Back to Journals » Breast Cancer: Targets and Therapy » Volume 14
The Notch Signaling Pathway Contributes to Angiogenesis and Tumor Immunity in Breast Cancer
Authors Jiang N ![]() , Hu Y
, Hu Y ![]() , Wang M
, Wang M ![]() , Zhao Z
, Zhao Z ![]() , Li M
, Li M
Received 1 June 2022
Accepted for publication 8 September 2022
Published 27 September 2022 Volume 2022:14 Pages 291—309
DOI https://doi.org/10.2147/BCTT.S376873
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Pranela Rameshwar
Nina Jiang,1,* Ye Hu,1,* Meiling Wang,2,* Zuowei Zhao,2 Man Li1
1Department of Oncology, the Second Hospital of Dalian Medical University, Dalian, Liaoning, People’s Republic of China; 2Department of Breast Surgery, the Second Hospital of Dalian Medical University, Dalian, Liaoning, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zuowei Zhao, Department of Breast Surgery, the Second Hospital of Dalian Medical University, Dalian, Liaoning, People’s Republic of China, Tel +86-0411-84671291, Fax +86-0411-84671230, Email [email protected] Man Li, Department of Oncology, the Second Hospital of Dalian Medical University, Dalian, Liaoning, People’s Republic of China, Tel +86-0411-84671291, Fax +86-0411-84671230, Email [email protected]
Abstract: Breast cancer in women is the first leading tumor in terms of incidence worldwide. Some subtypes of BC lack distinct molecular targets and exhibit therapeutic resistance; these patients have a poor prognosis. Thus, the search for new molecular targets is an ongoing challenge for BC therapy. The Notch signaling pathway is found in both vertebrates and invertebrates, and it is a highly conserved in the evolution of the species, controlling cellular fates such as death, proliferation, and differentiation. Numerous studies have shown that improper activation of Notch signaling may lead to excessive cell proliferation and cancer, with tumor-promoting and tumor-suppressive effects in various carcinomas. Thus, inhibitors of Notch signaling are actively being investigated for the treatment of various tumors. The role of Notch signaling in BC has been widely studied in recent years. There is a growing body of evidence suggesting that Notch signaling has a pro-oncogenic role in BC, and the tumor-promoting effect is largely a result of the diverse nature of tumor immunity. Immunological abnormality is also a factor involved in the pathogenesis of BC, suggesting that Notch signaling could be a target for BC immunotherapies. Furthermore, angiogenesis is essential for BC growth and metastasis, and the Notch signaling pathway has been implicated in angiogenesis, so studying the role of Notch signaling in BC angiogenesis will provide new prospects for the treatment of BC. We summarize the potential roles of the current Notch signaling pathway and its inhibitors in BC angiogenesis and the immune response in this review and describe the pharmacological targets of Notch signaling in BC, which may serve as a theoretical foundation for future research into exploring this pathway for novel BC therapies.
Keywords: Notch signaling pathway, angiogenesis, tumor immunity, breast cancer, PD-1/PD-L1
Introduction
Breast cancer (BC) is the most common malignant tumor in women. As of 2020, global tumor data indicated that BC incidence surpassed that of lung cancer and has developed into the major malignant tumor in women around the world.1 Clinically, different subtypes of BC lack specific molecular targets and exhibit drug resistance, and treatment is less effective. Therefore, the search for a new specific target molecule for BC therapy is a current therapeutic challenge. The previous studies demonstrated that BC is the first solid tumor in which associated with Notch was identified, and there is also an emerging opinion that Notch signaling is one of the three main culprits of breast cancer.2,3 Studies have found increased expression of members of the Notch family in BC tissues, compared to normal control tissues. Triple-negative BC (TNBC) exhibits abnormally activated Notch signaling. The overexpression of the Notch receptor is highly correlated with the invasiveness, metastasis, and drug resistance of TNBC.4,5 And several studies have indicated a crucial association between BC and members of the Notch family [Table 1].
