Back to Journals » Journal of Inflammation Research » Volume 18
The Pomegranate Peel Polyphenols Protects Acne Vulgaris by Regulating Inflammation Through Notch/NF-κB Signal Pathway
Authors Wang Y ![]() , Zhu M
, Zhu M ![]() , Wu S, Zhao D
, Wu S, Zhao D ![]() , Bai Q, Liu Y
, Bai Q, Liu Y
Received 18 June 2025
Accepted for publication 24 September 2025
Published 10 October 2025 Volume 2025:18 Pages 14113—14129
DOI https://doi.org/10.2147/JIR.S545787
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Anish R. Maskey
Yun Wang,1,2 Mingfang Zhu,1 Shuhui Wu,1 Dan Zhao,1 Qi Bai,1 Ying Liu1
1Department of Dermatology, the Second Affiliated Hospital of Hunan University of Chinese Medicine, Changsha, People’s Republic of China; 2Department of Dermatology, the First Hospital of Hunan University of Chinese Medicine, Changsha, People’s Republic of China
Correspondence: Mingfang Zhu, Department of Dermatology, the Second Affiliated Hospital of Hunan University of Chinese Medicine, 233 Cai E North Road, Kaifu District, Changsha, People’s Republic of China, Tel +86-13907317358, Fax +86-0731-84917727, Email [email protected]
Background: Pomegranate peel is a kind of traditional Chinese medicine for treating acne. Its main active ingredient is pomegranate peel polyphenols(PPPs), but its mechanism is not clear. Inflammatory response is still the focus of current treatment. Interestingly, Notch signaling is considered to be a key regulator of macrophage activation, which can promote the occurrence and development of Acne vulgaris (AV) by activating nuclear factor kappa-B (NF-κB) to produce a large number of inflammatory factors.
Objective: This study aims to explore the potential mechanism of PPPs for treating AV.
Methods: An in-vivo model of acne was established by injecting SD rats with Cutibacterium acnes(C. acnes),and the RAW264.7 Cells were induced by lipopolysaccharide(LPS) in vitro. After treatment, the levels of inflammatory factors and Notch/NF-κB signaling pathway were observed.
Results: PPPs improved the levels of IL-1α, TNF-α, IL-6, IL-8 and IL-12 in serum and pathological skin lesions of SD rats induced by C. acnes, and decreased the expression of macrophages in skin lesions. Notch, NF-κB, IL-1α, IL-6, TNF-α protein and mRNA expression and NF-κB phosphorylation were inhibited.
Conclusion: PPPs can inhibit Notch/NF-κB signaling pathway, reduce inflammatory response, and exert anti-acne effects.
Keywords: acne vulgaris, Notch/NF-κB signaling pathway, pomegranate peel polyphenols, inflammation, RAW264.7 cells
Introduction
Acne vulgaris (AV) is a common chronic inflammatory disorder of the sebaceous follicles. It mainly involves the following four pathogenesis: inflammatory response, colonization of Cutibacterium acnes (C. acnes), increased lipid secretion and excessive keratosis of follicular orifice.1,2 Although the pathogenesis of AV has not been fully elucidated, inflammation and immune response are the main pathological features of AV, and the inflammatory response runs through the whole process of the disease.3 Macrophages are important cells of innate immunity, which play an important role in acne inflammation.4 Notch signaling is a key regulatory factor in the biological function of macrophages. Activation of Notch signaling can regulate the differentiation of macrophages to M1, thus promoting the occurrence and development of inflammation.5,6 Nuclear factor kappa-B (NF-κB) is a key player in the cellular environment that interacts with Notch signaling under physiological or pathological conditions,7 and Notch signaling can promote the development of AV by activating NF-κB to produce a large number of inflammatory factors.8 Studies have shown that the NF-κB pathway is one of the important pathways promoting AV inflammation,9 but the regulatory mechanism of Notch/NF-κB signaling pathway on AV inflammation remains unclear.Pomegranate peel polyphenols(PPPs) are the main active substances in pomegranate peel,10 which are rich in content and have high bioavailability.The results of previous studies suggest that PPPs not only have anti-inflammatory effects11 and inhibit the proliferation of C. acnes,12 but also reduce inflammation by inhibiting the polarization of macrophages. However, the mechanism of PPPs in treating AV remains to be further elucidated.In this study, the AV model in vivo was established by injecting C. acnes into the auricle skin and intraperitoneally, and lipopolysaccharide(LPS) induced macrophages to establish the AV model in vitro. The potential mechanism of PPPs in treating acne was investigated by observing the levels of inflammatory indicators such as interleukn(IL)-6, IL-1α and Notch/NF-κB signaling pathway.
Materials and Methods
Materials
1-Stearoyl-rac-glycerol (Cat#123-94-4),Sodium dodecylbenzenesulfonate (Cat#25155-30-0), Laurocapram (Cat#59227-89-3), and Glycerol (Cat#56-81-5) came from Macklin Biochemical Co.,Ltd. (Shanghai, China). Cutibacterium acnes (Cat#6919) came from American Type Culture Collection Co.,Ltd. Pomegranate peel extract (70% polyphenol) (Cat#CO322107106) came from Kepler Biological Co.,Ltd. (Shanxi, China). Octadecanoic acid (Cat#30171118),Octadecanol (Cat#30137128),Liquid paraffin (Cat#30139860), and White vaseline (Cat#69047618) came from Hushi Laboratory Equipment Co.,Ltd. (Shanghai, China). Clindamycin hydrochloride ointment (Cat#H20040314) came from GoldenSun Group Co.,Ltd. (Fujian, China). Interleukn-6 (IL-6) ELISA KIT (Cat#JYM0646Ra), Interleukn-8 (IL-8) ELISA KIT (Cat#JYM0583Ra), Interleukn-12 (IL-12) ELISA KIT (Cat#JYM1006Ra), Interleukin-1α (IL-1α) ELISA KIT (Cat#JYM0649Ra),and Tumor Necrosis Factor-α (TNF-α) ELISA KIT (Cat#JYM0635Ra) came from Wuhan Jiyinmei Biotech Co.,Ltd. (Wuhan, China). RIPA lysis buffer (Cat#SW104-02), a loading buffer (Cat#SW-163-01), and BCA protein assay kit (Cat#SW101-02) were purchased from Seven Innovation Biotechnology Co.,Ltd. (Beijing, China.) a protease inhibitor (Cat#K1007) came from APExBIO Technology LLC. (Houston,America). ECL chemiluminescence reagent (Cat#BL523B) came from Biosharp Biotechnology Co.,Ltd. (Hefei, China). Antibodies toward the following proteins were used: Notch1 (Cat#4380T), Jagged1 (Cat#70109T), NF-κB p65 (Cat#8242T), Phospho-NF-κB p65 (Cat#3033T), TNF-α (Cat#11948T), NF-κB p65 (L8F6) Mouse mAb (Cat#6956T) and Phospho-NF-κB p65 (Ser536) (93H1) Rabbit mAb (Cat#3033T) came from Cell Signaling Technology (MA, USA). IL-6(Cat#DF6087) was from Affinity Biosciences (Jiangsu, China). IL-1 Alpha polyclonal antibody (Cat#16765-1-AP), and Beta Actin Rabbit RecAb (Cat#81115-1-RR) were obtained from Proteintech Biotechnology Co.,Ltd. (Wuhan, China). Anti-CD68(Cat#Ab283654) was from Abcam.(Shanghai, China). TRIzol™ Reagent (Cat#5003050) was obtained from Simgen Biological Reagent Development Co.,Ltd. (Hangzhou, China). NovoScript®Plus All-in-one 1st Strand cDNA Synthesis SuperMix (Cat#E047-01B) and NovoStart® SYBR High-Sensitivity qPCR SuperMix (Cat#E099-01A) were obtained from Novoprotein Technology Co.,Ltd. (Suzhou, China). DAPT (Cat#208255-80-5) and PEG300 (Cat#HY-Y0873) came from Med Chem Express (Shanghai, China). Cell Counting Kit-8 (CCK-8) (Cat#E-CK-A362) was from Elabscience Biotechnology Co., Ltd. (Wuhan, China). RAW264.7 Cell medium (Cat#CM-0190) was obtained from Procell (Wuhan, China). Dimethyl sulfoxide(DMSO) (Cat#67-68-5) and Lipopolysaccharide (LPS) (Cat#L2630-10MG) were obtained from Sigma-Aldrich(Shanghai, China). Fixable Viability Stain 450 (Cat#562247) was obtained from BD Pharming(Shanghai, China). PE anti-mouse CD80 (Cat#104707) was obtained from Biolegend (Beijing, China).
