Polymerase Chain Reaction (Rt-qPCR) using the Roche LightCycler 480 Detection System. The real-time PCR program was as follows: initial denaturation at 95 °C for 2 min, denaturation at 95 °C for 15s, annealing at 60 °C for 30s, and extension at 72 °C for 30s, a total of 40 cycles. Target mRNA levels were normalized to GAPDH levels, and the relative gene expression was determined by the 2−ΔΔCT method. The primers of COL-2, ACAN, MMP-13, ADAMTS-5, iNOS, COX-2, and GAPDH were designed by NCBI Primer-Blast tool, which was as follows: COL-2: forward: CAGGATGCCCGAAAATTAGGG, reverse: ACCACGATCACCTCTGGGT; ACAN: forward: GTGGAGCCGTGTTTCCAAG, reverse: AGATGCTGTTGACTCGAACCT; MMP-13: forward: TGTTTGCAGAGCACTACTTGAA, reverse: CAGTCACCTCTAAGCCAAAGAAA; ADAMTS-5: forward: CCACGACCCTCAAGAACTTTT, reverse: CTGCATCGTAGTGCTCTTCG; iNOS: forward: GGAGTGACGGCAAACATGACT, reverse: TCGATGCACAACTGGGTGAAC; COX-2: forward: TTCCAATCCATGTCAAAACCGT, reverse: AGTCCGGGTACAGTCACACTT; GAPDH: forward: TGACCTCAACTACATGGTCTACA, reverse: CTTCCCATTCTCGGCCTTG.
Western Blotting Analysis
The mouse chondrocytes were washed with ice-cold PBS twice, and then the protein was extracted from the mixture of RIPA buffer and PMSF (100:1). The protein content was determined by high-speed centrifugation at 4 °C (14,400 rpm, 15 min) after 30 min of ice cracking. The protein concentration of the samples was determined by the BCA protein assay kit. The same amount of protein was separated by prefabricated gel with denatured protein and transferred to the PVDF membrane (Millipore, United States). After sealing the membrane with 5% non-fat milk at room temperature for 2 h, the membrane was washed with TBST three times for 10 min and incubated with primary antibody: COL-2 (diluted 1:3000, #28459-1-AP, Proteintech Group), ACAN (diluted 1:1000, #DF7561, Affinity, Cincinnati, OH, USA), MMP-13 (diluted 1:2500, #18165-1-AP, Proteintech Group), anti-ADAMTS-5 (diluted 1:1000, ab41037, Abcam Corporation, USA), anti-iNOS (diluted 1:1000, ab178945, Abcam Corporation, USA), COX-2 (diluted 1:2000, #12375-1-AP, Proteintech Group), GAPDH (diluted 1:50,000, #60004-1-IG, Proteintech Group), p38 MAPK (diluted 1:1000, #14064-1-AP, Proteintech Group), Phospho-p38 MAPK (diluted 1:2000, #28796-1-AP, Proteintech Group), IκBα (diluted 1:2000, #10268-1-AP, Proteintech Group), anti-p-IκBα (diluted 1:1000, ab133462, Abcam Corporation, USA), NF-κB p65 (diluted 1:2000, #10745-1-AP, Proteintech Group), anti-NF-κB p-p65 (diluted 1:1000, ab76302, Abcam Corporation, USA), Lamin B1 (diluted 1:3000, #12987-1-AP, Proteintech Group), at 4 °C overnight. The membrane was washed with TBST three times for 10 min each and incubated with the secondary antibody (goat Anti-Rabbit IgG, diluted 1:3000, #ZB-2301, Zhongshan Golden Bridge Biotechnology; goat Anti-Mouse IgG, diluted 1:50,000, #HA1006, HuaAn Biotechnology) at room temperature for 2 h. Then, the membranes were washed with TBST three times, reactive protein bands were found using enhanced chemiluminescence (ECL), and the intensity of these membranes was determined using ChemiDocTM MP Imaging System (Bio-Rad, United States).
Immunofluorescence Assay
F1 generation chondrocytes were cultured in confocal dishes for 12 h, treated with IL-1β and SeNPs or without SeNPs for 24 h, washed once with PBS, then fixed with 4% paraformaldehyde for 15 min, washed three times with PBS, and washed for 3 min each time. The samples were incubated with 0.2%Trition X-100 (prepared with PBS) for 30 min, then added with 5% normal goat serum, and blocked for 30 min at room temperature. Chondrocytes were mixed with COL-2 (1:100), and MMP-13 (1:100) antibody was incubated at 4 °C overnight. On the next day, the cells were washed with PBS, incubated with fluorescein-conjugated goat anti-rabbit IgG antibody (diluted 1:200, #ZF-0311, Zhongshan Golden Bridge) for 1 h, then stained with Hoechst for 15 min, washed with PBS for 3 times, and observed under the laser confocal microscopy (Leika TCS SP8 Confocal, LEIKA Inc, California, United States).
Mice OA Models
Thirty 8-week-old C57BL/6 male wild type (WT) mice were purchased from Slack Laboratory Animal Co., LTD., Shanghai. All experiments were carried out by the requirements of the animal Ethics and Use Committee of Zhejiang Provincial People’s Hospital. All procedures associated with the care of animals were performed according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee. The medial meniscus (DMM) was surgically destroyed, and the tibial ligament of the medial meniscus was severed to establish a mouse model of osteoarthritis. Then, they were randomly divided into the sham control group (n=10), the OA group (n=10), and the SeNPs treatment group (n=10). Four weeks after the DMM operation, 5μL solution containing 50 nM SeNPs was injected into the knee joint of the mouse in the SeNPs treatment group, and the administration lasted for 5 weeks every week. All mice were sacrificed 8 weeks after DMM surgery to preserve joint tissue for further analysis.
Experimental Design
For measuring the cartilage protection of various concentrations of SeNPs in vitro, chondrocytes were treated with IL-1β and different concentrations of SeNPs (20 nM, and 50 nM). The negative control group was treated with 5 ng/mL IL-1β, while the blank control group received no treatment. The cells were taken 24 h after being incubated.
For in vivo experiments, the medial collateral ligament and medial meniscus of the mouse knee joint were removed as previously mentioned, destabilizing the joint stability of experimental animals. Each mouse in the OA and SeNPs therapy groups had the medial meniscus of their knee joint surgically destabilized. In the sham control group, the knee joint was cut without removing the medial collateral ligament and the medial meniscus. Every five days, SeNPs were injected into the joint cavity in the therapy group. Sterile normal saline was administered identically to both the OA group and the sham control group. For specimen preparation and histological examination, joint tissue was saved.
Histopathologic Analysis and Immunohistochemistry
Samples obtained from mouse knee joints were immobilized in 4% paraformaldehyde for 48 h and then recalculated in 10% EDTA solution for 4 weeks. Then, the tissues were embedded in paraffin wax and cut into slices with a thickness of 6μm. The sections were stained with hematoxylin and Safranin-O/Fast Green and the histological changes of stained sections were evaluated by an optical microscope and the International Society for the Study of Osteoarthritis (OARSI) scoring system. COL-2 and MMP-13 were incubated at 4 °C for the duration of the night in immunohistochemistry. After that, tissue sections were treated with secondary antibodies for two hours at room temperature. The sections were examined and photographed under an optical microscope (Nikon Ti, Japan). Calculate the percentage of positive cells by counting the number of stained cells that are positive across the entire joint surface of each specimen.