 |
Table 1 Relationship Between Notch Receptors and Different Types of Breast Cancer |
Notch1 overexpression is associated with a poor prognosis and BC progression and the transition from ductal carcinoma in situ to invasive BC.6,7 Mutations in Notch2 can increase the incidence of BC, and the mutation rate is positively correlated with the progression of HER2-positive BC. Furthermore, the susceptibility to BC has been strongly associated with the Notch2 variant rs11249433.8–10 Notch3 overexpression is related to BC invasion and distant metastasis and is also related to the advancement of HER2-negative BC.11,12 TNBC has been reported to exhibit Notch4 expression in 55.6% of cases, while HER2-positive cases occur in 45.8% and in 25.5% of luminal cases.13,14 Highly metastatic BC cells can exhibit higher P2Y2 receptors (P2Y2R), and P2Y2R is significantly associated with Notch4 in BC.15 Numb is a negative regulator of the Notch signaling system, and the interference with the inhibition of the Notch pathway by Numb is linked to the development of a variety of cancers, most notably BC. Studies have also shown that increased expression of the Notch1 protein is associated with poor BC differentiation and survival, which may be due to the fact that half of BCs have low expression of Numb and Numb-like proteins.16,17 In general, Notch is highly expressed in normal breast tissue, and when the negative regulatory effect of Numb is inhibited, this will lead to the appearance of BC.13 Activation of Notch signaling maintains stem cell stemness in BC, and aberrant Notch signaling can affect the onset of BC by interfering with other signaling pathways in BC, indicating that Notch signaling may be negatively regulated to maintain normal mammary gland development. Mouse mammary tumor virus (MMTV)-integration of Notch1 or Notch4 into the genome results in hyperactive Notch signaling, which leads to an overactivated mammary gland and carcinogenesis.18 DLL1, DLL3, DLL4, Jagged1, and Jagged2 are all ligands for Notch receptors. DLL1-expressing breast tumor stem cells can drive chemoresistance in BC through the NF-κB pathway, and high expression of DLL3 is associated with a poor prognosis and immune infiltration for patients with invasive BC.19,20 Furthermore, the high expression of DLL4 mRNA has been associated with advanced stage BC. In addition, interference with the Jagged1/Notch1 signaling pathway has been shown to control the invasion and migration of TNBC via BRD4. Jagged2 has also been reported to regulate the expression of microRNA-200, which facilitates tumor stem cell properties and resistance to paclitaxel in TNBC cells.21–23 Thus, Notch receptors and ligands are heavily involved in BC, and abnormalities in Notch signaling can lead to the development of BC, suggesting that the Notch signaling pathway contributes significantly to BC pathogenesis.
The diversity of the tumor microenvironment results in either tumor-promoting or tumor-suppressive effects of Notch signaling in different carcinomas, but mainly exerts a pro-oncogenic effect on BC. A crucial process that affects the occurrence and development of BC is the immune response in the microenvironment, which is also closely associated with Notch signaling. These findings show that Notch pathway inhibitors may become a novel target for BC immunotherapy. We will discuss how the Notch signaling pathway regulates angiogenesis and immunity in BC diseases, as well as the potential applications of Notch pathway inhibitors, which may provide a theoretical foundation for future research into making Notch signaling an attractive target for novel BC therapies.
Notch Signaling Pathway
Unlike most other signaling pathways, both the Notch ligands and the receptors are transmembrane protein 1; therefore, the Notch pathway is initiated by a receptor–ligand interaction between two neighboring cells.24 It has also been reported that the Notch signaling pathway can also occur in the cytoplasmic membrane of the same cell.25 In mammals, there are more members of the Notch family, which consists mostly of four transmembrane protein receptors (Notch1-4) and five transmembrane protein ligands (DLL1, DLL3, DLL4, Jagged1, and Jagged2).26,27 The Notch signaling pathway is composed of Notch receptors, Notch ligands, downstream target genes, and other effectors, and it is activated by two pathways. The most recognized one is the classical CBF-1/RBP-Jκ-dependent pathway [Figure 1], which consists of the interaction of ligands and receptors from two adjacent cells. Subsequently, proteolytic cleavage of the receptor is initiated, and after the Notch receptor protein undergoes two consecutive proteolytic cleavages, the intracellular segment (NICD) is released into the cytoplasm by interacting with the transcriptional repressor family CSL (CBF1/SU(H)/LAG-1) and recruiting the nuclear transcriptional activator protein family MAML (mastermind-like), generating a ternary complex transcriptional activator (NICD-CSL-MAML), which then activates transcription of downstream target genes (Hes and Hey protein family), therefore controlling downstream transcription factor expression levels.28–36 The Notch signaling system is a receptor-ligand cascade that is extremely conserved. After Notch signaling is turned on, it influences normal cellular morphogenesis, including cell differentiation, proliferation, apoptosis, and cell border creation, among other things.37,38
 |
Figure 1 Classical mechanism of the Notch signaling pathway. Abbreviations: DLL1, Delta-like 1; DLL3, Delta-like 3; DLL4, Delta-like 4; NICD, intracellular segment; ADAM10, a disintegrin and metalloproteinase domain 10; TACE, TNF-alpha converting enzyme. |
Notch Signaling Mediates Tumor Angiogenesis in Breast Cancer
The mechanisms involved in Notch signaling-mediated angiogenesis in BC are depicted in Figure 2.