Preparation of Pomegranate Peel Polyphenols Cream
Prepare the cream base according to the method in Article Mechanisms of Pomegranate Peel Polyphenol Ointment in Preventing Acne Scar Formation by Mediating TGF-β/Smad Signaling Pathway.13 According to Huang Jihan’s Dose conversion among different animals and healthy volunteers in pharmacological study,14 the high, medium and low doses of PPPs in rats could be calculated as 0.28 g/(d·kg), 0.14 g/ (d·kg) and 0.07 g/ (d·kg).
Animal Experiments
The First Animal Experiments
A total of 48 SPF-grade healthy SD rats,Half male and half female, (No. 430727231101308048) (weighing about 200±20g) were obtained from Hunan Silaikejingda Experimental Animal Co.,Ltd (Changsha,China). These rats were accommodated at the Laboratory Animal Center affiliated with the Hunan University of Chinese Medicine (HNUCM) (License No. SYXK [Hunan] 2019–0004), ensuring continuous access to feed and water. C. acnes propionate solution (6×107cfu/mL) was injected intracutaneously into the two sides auricle of each rat once a day for 3 days. On the third day, each rat was given C. acnes propionate solution (6×107cfu/mL) 1mL intraperitoneally once a day for 7 consecutive days. The rats with C. acnes-induced acne vulgaris (CIAC) model was employed to assess the significance of PPPs on AC. Then, the CIAC models were randomly divided into groups: model group, cream base group, PPPs group (0.28,0.14,0.07 g/kg by external use), Clindamycin cream group (by external use), blank group and model group were treated with equal amount of normal saline externally. The drug was administered twice a day for 7 days.
The second Animal Experiments
The CIAC models were randomly divided into groups: model group, cream base group, PPPs group (0.28 g/kg for external use, the optimal concentration of PPPs in 2.3.1 group was selected), DAPT group (500 ug/100g by intraperitoneal injection). Blank group and model group were treated with equal amount of normal saline externally. DAPT was given once a day, and the other drug was administered twice a day for 7 days.
Cell Culture and Treatment
RAW264.7 were obtained from Procell Life Science&Technology Co., Ltd. The cells were cultured in RAW264.7 cell-specific medium and maintained in a CO2 incubator. The cells were passaged every day depending on the cell conditions. When grown to 80%–90% confluence, cells were randomly divided into a control group, LPS group (100 ng/mL, 500 ng/mL and 1000 ng/mL), to screen the optimal concentration of LPS (Figure 10C and D). Then, the cells were randomly divided into a blank group (RAW264.7 cell-specific medium for 24 h), model group (The optimal concentration of LPS for 24 hours), DMSO group (The cells were incubated with DMSO at the same dose as the model group 2 hours before LPS induction), PPPs-20ug/mL group (Two hours before LPS-induced cells were incubated with 20 μg/mL PPPs), DAPT group (Two hours before LPS-induced cells were incubated with 10 umol/L DAPT).
Morphological Observation
From the date of modeling, the changes of the auricle skin of rats were observed and recorded daily, including whether the skin surface was red, swollen, papules, pustules, cysts, enlarged pores and scales, and whether the skin was thickened or hardened.
Hematoxylin-Eosin (HE) Staining
The rat skin tissues were fixed with 4% paraformaldehyde after dehydration with gradient alcohol, xylene transparency, xylene, and paraffin immersion, embedded in paraffin, and cut into 5-μm slices. Next, the slices were stained with hematoxylin and eosin. Then, the slices were mounted for observation under a microscope (DMI8, Leica, Germany) and measured using Image-Pro Plus 6.0.
ELISA Assays
Take out the spare rat serum and follow the instructions of the kits.
Immunohistochemistry Assays
The rat skin tissues paraffin-embedded sections from each group were dewaxed, placed in Citrate buffer (pH 6.0) for antigen repair, and incubated in 1% periodic acid for 10 min at room temperature. The sections were immunoblotted with a primary antibody: Notch1,Jagged1,NF-κBp65, overnight at 4 °C, followed by incubation with the corresponding secondary antibody for one hour. The color was developed with DAB chromogenic, and the sections were counterstained with hematoxylin. The sections were observed under a microscope (DMI8, Leica, Germany), and ImagePro Plus 6.0 was employed to measure OD.
Immunofluorescence Detection
The rat skin tissues paraffin-embedded sections from each group were dewaxed, placed in Citrate buffer (pH 6.0) for antigen repair, and incubated in 1% periodic acid for 10 min at room temperature. The sections were immunoblotted with a primary antibody: p-NF-κBp65, NF-κBp65, CD68, overnight at 4 °C, followed by incubation with the corresponding secondary antibody for one hour. DAPI stain and anti-fading mountant forfluorescence quenching. The sections were observed under a microscope (DMI8, Leica, Germany).
Cells Proliferation
Cells in the logarithmic growth phase were collected and plated at a density of 5×105/mL cell suspension, then inoculated into a 96-well cell culture plate with a volume of 100 μL per well. After cell culture and treatment, 10 μL CCK-8 solution was added to each well. The optical density (OD) at 450 nm of each well was measured using a microplate reader (Varioskan LUX, Thermo Fisher Scientific, USA). Three independent experiments were carried out.
Flow Cytometry
Cells in the logarithmic growth phase were collected and plated at a density of 5× 105/mL cell suspension, then inoculated into a 6-well cell culture plate with a volume of 2 mL per well. The cells were treated with LPS of 100 ng/mL, 500 ng/mL and 1000 ng/mL for 24 hours, respectively. The cells were collected, incubated with antibodies, and counted by laser flow cytometry (Becton Dickinson and Company, Shanghai, China).
Western Blot Analysis
Protein concentration in the rat skin tissues and RAW264.7 was determined via BCA assay. Then the extracted protein was added to the loading buffer and boiled. Equal amounts of protein were resolved on an SDS-PAGE (8%, 10%, 12%) and transferred onto polyvinyl difluoride (PVDF) membranes. At room temperature, the membranes were blocked with 5% skimmed milk for one hour and then immunoblotted with a primary antibody: Notch-1 (1:1000), Jagged1 (1:1000), NF-κBp65 (1:1000), p-NF-κBp65 (1:1000), TNF-α (1:1000), IL-6 (1:1000), IL-1α (1:1000), β-actin (1:5000) overnight at 4 °C. Next, the membranes were incubated with the corresponding secondary antibody for one hour, and the specific protein bands were visualized with an ECL chemiluminescence reagent. Western blot bands were analyzed by Bio-rad molecular imager (Molecular Imager® ChemiDoc™ XRS+ Imaging System, Bio-rad, USA) and quantified using Quantity One software.