Statistical Analysis
All data are presented as mean ± standard deviation. Multiple groups of data were compared using the one-way analysis of variance (ANOVA). Comparisons between the two sets of data were made using the Student’s t-test. A P-value < 0.05 is considered to be a significant difference.
Results
Particle Characterization of SeNPs
We measured the size of nanoparticles using dynamic light scattering to investigate the particle characteristics of SeNPs. According to findings, SeNPs have an average particle size of 222.9 nm (Figure 1A). The Zeta potential of SeNPs was measured with an average value of −18.7 mV (Figure 1B). Transmission electron microscopy indicated that nanoparticles were spherical (Figure 1C), and the particles were monodispersed in the solution without agglomeration. Energy Dispersion X-ray spectroscopy exhibits that the particles primarily comprise selenium (Figure 1D), indicating that the synthesized nanoparticles had high purity. These results lay a foundation for the follow-up experiments.
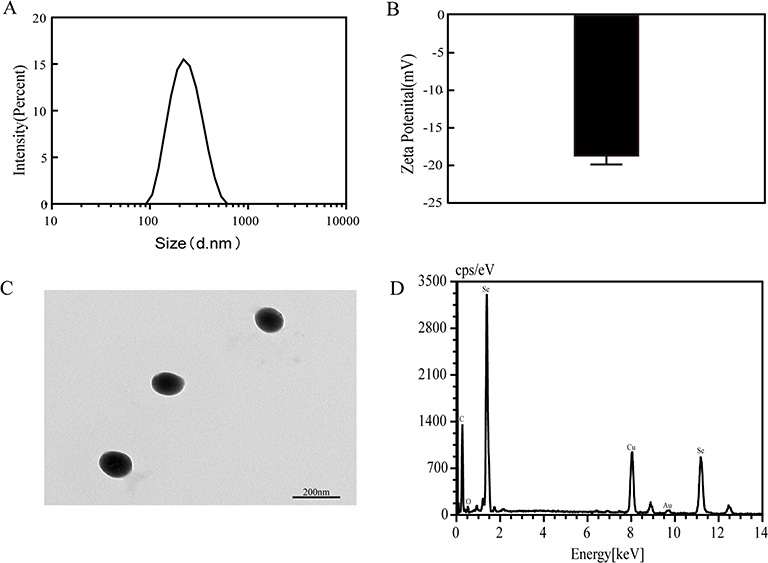 |
Figure 1 Particle characterization of SeNPs. (A) The corresponding size distribution of SeNPs. (B) The average zeta potential value of SeNPs. (C) A transmission electron microscopy image of SeNPs. (D) Energy Dispersion X-ray spectroscopy (EDX) of SeNPs. Abbreviation: SeNPs, selenium nanoparticles. |
Effect of SeNPs on the Viability of Mouse Chondrocytes
Chondrocytes were exposed to a range of concentrations (10 nM, 20 nM, 50 nM, 100 nM, 200 nM, 400 nM, 800 nM, 1200 nM, 1600 nM, and 2000 nM) of SeNPs for 24 or 48 h to assess the cytotoxic effects of SeNPs on mouse chondrocytes. The CCK-8 assay was used to measure cell viability. SeNPs have no cytotoxicity to chondrocytes after 24 h of treatment when the concentration is less than 50 nM. In contrast, cell viability was significantly inhibited at concentrations of 100 nM or higher (Figure 2A and Figure 2B). Similar results were observed when chondrocytes were treated with SeNPs for 48 h compared to untreated cells (P < 0.05). These results indicated that SeNPs lower concentrations do not affect the cell vitality of chondrocytes. Therefore, 20 nM and 50 nM of SeNPs were selected for subsequent experiments.
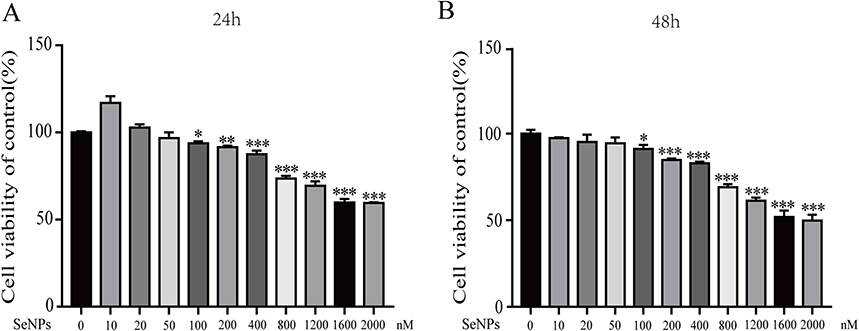 |
Figure 2 Effect of SeNPs on the viability of mouse chondrocytes. Cells were treated with different concentrations of SeNPs (10 nM, 20 nM, 50 nM, 100 nM, 200 nM, 400 nM, 800 nM, 1200 nM, 1600 nM, 2000 nM) for 24 h (A) or 48 h (B) and analyzed using a CCK-8 assay. All data are presented as mean ± standard deviation (n=3). *P < 0.05 vs the control group; **P < 0.01 vs the control group; ***P < 0.001 vs the control group. Abbreviation: SeNPs, selenium nanoparticles. |
Effect of SeNPs on ROS Generation and Glutathione Peroxidase Activity
Oxidative stress is an imbalance between the production of ROS and scavenging by the antioxidant defense system and substantially impacts OA.37 IL-1β was used for stimulating the generation of ROS and oxidative stress in chondrocytes to explore whether SeNPs could alleviate oxidative stress. Chondrocytes treated with IL-1β exhibited a significantly higher ROS production than untreated controls (Figure 3A). However, SeNPs subdued the ROS production induced by IL-1β. Glutathione peroxidase (GPx), a crucial part of the body’s antioxidant defense mechanism, is strongly regulated by selenium in its expression and activity.38 Then, we detected GPx activity to confirm the effect of SeNPs on GPx in chondrocytes after treatment with or without IL-1β. The results revealed that GPx activity was dramatically suppressed after IL-1β administration compared to the control group. However, SeNPs can significantly reduce the inhibitory effect of IL-1β on GPx activity (Figure 3B). These results suggest that SeNPs can counteract excessive ROS generation and increase GPx activity in chondrocytes.