 |
Figure 2 Role of the Notch signaling pathway in angiogenesis of breast cancer. Abbreviations: DLL4, Delta-like 4; NICD, intracellular segment; TNF-α, tumor necrosis factor-alpha; VEGF, vascular endothelial growth factor; VEGF2, vascular endothelial growth factor receptor 2. |
Relevance of Notch Signaling and Breast Cancer Angiogenesis
Tumor angiogenesis, the formation of new blood vessels, is a crucial process for tumor growth and metastasis.39,40 Normally, pro-angiogenic and anti-angiogenic factors are in a tightly balanced mechanism. Once the balance is disturbed, proliferation, infiltration, and metastasis of the tumor will occur. The theory that tumor growth depends on angiogenesis has been proposed by many scholars earlier; therefore, anti-angiogenic therapy has also become an efficient strategy for BC therapy.39,41 Due to various reasons such as the variability of tumor cells, lack of targeting, and VEGF gene polymorphism, single anti-angiogenic drug therapy is not effective for BC patients, thus finding new therapeutic targets for anti-angiogenesis is urgently needed.
The Notch signaling pathway plays a crucial role in angiogenesis and is necessary for the development of normal arterial and venous systems.42,43 Mice lacking Notch1 die on embryonic day 9.5 and present severe vascular abnormalities, while Notch1 at a removing later stage of development would lead to vascular rupture.44 Notch receptors and ligands located in the arteries are critical for arterial differentiation during early embryonic stages, which is mainly completed by activation of the FOXC2/DLL4/Notch1 pathway.45,46 FOXC2 is a transcription factor that initiates DLL4 transcription and translation by binding to the DLL4 promoter region and increased expression of DLL4, which, in turn, binds to the Notch1 receptor, to primarily determine embryonic vascular differentiation into arterioles or venules during the embryonic stage.47,48 A previous study found that binding of Notch1 and DLL4 in BC cells induced transcription of the downstream target gene Hey2 and FOXC2 interacted with the Notch transcriptional activation complex containing intracellular Notch domains to increase Hey2 promoter activity, while binding of FOXC2 and the vascular endothelial growth factor (VEGF) induced activation of DLL4 and Hey2 promoters.46 Furthermore, the higher the malignancy of human BC, the higher the level of FOXC2, which may suggest that a poor prognosis of BC is associated with Notch signaling involving in FOXC2.49
To further demonstrate the proangiogenic effect of FOXC2/DLL4/Notch1 in BC, researchers constructed a FOXC2-overexpressing vector in MCF-7 with low FOXC2 expression. They found that the supernatant of the MCF-7 cell line overexpressing FOXC2 promoted angiogenesis in human umbilical vein endothelial cells (HUVEC). Further experiments verified that FOXC2 overexpression increased the expression levels of DLL4 and Notch1 mRNA, suggesting that the proangiogenic effect of FOXC2 on BC may be achieved through Notch signaling.50
By knocking down Linc-OIP5 expression in mammary carcinoma cells, Zhu et al reported that the cocultured of HUVEC with cancer cells resulted in the downregulation of YAP1 and Jagged1 expression. Furthermore, angiogenesis was found to be inhibited in cocultured HUVECs under the same conditions, indicating that Linc-OIP5 could have a regulatory effect on BC angiogenesis by suppressing Notch signaling.51
As an overexpressed gene and tumor endothelial marker in BC cells, HEYL can regulate tumor angiogenesis by modulating the expression of the breast tumor epithelium and endothelium. HEYL acts as a downstream target of Notch signaling, suggesting that regulation of the Notch pathway can affect angiogenesis in breast cancer. Based on this evidence, there is an essential association between the Notch signaling pathway and angiogenesis in BC.