RNA Isolation and RT-qPCR Analysis
Total RNA from tissues and cells was extracted using TRIzol™ Reagent and was reverse transcribed into cDNA. The reverse transcription was performed at 50 °C for 15 min, followed by 85 °C for 5 min cDNA was used for RT-qPCR analysis using the UltraSYBR Mixture. The amplification procedures included pre-denaturation at 95 °C for 10 min; 40 cycles of denaturation at 95 °C for 15s; annealing at 60 °C for 30s; extension at 72 °C for 10 min. The primers for animal experiments were synthesized by Beijing Tsingke Biotech Co., Ltd. (Beijing, China): Notch1, forward GCTGCCTCTTTGATGGCTTCGA and reverse CACATTCGGCACTGTTACAGCC; Jagged1, forward TGCGTGGTCAATGGAGACTGGT and reverse TCGCACCGATACCAGTTGTCTC; NF-κBp65, forward TCCTGTTCGAGTCTCCATGCAG and reverse GGTCTCATAGGTCCTTTTGCGC; TNF-α, forward GGTGCCTATGTCTCAGCCTCTT and reverse GCCATAGAACTGATGAGAGGGAG; IL-1α, forward ACGGCTGAGTTTCAGTGAGACC and reverse CACTCTGGTAGGTGTAAGGTGC; IL-6, forward TACCACTTCACAAGTCGGAGGC and reverse CTGCAAGTGCATCATCGTTGTTC. The primers for cell experiments were synthesized by Beijing Tsingke Biotech Co., Ltd. (Beijing, China): Notch1, forward CTGAGCGTTTCAAAGTGTC and reverse TCGTTCTGATTGTCGTCC; NF-κBp65, forward CAGAGGCGTGTATTAGGGG and reverse GTGCTGTCAGGGAGGAAGG; TNF-α, forward GCGGTGCCTATGTCTCAG and reverse TCCTCCACTTGGTGGTTT; IL-1α, forward ACAACATACTGCTAACCGAC and reverse CTGGGGCATCACTTCTACC;IL-6,forward ACAACATACTGCTAACCGAC and reverse CTGGGGCATCACTTCTACC. The relative mRNA expression levels of Notch-1, Jagged1, NF-κBp65, IL-1α, IL-6 and TNF-α were determined via the 2−ΔΔCt analysis.
Statistical Analysis
All data were analyzed by SPSS 21.0 statistical software, and the measured data were processed by mean ± standard deviation. To compare the differences between multiple groups, one-way ANOVA and LSD were used when the data conformed to the normal distribution and homogeneity of variance. Conversely, the non-parametric test was used. Statistical significance was defined as *P < 0.05.
Results
Visible and Pathological Changes of Auricle Skin of Rats in Each Group
Compared with the blank group, the skin of the model group was red, swollen, thickened, with scales on it, and the follicular orifice was obviously enlarged, some of the follicular orifice was blocked into small cysts, and the touch was rough and hard, and the above symptoms were improved to varying degrees after various concentrations of PPPs cream and clindamycin cream (Figure 1A). The HE staining (Figure 1B) revealed a normal skin structure in blank group rats, the boundary between epidermis, dermis and subcutaneous tissue was clear, no obvious inflammatory cell infiltration was observed, the sebaceous gland volume was normal, and no capillary dilatation was observed. Meanwhile, in the model group, hyperkeratosis of the epidermis, hyperplasia of the sebaceous glands, enlargement of the follicle mouth, accumulation of a large amount of keratinized substances, dilatation of capillaries, and local infiltration of a large number of lymphocytes and histiocytic cells were observed. These conditions were alleviated to varying degrees using PPPs or clindamycin.
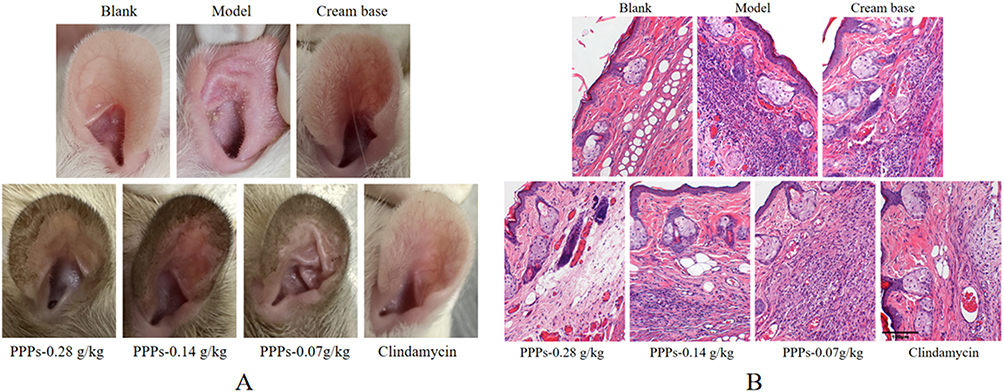 |
Figure 1 Obvious changes in the auricular skin of rats in different groups (A) and Pathological changes of auricle of rats in each group (B) (HE×200). |
PPPs (0.28g/kg, 0.14g/kg, 0.07g/Kg) Reduced IL-1α, IL-6, TNF-α, IL-8 and IL-12 Levels in the Serum of SD Rats with C. Acnes-Induced
Serum IL-1α, IL-6, TNF-α, IL-8 and IL-12 levels were upregulated in the model group compared to blank group; however, administration of PPPs cream (0.28g/kg, 0.14g/kg, 0.07g/kg) and Clindamycin cream decreased serum IL-1α, IL-6, TNF-α, IL-8, and IL-12 levels. (Figure 2A–E).
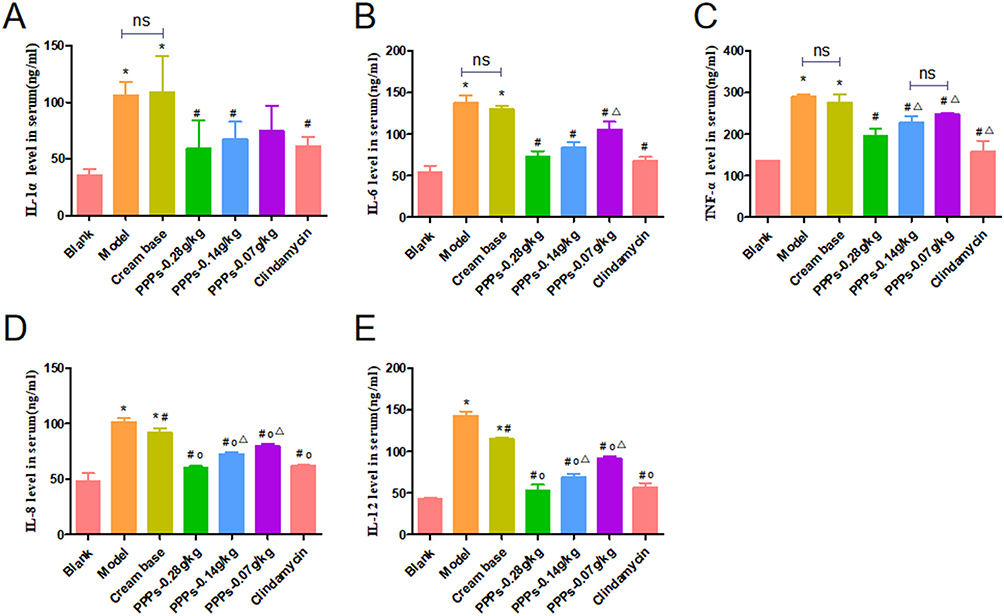 |
Figure 2 PPPs reduced IL-1α, IL-6, TNF-α, IL-8 and IL-12 content in the serum of SD rats with C. acnes-induced. (A–E)*P < 0.05 vs Blank group; #P < 0.05 vs Model group; ΔP < 0.05 vs Cream base group; op<0.05 vs. PPPs-0.28g/kg group; ns: P > 0.05. |
Figure 2 PPPs reduced IL-1α, IL-6,TNF-α, IL-8 and IL-12 content in the serum of SD rats with C. acnes-induced.*P < 0.05 vs Blank group; #P < 0.05 vs Model group; ΔP < 0.05 vs Cream base group;op<0.05vs. PPPs-0.28g/kg group;ns:P > 0.05.