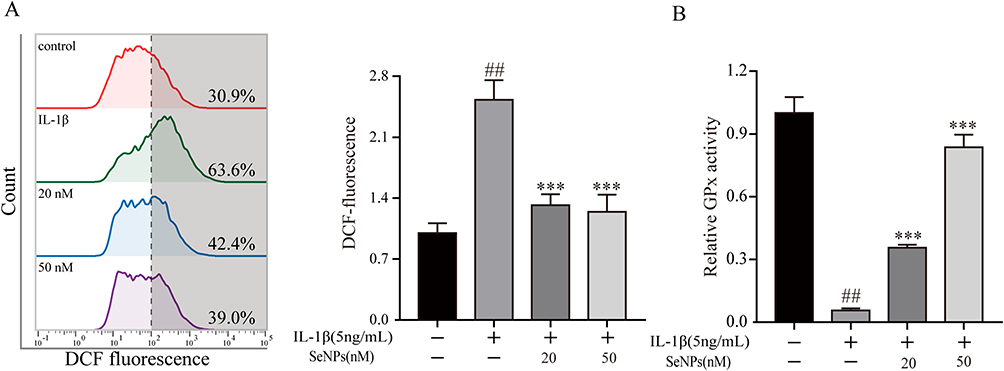 |
Figure 3 Effects of SeNPs on the level of reactive oxygen species (ROS) and the activity of glutathione peroxidase (GPx) in mouse chondrocytes induced by IL-1β. (A) Intracellular ROS levels were detected by DCFH-DA staining and flow cytometry. (B) The activity of GPx was determined by the glutathione (GSH) assay kit. All data are presented as mean ± standard deviation (n=3). ##P < 0.0001 vs compared with the control group; ***P < 0.001 vs compared with IL‐1β group. Abbreviations: SeNPs, selenium nanoparticles; IL, interleukin; ROS, reactive oxygen species; GPx, glutathione peroxidase; DCFH-DA, 2, 7-dichlorodi-hydrofluorescein diacetate. |
Effects of SeNPs on IL-1β-Induced ECM Synthesis and Degradation in Mouse Chondrocytes
MMPs and ADAMTS overexpression leads to ECM degradation, resulting in reduced collagen and aggrecan synthesis in the pathogenesis of OA.39 Western blot and Rt-qPCR analysis were conducted to evaluate whether SeNPs have an impact on the expression of COL-2, ACAN, MMP-13, and ADAMTS-5 with or without IL-1β treatment to confirm whether SeNPs can protect against IL-1β-induced ECM degradation in chondrocytes. After IL-1β treatment, mRNA and protein expression of ADAMTS-5 and MMP-13 increased significantly, and SeNPs can decrease their up-regulation caused by IL-1β (Figure 4A–E). Meanwhile, SeNPs dramatically decreased the mRNA and protein downregulation of COL-2 and ACAN caused by IL-1β stimulation. Immunofluorescence results consistently presented that SeNPs markedly inhibited COL-2 protein degradation and MMP-13 expression (Figure 4F and Figure 4G). Thus, these results reveal that SeNPs prevent ECM degradation in chondrocytes.
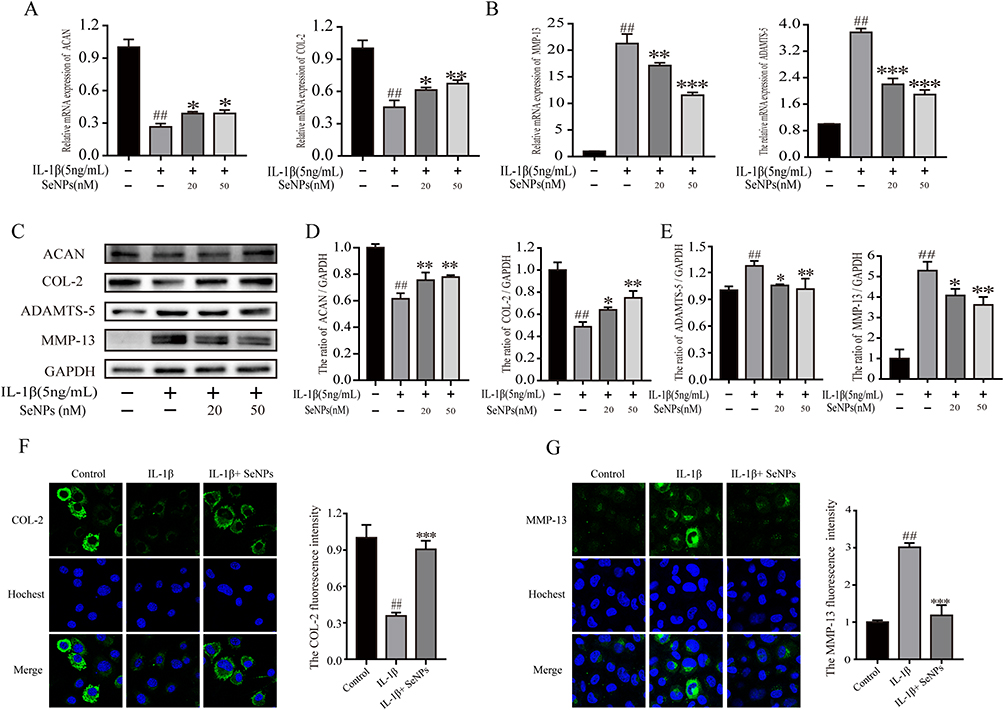 |
Figure 4 Effect of SeNPs on extracellular matrix degradation of mouse chondrocytes induced by IL-1β. (A and B) The mRNA expression levels of ACAN, COL-2, ADAMTS-5, and MMP-13 in mouse chondrocytes were measured by Rt-qPCR. (C–E) The protein expression of ACAN, COL-2, ADAMTS-5, and MMP-13 was examined by Western blot analysis. (F and G) Immunofluorescence was used to assess the amounts of COL-2 and MMP-13 protein expression. All data are presented as mean ± standard deviation (n=3). ##P < 0.0001 vs compared with the control group; *P < 0.05 vs compared with IL‐1β group; **P < 0.01 compared with IL‐1β group; ***P < 0.001 vs compared with IL‐1β group. Abbreviations: SeNPs, selenium nanoparticles; IL, interleukin; Rt-qPCR, Real-time quantitative Polymerase Chain Reaction; ACAN, aggrecan; COL-2, type II collagen; ADAMTS, a disintegrin and metalloproteinase thrombospondin motifs; MMP, matrix metalloproteinase. |
Effect of SeNPs on IL-6, NO, iNOS, and COX-2 Expression of Mouse Chondrocytes
Next, we investigated the effects of SeNPs on IL-1β-induced production of inflammatory mediators, including COX-2, iNOS, IL-6, and NO, in chondrocytes. The cells were treated with different concentrations of SeNPs combined with IL-1β for 24 h. Griess reaction and ELISA kit were used to determine the concentrations of endogenous NO and IL-6 in the cell suspension. The results revealed that NO and IL-6 levels decreased significantly dose-dependently when combined with SeNPs compared to chondrocytes treated with IL-1β alone (Figure 5A). Then, we evaluated the effect of SeNPs on iNOS and COX-2 expressions in IL-1β-stimulated mouse chondrocytes using Western blot and Rt-qPCR. The findings demonstrated that IL-1β dramatically boosted COX-2 and iNOS expressions at the mRNA and protein levels. However, iNOS and COX-2 expressions produced by IL-1β were reduced by SeNPs in a dose-dependent manner (Figure 5B and Figure 5C). These results indicated that SeNPs could inhibit the production of IL-1β-induced inflammatory mediators and cytokines in chondrocytes in a concentration-dependent manner.
 |
Figure 5 Effects of SeNPs on the expression of NO, iNOS, COX2, and IL-6 in mouse chondrocytes induced by IL-1β. (A) The production of IL-6 was measured by ELISA and NO was detected by Griess. (B) The mRNA expression levels of iNOS and COX-2 were measured by Rt-qPCR. (C) The protein expression of iNOS and COX‐2 assessed by Western blot and quantification analysis. All data are presented as mean ± standard deviation (n=3). ##P < 0.0001 vs compared with the control group; *P < 0.05 vs compared with IL‐1β group; **P < 0.01 compared with IL‐1β group; ***P < 0.001 vs compared with IL‐1β group. Abbreviations: SeNPs, selenium nanoparticles; IL, interleukin; Rt-qPCR, real-time quantitative Polymerase Chain Reaction; NO, nitric oxide; iNOS, nitric oxide synthase; COX-2, cyclooxygenase-2. |
Effects of SeNPs on IL-1β-Induced NF-κB and P38/MAPK Pathways
We examined the phosphorylation status of signaling proteins in primary IL-1β-stimulated chondrocytes to determine whether the inhibitory effect of SeNPs was associated with IL-1β-induced changes in signaling pathways. Western blot demonstrated that IL-1β stimulates translocation of NF-κB p65 from the cytoplasm to the nucleus and IκBα degradation in chondrocytes. SeNPs significantly inhibited the IL-1β regulation on mouse chondrocytes dose-dependently (Figure 6A). The p38/MAPK pathway is also activated when IL-1β stimulates OA chondrocytes, resulting in MMPs production and inflammatory mediator release. IL-1β greatly increased p38 phosphorylation, while SeNPs treatment suppressed it (Figure 6B). Consequently, SeNPs may regulate NF-κB and p38/MAPK signaling pathways to limit MMPs expression and act as an anti-inflammatory.