Relevance of Notch Signaling and VEGF in Breast Cancer Angiogenesis
A feedback loop seemingly between Notch signaling and VEGF, with Notch signaling located downstream of VEGF, can down-regulate VEGFR2 expression when Notch signaling is activated.43,52 Inhibition of VEGFR2, in turn, leads to inhibition of tumor vascular sprouting and angiogenesis. Conversely, DLL4 expression decreases rapidly and significantly when VEGF is blocked, suggesting that DLL4 expression is dependent on sustained VEGF signaling in tumor vessels.53 HUVECs overexpressing DLL4 using retroviruses were engineered in one study. Compared to controls, proliferation and migration of the cells overexpressing DLL4 were inhibited, while overexpression of DLL4 negatively regulated the expression of VEGFR2, which is the main signaling receptor of VEGF-A.54 To determine whether this change occurred through the Notch signaling pathway, a recombinant human DLL4 (RhDLL4) lacking NICD was introduced. RhDLL4 not only inhibited VEGF-A proliferation but also significantly decreased VEGFR2 expression, indicating that the Notch extracellular region was sufficient to inhibit signaling.54
The interdependent relationship between Notch and VEGF also has a role in angiogenesis of BC. Leptin stimulates angiogenesis of BC by increasing the expression of VEGF/VEGFR2 via the Notch pathway.55 Cao et al found that the expression of VEGF and Notch1 in BC tissues was higher than in normal breast tissue adjacent to cancer tissues and that the expression of VEGF and Notch1 in BC tissues exhibited a consistent positive correlation.56 Furthermore, the expression of DLL4 mRNA is positively associated with VEGF levels in BC tissues.57
Taken together, the above findings show that both Notch and VEGF receptors and ligands are significantly co-expressed in BC. This indicates that Notch signaling and VEGF play a cross-linked role in BC, and both may be involved in the regulation of angiogenesis of BC. The findings also imply that inhibiting the Notch signaling system in combination with VEGFR inhibitors could lead to novel BC therapy options.
Down-Regulation of DLL4 Has an Inhibitory Effect on Breast Cancer Angiogenesis
Highly expressed in the tumor vasculature, DLL4 and Jagged1 are the best studied Notch ligands and are considered competitive regulators of tumor angiogenesis.58,59 Although DLL4 expression is required for proper embryonic angiogenesis, it is not detected in normal breast tissues. DLL4 is thus not expressed in normal breast tissue but is significantly expressed in BC cells, according to in situ hybridization and quantitative polymerase chain reaction assays, and the expression of DLL4 mRNA and VEGF was positively correlated.21,57
Mailhos et al grafted a DLL4-knockout human BC MCF-7 cell line subcutaneously in mice and found higher expression of DLL4 mRNA in the mouse mammary carcinoma vascular system.60 DLL4, which is expressed in endothelial cells, has been linked to the differentiation of tip and stem cells during angiogenesis. DLL4 inhibits endothelial cell growth, proliferation, and metastasis by acting as a negative regulator.61–64 As a negative regulatory factor for blood vessel growth, enhancement of DLL4 can inhibit excessive sprouting and branching of blood vessels. Conversely, blocking the DLL4/Notch signaling pathway will enhance the sprouting and branching activity of tumor blood vessels, which will lead to a significant increase in tumor blood vessel density. Because the increase is “nonproductive”, both the function of blood vessels and tumor growth is inhibited.65 Zhou et al generated a bispecific monoclonal antibody (HB-32) with a high affinity for VEGF and DLL4 and can block DLL4 and human VEGF. Xenograft experiments demonstrated that treatment with HB-32 decreased BC cell growth and promoted tumor cell death more effectively than an anti-VEGF or anti-DLL4 antibody alone.66 In another study, Wang et al developed a new DLL4 targeted antibody coupling small molecular cytotoxic drugs that also exhibit an effective antitumor effect.67 Downregulation of DLL4 expression and the development of novel DLL4 blockers that inhibit vascular growth may represent a new therapeutic approach to inhibit tumor growth of BC.
