Back to Journals » Journal of Experimental Pharmacology » Volume 14
The Oil Formulation Derived from Moringa Oleifera Seeds Ameliorates Behavioral Abnormalities in Water-immersion Restraint Stress Mouse Model
Authors Purwoningsih E, Arozal W, Lee HJ, Barinda AJ ![]() , Sani Y, Munim A
, Sani Y, Munim A
Received 19 August 2022
Accepted for publication 8 December 2022
Published 22 December 2022 Volume 2022:14 Pages 395—407
DOI https://doi.org/10.2147/JEP.S386745
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Jianbo Sun
Emni Purwoningsih,1,2 Wawaimuli Arozal,3 Hee Jae Lee,4 Agian Jeffilano Barinda,3,5 Yulvian Sani,6 Abdul Munim7
1Doctoral Program in Biomedical Science, Faculty of Medicine, Universitas Indonesia, Jakarta, Indonesia; 2Faculty of Medicine, Universitas Muhammadiyah Sumatera Utara, Medan, Indonesia; 3Department of Pharmacology and Therapeutics, Faculty of Medicine, Universitas Indonesia, Jakarta, Indonesia; 4Department of Pharmacology, School of Medicine, Kangwon National University, Chuncheon, Republic of Korea; 5Metabolic, Cardiovascular, and Aging Cluster, Indonesia Medical Education and Research Institute (IMERI), Faculty of Medicine, Universitas Indonesia, Jakarta, Indonesia; 6Research Center for Biomedical Research, National Research and Innovation Agency, Jakarta, Indonesia; 7Department of Pharmacognosy-Phytochemistry, Faculty of Pharmacy, Universitas Indonesia, Depok, West Java, Indonesia
Correspondence: Wawaimuli Arozal, Department of Pharmacology and Therapeutics, Faculty of Medicine, Universitas Indonesia, Jl. Salemba Raya No. 6, Jakarta, Indonesia, 10430, Tel +62-21-31930481, Fax +62-21-3920947, Email [email protected]; [email protected] Hee Jae Lee, Department of Pharmacology, School of Medicine, Kangwon National University, Chuncheon Campus 1, Gangwondaehakgil, Chuncheon-si, Gangwon-do, 24341, Republic of Korea, Tel/Fax +82-33-250-8850, Email [email protected]
Purpose: Repeated stress events are well known to be associated with the onset of behavioral abnormalities including depression, anxiety and memory impairment. In spite of the traditional uses of Moringa oleifera (MO), no experimental evidence for its use against chronic stress exists. Here, we investigated whether seed oil from MO (MOO) could improve behavior abnormalities of chronic stress mice induced by water-immersion restraint stress (WIRS) and the underlying mechanism.
Methods: BALB/C male mice at 12 weeks of age were exposed to chronic WIRS for two weeks and divided in to four groups: normal group, WIRS group, WIRS+MOO1 group (treated with MOO at the dose of 1 mL/kg BW), and WIRS+MOO2 group (treated with MOO 2 mL/kg BW). The MOO treatment was given orally for 23 days. On day 24, we checked the behavior parameters, the plasma level of cortisol, acetylcholinesterase (AChE) activity in hippocampus, mRNA expression level of brain-derived neurotrophic factor (BDNF) and oxidative stress parameters in brain tissues. In addition, we also checked the histopathological features of the gastric mucosa wall.
Results: Administration of MOO ameliorated anxiety-like, depression-like and memory impairment phenotypes in the WIRS mouse model although the plasma cortisol concentrations were comparable among the groups. Of note, MOO both in two doses could suppress the AChE activity in hippocampus tissue and ameliorated the MDA level in prefrontal cortex tissue in mice exposed to WIRS. Although only WIRS+MOO2 group could increase the mRNA expression of BDNF, the histopathological gastric mucosa wall features were improved in all MOO groups.
Conclusion: Taken together, these finding suggested that MOO may have a neuroprotective effect in the mouse model of WIRS as evidenced by improving the abnormal behaviors through enhancing mRNA expression level of BDNF, inhibited AChE activity, and prevented the increase of MDA level in the brain.
Keywords: Moringa oleifera, behavioral abnormality, WIRS, AChE activity, oxidative stress, BDNF
Introduction
Stress is an essential part of organism life, and it is known that many organs and systems, especially the brain, are affected by stress, causing the natural integrity and homeostasis of the organism to be threatened. Stress causes structural changes in the hippocampus, prefrontal cortex, amygdala, anterior cingulate cortex, and basal ganglia.1 Excessive external stress is a widely accepted theory in the development of mood disorders such as depression and anxiety.2 Stress negatively affects many parts of the brain which are involved in the regulation of emotion including the cortex and hippocampus.3,4 In addition, for decades the idea that maladaptive stress impairs cognitive function has been a cornerstone to be explored in basic and clinical research. In human studies, acute stressors can cause short-term, but reversible deficits in tasks of memory, while chronic stress could lead to irreversible loss of hippocampal neurons and cognitive impairment.5 Results obtained from animal studies also confirmed a significant effect of stress on global cognitive performance.6 In view of the unfavorable chronic stress effects on cognitive function and mood disorder, it is important to explore the possible therapeutic agent, including herbal supplementation to ameliorate these phenomena.