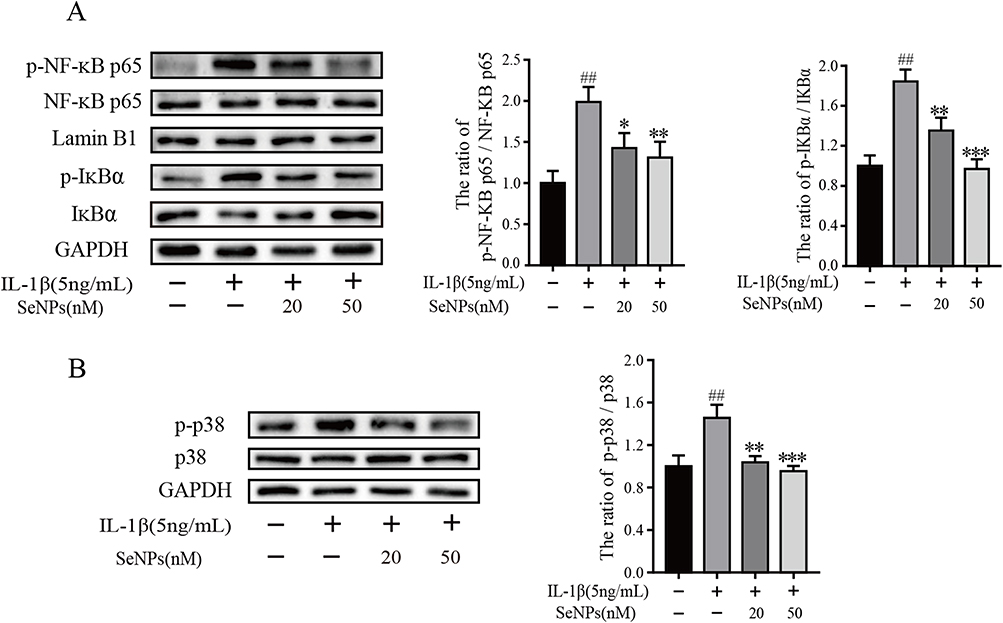 |
Figure 6 Effect of SeNPs on IL-1β-induced NF-κB and p38 signaling pathways. Chondrocytes were pretreated with different concentrations of SeNPs for 2 h and then stimulated with IL-1β (5 ng/mL) for 30 min. The expression of phosphorylated NF-κB p65 (p-NF-κB p65), total NF-κB p65, phosphorylated 38 (p-p38), total p38, phosphorylated IκBα (p-IκBα), total IκBα were determined by Western blot and quantification analysis (A and B). All data are presented as mean ± standard deviation (n=3). ##P < 0.0001 vs compared with the control group; *P < 0.05 vs compared with IL‐1β group; **P < 0.01 compared with IL‐1β group; ***P < 0.001 vs compared with IL‐1β group. Abbreviations: SeNPs, selenium nanoparticles; IL, interleukin; IκB, inhibitor of kappa B; NF-κB, nuclear factor κB. |
Histopathological Analysis and Morphological Evaluation
DMM OA model was established in mice to study whether SeNPs have a protective effect on the cartilage developed by OA in vivo. The sham surgery control group’s cartilage surface was stained with Safranin-O and hematoxylin-eosin, exhibiting a smooth and complete cartilage surface (Figure 7A), while the OA group’s cartilage surface was severely damaged. Aggrecan decreased significantly, and cell density decreased. Comparatively, the SeNPs treatment group alleviated the destruction of articular cartilage, maintained aggrecan in cartilage, and reduced the damaging effects. The Safranin O staining results revealed that the OARSI score of the OA group was significantly higher than that of the sham operation control group, whereas the OARSI score of the SeNPs group was significantly lower than that of the OA group (Figure 7B). Immunohistochemistry was also used to assess COL-2 and MMP-13 expressions in the cartilage matrix. COL-2 positive cells were reduced, MMP-13 positive cells increased in the OA group compared to the sham surgery control group. After receiving a SeNPs treatment, COL-2 positive cells rose, while MMP-13 positive cells reduced (Figure 7C and Figure 7D). The results demonstrated that SeNPs could effectively shield chondrocytes of arthritis mice, reduce the loss of ECM, promote ECM synthesis protein (COL-2) expression, and inhibit the ECM degrading enzyme (MMP-l3) expression.
 |
Figure 7 SeNPs alleviated the development of OA in a surgically induced mouse DMM model. (A) Histological analysis was performed by safranin O staining, Haematoxylin-Eosin, and evaluation of OA. Stain, H-E, safranin O, magnification, 10×; scale bar, 200 μm. (B) Sham surgery, DMM, and DMM + SeNPs were scored by the International Society for Osteoarthritis Research (OARSI). (C and D) The effects of SeNPs on cartilage matrix degradation in OA model mice were evaluated by immunohistochemistry of COL-2 and MMP‐13. Stain, immunohistochemistry, magnification, 40×; scale bar, 50 μm. All data are presented as mean ± standard deviation (n=10). ##P < 0.0001 vs Sham group; **P < 0.01 vs DMM group; ***P < 0.001 vs DMM group. Abbreviations: SeNPs, selenium nanoparticles; OA, osteoarthritis; DMM, destabilization of the medial meniscus; COL-2, type II collagen; MMP, matrix metalloproteinase; DMM, the destabilization of medial meniscus. |
Discussion
OA, a common chronic degenerative joint ailment, is primarily caused by an imbalance between catabolic and anabolic processes.40 Currently, the primary goal of OA treatment is to reduce OA patients’ complaints of joint pain, swelling, and dysfunction.41,42 Other OA medications and NSAIDs are only momentarily beneficial, and excessive use might have negative side effects.43 Therefore, it is necessary to discover safe and effective strategies to alleviate the clinical symptoms of OA. Nanoparticle technology, widely used in the healthcare industry, can reduce the negative effects on the body.44 Micronutrient selenium, needed for chondrogenesis, has anti-inflammatory properties.45 Selenium is a dual-purpose element since it has slightly greater toxic concentrations than those needed for basic physiological processes.46 The toxicity of high concentrations limits the long-term use of selenium products to treat Kashin-Beck disease, osteoarthropathy, or other illnesses.47 Meanwhile, nano-sized elemental selenium exhibits potential biological effects and reduces the risk of selenium toxicity.48 Therefore, this study created a type of SeNPs utilizing less harmful and more bioavailable microorganisms. We confirmed that the effect of SeNPs (0–50 nM) on mouse OA chondrocytes could not be attributed to cytotoxicity. We discovered that SeNPs significantly inhibited iNOS, COX-2, MMP-13, and ADAMTS-5 expressions in IL-1β-induced inflammatory responses, promoting COL-2 and ACAN synthesis in chondrocytes. Furthermore, our findings demonstrated that SeNPs could effectively inhibit NF-κB p65 activation and p38/MAPK phosphorylation caused by IL-1β and had a protective effect on cartilage degradation in mice with DMM-induced OA.