Jagged1 and DLL4 Antagonize Each Other to Regulate Vascular Growth in Breast Cancer Angiogenesis
Jagged1 has an essential regulatory role in tumor angiogenesis, and the lack of Jagged1 results in the death of these mutants on embryonic day 10.5.68 Jagged1 expression is associated with the density of blood vessels in tumors, and estrogen has been found to upregulate the expression of Jagged1 and Notch1 in MCF-7 cells, which in turn promotes angiogenesis of BC and accelerates the progression of BC.69,70 A mouse model expressing Jagged1 cDNA showed that overexpression of Jagged1 promotes sprouting angiogenesis.63 Similar to DLL4, Jagged1 has a function in vascular outgrowth, proliferation, and metastasis, and it also acts as a proangiogenic factor.61–64 DLL4, which down-regulates DLL4/Notch signaling and contributes to the maintenance of VEGFR expression, may be efficiently antagonized by Jagged1 – necessary to maintain the equilibrium of mammary angiogenesis. Furthermore, Sainion et al found that tumor necrosis factor-alpha (TNF-α) decreases DLL4 expression and increases Jagged1 expression, which also promotes BC.71
Taken together, the above findings may indicate that Jagged1, a proangiogenic factor, promotes angiogenesis of BC. A new therapeutic strategy may be provided by appropriately regulating the level of DLL4 and antagonizing Jagged1 expression for Jagged1-overexpressing breast tumors.
Correlation of Notch and Major Immune Cells in Breast Cancer
Immune cells are important for normal tissue homeostasis and tumor development since they are one of the most visible components of the immune microenvironment. As an extremely “cold” immune microenvironment, the clinical prognosis of breast cancer is inextricably linked to immune infiltration, with the most critical regulators of breast cancer formation and progression being tumor-associated macrophages (TAM), followed by CD8+ T cells, CD4+ T cells, B cells, NK cells, and macrophages.72 These play a major role in resistance to tumor cells by providing important innate and acquired immunity to the breast epithelium. [Figure 3]
 |
Figure 3 (A) The mechanism of PD-1/PD-L1 signaling pathway and its role in T cells; (B)Relationship between immune cells and Notch signaling pathway in breast cancer: ① Notch signaling mediates the anti-tumor effect of IFN-γ secretion by NK cells on breast cancer. ② Notch pathway activation promotes the secretion of INF-γ by CD4+ T cells to produce pro-cancer effects. ③ Notch pathway promotes SLEC differentiation of CD8+ T cells through the Hes1-PTEN axis, thus affecting the development of breast cancer. ④ Notch pathway increases the percentage of Treg cells by upregulating the expression of FOXP3, which in turn affects the development of breast cancer. ⑤ Notch pathway activation upregulates the expression of PD-1/PD-L1, which leads to immune escape from breast cancer. ⑥ Notch pathway upregulates HES1 expression to promote T-cell development, which in turn affects the development of breast cancer. ⑦ Silencing of the Notch pathway promotes the polarization level of M2 and down-regulates the polarization level of M1, which in turn affects the development of breast cancer. Abbreviations: BC, breast cancer; NK, natural killer cells; Treg, regulatory cells; PD-1, programmed death-1; PD-L1, programmed cell death-Ligand 1; M1, M1 macrophages; M2, M2 macrophages; INF-γ, interferon-gamma. |
Notch Regulates Tumor-Associated Macrophages (TAMs) in Breast Cancer
Macrophages are the main facilitators of the innate immune response. In addition to secreting lysosomal enzymes, myeloperoxidases, and cytotoxic factors to kill target cells, they can also be altered by substances secreted by tumor cells to become tumor-associated macrophages (TAM) that regulate tumor development.73 TNBC is characterized by a large invasion of immune cells. The number of TAMs and the infiltration patterns are directly associated with the prognosis of TNBC, and high levels of TAM are inversely correlated with BC angiogenesis and survival.74 The Notch pathway, which controls the production of potent pro-inflammatory proteins in tumor cells, is linked to a poor prognosis in BC and is an important signaling pathway that defines the innate immune response of the tumor.75–78 Furthermore, Notch also moderates TAM activation in response to transforming growth factor β (TGFβ)-mediated tumor cells. Inhibition of the Notch pathway leads to malignant infiltration of TAMs and overproduction of cytokines, and TAM facilitates BC progression by further stimulating tumor angiogenesis, tumor cell migration, and invasion.79,80 By increasing Notch signaling in BC cells, Lin et al determined that lactic acid can convert human macrophages into TAMs and increase CCL5 production to promote BC metastasis and epithelial–mesenchymal transition.81
TAMs in BC include M1 and M2 types, both of which exert corresponding inhibitory or promotive effects on tumors mainly through polarization. Typically, M1 exerts a killing effect on BC cells by mainly secreting IL-12, while M2 contributes to the development of BC by primarily secreting IL-10.82,83