The hypothalamic-pituitary-adrenal (HPA) axis is implicated in the pathogenesis of a wide range of neuropsychiatric disorders.7–9 In depression, hyperactivity of the HPA axis occurs from inadequate negative feedback mediated by the glucocorticoid system.7 This is due to cortisol hypersecretion in chronic stress,10 and cortisol levels have been shown to decrease verbal and working memory, lowering cognitive performance in depression.7 Brain-derived neurotrophic factor (BDNF) is a crucial modulator of neuroplasticity, which includes brain remodeling and neurogenesis.11 In addition, previous research has shown strong correlation between chronic stress-induced depression and changes in BDNF.12,13 Previous studies have found that exposure to various types of chronic stress could decrease BDNF expression level in the hippocampus and prefrontal cortex and BDNF activity in the limbic brain areas.13,14
It has been established that free radicals such as reactive oxygen species (ROS) and reactive nitrogen species (RNS) play an important role in the pathophysiology of many diseases including neuropsychiatric diseases with the symptom of depression and cognitive impairment.15,16 The increase in ROS and RNS production or the decrease in antioxidant mechanisms generates conditions called oxidative and nitrosative stress, respectively, defined as the imbalance between pro- and antioxidants in favor of the oxidants, which is detrimental to cells.16
Moreover, the reduced ability of antioxidant defense mechanisms in the central nervous system (CNS) for eliminating excess ROS and RNS leads to tissue damage, inflammation, and neurodegeneration in the CNS,17 thus consequently affect the brain functions, including emotion, learning, and memory. In addition, depression-mediated cholinergic dysfunctions might also explain the emergence of cognitive symptoms. Cognitive impairment in depression status, such as poor concentration and attention as well as impaired memory and information processing, point to deficits in cholinergic function, which is mainly regulated by acetylcholine (ACh).18 The evidence from previous research has confirmed that a higher level of ACh in the hippocampus was correlated with increased depression.19
Moringa oleifera (MO) has various biological activities including anti-inflammatory, antioxidant and neuroprotective properties.20,21 These pharmacological effects are associated with ingredients such as phenolic acids, flavonoids (quercetin, iso quercetin, isothiocyanates and glycosides, apigenin, myricetin, rhamnetin, kaempferol), alkaloids, phytosterols, glucosinolates, glycosides, minerals, organic acids, and vitamins.22,23 MO supplemented diet significantly ameliorated oxido-inflammatory stress, restored cholinergic transmission via acetylcholinesterase inhibition and maintains neuronal integrity in the brain in scopolamine-induced spatial memory deficit in mice.24 Moreover, long-term consumption of MO progressively increases the body weight after the 12 weeks, improved spatial and non-spatial memory performance, protect against oxidative stress, inhibit AChE activity and suppresses neuronal degeneration in the hippocampus of rats.25
In our previous study,26 we have reported that palmitic acid, oleic acid, stearic acid, stigmasterol, and β sitosterol were abundant on the seed oil from MO (MOO), and confirmed the neuroprotective effects of MOO by improved the memory impairment caused by scopolamine injection through the inhibition of AChE activity and overexpression of NF-κB protein level, and modulate the level of Trk-B expression. Similarly, the administration of 70% ethanol extract of MO seeds could improve memory and cognitive impairment in scopolamine mice model. These effects were mediated through enhancement of the cholinergic neurotransmission system and neurogenesis via activation of the Akt, ERK 1/2, and CREB signaling pathways.27
To recapitulate an external chronic stress in the animals, we used water immersion restraint-induced stress (WIRS) model. WIRS is a severe stress model that combines psychological and physical stress.28 Previous studies reported WIRS induction was used to investigate various effects resulting from chronic stress in animal model, such as changes in emotional behavior,29 memory function and learning,30 gastric mucosal lesion,31 and post-traumatic stress disorder.32 Moreover, repeated of WIRS procedure, may also exacerbate depression and sleep disturbance.33
The aim of this study was to investigate the effects of MOO on anxiety-like, depression-like and memory impairment phenotypes in WIRS mice model. Mechanistically, oxidative stress related parameters, AChE activity and BDNF expression level in the brain were also investigated. In addition, we checked the histopathological features in the gastric mucosal wall to further identify the impact of stress in targeted organ on the WIRS rodent model.
Materials and Methods
Animals, Grouping, and Treatment
The animal experiments were approved by the local ethics committee of the Faculty of Medicine of Universitas Indonesia with the reference number KET-1405/UN2.F/ETIK/PPM.00.02/2020. Twenty-four male BALB/c mice weighing 25–30 g, were obtained from the animal house of Biopharma laboratory and animal breeding, Bandung, Indonesia. All mice were kept in closed system cages in an environment with a constant temperature of around 25°C and a 12-hour light/12-hour dark cycle at the Animal Research Facility, Institute of Medical and Research, Faculty of Medicine Universitas Indonesia, Jakarta, Indonesia. Food and water were available ad libitum. The mice were divided into four groups (six animals per group) as follows: normal (control, without WIRS intervention) group, WIRS (mice with WIRS intervention, treated with vehicle) group, WIRS+MOO1 (mice with WIRS intervention and treated with MOO at 1 mL/kg BW/day) group, and WIRS+MOO2 (mice with WIRS intervention and treated with MOO at 2 mL/kg BW/day) group. The doses of MOO were chosen based on our previous report.23 The MOO was obtained from PT Keloria (Medan, Indonesia) as previously reported.23 In brief, the fresh MO seeds were oven-dried for seven days and were separately blended with a seed blender, and the melted seeds were independently poured into an oil extraction machine that separated the oil from the residue at high temperature and pressure. The oil was then kept at about 60–100°C under degraded pressure to maintain the constant temperature in the oil.23 One milliliter of MOO was diluted with 10 mL sunflower oil. The MOO and vehicle were administered orally for 23 days. The time line of this study was illustrated in Figure 1.
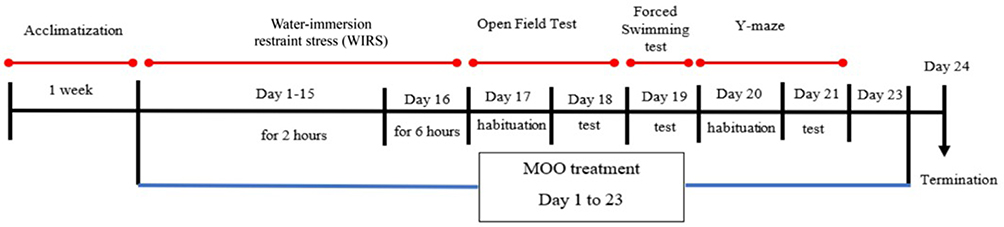 |
Figure 1 Schematic representation of the in vivo experiment. Normal group: mice control, without WIRS intervention (N=6); WIRS group: mice with WIRS intervention, treated with vehicle (N=6); WIRS+MOO1: mice with WIRS intervention, treated with MOO at dose 1 mL/kg BW (N=6); WIRS+MOO2: mice with WIRS intervention, treated with MOO at dose 2 mL/kg BW (N=6). Abbreviations: WIRS, water-immersion restraint stress induced; MOO, Moringa oleifera seed oil. |
Stress Protocols
In this study, we conducted the WIRS protocol to produce models of stress animals, since WIRS is a combination of restraint stress and water immersion stress, which leads to the emergence of behaviors such as depression34 and includes psychological and physical stimulations.35 Mice in the groups WIRS, WIRS+MOO1 and WIRS+MOO2 were subjected to WIRS protocol as previously reported by Yasugaki et al, with minor modifications.33 In brief, mice were restrained by a 50 mL conical polypropylene centrifuge tube containing multiple holes. Each tube was placed on a modified plastic tube rack, and then the tube was immersed in a vertical position in the water, with water height to the limit of xiphoid. Water temperature was adjusted to 22.0±1.0°C. Mice were subjected to this stress session once a day for 15 days in the duration of two hours. On day 16, mice were exposed to this session for six hours (Figure 1). For control mice (normal group), instead of exposure to two hours of WIRS, the mice were placed in a novel cage for two hours with no food or water.
Behavior Analysis
The animals were subjected to the behavior analysis during the light phase. Behavior tests were performed one day after WIRS protocol (Figure 1) as follows: open field test (OFT), followed by forced swim test (FST), and finally a Y-maze test.
Anxiety-like Measurement in OFT
We performed OFT to assess spontaneous locomotor activity and anxiety-like behavior of animals in new surroundings.36 The OFT was performed as described previously by Seibenhener and Wooten, with some modifications.37 The OFT was consisted of an area of an apparatus measured 40 cm (length) × 40 cm (width) × 40 cm (height) with a video camera installed above the apparatus. Red stripes were drawn on the floor with markers. The mice were placed in the middle of the bottom surface of the apparatus, and their ambulatory movements were recorded for 10 min and analyzed using ANY-maze® software. The anxiety level was determined by calculated the time spent in the central area (TCA) and the number of returns to the center (NRC).38
Depression-like Measurement in FST
To analyze the depression-like behavior in our experimental mice, we next performed FST following OFT. The protocol was carried out according to Yankelevitch-Yahav et al, with some modifications.39 In brief, we used an apparatus consisting of transparent cylindrical glass containers measuring 25 cm in height and diameter of 15 cm. The immobility time (in second) parameters were observed and recorded during four-minute sessions. Immobility time described as a time that the animal spends without movement, only moves to keep its head out of the water.40 Physical immobility is an indicator of behavioral despair, and the percentage of immobility time represent a depressive-like response.41,42
Spatial Working Memory Measurement by Y-maze Test
Finally, we also conducted the Y-maze test to check the spatial working memory in our test animals according to method described by Kraeuter et al.43 The apparatus was made of black colors fiber plastic, a Y-shaped compartment (35×15×17 cm), with an angle between arms of 120°. The percent (%) of alternation was calculated using the formula: % alternation=(the number of alternations/total number of arm entries–2)*100.