ROS are oxygen-containing free radicals, including superoxide anion (O2−), hydrogen peroxide (H2O2), and hydroxyl radical (OH).49 Previous research has demonstrated that OA progression is significantly related to oxidative stress.50 High ROS concentrations can cause oxidative stress, resulting in mitochondrial dysfunction and a breakdown of the epigenetic controls over cell signaling and gene expression.51,52 Oxidative stress causes inflammation, chondrocyte apoptosis, cartilage degradation, subchondral bone dysfunction, and unbalanced bone resorption and remodeling.53,54 When ROS production increases, the cellular defense systems against oxidative stress are activated, effectively removing ROS molecules from the cell.55 GPxs are a family of selenium-dependent and independent antioxidant enzymes that catalyzes the conversion of harmful H2O2 and numerous hydroperoxides, including lipid peroxide and DNA peroxide, into water and alcohol, respectively.56 This process frequently employs GSH as a reducing agent, shielding biofilm and cellular components from oxidative stress.57 We stimulated chondrocytes with IL-1β to decrease glutathione concentration and increase ROS levels. When OA chondrocytes were treated with SeNPs, we observed a significant decrease in ROS levels while increasing GPx activity, indicating that SeNPs may protect chondrocytes by reducing ROS production while increasing GPx activity.
ACAN and COL-2 are the primary structural components of the chondrocyte extracellular matrix, and they are crucial to preserve structural integrity.58 ACAN and COL-2 degradation accelerate OA development.59 Therefore, OA might be treated by preventing ACAN and COL-2 degradation. MMPs are crucial regulators in OA development, preventing ACAN and COL-2 synthesis.60 Among all MMPs, MMP-13 plays a major role in OA progression due to its ability to break down COL-2, constituting the primary structure of the cartilage ECM.61 ADAMTS family plays a key role in cartilage degradation in OA.62 ADAMTS-5 is a key enzyme associated with arthritis and participates in the cleavage of ACAN in OA.63 IL-1β is a proinflammatory cytokine widely used to stimulate the OA microenvironment in vitro.64 We evaluated the protective impact of SeNPs on IL-1β-induced chondrocyte damage using IL-1β to produce an oxidative stress microenvironment. Our findings demonstrated that chondrocytes treated with IL-1β dramatically raised MMP-13 and ADAMTS-5 mRNA and protein expression levels, while ACAN and COL-2 mRNA and protein levels dropped. SeNPs can increase ACAN and COL-2 synthesis while inhibiting MMP-13 and ADAMTS-5 synthesis in cells.
According to a report, IL-1β promotes iNOS and COX-2 expressions in OA chondrocytes, which are responsible for NO and PGE2 up-regulation, respectively.65 The inflammatory mediators, NO and PGE2, are crucial in OA development.66 Previous research has demonstrated that decreasing inflammatory cytokines, including NO and PGE2, can weaken OA progression because NO and PGE2 can inhibit ECM synthesis and induce ECM degradation.67,68 Our results revealed that SeNPs inhibited IL-1β-induced NO and IL-6 production, as well as iNOS and COX-2 expression in chondrocytes. These findings suggested that SeNPs had anti-inflammatory effects on IL-1β stimulated chondrocytes.
NF-κB exists in the cytoplasm and binds to NF-κB (IκB) repressor.69 IL-1β stimulation can trigger phosphorylation of IκBα and p65, IκB protein is degraded by phosphorylation, and NF-κB p65 is transferred from cytoplasm to the nucleus to regulate inflammatory mediator production.70 The MAPKs family is divided into at least three components: extracellular signal-regulated kinase 1/2 (ERK 1/2), c-Jun interterminal kinase (JNK), and p38/MAPK, activated by phosphorylation of specific tyrosine and threonine residues by upstream kinases in response to inflammation and other extracellular signals.71–73 According to previous studies, NF-κB and MAPK are the most important transcription factors in regulating MMPs, COX-2, and iNOS expressions in OA.74,75 IL-1β can easily activate the MAPK pathway (ERK, P38, and JNK) and NF-κB p65 phosphorylation, inducing inflammatory mediator expressions and leading to ECM degradation.76 In this study, we treated chondrocytes with IL-1β enhanced p38/MAPK and IκBα phosphorylation, promoting the translocation of NF-κB p65 from the cytoplasm to the nucleus. However, SeNPs treatment suppressed IκBα and p38/MAPK phosphorylation and attenuated the nuclear translocation of NF-κB p65. The results presented that SeNPs could inhibit the inflammatory reaction induced by IL-1β via NF-κB and p38/MAPK signals, thus playing a protective role in chondrocytes.
Besides in vitro study, we employed the DMM model to evaluate the protective effect of SeNPs on cartilage degradation in vivo. The DMM model is widely used to evaluate the efficacy of drug treatment for OA due to its mechanical instability.77 A weekly intra-articular injection of SeNPs reduces cartilage degeneration during OA progression. The morphological and histological evaluation confirmed knee cartilage degeneration and structural changes were less severe in the SeNPs treatment group than in the control group. These in vivo results are consistent with the in vitro effects of SeNPs and support the consistency and reproducibility of this OA model.
Conclusion
Our results suggest that SeNPs inhibit IL-1β-induced inflammatory mediator expression by inhibiting NF-κB and p38/MAPK activation in chondrocytes (Figure 8). Meanwhile, SeNPs can increase the synthesis of COL-2 and ACAN. In vivo experiments, SeNPs can suppress ECM degradation and chondrocyte loss, and reduce the degree of articular cartilage irregularity and knee joint stenosis in mice model of OA. These findings suggest that SeNPs may be a potential anti-inflammatory agent for treating OA.