A strong relationship between Notch signaling and polarization of M1 macrophages has been identified. After inoculating tumor-bearing mice with macrophages generated from bone marrow monocytes cultured in vitro, the expression of M1 polarization in tumors is accompanied by the presence of additional Notch ligands and receptors.84 In addition, researchers observed significantly enhanced IL-12 expression by stimulating strong activation of Notch signaling, showing a stronger M1 polarization phenomenon.84 Yin et al found that esophageal M1 polarization was decreased following high-frequency mechanical ventilation when Notch signaling was inhibited.85
In an in vitro study, DLL4-mediated Notch signaling enhanced MCP-1 expression through NF-kB, which also caused macrophages to polarize to M1.86 Forced activation of Notch signaling may facilitate the polarization of M1, and activation of macrophages lacking Notch signaling may exhibit the M2 phenotype, with most TAMs exhibiting M2.84 Polarization of M2 polarization is a major participant in BC malignancy and metastasis. Unlike M1 polarization, Notch signaling negatively regulates M2 polarization. Singla et al reported higher levels of M2 polarization after blocking the Notch1 pathway, while Pagie et al determined that DLL4 not only promoted M1 polarization through the Notch signaling pathway but also promoted M2 polarization by inhibiting M2-specific genes or promoting their apoptosis.87,88 Tao et al discovered that Linc00514 was highly expressed in clinical BC tissues and BC cell lines and that elevated Linc00514 not only promoted proliferation and invasion of BC cells but also increased the percentage of M2 macrophages by analyzing candidate long-stranded noncoding RNAs associated with BC and performing functional tests using the GEO database.89 Therefore, according to the above studies, TAMs when combined with Notch signaling appear to have a major impact on BC development.
Notch and Natural Killer Cells
In the immunological environment of BCs, natural killer cells (NK), the principal effector cells of innate immunity, are diverse.90 NK cells not only predict BC remission rates and overall survival, but different subtypes have substantial ramifications for mammary gland growth and tolerance to treatment.91 In BC, Notch signaling is critical for the formation and function of NK cells.
RBP-J is a well-known Notch signaling transcriptional effector. Both activated NK cells and NK cells were found to be considerably decreased in peripheral blood from mice lacking RBP-J, suggesting that Notch signaling is involved in controlling NK cell development.92 Recently, in vitro experiments have shown that Jagged1, DLL1, and DLL4 can induce HUVECs to expand functional NK cells.93 Notch regulates embryonic plasticity and maturation of human NK cells in a stage-specific and stroma-dependent manner.94
MRPL13 is an influential molecule that promotes BC progression and is involved in several regulatory signaling processes in BC, including the Notch signaling pathway.95 Inhibiting MRPL13 expression reduces the ability of BC cells to invade by activating intracellular Notch signaling.
Increasing the number of NK cells96 activates the Notch pathway to increase the secretion of interferon (INF), a pleiotropic cytokine. Thus, Notch signaling can exert anti-tumor effects on BC by mediating IFN-secretion by NK cells, which implies that Notch signaling can exert anti-tumor effects on BC by mediating IFN-secretion of NK cells.97,98 Altogether, this evidence suggests that NK cells can control BC via Notch signaling.
Notch Signaling and Tumor-Specific T Cell Responses in Breast Cancer
Tumor-specific T lymphocytes, a key target in immunotherapy, contribute to tumor cell proliferation by promoting growth and clearance. T cells in their early stages are known to be double-negative (CD4- and CD8-) and can only develop into double-positive T cells (CD4+, CD8+) under particular conditions.99 The Notch signaling pathway is crucial for proper thymic T cell development before differentiation into double-positive cells, after which Notch signaling strength declines.99 Notch signaling is required for appropriate T cell development, according to several studies. Many Notch ligands are expressed in T cells at various stages and multiple transcription factors involved in T cell development, such as GATA3 and HES1, have key synergistic connections with Notch signaling. GATA3 can initiate T cell development by inhibiting the activity of the Notch pathway, and in turn, GATA3 can also promote T cell activity by inhibiting the activity of the Notch pathway.100,101 GATA3, an important transcription factor affecting BC development, progression, and metastasis, has an important link to Notch-mediated T cell activity, suggesting that Notch signaling may influence BC fate via T cell/GATA3.