Blood and Tissues Collection
On the day 24, mice were decapitated 30 min following an intraperitoneal injection of ketamine/xylazine, and blood was collected for further biochemical parameter analyses. The whole brains and stomach tissues were quickly removed, the hippocampus and the prefrontal cortex were carefully separated from the whole brain, rinsed with ice-cold buffered saline and stored at −80°C. Plasma was separated by centrifugation at 3000 rpm for five minutes at 4°C.
Cortisol Plasma Level
To measure the concentration of cortisol, we used a Mouse/Rat Cortisol kit (USA R&D system®, Inc.) according to the manufacturer’s protocol, using a microplate reader at 450 nm.
Acetylcholine Esterase (AChE) Activity in Hippocampus Tissue
The hippocampus tissues were checked for their AChE activities using a mouse-ELISA kit (Sigma-Aldrich, St Louis, MO, USA), according to manufacturer’s protocol. The hippocampus tissues were homogenized in 0.1 M phosphate buffer, pH 7.5 followed by centrifugation at 14.000 rpm for five minutes. The color intensity obtained was proportional to the AChE activity in each sample and was expressed as a unit of enzymatic activity (U/L). One unit of AChE was an enzyme that catalyzes the production of 1.0 μ moles of thiocholine per minute at room temperature at pH 7.5.
Assessment of Malondialdehyde (MDA) Level and Superoxide Dismutase (SOD) Activity
In this study, the lipid peroxidation of the homogenized prefrontal cortex tissue samples was determined by the method described by Wills.44 Prefrontal cortex MDA was estimated according to the procedure of MDA-microplate assay kit (MyBiosource, Inc., San Diego, USA, 822354). The MDA level of each sample was calculated according to the protein concentration of each sample and expressed in nmol/mg protein. The protein in each sample was checked using the Bradford method.45 The SOD activity in the prefrontal cortex tissue was determined using the commercial kit (ab65354, colorimetry, Abcam, Cambridge, UK) according to manufacturer’s protocol. SOD activity was expressed as the inhibition rate (%) of the formation of water-soluble formazan from water-soluble tetrazolium and a superoxide anion generated from the xanthine-xanthine oxidase system.46
BDNF mRNA Expression
Total RNA was extracted from the hippocampus tissue sample using a total RNA mini kit (Geneaid) according to the manufacturer’s instructions. The total RNA was quantified on a NanoDrop spectrophotometer (BioDrop, UK) with a wavelength of 260 nm. Quantitative real time-polymerase chain reaction (qRT-PCR) assays were performed using similar procedure from Nugroho, et al.47 The BDNF mRNA expression analysis was performed using qRT-PCR applied Biosystem with ReverTra® qPCR RT Master Mix/gDNA remover kit (Toyobo Bio-Tech). The presence of BDNF and GAPDH was detected with a primary from: BDNF-NeuroD1, forward 5’-AAGCCATGAATGCAGAGGAGGACT-3’; reverse: AGCTGCAGGCAGCCGGCGACC), GAPDH forward 5’-TGCACCACCAACTGCTTAGC-3’ and reverse 5’-GGCATGGACTGTGGTCATGAG-3’. GAPDH and BDNF primers were the same as in Mir et al.48 Primer was design and manufactured by Integrated DNA Technologies, Inc. (Coralville, IA, USA). The 2−ΔΔCT method was used to evaluate relative expression level.
Histopathological Analysis of Gastric Sample
The stomach of the mice was separately fixed in paraformaldehyde, dehydrated using a series of alcohol, cleared in xylene and then treated with paraffin imbedding. Hematoxylin and eosin staining was performed to observe the histopathological changes of these stomach samples under a light microscope. To evaluate the gastric mucosal wall injury, we determined the ulcer scores (0 to 5 points), blindly, according to previous report.49
Statistical Analysis
The results were presented as mean ±standard error of the mean (SEM). Data were analyzed using one-way analysis of variance was used, followed post hoc analysis using by Tukey’s test. Differences with p-value less than 0.05 was considered significant. The data were analyzed using SPSS version 26 for statistical analysis and graph were presented in GraphPad version 9.00.
Results
Open Field Test Analysis
Analysis of the locomotor activity of rats in the OFT showed different behavioral pattern in the normal, WIRS, WIRS+MOO1 and WIRS+MOO2 groups as shown in representative traces of ambulatory movements in the area used of this test (Figure 2A). In addition, mice induced with WIRS protocol for two weeks spent significant less TCA and NRC compared to normal group (p=0.001), indicating an anxiety-like behavior. Administration of MOO for 23 days at the dose of 1 mL/kgBW (WIRS+MOO1 group) ameliorated these effects by increased the TCA and NRC significantly (p=0.014 and p=0.0018, respectively) compared to the WIRS group. However, we found no significant difference in TCA and NRC values in WIRS+MOO2 group compared to WIRS group (Figure 2B and C). See also the Supplementary Figures.
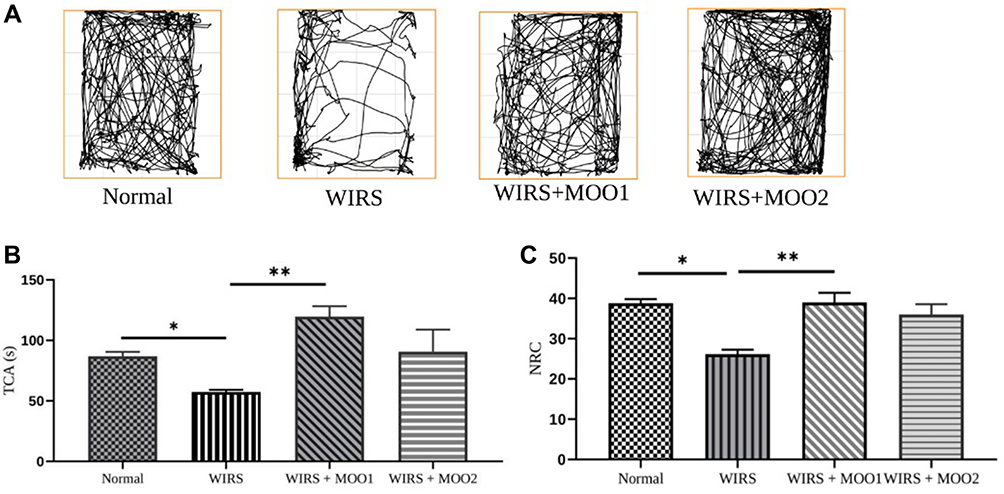 |
Figure 2 (A) Representative traces in the OFT during a period of 10 min. Behavioral tests were performed on day 24; effect of MOO on anxiety-like measurement as determined by OFT (B) TCA; (C) NRC. The values are presented as means ±SEM; *p<0,05 compared to the normal group, and **p<0.05 compared to WIRS group. Abbreviations: MOO, Moringa oleifera seed oil; NRC, number of returns to the center; OFT, open field test; TCA, time spent in the central area; WIRS, water-immersion restraint stress-induced, WIRS+MOO1, WIRS+Moringa seed oil 1 mL/kg BW); WIRS+MOO2 (WIRS+Moringa seed oil 2 mL/kg BW) groups. |
Forced Swim Test Analysis
Exposure to stress using WIRS protocol caused significant increase of immobility time compared to normal group (p=0.001). Interestingly, WIRS mice treated with MOO at two different doses provoked significant decrease in mobility time of mice compared to the WIRS group (p=0.07) (Figure 3A). This result can be assumed that the behavioral despair was ameliorated.