PPPs Cream (0.28g/kg,0.14g/kg,0.07g/Kg) Inhibits IL-1α,IL-6,TNF-α, Notch1, Jagged1, NF-κBp65 Protein Expression in the SD Rats with C. Acnes-Induced
Western blot showed that the IL-1α, IL-6, TNF-α, Notch1, Jagged1, NF-κBp65 and p-NF-κBp65 protein expression increased in the model group and cream base group compared with the blank group. After treatment with PPPs cream (0.28g/kg, 0.14g/kg, 0.07g/kg) group or Clindamycin cream, the expression level of IL-1α,IL-6, TNF-α, Notch1, Jagged1, NF-κBp65 and p-NF-κBp65 decreased compared to the model group and cream base group. Among them, the 5.6% ppps cream group showed the most significant decline; however, there was no statistical difference in the rise and fall of Jagged among the groups (Figure 3A–G).
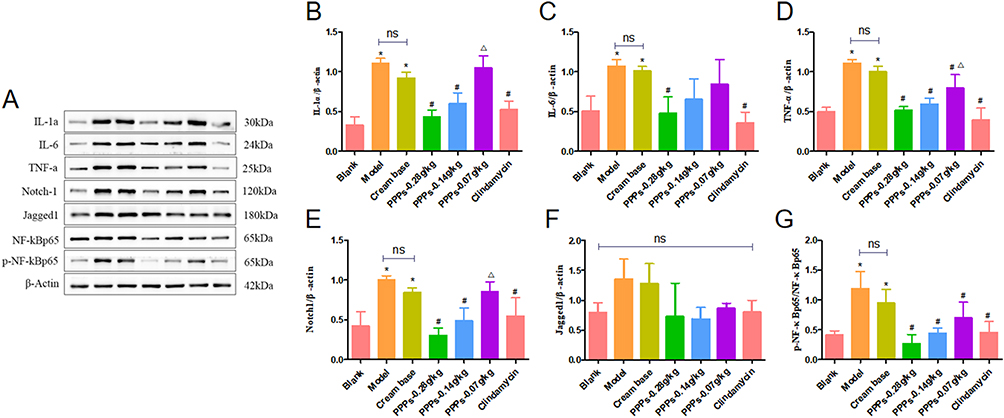 |
Figure 3 PPPs reduced IL-1α, IL-6, TNF-α, Notch1, Jagged1 and NF-κBp65 protein expression in the SD rats with C. acnes-induced. (A–G) *P < 0.05 vs Blank group; #P < 0.05 vs Model group; ΔP < 0.05 vs ppps-0.28g/kg group; ns: P > 0.05. |
PPPs (0.28g/kg, 0.14g/kg, 0.07g/Kg) Inhibits IL-1α, IL-6, TNF-α, Notch1, Jagged1 and NF-κBp65 mRNA Expression in the SD Rats with C. Acnes-Induced
In model group and cream base group, IL-1α, IL-6, TNF-α, Notch1 and NF-κBp65 mRNA expression in the Skin tissue increased compared to blank group, which decreased with PPPs cream (0.28g/kg, 0.14g/kg, 0.07g/kg) or Clindamycin cream for external group. Among them, the 5.6% ppps cream group showed the most significant decline; however, there was no statistical difference in the rise and fall of Jagged among the groups (Figure 4A–F).
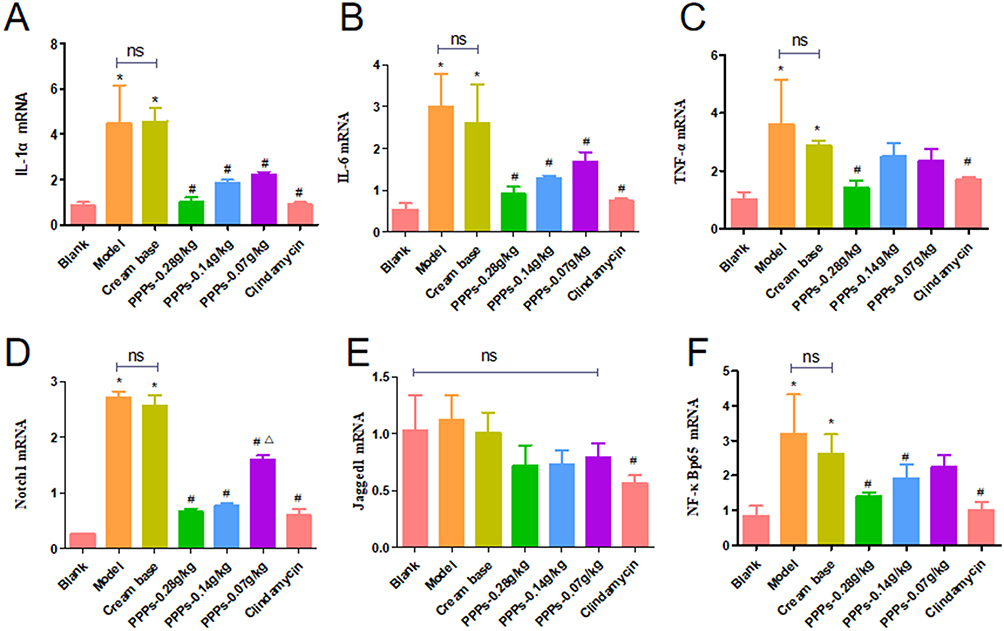 |
Figure 4 PPPs reduced IL-1α, IL-6, TNF-α, Notch1, Jagged1 and NF-κBp65 mRNA expression in the SD rats with C. acnes-induced. (A–F)*P < 0.05 vs Blank group; #P < 0.05 vs Model group; ΔP < 0.05 vs ppps-0.28g/kg group; ns:P > 0.05. |
Visible and Pathological Changes of Auricle Skin of Rats in Each Group
Compared with the blank group, the skin of the model group was red, swollen, thickened, with scales on it, and the follicular orifice was obviously enlarged, some of the follicular orifice was blocked into small cysts, and the touch was rough and hard, and the above symptoms were improved after ppps-0.28g/kg and DAPT (Figure 5A). The HE staining (Figure 5B) revealed a normal skin structure in blank group rats, the boundary between epidermis, dermis and subcutaneous tissue was clear, no obvious inflammatory cell infiltration was observed, the sebaceous gland volume was normal, and no capillary dilatation was observed. Meanwhile, in the model group, hyperkeratosis of the epidermis, hyperplasia of the sebaceous glands, enlargement of the follicle mouth, accumulation of a large amount of keratinized substances, dilatation of capillaries, and local infiltration of a large number of lymphocytes and histiocytic cells were observed. These symptoms can be alleviated after using PPPs and DAPT.