HES1 has been reported to be negatively expressed in normal breast tissue but highly expressed in invasive ductal carcinoma of the breast as evidenced by immunohistochemistry. Further, knocking down HES1, an important downstream target gene of Notch, can suppress T-cell development, while overexpression of HES1 promotes T-cell development, suggesting that Notch can influence BC T-cell responses by regulating HES1 expression.102–104 These findings suggest that Notch signaling is related to the T-cell response in BC.
Notch and the Proliferation and Differentiation of CD4+ T Cells
CD4+ T cells are critical components of the human immune system. They are developed primarily into helper T cells like Th1, Th2, Th17, and Treg cells when they are young. In most helper T cells, Notch signaling is active. The exact mechanism by which Notch and T cells develop is not well understood. Rutz et al found that the knockdown of Notch1 could inhibit the ability of CD4+ T cells to activate, proliferate, and differentiate, although CD4+ T cells did not show the same situation when RBP-J expression was knocked down, which may indicate that Notch signaling has an effect on CD4+ T cells upstream of Notch signaling.105 Th1 and Th2, as the main helper T cells, mainly secrete IFN-γ and IL-4 and activation of the DLL4-Notch signaling pathway, which then increases the secretion of IFN-γ and IL-4 by CD4+ T cells. As mentioned above, IFN-γ is an important cytokine in the development of BC, and IFN-γ and IL-4 can promote or inhibit the expression of ER protein in BC cell lines, which affects the growth of BC.105,106 Experiments using CD4+ T cell subsets derived from the tumor microenvironment of patients with inflammatory BC demonstrated that DLL4 overexpression enhances Th17 cell growth and that Notch-mediated Th17 polarization is therapeutically essential for immune modulation of BC.107 Furthermore, researchers have examined normal breast tissue from parametrial or prophylactic mastectomy specimens and found that CD4+ and CD8+ were more common in BC tissue.108 Overall, these findings suggest that Notch signaling regulates the tumor immune response in BC by modulating the activity of CD4+ T cells.
Notch Regulates SLEC Differentiation of CD8+ T Cells
CD8+ T cells specifically secrete various cytokines involved in immune responses and constitute an important line of defense for antitumor and antiviral immune responses together with NK cells. After the initial proliferation, nearly 90% of CD8+ T cells die, and the rest mainly form memory T cells, which can also differentiate into short-term effector T cells (SLEC). The close link between the correct differentiation of CD8+ T cells and Notch signaling has been previously reported, but the specific mechanism is not clear.109
Because activated CD8+ T cells primarily express Notch1 and Notch2, CTL activity for antigen presentation is severely impaired in Notch1 knockout mice models. Analyses of the transcriptomes of activated CD8+ T cells revealed that more than 40% of SLEC-specific genes were reduced, implying that the Notch pathway is a key factor in SLEC differentiation of T cells.109 De Sousa et al activated the expression of HES1, an important downstream gene of the Notch signaling pathway. The inhibition of PTEN transcription revealed that Notch signaling interfered with SLEC differentiation via the HES1-PTEN axis.110
CD8+ T lymphocytes have been shown to be positively linked to the survival of BC patients. The expression of CD8+ T cells is highest in TNBC, and the expression of CD8 + T cells also predicts a decreased rate of recurrence and metastasis in TNBC, suggesting that the pro-differentiation impact of Notch on CD8+ T cells may affect BC development.111 Notch’s pro-differentiation effect on CD8+ T cells might lead to a new approach to BC immunotherapy.