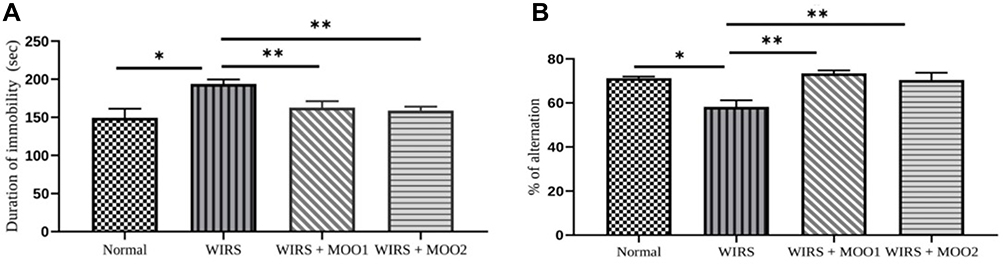 |
Figure 3 Effect of MOO on depression like measurement as determined by FST. (A) Immobility time in seconds; and effect of MOO on spatial working memory measurement as determined by Y-maze test (B) Spontaneous alternation percentage. The values are presented as means ±SEM; *p<0,05 compared to the normal group, and **p<0.05 compared to WIRS group. Abbreviations: FST, forced swimming test; MOO, Moringa oleifera seed oil; WIRS, water- immersion restraint stress-induced, WIRS+MOO1, WIRS+Moringa seed oil 1 mL/kg BW); WIRS+MOO2 (WIRS+Moringa seed oil 2 mL/kg BW) groups. |
Analysis of Spatial Memory Using Y-maze Test
Y-maze test was carried out to determine spatial memory ability of mice. Exposure mice to WIRS protocol caused significantly decreased of alternation percentage compared to the normal group (p=0.001). The significant increase of alternation percentage in mice treated with MOO (in both doses, p=0.000 and p=0.002, respectively) compared with WIRS group indicating the improvement of the spatial memory ability of those mice (Figure 3B).
Plasma Cortisol Concentration in WIRS Mice Model
To analyzed the hormonal alteration in this animal stress model, we next examined the plasma cortisol level. Unexpectedly, we could not find any significant difference between groups (p=0.975) in level of plasma cortisol (Figure 4).
 |
Figure 4 Effect of MOO on plasma cortisol concentration (ng/mL). The values are presented as means ±SEM. Abbreviations: MOO, Moringa oleifera seed oil; WIRS, water-immersion restraint stress-induced, WIRS+MOO1, WIRS+Moringa seed oil 1 mL/kg BW); WIRS+MOO2 (WIRS+Moringa seed oil 2 mL/kg BW) groups. |
The Effect of MOO on AChE Activity in Hippocampus Tissue
To analyze whether the cholinergic system was involved in this animal model and to check the MOO ability to modify this enzyme activity, we next performed the AChE activity in hippocampus tissues. As shown in Figure 5, the AChE activity in WIRS group increased significantly in comparison with the normal group (p=0.001). As expected, the activity of AChE in the WIRS+MOO1 and WIRS+MOO2 groups showed significant decreased in comparison with WIRS group (p=0.042 and p=0.019), respectively.
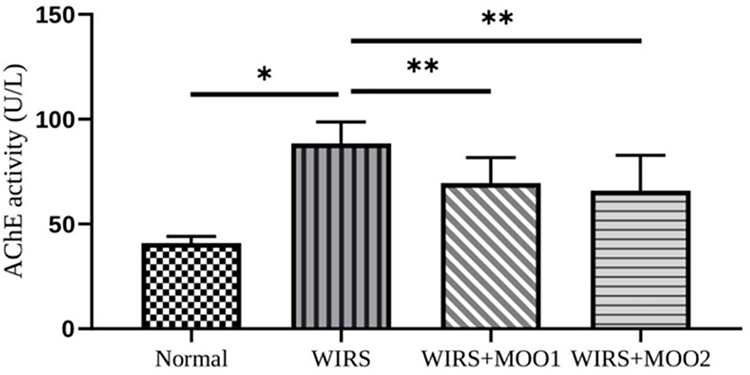 |
Figure 5 Effect of MOO on AChE activity (U/L) in hippocampus tissues. The values are presented as means ±SEM. *p<0,05 compared to the normal group, and **p<0.05 compared to WIRS. Abbreviations: AChE, acetylcholine esterase; MOO, Moringa oleifera seed oil; WIRS, water-immersion restraint stress-induced, WIRS+MOO1, WIRS+Moringa seed oil 1 mL/kg BW); WIRS+MOO2 (WIRS+Moringa seed oil 2 mL/kg BW) groups. |
Analysis of MDA Level and SOD Activity in Prefrontal Cortex Tissue
Mice exposed to the stress without treatment showed a significant increase of MDA level in the prefrontal cortex tissue compared to the normal group. However, the SOD activity was also increase in WIRS group compared to the normal group. Cotreatment with MOO, both in two doses ameliorated the MDA level, but did not affect the SOD activity in those groups (Table 1).
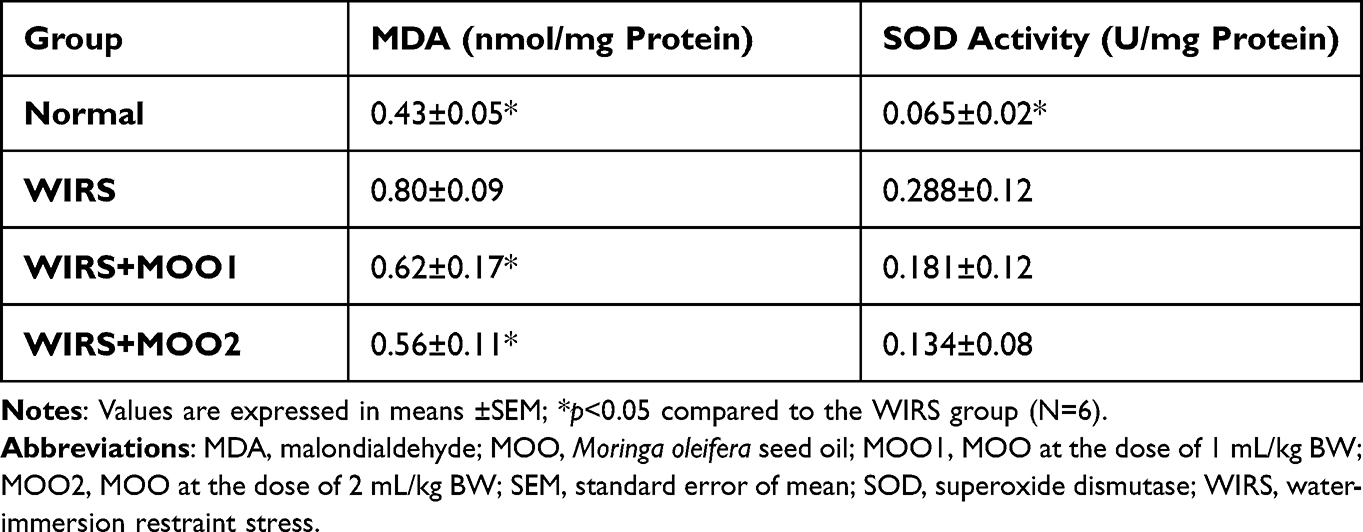 |
Table 1 Effect of Moringa Oleifera Seed Oil on Oxidative Stress Parameters in Prefrontal Cortex Tissues of Mice |
The Effect of MOO in BDNF mRNA Expression Level in Hippocampus Tissue Derived from WIRS Mice Model
The transcription of BDNF gene in the mouse hippocampus was comparable between WIRS group and normal group. However, oral administration of MOO at the dose of 2 mL/kg BW was significantly enhanced the BDNF mRNA expression level. Meanwhile, the WIRS+MOO1 group was not able to enhance the mRNA expression level of BDNF (Figure 6).