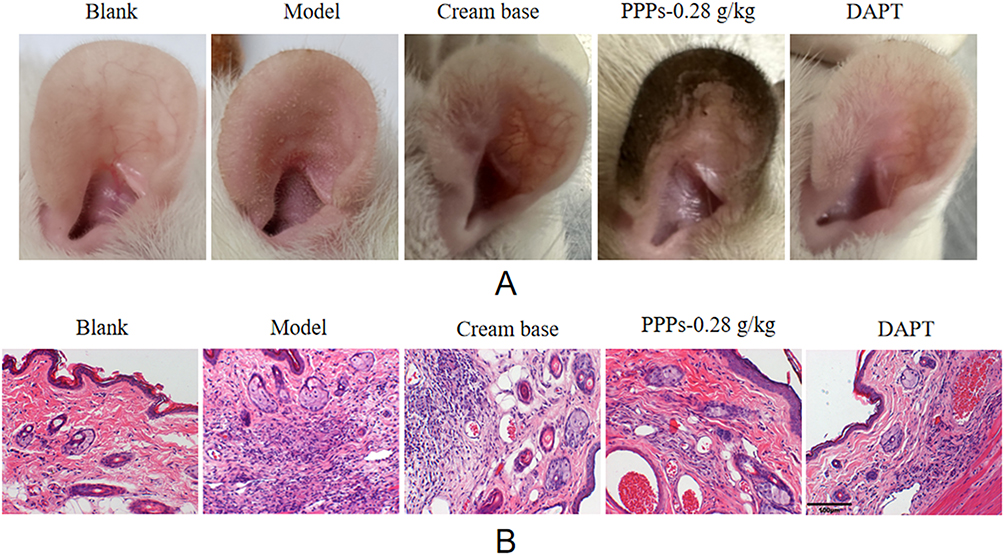 |
Figure 5 Visible changes of auricle of rats in each group (A) and Pathological changes of auricle of rats in each group (B) (HE×200). |
PPPs Cream (0.28g/Kg) Reduced IL-1α, IL-6, TNF-α, IL-8 and IL-12 Levels in the Serum of SD Rats with C. Acnes-Induced
Serum IL-1α, IL-6, TNF-α, IL-8 and IL-12 levels were upregulated in the model group compared to blank group; however, administration of PPPs cream (0.28g/kg) and DAPT decreased serum IL-1α, IL-6, TNF-α, IL-8, and IL-12 levels (Figure 6A–E).
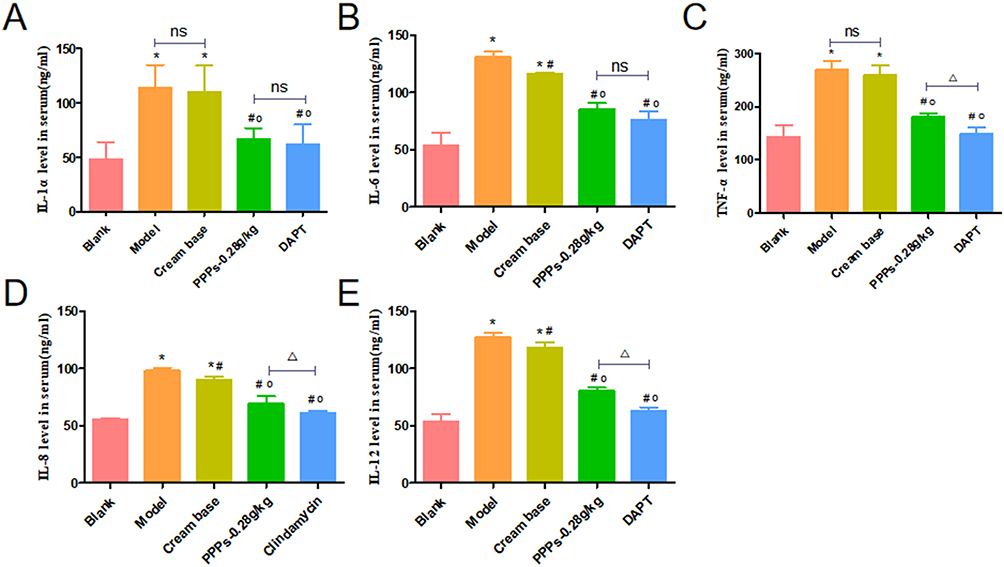 |
Figure 6 PPPs reduced IL-1α, IL-6, TNF-α, IL-8 and IL-12 content in the serum of SD rats with C. acnes-induced. (A–E)*P < 0.05 vs Blank group; #P < 0.05 vs Model group; op<0.05vs. Cream base group; ΔP < 0.05; ns:P > 0.05. |
PPPs (0.28g/Kg) Inhibits Notch1, NF-κBp65, IL-6, TNF-α and IL-1α mRNA Expression in the SD Rats with C. Acnes-Induced
In model group and cream base group, Notch1, NF-κBp65, IL-6, TNF-α and IL-1α mRNA expression in the Skin tissue increased compared to blank group, which decreased with PPPs cream (0.28g/kg) or DAPT (Figure 7A–F).
 |
Figure 7 PPPs reduced Notch1,NF-κBp65,IL-6,TNF-α and IL-1α mRNA expression in the SD rats with C. acnes-induced. (A–E)*P < 0.05 vs Blank group; #P < 0.05 vs Model group; op<0.05vs. Cream base group;ΔP < 0.05;ns:P > 0.05. |
PPPs Inhibited Notch1, NF-κBp65, IL-1α, IL-6 and TNF-α Protein Expression in the SD Rats with C. Acnes-Induced
Immunohistochemistry analysis showed that in the SD rats with C. acnes-induced, the Notch1 and NF-κBp65 positive protein expression in the skin increased compared to blank group. The expression level decreased following treatment with PPPs cream (0.28g/kg) or DAPT (Figure 8A–D). Jagged1 was not expressed in all groups (Figure 8E). Western blot showed that the Notch1, NF-κBp65, p-NF-κBp65, IL-1α, IL-6, TNF-α protein expression and p-NF-κB/ NF-κB increased in the model group compared with the blank group. Following treatment using PPPs cream (0.28g/kg) or DAPT, the Notch1, NF-κBp65, p-NF-κBp65, IL-1α,IL-6, TNF-α protein expression and p-NF-κB/ NF-κB decreased compared with the model group (Figure 8F and G). There was no statistical significance in Jagged expression.
 |
Figure 8 PPPs inhibited Notch1 and NF-κBp65 protein expression in the skin of SD rats with C. acnes-induced. (A–D) Jagged1 was not expressed in all groups. (E) Representative immunohistochemistry images and semiquantitative analysis of skin of rats stained with Notch1 and NF-κBp65 antibody. Western blotting and quantitative analysis of Notch1, NF-κBp65, p-NF-κBp65, IL-1α, IL-6 and TNF-α in SD rats. (F and G). Among them, the expression of Jadded was not statistically significant.*P < 0.05 vs Blank group; #P < 0.05 vs Model group; ΔP < 0.05;ns:P > 0.05. |
Immunofluorescence Results of NF-κBp65, p-NF-κBp65, CD68
As shown in Figure 9A–C, the model group and cream base group exhibited strong green fluorescence in NF-κBp65 and strong red fluorescence in p-NF-κBp65, while minimal fluorescence was observed in PPPs and DAPT-treated NF-κBp65 and p-NF-κBp65. As shown in Figure 9B–D, the model group and cream base group exhibited strong red fluorescence in CD68, while minimal fluorescence was observed in PPPs and DAPT-treated CD68.
 |
Figure 9 Immunofluorescence staining and FI values of NF-κBp65 (green), p-NF-κBp65 (red), and nuclei (blue) on the skin of SD rats with C. acnes-induced. (A–C) (X40); Immunofluorescence staining and FI values of CD68 (red), and nuclei (blue) on the skin of SD rats with C. acnes-induced. (B and D)(X40); *P < 0.05 vs Blank group. #P < 0.05 vs Model group; ΔP < 0.05; ns: P > 0.05. |
Macrophages Induced by LPS
After 24 hours of LPS induction, RAW264.7 cells became significantly larger in volume and grew pseudopodia. Pseudopodia pulling made the cells appear to be spindle shaped, and the vacuoles in the cytoplasm gradually increased (Figure 10A). LPS (100 ng/mL, 500 ng/mL,1000 ng/mL) induced macrophage proliferation, but there was no statistical difference in survival rate between the three concentrations (Figure 10B). When macrophages were polarized towards the M1 phenotype using LPS, robust upregulation in the green fluorescence intensity of CD80 was observed. Flow cytometry showed that when LPS was used to polarize macrophages into M1 phenotype, the expression of CD80 was significantly increased, and the proportion of M1 macrophages induced by LPS (500ng/mL and 1000ng/mL) was superior to that induced by LPS (100ng/mL). There was no difference between LPS (500ng/mL)and LPS (1000ng/mL) (Fig.C&D).