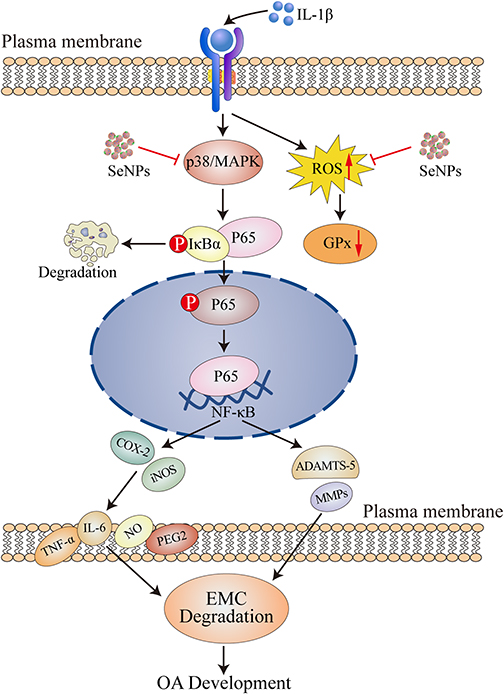 |
Figure 8 Molecular mechanism of SeNPs on IL-1β-induced mouse chondrocytes. Abbreviations: OA, osteoarthritis; SeNPs, selenium nanoparticles; IL, interleukin; MAPK, mitogen-activated protein kinase; ROS, reactive oxygen species; GPx, glutathione peroxidase; IκB, inhibitor of kappa B; NF-κB, nuclear factor κB; ACAN, aggrecan; COL-2, type II collagen; ADAMTS, a disintegrin and metalloproteinase thrombospondin motifs; MMP, matrix metalloproteinase; NO, nitric oxide; iNOS, nitric oxide synthase; COX-2, cyclooxygenase-2. |
Acknowledgments
This research was funded by grants from National Science Foundation of China (Grant No. 81672769), Health Technology Project of Zhejiang Province (2020KY029), Medical Health Science and Technology Project of Zhejiang Provincial Health Commission (2021KY028) and basic public welfare research plan of Zhejiang Province (LGF22H060029).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Kang DG, Lee HJ, Lee CJ, Park JS. Inhibition of the expression of matrix metalloproteinases in articular chondrocytes by resveratrol through affecting nuclear factor-kappa B signaling pathway. Biomol Ther. 2018;26(6):560–567. doi:10.4062/biomolther.2018.132
2. Li X, He P, Hou Y, et al. Berberine inhibits the interleukin-1 beta-induced inflammatory response via MAPK downregulation in rat articular chondrocytes. Drug Dev Res. 2019;80(5):637–645. doi:10.1002/ddr.21541
3. Lin Z, Fu C, Yan Z, et al. The protective effect of hesperetin in osteoarthritis: an in vitro and in vivo study. Food Funct. 2020;11(3):2654–2666. doi:10.1039/c9fo02552a
4. Park DR, Kim J, Kim GM, et al. Osteoclast-associated receptor blockade prevents articular cartilage destruction via chondrocyte apoptosis regulation. Nat Commun. 2020;11(1):4343. doi:10.1038/s41467-020-18208-y
5. Chen C, Yang M, Chen Y, et al. Astilbin-induced inhibition of the PI3K/AKT signaling pathway decelerates the progression of osteoarthritis. Exp Ther Med. 2020;20(4):3078–3083. doi:10.3892/etm.2020.9048
6. Mao L, Wu W, Wang M, et al. Targeted treatment for osteoarthritis: drugs and delivery system. Drug Deliv. 2021;28(1):1861–1876. doi:10.1080/10717544.2021.1971798
7. Wang Z, Xu M, Bai J, et al. Vildagliptin reduced extracellular matrix degradation in human primary chondrocytes. Eur J Pharmacol. 2019;844:49–55. doi:10.1016/j.ejphar.2018.11.020
8. Wang ZW, Chen L, Hao XR, et al. Elevated levels of interleukin-1β, interleukin-6, tumor necrosis factor-α and vascular endothelial growth factor in patients with knee articular cartilage injury. World J Clin Cases. 2019;7(11):1262–1269. doi:10.12998/wjcc.v7.i11.1262
9. Li X, Lin J, Ding X, et al. The protective effect of sinapic acid in osteoarthritis: in vitro and in vivo studies. J Cell Mol Med. 2019;23(3):1940–1950. doi:10.1111/jcmm.14096
10. Durigova M, Soucy P, Fushimi K, Nagase H, Mort JS, Roughley PJ. Characterization of an ADAMTS-5-mediated cleavage site in aggrecan in OSM-stimulated bovine cartilage. Osteoarthritis Cartilage. 2008;16(10):1245–1252. doi:10.1016/j.joca.2008.02.013
11. Fu Y, Lei J, Zhuang Y, Zhang K, Lu D. Overexpression of HMGB1 A-box reduced IL-1β-induced MMP expression and the production of inflammatory mediators in human chondrocytes. Exp Cell Res. 2016;349(1):184–190. doi:10.1016/j.yexcr.2016.10.014
12. Khan NM, Haseeb A, Ansari MY, Devarapalli P, Haynie S, Haqqi TM. Wogonin, a plant derived small molecule, exerts potent anti-inflammatory and chondroprotective effects through the activation of ROS/ERK/Nrf2 signaling pathways in human Osteoarthritis chondrocytes. Free Radic Biol Med. 2017;106:288–301. doi:10.1016/j.freeradbiomed.2017.02.041
13. Sutipornpalangkul W, Morales NP, Harnroongroj T. Free radicals in primary knee osteoarthritis. J Med Assoc Thai. 2009;92(Suppl 6):S268–74.
14. Ma Z, Piao T, Wang Y, Liu J. Astragalin inhibits IL-1β-induced inflammatory mediators production in human osteoarthritis chondrocyte by inhibiting NF-κB and MAPK activation. Int Immunopharmacol. 2015;25(1):83–87. doi:10.1016/j.intimp.2015.01.018
15. Ding QH, Ye CY, Chen EM, Zhang W, Wang XH. Emodin ameliorates cartilage degradation in osteoarthritis by inhibiting NF-κB and Wnt/β-catenin signaling in-vitro and in-vivo. Int Immunopharmacol. 2018;61:222–230. doi:10.1016/j.intimp.2018.05.026
16. Gundogdu G, Demirkaya Miloglu F, Gundogdu K, et al. Investigation of the efficacy of daidzein in experimental knee osteoarthritis-induced with monosodium iodoacetate in rats. Clin Rheumatol. 2020;39(8):2399–2408. doi:10.1007/s10067-020-04958-z
17. Scott DL, Berry H, Capell H, et al. The long-term effects of non-steroidal anti-inflammatory drugs in osteoarthritis of the knee: a randomized placebo-controlled trial. Rheumatology. 2000;39(10):1095–1101. doi:10.1093/rheumatology/39.10.1095
18. Wrobel JK, Power R, Toborek M. Biological activity of selenium: revisited. IUBMB Life. 2016;68(2):97–105. doi:10.1002/iub.1466
19. Malhotra S, Welling MN, Mantri SB, Desai K. In vitro and in vivo antioxidant, cytotoxic, and anti-chronic inflammatory arthritic effect of selenium nanoparticles. J Biomed Mater Res B Appl Biomater. 2016;104(5):993–1003. doi:10.1002/jbm.b.33448
20. Hariharan S, Dharmaraj S. Selenium and selenoproteins: it’s role in regulation of inflammation. Inflammopharmacology. 2020;28(3):667–695. doi:10.1007/s10787-020-00690-x
21. Effraimidis G, Wiersinga WM. Mechanisms in endocrinology: autoimmune thyroid disease: old and new players. Eur J Endocrinol. 2014;170(6):R241–52. doi:10.1530/eje-14-0047
22. Zubair M, Ali M, Ahmad M, Sajid M. Effect of Selenium and Vitamin E on cryopreservation of semen and reproductive performance of animals (a review). 2015.