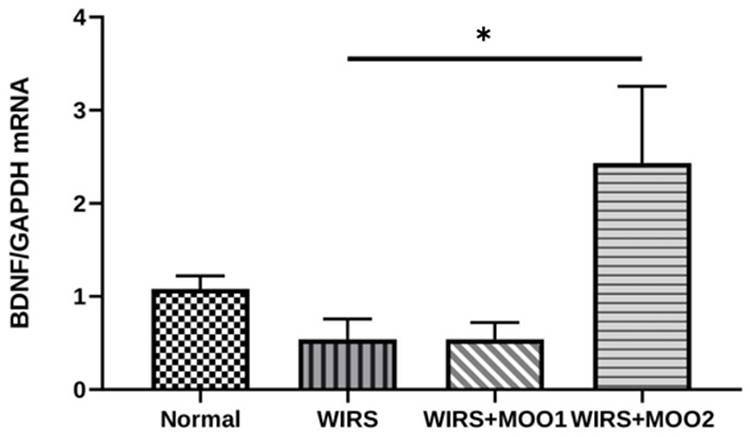 |
Figure 6 Effect of MOO on BDNF mRNA expression level in hippocampus tissues. The values are presented as means ±SEM. *p<0.05 compared to WIRS. Abbreviations: BDNF, brain-derived neurotrophic factor; MOO, Moringa oleifera seed oil; WIRS, water-immersion restraint stress-induced, WIRS+MOO1, WIRS+Moringa seed oil 1 mL/kg BW); WIRS+MOO2 (WIRS+Moringa seed oil 2 mL/kg BW) groups. |
The Effect of MOO in Ulcer Lesion Index and Histology Analysis
We next investigated whether the WIRS did indeed induce the stress phenotype by analyzing the histopathology features in the target organ such as gastric mucosal wall. Of note, gastric isolated from WIRS group (Figure 7B) showed the ulcer appearance, neutrophil infiltration, and sub mucosal dilatation while these phenotypes did not occur in the normal group (Figure 7A). Meanwhile, WIRS mice model treated with MOO exhibited minimal ulcer phenotype and less neutrophil infiltration (Figure 7C and D). These results were further confirmed by the significant improvement of ulcer lesion index in both WIRS+MOO1 and WIRS+MOO2 groups compared with the WIRS group (p<0.05 and p<0.05), respectively (Figure 8).
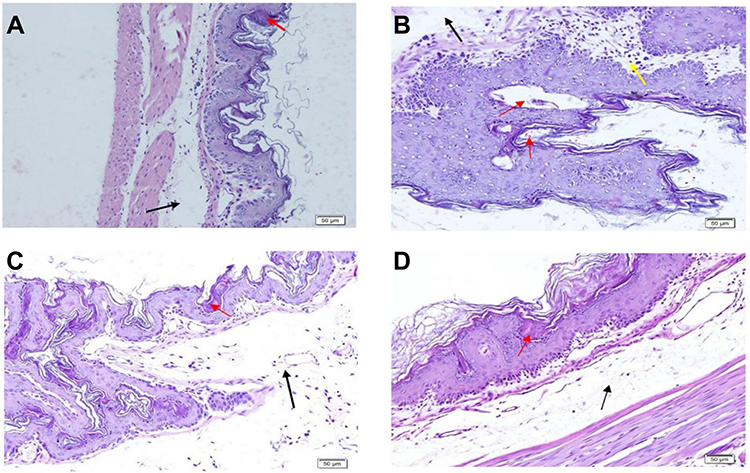 |
Figure 7 Effect of MOO on gastric ulcer induced by WIRS model assessed by hematoxylin and eosin staining in gastric mice (A) normal, (B) WIRS, (C) WIRS+MOO1, and (D) WIRS+MOO2 groups. The black arrow indicates the dilatation of the submucosal layer, red arrow shows ulcer appearance, and yellow arrow represents mononuclear cell and neutrophil infiltration. Bars: 50 µm. Abbreviations: MOO, Moringa oleifera seed oil; WIRS, water-immersion restraint stress-induced, WIRS+MOO1, WIRS+Moringa seed oil 1 mL/kg BW); WIRS+MOO2 (WIRS+Moringa seed oil 2 mL/kg BW) groups. |
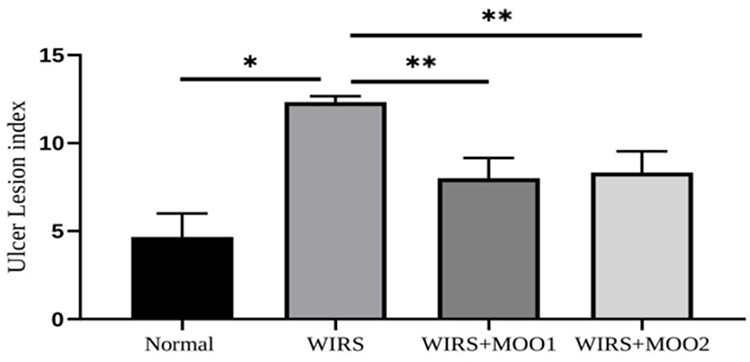 |
Figure 8 Effect of MOO on gastric ulcer calculated by ulcer lesion index. The values are presented as means ±SEM. *p<0,05 compared to the normal group, and **p<0.05 compared to WIRS. Abbreviations: MOO, Moringa oleifera seed oil; WIRS, water-immersion restraint stress-induced, WIRS+MOO1, WIRS+Moringa seed oil 1 mL/kg BW); WIRS+MOO2 (WIRS+Moringa seed oil 2 mL/kg BW) groups. |
Discussion
The present study demonstrated that an oil formulation derived from MO seeds showed neuroprotective effects by amelioration anxiety-like, depression-like and memory impairment phenotypes in the WIRS mice model via enhancing mRNA expression level of BDNF, inhibiting the AChE activity, and improving the antioxidant capacity. Furthermore, MOO may also have additional value to minimize the gastric lesion caused by WIRS induction. Given that the interrelation between oxidative stress, cholinergic pathway, and neuroplasticity is involved in the pathogenesis of stress induced behavioral impairments, we propose that MOO may be a valuable remedy for symptom-modifying supplement against stress-induced behavioral abnormalities.