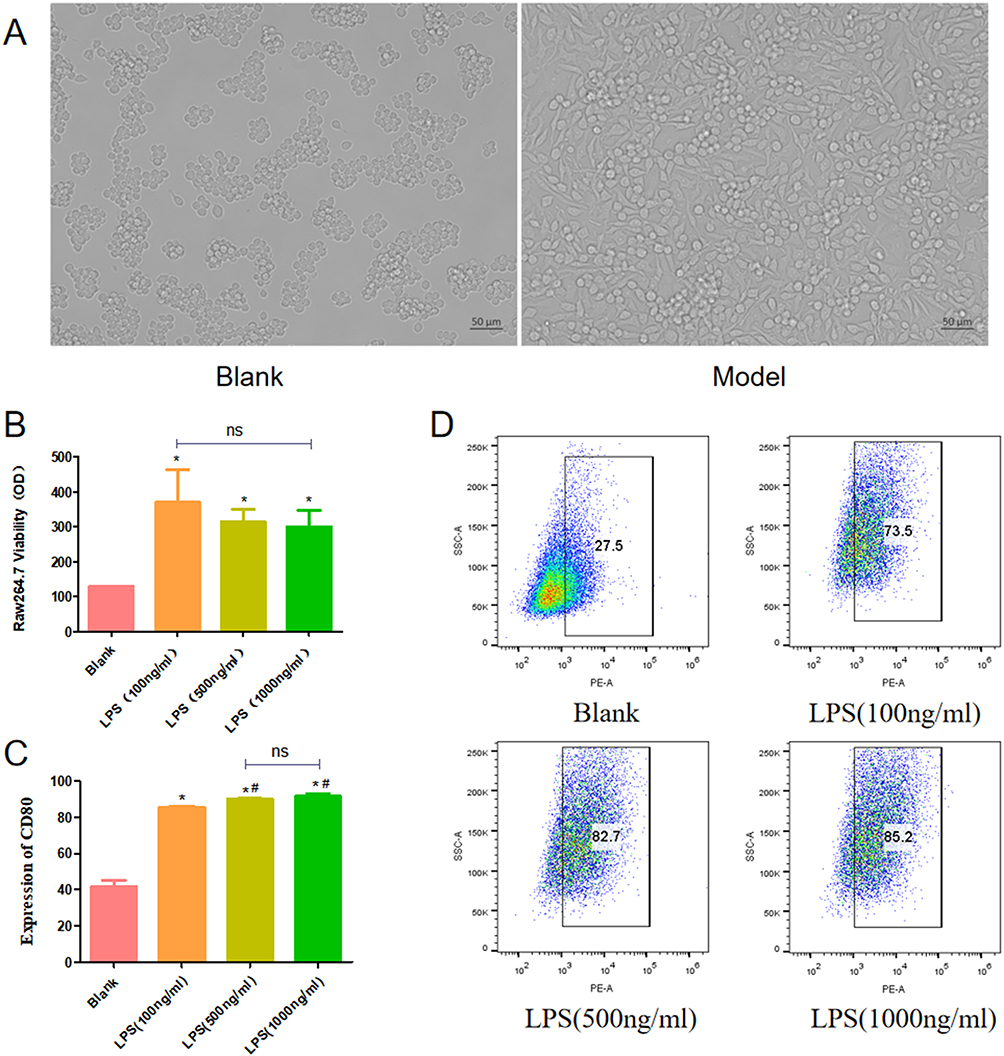 |
Figure 10 Morphology of normal macrophages and LPS-induced macrophages (A). Effects of LPS on proliferation of RAW264.7 cells (B). Percentages of CD80 positive macrophages were analyzed by flow cytometry (C and D).*P < 0.05 vs Blank group; #P < 0.05 vs LPS (100ng/mL); ns: P > 0.05. |
PPPs Inhibits Notch1, NF-κBp65, IL-1α, IL-6 and TNF-α mRNA Expression in the RAW264.7 Induced by LPS
In RAW264.7 induced by LPS, Notch1, NF-κBp65,IL-1α, IL-6 and TNF-α mRNA expression increased in the model group compared with the blank group. After treatment using PPPs or DAPT, the Notch1, NF-κBp65, IL-1α, IL-6 and TNF-α mRNA expression decreased compared with the model group (Figure 11A–E).
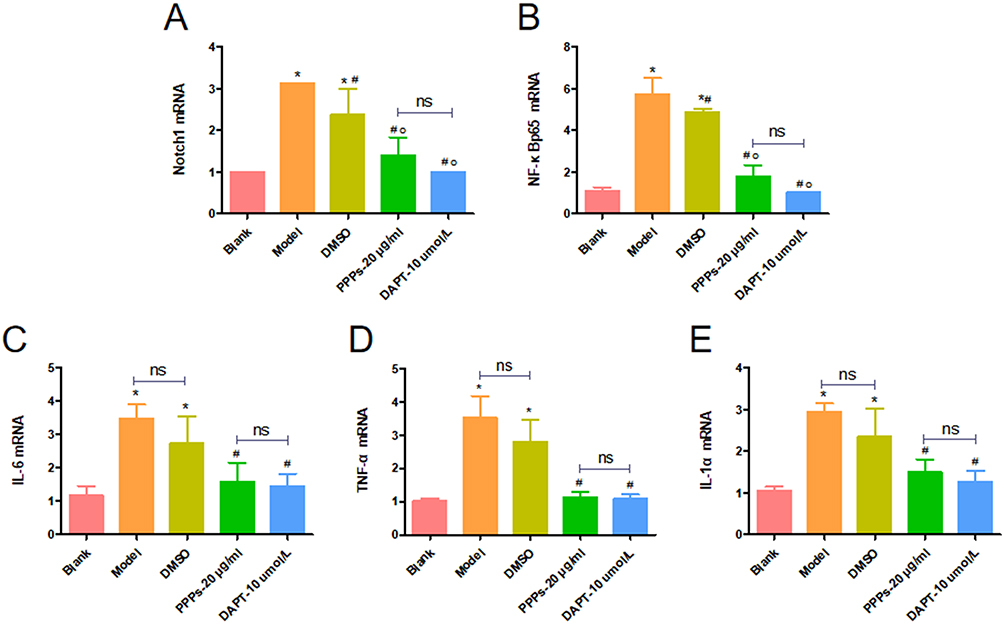 |
Figure 11 PPPs inhibited Notch1, NF-κBp65, IL-1α, IL-6 and TNF-α mRNA expression in the RAW264.7 induced by LPS. (A–E) *P < 0.05 vs Blank group; #P < 0.05 vs Model group; op<0.05 vs. DMSO group; ns: P > 0.05. |
PPPs Inhibits Notch1, NF-κBp65, IL-1α, IL-6 and TNF-α Protein Expression in the RAW264.7 Induced by LPS
In RAW264.7 induced by LPS, Notch1, NF-κBp65, IL-1α, IL-6 and TNF-α Protein expression increased in the model group compared with the blank group. After treatment using PPPs or DAPT, the Notch1, NF-κBp65, IL-1α, IL-6 and TNF-α Protein expression decreased compared with the model group (Figure 12A–F).
 |
Figure 12 PPPs inhibited Notch1, NF-κBp65, IL-1α, IL-6 and TNF-α protein expression in the RAW264.7 induced by LPS. (A–F) *P < 0.05 vs Blank group; #P < 0.05 vs Model group; ns: P > 0.05. |
Discussion
Although the pathogenesis of acne is complex, the theory of inflammation is still the mainstream today.15,16 Many scholars believe that C. acnes is the main force inducing AV inflammation,17 but some scholars believe that AV inflammation is not a subsequent event, but in the early stage of AV inflammation.4 For AV inflammation, antibiotics are generally used for treatment. Although antibiotics have made great contributions to the development of medicine, their drug resistance has a serious negative impact on medicine, human health and even society. It is of great practical significance and application value to find suitable antibiotic substitutes. More and more studies at home and abroad have found that plant-derived natural products have good effects in antibacterial and anti-inflammatory aspects, and PPPs are the main active substances in pomegranate peel, which is a good natural anti-inflammatory drug and can play a role in a variety of inflammatory diseases.