23. Medeiros DM. Copper, iron, and selenium dietary deficiencies negatively impact skeletal integrity: a review. Exp Biol Med. 2016;241(12):1316–1322. doi:10.1177/1535370216648805
24. Li G, Cheng T, Yu X. The impact of trace elements on osteoarthritis. Front Med. 2021;8:771297. doi:10.3389/fmed.2021.771297
25. Cheng AWM, Bolognesi M, Kraus VB. DIO2 modifies inflammatory responses in chondrocytes. Osteoarthritis Cartilage. 2012;20(5):440–445. doi:10.1016/j.joca.2012.02.006
26. Vieira AT, Silveira KD, Arruda MC, et al. Treatment with Selemax®, a selenium-enriched yeast, ameliorates experimental arthritis in rats and mice. Br J Nutr. 2012;108(10):1829–1838. doi:10.1017/s0007114512000013
27. Kahya MC, Naziroğlu M, Çiğ B. Melatonin and selenium reduce plasma cytokine and brain oxidative stress levels in diabetic rats. Brain Inj. 2015;29(12):1490–1496. doi:10.3109/02699052.2015.1053526
28. Jia M, Zhang Y, Huang B, Zhang H. Source apportionment of selenium and influence factors on its bioavailability in intensively managed greenhouse soil: a case study in the east bank of the Dianchi Lake, China. Ecotoxicol Environ Saf. 2019;170:238–245. doi:10.1016/j.ecoenv.2018.11.133
29. Boostani A, Sadeghi AA, Mousavi SN, Chamani M, Kashan N. The effects of organic, inorganic, and nano-selenium on blood attributes in broiler chickens exposed to oxidative stress. Acta Scientiae Veterinariae. 2015;43(1):154.
30. Xiao L, Cui J, Sun Z, Liu Y, Zheng J, Dong Y. Therapeutic potential of nanotechnology-based approaches in osteoarthritis. Front Pharmacol. 2022;13:920824. doi:10.3389/fphar.2022.920824
31. El-Ghazaly MA, Fadel N, Rashed E, El-Batal A, Kenawy SA. Anti-inflammatory effect of selenium nanoparticles on the inflammation induced in irradiated rats. Can J Physiol Pharmacol. 2017;95(2):101–110. doi:10.1139/cjpp-2016-0183
32. Dong X, Price M, Dai Z, Xu M, Pelton R. Mineral-mineral particle collisions during flotation remove adsorbed nanoparticle flotation collectors. J Colloid Interface Sci. 2017;504:178–185. doi:10.1016/j.jcis.2017.05.050
33. Qiao K, Xu L, Tang J, et al. The advances in nanomedicine for bone and cartilage repair. J Nanobiotechnology. 2022;20(1):141. doi:10.1186/s12951-022-01342-8
34. Li X, Dai B, Guo J, et al. Nanoparticle-Cartilage Interaction: pathology-Based Intra-articular Drug Delivery for Osteoarthritis Therapy. Nanomicro Lett. 2021;13(1):149. doi:10.1007/s40820-021-00670-y
35. Wang Q, Mynar JL, Yoshida M, et al. High-water-content mouldable hydrogels by mixing clay and a dendritic molecular binder. Nature. 2010;463(7279):339–343. doi:10.1038/nature08693
36. Wang Y, Fu L. Forms of selenium affect its transport, uptake and glutathione peroxidase activity in the Caco-2 cell model. Biol Trace Elem Res. 2012;149(1):110–116. doi:10.1007/s12011-012-9395-8
37. Qin R, Sun J, Wu J, Chen L. Pyrroloquinoline quinone prevents knee osteoarthritis by inhibiting oxidative stress and chondrocyte senescence. Am J Transl Res. 2019;11(3):1460–1472.
38. Schnabel R, Lubos E, Messow CM, et al. Selenium supplementation improves antioxidant capacity in vitro and in vivo in patients with coronary artery disease The SElenium Therapy in Coronary Artery disease Patients (SETCAP) Study. Am Heart J. 2008;156(6):1201.e1–11. doi:10.1016/j.ahj.2008.09.004
39. Zheng W, Chen C, Zhang C, Cai L, Chen H. The protective effect of phloretin in osteoarthritis: an in vitro and in vivo study. Food Funct. 2018;9(1):263–278. doi:10.1039/c7fo01199g
40. Wu Y, Lu X, Shen B, Zeng Y. The therapeutic potential and role of miRNA, lncRNA, and circRNA in osteoarthritis. Curr Gene Ther. 2019;19(4):255–263. doi:10.2174/1566523219666190716092203
41. Moghimi N, Nasseri S, Ghafouri F, Jalili A. Frequency of growth differentiation factor 5 rs143383 and asporin D-repeat polymorphisms in patients with hand and knee osteoarthritis in Kurdistan province, Iran. Int J Rheum Dis. 2021;24(5):694–700. doi:10.1111/1756-185x.14097
42. Majeed M, Majeed S, Narayanan NK, Nagabhushanam K. A pilot, randomized, double-blind, placebo-controlled trial to assess the safety and efficacy of a novel Boswellia serrata extract in the management of osteoarthritis of the knee. Phytother Res. 2019;33(5):1457–1468. doi:10.1002/ptr.6338
43. Zhou K, Hu L, Liao W, Yin D, Rui F. Coptisine prevented IL-β-induced expression of inflammatory mediators in chondrocytes. Inflammation. 2016;39(4):1558–1565. doi:10.1007/s10753-016-0391-6
44. Ouvinha de Oliveira R, De santa maria LC, Barratt G. Nanomedicine and its applications to the treatment of prostate cancer. Ann Pharm Fr. 2014;72(5):303–316. doi:10.1016/j.pharma.2014.04.006
45. Shahabi R, Anissian A, Javadmoosavi SA, Nasirinezhad F. Protective and anti-inflammatory effect of selenium nano-particles against bleomycin-induced pulmonary injury in male rats. Drug Chem Toxicol. 2021;44(1):92–100. doi:10.1080/01480545.2018.1560466
46. Rocourt CR, Cheng WH. Selenium supranutrition: are the potential benefits of chemoprevention outweighed by the promotion of diabetes and insulin resistance? Nutrients. 2013;5(4):1349–1365. doi:10.3390/nu5041349
47. Zhang J, Wang X, Xu T. Elemental selenium at nano size (Nano-Se) as a potential chemopreventive agent with reduced risk of selenium toxicity: comparison with se-methylselenocysteine in mice. Toxicol Sci. 2008;101(1):22–31. doi:10.1093/toxsci/kfm221
48. Han J, Guo X, Lei Y, Dennis BS, Wu S, Wu C. Synthesis and characterization of selenium-chondroitin sulfate nanoparticles. Carbohydr Polym. 2012;90(1):122–126. doi:10.1016/j.carbpol.2012.04.068
49. Bhattacharjee S. ROS and oxidative stress: origin and implication. 2019.
50. Zahan OM, Serban O, Gherman C, Fodor D. The evaluation of oxidative stress in osteoarthritis. Clujul Med. 2020;93(1):74.