In our study, we performed three behavioral tests to explore the symptoms modifying potency of MOO against WIRS-induced stress in mice. Mice exposed to stress showed decreased values of TCA and NRC in the OFT procedure, indicating the anxiety-like behaviors;36 increased the immobility time in the FST procedure, indicating the depressive-like performance39 and further decreased the percentage of alternation in the Y-maze test, indicating the impairment of spatial working memory.43 Interestingly, oral administration of MOO significantly increased TCA and NRC, and decreased the immobility time along with improved spatial working memory in the mice exposed to WIRS protocol (Figure 2). These improvements of such behavioral tests indicate that MOO may have an anxiolytic, and antidepressant effect and be found to alleviate memory deficits caused by WIRS protocol. In line with the result of our study, previous study reported that administration of alcoholic extract of MO, in combination with low dose fluoxetine, has an additional antidepressant effect in depressive mice.50
Accruing data suggested an established link between ROS and the pathophysiology of behavioral disorders.17 Therefore, the affective disorder and the impaired memory observed in our study following WIRS protocol were further accompanied by increased ROS as evidenced by increased lipid peroxidation (MDA level) significantly in the prefrontal cortex tissue of mice (Table 1). It is well-known that the brain is vulnerable to ROS due its immense oxygen utilization with a minimal antioxidant defense.51 As observed in this study, we found that administration of MOO exposed with WIRS protocol significantly reduced the MDA level in prefrontal cortex tissue and as a consequence improved the memory function and emotional behavior. In contrast with the MDA level, we observed that the SOD activity was increased both in the WIRS group and the group with MOO treatment. The activity level of antioxidant enzymes in the brain seemed to vary depending on the its pathologic condition.52 A study by Popovic et al showed that chronic restraint stress in mice increased the activity of monoamine oxidase (MAO) A and MAO B, as well as increased protein levels of catechol-O-methyltransferase probably indicated increased catecholamine degradation, which was followed by increased activity of SOD1, SOD2 and catalase, as well as decreased activity of glutathione peroxidase under stress conditions.53 The lack of the performing component of SOD (SOD1 and SOD2) in our study, either in mRNA or in protein expression was considered a limitation of this study.
This neuroprotective effect of MOO might be due to the active components of MOO namely palmitic acid, oleic acid, stearic acid, stigmasterol, and β sitosterol which has been reported in our previous study.26 Similarly, MO leaves extract seemed to have a potential cognitive enhancer via the decreased oxidative stress and the enhanced cholinergic function.24,25,54,55
In our study, we evaluated the activity of AChE and mRNA expression level of BDNF in hippocampus tissue of the mice. Central cholinergic system particularly in hippocampus plays a significant role in regulation of learning and memory, and it was reported that chronic predictable stress influenced the AChE activity in hippocampus along with the impairment of cognitive function.56 In addition, BDNF is an important factor for neuroprotection and participated as a mediator in survival neurons from neuronal damage resulting from stress.57 Consistently, mice exposed with WIRS protocol showed a significant increase AChE activity, even though the mRNA expression level of BDNF was similar to the normal group (Figure 6). Of note, administration of MOO inhibited the activity of AChE and enhanced the mRNA expression of BDNF. The impact on AChE activity and BDNF expression after administration of MOO suggested that MOO may have the ability to modify central cholinergic system and improved neuronal perturbation caused by stress. In line with our study, Zhou et al reported administration of MO seed extract may improve cognitive in scopolamine-induced mice via cholinergic neurotransmission system pathways and neurogenesis.27
To analyze the hormonal alteration in this stress animal model and support the other stress phenotype such as gastric ulcer, we checked the plasma cortisol level along with histopathological analysis of the gastric mucosa wall, since WIRS has previously been utilized to induce a gastric ulcer.58–61 Unexpectedly, we could not find any significant difference in the plasma cortisol level between groups. This contradictive data might be due to the dynamic of cortisol level in stress condition, since previous experimental study in animals reported that corticosterone hormone was more remarkably affected than cortisol level under stressful conditions.62 The lack to performing the corticosterone level in our study was considered a limitation of this study.
In contrast, we detected an anti-ulcer effect of MOO by shown the improvement of ulcer index and histopathological features in gastric (Figures 7 and 8). Results from previous study also reported leaves extract of MO improved the healing process of gastric and duodenal ulcers in rats.63
Conclusion
In conclusion, these finding suggested that MOO may has a neuroprotective effect in mice model of WIRS-induced stress as evidenced with improving the abnormal behaviors through enhancing mRNA expression level of BDNF, inhibited AChE activity, and reduced the MDA level in the brain. Future research is needed to explore the effect of MOO in the downregulation of the BDNF signaling pathway along with the details of hormones that regulated physical and physiological stresses.
Acknowledgments
The authors thank Christian Marco and Annisa Ulhuriyah for their excellent technical assistance.
Author Contributions
All authors made a significant contribution statement to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by grants from Kangwon National University 2019 and Hibah Penelitian Disertasi Direktorat Jenderal Pendidikan Tinggi, Riset, and Teknologi Republic Indonesia (NKB-923/UN2.RST/HKP.05.00/2022).
Disclosure
Mrs Emni Purwoningsih reports grants from Republic of Indonesia, during the conduct of the study; grants from Republic of Indonesia and Kangwon National University, outside the submitted work. Professor Abdul Munim reports grants from Ministry of Education, outside the submitted work. The authors report no other conflicts of interest in this work.