Studies have shown that C. acnes plays an important role in the activation of AC inflammation, and can play a pro-inflammatory role by stimulating macrophages, sebaceous cells and keratinocytes to secrete IL-1, IL-6, IL-8, IL-12, TNF-α and other factors. Among them, the IL-1 family plays a key role in the initiation and development of inflammation.18 In addition to causing excessive keratosis of follicular orifice, IL-1 can recruit neutrophils and induce the secretion of IL-6 and IL-8 to mediate inflammation. IL-1α is the main form of its presence in AC lesions. Relevant studies have found that high expression of IL-1α is detected not only during inflammatory skin lesions such as papules and pustules that occur after substantial colonization of C. acnes, but can already be identified at the microcomedone stage.19 It can lead to abnormal follicle keratosis, sebum excretion difficulty, promote the occurrence and development of inflammatory response. IL-6 and TNF-α are key to initiating the innate immune response.20 IL-6 is a critical mediator in inflammatory responses, not only directly participating in inflammation but also chemoattracting and activating neutrophil-dominated inflammatory cell infiltration during the early stages of inflammation, thereby exacerbating the progression of inflammatory reactions. This indicates that IL-6 serves as both an agonist and a catalyst for inflammation. TNF-α induces inflammatory responses not only by stimulating endothelial cells to produce adhesion molecules, which promote the aggregation of inflammatory cells to sites of cutaneous injury, but also by activating inflammatory cells to regulate the inflammatory cascade, ultimately leading to the onset of inflammatory reactions. IL-8 and IL-12 are also implicated in the inflammatory and immune responses of acne. Their mechanism likely involves C. acnes stimulating immune cells to activate intracellular signaling pathways, thereby triggering the release of pro-inflammatory cytokines, including IL-8 and IL-12.21 Our findings demonstrate that PPPs ameliorated symptom manifestations in CIAC, such as erythema, skin roughness, and follicular orifice dilation. Furthermore it reduced histopathological cellular infiltration and decreased serum levels of inflammatory factors, including IL-1α, IL-6, TNF-α, IL-8, and IL-12. Additionally, they downregulated protein and mRNA expression levels of IL-1α, IL-6, and TNF-α in skin tissues. Notably, these results confirmed the superior efficacy of the high-concentration polyphenol group compared to medium- and low-dose groups.
The Notch signaling pathway has emerged as a focal point in recent research on immune-inflammatory diseases and is highly expressed in various inflammatory skin disorders. NF-κB, a key transcriptional factor in acne-associated inflammatory responses, promotes inflammatory cascades in acne through the release of multiple pro-inflammatory cytokines upon activation. Notably, NF-κB also serves as a critical downstream target of Notch signaling, which regulates NF-κB expression via protein-protein interactions and gene transcriptional modulation. Studies report that active Notch1 strongly activates NF-κB by converting repressor complexes into activator complexes at the transcriptional level.22 Additionally, the Notch1 intracellular domain competes with IκBα for NF-κB binding, thereby enhancing NF-κB transcriptional activity.23 Research24,25 demonstrates that NF-κB activity diminishes upon Notch-1 silencing, indicating Notch-1’s regulatory role in NF-κB activation. Furthermore, synergistic interactions between Notch-1 and Notch-3 receptors maximize NF-κB activation.26 Activated NF-κB drives increased expression of inflammatory cytokines, including IL-6, TNF-α, IL-8, IL-12, and IL-1α, amplifying inflammatory cascades. While extensive literature implicates Notch signaling in NF-κB-mediated regulation of inflammatory diseases, its role in acne pathogenesis remains unexplored. Therefore, this study investigated the expression profiles of Notch signaling and NF-κB in AC. Results revealed upregulated expression of Notch1 and NF-κBp65 in acne-like lesions. Notably, PPPs significantly reduced Notch1 and NF-κBp65 expression in these lesions, suggesting Notch signaling may contribute to acne-associated inflammatory responses.
To further investigate whether Notch signaling exerts pro-inflammatory effects in acne by activating the NF-κB pathway, CIAC model rats were administered intraperitoneal injections of the Notch pathway inhibitor DAPT and topical PPPs. Results demonstrated significant reductions in both protein and mRNA expression of Notch-1 and NF-κBp65 in skin lesions of treated rats compared to the model group, though no statistically significant changes in Jagged1 expression were observed. Furthermore, nuclear levels of p-NF-κBp65 were markedly elevated in the model group, whereas interventions with DAPT and PPPs significantly reduced nuclear p-NF-κBp65 expression. These findings suggest that Notch signaling promotes NF-κB activation by facilitating its nuclear translocation, a mechanism potentially independent of its ligand Jagged1. In acne model rats, elevated expression of IL-6, TNF-α, IL-8, IL-12, and IL-1α was detected in both skin lesions and peripheral blood compared to the blank group. Following treatment with topical PPPs cream and intraperitoneal DAPT, these inflammatory factors were significantly downregulated in both compartments. These results further support the hypothesis that Notch signaling regulates AC-associated inflammatory cytokine expression via the NF-κB pathway, while PPPs exhibit anti-inflammatory effects by suppressing Notch signaling activity.
Recent studies27 integrating single-cell transcriptomic data have revealed a higher proportion of macrophages in acne patient skin compared to healthy controls, identifying macrophages as a potential therapeutic target for acne. Notably, the Notch signaling pathway, a critical regulator of macrophage biology, has been implicated in modulating the activation of M1-polarized macrophages.6 Our findings corroborate this hypothesis, demonstrating significantly elevated expression of CD68—a macrophage-specific marker—in AC-like lesions, which was reduced following DAPT intervention. This indicates macrophage infiltration in acne model rats and suggests Notch-dependent regulation of macrophage involvement in acne pathogenesis. In an “in vitro” acne model using LPS-stimulated RAW264.7 macrophages, LPS was observed to promote cellular proliferation and exacerbate inflammatory responses, effects that were reversed by PPPs. Further investigation into the molecular mechanisms revealed that under LPS induction, the Notch pathway specifically triggers NF-κB to activate macrophages, culminating in excessive production of inflammatory cytokines such as IL-1α, IL-6, and TNF-α, thereby aggravating acne-associated inflammation. Importantly, PPPs effectively suppressed Notch/NF-κB pathway activation.
Conclusion
In our study, PPPs ameliorated inflammatory responses in AC-like rat models by suppressing LPS-induced macrophage proliferation and M1 polarization, thereby attenuating inflammation. These mechanisms may be associated with the inhibition of the Notch/NF-κB signaling pathway. However, the findings of this study require further validation. Future investigations will include siRNA experiments or complementary approaches to strengthen the robustness of our conclusions. In summary, our research provides novel insights into potential therapeutic strategies for acne.