51. Portal-Núñez S, Esbrit P, Alcaraz MJ, Largo R. Oxidative stress, autophagy, epigenetic changes and regulation by miRNAs as potential therapeutic targets in osteoarthritis. Biochem Pharmacol. 2016;108:1–10. doi:10.1016/j.bcp.2015.12.012
52. Singh A, Kukreti R, Saso L, Kukreti S. Oxidative stress: a key modulator in neurodegenerative diseases. Molecules. 2019;24(8):1583. doi:10.3390/molecules24081583
53. Bolduc JA, Collins JA, Loeser RF. Reactive oxygen species, aging and articular cartilage homeostasis. Free Radic Biol Med. 2019;132:73–82. doi:10.1016/j.freeradbiomed.2018.08.038
54. Lepetsos P, Papavassiliou AG. ROS/oxidative stress signaling in osteoarthritis. Biochim Biophys Acta. 2016;1862(4):576–591. doi:10.1016/j.bbadis.2016.01.003
55. Ansari MY, Ahmad N, Haqqi TM. Oxidative stress and inflammation in osteoarthritis pathogenesis: role of polyphenols. Biomed Pharmacother. 2020;129:110452. doi:10.1016/j.biopha.2020.110452
56. Dabo AJ, Ezegbunam W, Wyman AE, et al. Targeting c-Src reverses accelerated GPX-1 mRNA decay in chronic obstructive pulmonary disease airway epithelial cells. Am J Respir Cell Mol Biol. 2020;62(5):598–607. doi:10.1165/rcmb.2019-0177OC
57. Vlahos R, Bozinovski S. Glutathione peroxidase-1 as a novel therapeutic target for COPD. Redox Rep. 2013;18(4):142–149. doi:10.1179/1351000213y.0000000053
58. Gao F, Zhang S. Salicin inhibits AGE-induced degradation of type II collagen and aggrecan in human SW1353 chondrocytes: therapeutic potential in osteoarthritis. Artif Cells Nanomed Biotechnol. 2019;47(1):1043–1049. doi:10.1080/21691401.2019.1591427
59. Yi H, Zheng Q, Jiang MM, et al. The effect of protease inhibitors on the induction of osteoarthritis-related biomarkers in bovine full-depth cartilage explants. PLoS One. 2015;10(4):25.
60. Zhang J, Zhang W, Shi J, Dai J, Shen SG. Dlx2 overexpression enhanced accumulation of type II collagen and aggrecan by inhibiting MMP13 expression in mice chondrocytes. Biochem Biophys Res Commun. 2018;503(2):528–535. doi:10.1016/j.bbrc.2018.05.066
61. Wang M, Sampson ER, Jin H, et al. MMP13 is a critical target gene during the progression of osteoarthritis. Arthritis Res Ther. 2013;15(1):R5. doi:10.1186/ar4133
62. Yang CY, Chanalaris A, Troeberg L. ADAMTS and ADAM metalloproteinases in osteoarthritis - looking beyond the ‘usual suspects’. Osteoarthritis Cartilage. 2017;25(7):1000–1009. doi:10.1016/j.joca.2017.02.791
63. Jiang L, Lin J, Zhao S, et al. ADAMTS5 in osteoarthritis: biological functions, regulatory network, and potential targeting therapies. Front Mol Biosci. 2021;8:703110. doi:10.3389/fmolb.2021.703110
64. Zeng RM. Cartilage destruction%Chondrocytes%Forkhead box M1 (FOXM1)%Inflammation%Interleukin-1β (IL-1β)%Osteoarthritis (OA). Int Immunopharmacol. 2018;68:74–80.
65. Chen X, Li Z, Hong H, et al. Xanthohumol suppresses inflammation in chondrocytes and ameliorates osteoarthritis in mice. Biomed Pharmacother. 2021;137:111238. doi:10.1016/j.biopha.2021.111238
66. Chen X, Zhang C, Wang X, Huo S. Juglanin inhibits IL-1β-induced inflammation in human chondrocytes. Artif Cells Nanomed Biotechnol. 2019;47(1):3614–3620. doi:10.1080/21691401.2019.1657877
67. Zheng W, Tao Z, Cai L, et al. Chrysin attenuates IL-1β-induced expression of inflammatory mediators by suppressing NF-κB in human osteoarthritis chondrocytes. Inflammation. 2017;40(4):1143–1154. doi:10.1007/s10753-017-0558-9
68. Qu Y, Shen Y, Teng L, et al. Chicoric acid attenuates tumor necrosis factor-α-induced inflammation and apoptosis via the Nrf2/HO-1, PI3K/AKT and NF-κB signaling pathways in C28/I2 cells and ameliorates the progression of osteoarthritis in a rat model. Int Immunopharmacol. 2022;111:109129. doi:10.1016/j.intimp.2022.109129
69. Lepetsos P, Papavassiliou KA, Papavassiliou AG. Redox and NF-κB signaling in osteoarthritis. Free Radic Biol Med. 2019;132:90–100. doi:10.1016/j.freeradbiomed.2018.09.025
70. Ozawa M, Nishida K, Yoshida A, et al. Hyaluronan suppresses mechanical stress-induced expression of catabolic enzymes by human chondrocytes via inhibition of IL-1β production and subsequent NF-κB activation. Inflamm Res. 2015;64(3–4):243–252. doi:10.1007/s00011-015-0804-2
71. Zhong HM, Ding QH, Chen WP, Luo RB. Vorinostat, a HDAC inhibitor, showed anti-osteoarthritic activities through inhibition of iNOS and MMP expression, p38 and ERK phosphorylation and blocking NF-κB nuclear translocation. Int Immunopharmacol. 2013;17(2):329–335. doi:10.1016/j.intimp.2013.06.027
72. Torres-Martínez M, Rubio-Infante N, García-Hernández AL, Nava-Acosta R, Ilhuicatzi-Alvarado D, Moreno-Fierros L. Cry1Ac toxin induces macrophage activation via ERK1/2, JNK and p38 mitogen-activated protein kinases. Int J Biochem Cell Biol. 2016;78:106–115. doi:10.1016/j.biocel.2016.06.022
73. Landeros RV, Jobe SO, Aranda-Pino G, Lopez GE, Zheng J, Magness RR. Convergent ERK1/2, p38 and JNK mitogen activated protein kinases (MAPKs) signalling mediate catecholoestradiol-induced proliferation of ovine uterine artery endothelial cells. J Physiol. 2017;595(14):4663–4676. doi:10.1113/jp274119
74. Jeong JW, Lee HH, Lee KW, et al. Mori folium inhibits interleukin-1β-induced expression of matrix metalloproteinases and inflammatory mediators by suppressing the activation of NF-κB and p38 MAPK in SW1353 human chondrocytes. Int J Mol Med. 2016;37(2):452–460. doi:10.3892/ijmm.2015.2443
75. Xi Y, Huang X, Tan G, et al. Protective effects of Erdosteine on interleukin-1β-stimulated inflammation via inhibiting the activation of MAPK, NF-κB, and Wnt/β-catenin signaling pathways in rat osteoarthritis. Eur J Pharmacol. 2020;873:172925. doi:10.1016/j.ejphar.2020.172925
76. Moon SM, Lee SA, Han SH, et al. Aqueous extract of Codium fragile alleviates osteoarthritis through the MAPK/NF-κB pathways in IL-1β-induced rat primary chondrocytes and a rat osteoarthritis model. Biomed Pharmacother. 2018;97:264–270. doi:10.1016/j.biopha.2017.10.130
77. Lorenz J, Grässel S. Experimental osteoarthritis models in mice. Methods Mol Biol. 2014;1194:401–419. doi:10.1007/978-1-4939-1215-5_23
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Network Analysis of Osteoarthritis Progression Using a Steiner Minimal Tree Algorithm
Xie Y, Shao F, Ji Y, Feng D, Wang L, Huang Z, Wu S, Sun F, Jiang H, Miyamoto A, Wang H, Zhang C
Journal of Inflammation Research 2024, 17:3201-3209
Published Date: 18 May 2024