References
1. Sen A, Kara AY, Koyu A, Simsek F, Kizildag S, Uysal N. The effects of chronic restraint stress on empathy-like behaviour in rats. Neurosci Lett. 2021;765:136255. doi:10.1016/j.neulet.2021.136255
2. Park C, Rosenblat JD, Brietzke E, et al. Stress, epigenetics and depression: a systematic review. Neurosci Biobehav Rev. 2019;102:139–152. doi:10.1016/j.neubiorev.2019.04.010
3. Sapolsky RM. Stress and plasticity in the limbic system. Neurochem Res. 2003;28(11):1735–1742. doi:10.1023/a:
4. Holmes A, Wellman CL. Stress-induced prefrontal reorganization and executive dysfunction in rodents. Neurosci Biobehav Rev. 2009;33(6):773–783. doi:10.1016/j.neubiorev.2008.11.005
5. Lupien SJ, Maheu F, Tu M, Fiocco A, Schramek TE. The effects of stress and stress hormones on human cognition: implications for the field of brain and cognition. Brain Cogn. 2007;65(3):209–237. doi:10.1016/j.bandc.2007.02.007
6. Moreira PS, Almeida PR, Leite-Almeida H, Sousa N, Costa P. Impact of chronic stress protocols in learning and memory in rodents: systematic review and meta-analysis. PLoS One. 2016;11(9):e0163245. doi:10.1371/journal.pone.0163245
7. Mikulska J, Juszczyk G, Gawrońska-Grzywacz M, Herbet M. HPA axis in the pathomechanism of depression and schizophrenia: new therapeutic strategies based on its participation. Brain Sci. 2021;11(10):1298. doi:10.3390/brainsci11101298
8. Peavy GM, Jacobson MW, Salmon DP, et al. The influence of chronic stress on dementia-related diagnostic change in older adults. Alzheimer Dis Assoc Disord. 2012;26(3):260–266. doi:10.1097/WAD.0b013e3182389a9c
9. Peavy GM, Salmon DP, Jacobson MW, et al. Effects of chronic stress on memory decline in cognitively normal and mildly impaired older adults. Am J Psychiatry. 2009;166(12):1384–1391. doi:10.1176/appi.ajp.2009.09040461
10. Qin D, Rizak J, Feng X, et al. Prolonged secretion of cortisol as a possible mechanism underlying stress and depressive behaviour. Sci Rep. 2016;6(1):30187. doi:10.1038/srep30187
11. Lu B, Nagappan G, Guan X, Nathan PJ, Wren P. BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases. Nat Rev Neurosci. 2013;14(6):401–416. doi:10.1038/nrn3505
12. Murakami S, Imbe H, Morikawa Y, Kubo C, Senba E. Chronic stress, as well as acute stress, reduces BDNF mRNA expression in the rat hippocampus but less robustly. Neurosci Res. 2005;53(2):129–139. doi:10.1016/j.neures.2005.06.008
13. Dwivedi Y. Brain-derived neurotrophic factor: role in depression and suicide. Neuropsychiatr Dis Treat. 2009;433. doi:10.2147/NDT.S5700
14. Hosang GM, Shiles C, Tansey KE, McGuffin P, Uher R. Interaction between stress and the BDNFVal66Met polymorphism in depression: a systematic review and meta-analysis. BMC Med. 2014;12(1):7. doi:10.1186/1741-7015-12-7
15. Bhatt S, Nagappa AN, Patil CR. Role of oxidative stress in depression. Drug Discov Today. 2020;25(7):1270–1276. doi:10.1016/j.drudis.2020.05.001
16. Ercan K, Gecesefa OF, Taysi ME, Ali Ali OA, Taysi S. Moringa oleifera: a review of its occurrence, pharmacological importance and oxidative stress. Mini Rev Med Chem. 2021;21(3):380–396. doi:10.2174/1389557520999200728162453
17. Maes M, Galecki P, Chang YS, Berk M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness. Prog Neuro Psychopharmacol Biol Psychiatry. 2011;35(3):676–692. doi:10.1016/j.pnpbp.2010.05.004
18. Dagytė G, Den Boer JA, Trentani A. The cholinergic system and depression. Behav Brain Res. 2011;221(2):574–582. doi:10.1016/j.bbr.2010.02.023
19. McCloskey MC, Young TJ, Anderson SM. Research article: the influence of acetylcholinesterase on anxiety- and depression-like behaviors in fluoxetine-treated male mice. Bios. 2017;88(1):29–38. doi:10.1893/BIOS-D-15-00013.1
20. Cretella ABM, Pawloski PL, Ruziska RM, et al. Expanding the anti-inflammatory potential of Moringa oleifera: topical effect of seed oil on skin inflammation and hyperproliferation. J Ethnopharmacol. 2020;254:112708. doi:10.1016/j.jep.2020.112708
21. Idoga ES, Ambali SF, Ayo JO, Mohammed A. Assessment of antioxidant and neuroprotective activities of methanol extract of Moringa oleifera Lam. leaves in subchronic chlorpyrifos-intoxicated rats. Comp Clin Path. 2018;27(4):917–925. doi:10.1007/s00580-018-2682-9
22. Alzoubi KH, Rawashdeh NQ, Khabour OF, et al. Evaluation of the effect of Moringa peregrina extract on learning and memory: role of oxidative stress. J Mol Neurosci. 2017;63(3–4):355–363. doi:10.1007/s12031-017-0986-x
23. Perviz S, Khan H, Pervaiz A. Plant Alkaloids as an Emerging Therapeutic Alternative for the Treatment of Depression. Front Pharmacol. 2016;15:7. doi:10.3389/fphar.2016.00028
24. Onasanwo SA, Adamaigbo VO, Adebayo OG, Eleazer SE. Moringa oleifera-supplemented diet protect against cortico-hippocampal neuronal degeneration in scopolamine-induced spatial memory deficit in mice: role of oxido-inflammatory and cholinergic neurotransmission pathway. Metab Brain Dis. 2021;36(8):2445–2460. doi:10.1007/s11011-021-00855-9
25. Adebayo OG, Wopara I, Aduema W, Ebo OT, Umoren EB. Long-term consumption of Moringa oleifera -supplemented diet enhanced neurocognition, suppressed oxidative stress, acetylcholinesterase activity and neuronal degeneration in rat’s hippocampus. Drug Metab Pers Ther. 2021;2021. doi:10.1515/dmdi-2020-0189
26. Arozal W, Purwoningsih E, Lee HJ, Barinda AJ, Munim A. Effects of Moringa oleifera in two independents formulation and as neuroprotective agent against scopolamine-induced memory impairment in mice. Front Nutr. 2022;1:9. doi:10.3389/fnut.2022.799127
27. Zhou J, Yang W, Suo D, et al. Moringa oleifera seed extract alleviates scopolamine-induced learning and memory impairment in mice. Front Pharmacol. 2018;24:9. doi:10.3389/fphar.2018.00389
28. Hayashi Y, Tanaka J, Morizumi Y, Kitamura Y, Hattori Y. Polyamine levels in brain and plasma after acute restraint or water-immersion restraint stress in mice. Neurosci Lett. 2004;355(1–2):57–60. doi:10.1016/j.neulet.2003.10.027
29. Iijima C, Muroyama A, Mitsumoto Y. Changes in emotional behavior induced by water immersion restraint stress in mice. Neurosci Res. 2010;68:e428. doi:10.1016/j.neures.2010.07.1897
30. Kunisawa K, Kido K, Nakashima N, Matsukura T, Nabeshima T, Hiramatsu M. Betaine attenuates memory impairment after water-immersion restraint stress and is regulated by the GABAergic neuronal system in the hippocampus. Eur J Pharmacol. 2017;796:122–130. doi:10.1016/j.ejphar.2016.12.007
31. Izgüt-uysal VN, Gemici B, Birsen İ, Acar N, Üstünel İ. The protective effect of apelin against water-immersion and restraint stress-induced gastric damage. J Physiol Sci. 2014;64(4):279–289. doi:10.1007/s12576-014-0317-8