Abbreviations
PPPs, Pomegranate peel polyphenols; AV, Acne vulgaris; NF-κB, Nuclear factor kappa-B; C. acnes, Cutibacterium acnes; LPS, Lipopolysaccharide; IL, Interleukn; CIAC, C. acnes-induced acne vulgaris.
Data Sharing Statement
The datasets used and/or analyzed in the current study are available from the corresponding author on reasonable request.
Ethical Approval
This study was performed under the approval of the Ethics Committee of Hunan University of Chinese Medicine (Protocol Number:LL2023032201). The name of the guideline followed for the welfare of laboratory animals is the “3R” guideline. All surgeries were performed under sodium pentobarbital anesthesia. Animals were housed in a temperature-controlled facility with a 12-h light/dark cycle and ad libitum access to food and water. Humane endpoints were strictly observed, and animals showing severe distress were euthanized immediately.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Natural Science Foundation Project in Hunan Province [grant number 2023JJ60485]; the Hunan Province High-Level Health Talents 225 Project (Xiangweihan [2019]196); Hunan Province “14th Five-Year” Chinese medicine leading talent project ([2022]4).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Leeming J-P, Holland K-T, Cuncliffe W-J. The microbial colonization of inflamed acne vulgaris lesions. Br J Dermatol. 1988;118(2):203–208. doi:10.1111/j.1365-2133.1988.tb01775.x
2. Leyden -J-J, McGinley K-J, Vowels B. Propionibacterium acnes colonization in acne and nonacne. Dermatology. 1998;196(1):55–58. doi:10.1159/000017868
3. Chinese-Society-Of-Dermatology Working Group For Acne Diseases. Guideline for diagnosis and treatment of acne (the 2019 revised edition). J Clin Dermatol. 2019;48(09):583–588.
4. Jeremy A-H, Holland D-B, Roberts S-G, et al. Inflammatory events are involved in acne lesion initiation. J Invest Dermatol. 2003;121(1):20–27. doi:10.1046/j.1523-1747.2003.12321.x
5. Fengyun Zhang, Zhao Z, Zhang Z. Effects of miR-34a on polarization and inflammatory factor content of mouse macrophage cell line RAW264.7 under high glucose conditions. Basic Clin Med. 2023;43(12):1808–1813.
6. Keewan E, Naser S-A. Notch-1 signaling modulates macrophage polarization and immune defense against mycobacterium avium paratuberculosis infection in inflammatory diseases. Microorganisms. 2020;8(7):1006. doi:10.3390/microorganisms8071006
7. Ferrandino F, Grazioli P, Bellavia D, et al. Notch and NF-kappaB: coach and players of regulatory T-cell response in cancer. Front Immunol. 2018;92165.
8. de Avila MJR, Lopez-Lopez S, Garcia-Blazquez A, et al. RND3 potentiates proinflammatory activation through NOTCH signaling in activated macrophages. J Immunol Res. 2024;20242264799.
9. Wen-li H, Jie W, Huang X, et al. Effect of acne granules as adjuvant therapy on peripheral blood NF-κB signaling pathways of patients with infectious acne. Chin J Nosocomiol. 2023;33(09):1380–1384.
10. Singh B, Singh J-P, Kaur A, et al. Phenolic compounds as beneficial phytochemicals in pomegranate (Punica granatum L.) peel: a review. Food Chem. 2018;26175–26186.
11. Xu J, Cao K, Liu X, et al. Punicalagin regulates signaling pathways in inflammation-associated chronic diseases. Antioxidants. 2021;11(1).
12. Zhang X-L, Ming-Fang Z, Liu Y, et al. Antibacterial effect of pomegranate peel polyphenols against propionibacterium acnes in vitro. J Hunan Univ Chin Med. 2015;35(09):23–25.
13. Yixuan YANG, Mingfang ZHU, Wang F, et al. Mechanisms of pomegranate peel polyphenol ointment in preventing acne scar formation by mediating TGF-β/smad signaling pathway. J Hunan Univ Chin Med. 2019;39(05):596–599.
14. Huang J, Xiaojun H, Zhiyang C, et al. Dose conversion among different animals and healthy volunteers in pharmacological study. Chi J Clin Pharmacol Therap. 2004;09:1069–1072.
15. Zhou J. Study on the Relationship Between Acne and Abnormal Glucose and Lipid Metabolism. Hunan University of Chinese Medicine; 2021.
16. Kim H-J, Lee B-J, Kwon A-R. The grease trap: uncovering the mechanism of the hydrophobic lid in cutibacterium acnes lipase. J Lipid Res. 2020;61(5):722–733. doi:10.1194/jlr.RA119000279
17. Parndaeng K, Pitakbut T, Wattanapiromsakul C, et al. Chemical constituents from streblus taxoides wood with their antibacterial and antityrosinase activities plus in silico study. Antibiotics. 2023;12(2).
18. Ingham E, Eady E-A, Goodwin C-E, et al. Pro-inflammatory levels of interleukin-1 alpha-like bioactivity are present in the majority of open comedones in acne vulgaris. J Invest Dermatol. 1992;98(6):895–901. doi:10.1111/1523-1747.ep12460324
19. Norris J-F, Cunliffe W-J. A histological and immunocytochemical study of early acne lesions. Br J Dermatol. 1988;118(5):651–659. doi:10.1111/j.1365-2133.1988.tb02566.x
20. Cao X. Medical Immunology. People’s Health Publishing House; 2021.
21. Firlej E, Kowalska W, Szymaszek K, et al. The role of skin immune system in acne. J Clin Med. 2022;11(6):1579. doi:10.3390/jcm11061579
22. Oswald F, Liptay S, Adler G, et al. NF-kappaB2 is a putative target gene of activated Notch-1 via RBP-Jkappa. Mol Cell Biol. 1998;18(4):2077–2088. doi:10.1128/MCB.18.4.2077
23. Shin H-M, Minter L-M, Cho O-H, et al. Notch1 augments NF-kappaB activity by facilitating its nuclear retention. EMBO J. 2006;25(1):129–138. doi:10.1038/sj.emboj.7600902
24. Palaga T, Buranaruk C, Rengpipat S, et al. Notch signaling is activated by TLR stimulation and regulates macrophage functions. Eur J Immunol. 2008;38(1):174–183. doi:10.1002/eji.200636999
25. Monsalve E, Ruiz-Garcia A, Baladron V, et al. Notch1 upregulates LPS-induced macrophage activation by increasing NF-kappaB activity. Eur J Immunol. 2009;39(9):2556–2570. doi:10.1002/eji.200838722
26. Lopez-Lopez S, Monsalve E-M, De-Avila-MJ R, et al. NOTCH3 signaling is essential for NF-kappaB activation in TLR-activated macrophages. Sci Rep. 2020;10(1):14839. doi:10.1038/s41598-020-71810-4
27. Zhao C-X, Wang S-L, Li H-X, et al. Integration of single-cell transcriptomics data reveal differences in cell composition and communication in acne. Clin Cosmet Investig Dermatol. 2023;163413–163426.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Real-World Case Studies Showing the Effective Use of Azelaic Acid in the Treatment, and During the Maintenance Phase, of Adult Female Acne Patients
Layton AM, Dias da Rocha MA
Clinical, Cosmetic and Investigational Dermatology 2023, 16:515-527
Published Date: 24 February 2023
Plasma Treatment – Results of Skin Microbiome Analysis
Watanabe C
Clinical, Cosmetic and Investigational Dermatology 2025, 18:1269-1279
Published Date: 26 May 2025
Safety and Clinical Efficacy of KūlarisTM, an Herbal Supplement for Mild to Severe Acne Vulgaris – A 12-Week Randomized, Double Blind, Placebo-Controlled Study
Sudeep HV, Chandradhara D, Shyamprasad K
Clinical, Cosmetic and Investigational Dermatology 2025, 18:2475-2488
Published Date: 26 September 2025