32. Tamaki K, Yamada K, Nakamichi N, Taniura H, Yoneda Y. Transient suppression of progenitor cell proliferation through NMDA receptors in hippocampal dentate gyrus of mice with traumatic stress experience. J Neurochem. 2008;105(5):1642–1655. doi:10.1111/j.1471-4159.2008.05253.x
33. Yasugaki S, Liu C-Y, Kashiwagi M, et al. Effects of 3 weeks of water immersion and restraint stress on sleep in mice. Front Neurosci. 2019;14:13. doi:10.3389/fnins.2019.01072
34. Miyata S, Koyama Y, Takemoto K, et al. Plasma corticosterone activates SGK1 and induces morphological changes in oligodendrocytes in corpus callosum. PLoS One. 2011;6(5):e19859. doi:10.1371/journal.pone.0019859
35. Fan F, Yang M, Geng X, Ma X, Sun H. Effects of restraint water-immersion stress-induced gastric mucosal damage on astrocytes and neurons in the nucleus raphe magnus of rats via the ERK1/2 signaling pathway. Neurochem Res. 2019;44(8):1841–1850. doi:10.1007/s11064-019-02818-0
36. Kraeuter A-K, Guest PC, Sarnyai Z. The open field test for measuring locomotor activity and anxiety-like behavior. Methods Mol Biol. 2019;99–103. doi:10.1007/978-1-4939-8994-2_9
37. Seibenhener ML, Wooten MC. Use of the open field maze to measure locomotor and anxiety-like behavior in mice. J Vis Exp. 2015;96. doi:10.3791/52434
38. Lamtai M, Azirar S, Zghari O, et al. Melatonin ameliorates cadmium-induced affective and cognitive impairments and hippocampal oxidative stress in rat. Biol Trace Elem Res. 2021;199(4):1445–1455. doi:10.1007/s12011-020-02247-z
39. Yankelevitch-Yahav R, Franko M, Huly A, Doron R. The forced swim test as a model of depressive-like behavior. J Vis Exp. 2015;97. doi:10.3791/52587
40. Valvassori SS, Varela RB, Quevedo J. Animal models of mood disorders: focus on bipolar disorder and depression. In: Animal Models for the Study of Human Disease. Elsevier; 2017:991–1001. doi:10.1016/B978-0-12-809468-6.00038-3
41. Carter M, Shieh J. Guide to Research Techniques in Neuroscience. Elsevier; 2015:407–408. doi:10.1016/C2013-0-06868-5
42. Can A, Dao DT, Arad M, Terrillion CE, Piantadosi SC, Gould TD. The mouse forced swim test. J Vis Exp. 2011;58. doi:10.3791/3638
43. Kraeuter A-K, Guest PC, Sarnyai Z. The Y-maze for assessment of spatial working and reference memory in mice. Methods Mol Biol. 2019;105–111. doi:10.1007/978-1-4939-8994-2_10
44. Wills E. Mechanisms of lipid peroxide formation in animal tissues. Biochem J. 1966;99(3):667–676. doi:10.1042/bj0990667
45. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72(1–2):248–254. doi:10.1016/0003-2697(76)90527-3
46. Carneiro M, Reis B, Azevedo J, et al. Glutathione transferases responses induced by microcystin-LR in the gills and hepatopancreas of the clam venerupis philippinarum. Toxins. 2015;7(6):2096–2120. doi:10.3390/toxins7062096
47. Nugroho CMH, Kurnia RS, Tarigan S, et al. Screening and purification of NanB sialidase from Pasteurella multocida with activity in hydrolyzing sialic acid Neu5Acα(2–6)Gal and Neu5Acα(2–3)Gal. Sci Rep. 2022;12(1):9425. doi:10.1038/s41598-022-13635-x
48. Mir S, Cai W, Andres DA. RIT1 GTPase regulates Sox2 transcriptional activity and hippocampal neurogenesis. J Biol Chem. 2017;292(6):2054–2064. doi:10.1074/jbc.M116.749770
49. Li Q, Yang L, Fan L, et al. Activity of Brucea javanica oil emulsion against gastric ulcers in rodents. Asian J Pharm Sci. 2018;13(3):279–288. doi:10.1016/j.ajps.2017.12.005
50. Kaur G, Invally M, Sanzagiri R, Buttar H. Evaluation of the antidepressant activity of Moringa oleifera alone and in combination with fluoxetine. J Ayurveda Integr Med. 2015;6(4):273. doi:10.4103/0975-9476.172384
51. Taniguti EH, Ferreira YS, Stupp IJV, et al. Neuroprotective effect of melatonin against lipopolysaccharide-induced depressive-like behavior in mice. Physiol Behav. 2018;188:270–275. doi:10.1016/j.physbeh.2018.02.034
52. Salim S. Oxidative stress and the central nervous system. J Pharmacol Exp Ther. 2017;360(1):201–205. doi:10.1124/jpet.116.237503
53. Popović N, Pajović S, Stojiljković V, et al. Prefrontal catecholaminergic turnover and antioxidant defense system of chronically stressed rats. Folia Biol. 2017;65(1):43–54. doi:10.3409/fb65_1.43
54. Sutalangka C, Wattanathorn J, Muchimapura S, Thukham-mee W. Moringa oleifera mitigates memory impairment and neurodegeneration in animal model of age-related dementia. Oxid Med Cell Longev. 2013;2013:1–9. doi:10.1155/2013/695936
55. Peñalver R, Martínez-Zamora L, Lorenzo JM, Ros G, Nieto G. Nutritional and antioxidant properties of Moringa oleifera leaves in functional foods. Foods. 2022;11(8):1107. doi:10.3390/foods11081107
56. Das A, Rai D, Dikshit M, Palit G, Nath C. Nature of stress: differential effects on brain acetylcholinesterase activity and memory in rats. Life Sci. 2005;77(18):2299–2311. doi:10.1016/j.lfs.2005.02.020
57. McEwen BS, Nasca C, Gray JD. Stress effects on neuronal structure: hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology. 2016;41(1):3–23. doi:10.1038/npp.2015.171
58. Ishihara T, Takada T, Shoji Y, Uedono Y, Takeyama N, Tanaka T. Hyperammonemia reduces water immersion–restraint stress gastric ulcers in rats. Gen Pharmacol Vasc Syst. 1998;31(1):87–91. doi:10.1016/S0306-3623(97)00407-2
59. Lu S, Wu D, Sun G, et al. Gastroprotective effects of Kangfuxin against water-immersion and restraint stress-induced gastric ulcer in rats: roles of antioxidation, anti-inflammation, and pro-survival. Pharm Biol. 2019;57(1):770–777. doi:10.1080/13880209.2019.1682620
60. Ahmad S, Najmi A, Kaundal M, Akhtar M. Gastroprotective effect of thymoquinone on water immersion restraint stress induced ulceration in rats. Drug Res. 2017;67(06):366–372. doi:10.1055/s-0043-103574
61. Guo S. Gastric mucosal damage in water immersion stress: mechanism and prevention with GHRP-6. World J Gastroenterol. 2012;18(24):3145. doi:10.3748/wjg.v18.i24.3145
62. Gong S, Miao Y-L, Jiao G-Z, et al. Dynamics and correlation of serum cortisol and corticosterone under different physiological or stressful conditions in mice. PLoS One. 2015;10(2):e0117503. doi:10.1371/journal.pone.0117503
63. Devaraj VC, Asad M, Prasad S. Effect of leaves and fruits of Moringa oleifera. on gastric and duodenal ulcers. Pharm Biol. 2007;45(4):332–338. doi:10.1080/13880200701212924
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Intranasal Administration of Brain-Derived Neurotrophic Factor Rescues Depressive-Like Phenotypes in Chronic Unpredictable Mild Stress Mice
Li XL, Liu H, Liu SH, Cheng Y, Xie GJ
Neuropsychiatric Disease and Treatment 2022, 18:1885-1894
Published Date: 29 August 2022
Therapeutic Potential of Moringa oleifera in Alzheimer’s Disease: A Review of Neuroprotective Mechanisms
Yu H, Abbasi AA, Khan H, Hu H, Lu P
Journal of Inflammation Research 2026, 19:606644
Published Date: 19 June 2026