Notch Signaling Regulates Tumor-Associated Fibroblasts (CAFs) in Breast Cancer
CAFs are one of the most numerous components of the BC microenvironment, and they influence all aspects of breast cancer development, growth, and metastasis, and they mainly secrete various growth factors, chemokines, and cytokines to exert pro-tumorigenic effects.112 Many studies have been published on CAFs and Notch signaling. In vitro, activation of the Notch pathway in human melanoma fibroblasts (MAFs) inhibited tumor growth significantly, according to Shao et al.113 In their study on melanoma, Du et al found that CAFs regulate the plasticity of cancer stem cells through the Notch1 signaling pathway in cells.114 In BC cells and CAFs, knocking down GPER or employing the Notch transcriptional co-activator Master-mind like-1 (DN-MAML-1) revealed that Notch signaling is involved in mediating the effects of GPER signaling.115 Pelon et al discovered four CAF subpopulations in metastatic BC lymph nodes, and further experiments revealed that heterogeneity of CAFs in axillary metastatic lymph nodes can drive BC metastasis via the Notch pathway.116 Taken together, this suggests that modulating CAFs of BC may be able to achieve better therapeutic effects by targeting Notch.
Mechanisms of the Related Pathway Between Notch and PD-1/PD-L1
PD-1/PD-L1 Pathway
Activated T cells, NK cells, and other immune effector cells express the cell surface receptor programmed cell death protein 1 (PD-1); however, tumor-specific T cells express PD-1 in significant amounts.117 PD-1, a key immunosuppressive molecule, regulates the activation of T lymphocytes, whose ligands are PD-L1 and PD-L2, and thus plays a significant role in the immunosuppressive response.118 Programmed cell death ligand 1 (PD-L1) is a transmembrane protein that binds to PD-1, which is produced in all human cells and functions as a pro-tumor factor in cancer cells by connecting to tumor cell receptors and activating their proliferation and survival signaling pathways.118,119 When PD-1 and PD-L1 are combined, the proliferation of PD-1 cells is inhibited, and cytokine secretion is also inhibited. When the inhibited cells are T cells, this indicates that the programmed death of T cells is initiated, and the tumor will acquire the function of immune escape.120–122 PD-L1 has the potential to play a role in BC, particularly TNBC. These findings suggest that the link between PD-1/PD-L1-related pathways and Notch signaling should be investigated in BC.
The Notch Pathway and the PD-1/PD-L1 Pathway Synergistically Regulate Breast Cancer
In general, BC is not a particularly immunogenic tumor, but TNBC exhibits chemoresistance and a poor prognosis, and these tumors can also express some immunotherapeutic molecular targets, suggesting that they are immunogenic, and providing a rationale for immunotherapy as a treatment of choice for these patients.123,124
PD-L1, a vital immune checkpoint molecule, has been identified in different types of tumor cells, and the relationship between PD-L1 and BC is still a hot topic. Activation of the PD-1/PD-L1 pathway can downregulate the immune system by expanding the expression of PD-L1 in BC.125 TNBCs that express PD-L1 are more aggressive in general, and there is a negative relationship between the degree of PD-L1 expression and BC survival.126–128
Multiple signaling pathways are controlled, including PD-1/PD-L1. The Notch signaling system, which is critical for T cell maturation, has been linked to PD-1/PD-L1 in several studies. Chen et al found that Notch and PD-L1 were linked by the long noncoding RNA HOTAIRM1 on one axis, meaning that PD-L1 is elevated upon activation of Notch signaling.129 Pan et al discovered that Notch signaling was involved in the regulation of PD-1 expression in previous studies related to sepsis.130 Although immune checkpoint inhibitors have produced promising results in many tumors, only a small proportion of patients with BC respond well. Whether the Notch signaling pathway, one of the many pathways that regulates the PD-1/PD-L1 axis, is mechanistically associated with PD-1/PD-L1 in BC is not yet clear, but a study by Meng et al found that treatment with anti-PD-1 antibodies combined with Notch inhibitors significantly inhibited tumor growth in BC models compared to the group administered Notch inhibitors alone, which may suggest a synergistic effect of Notch signaling with PD-1/PD-L1 in BC.131 PD-L1 overexpression in BC stem cells is partially mediated the Notch3/mTOR axis, suggesting that anti-PD-L1 therapy in combination with Notch inhibitors can successfully improve immunotherapy in BC. According to Mansour et al,132 Notch pathway inhibitors in combination with PD-1/PD-L1 inhibitors are expected to overcome the problem of immune resistance in BC.
Notch Signaling Pathway Maintains Th17/Treg Homeostasis in Breast Cancer
Treg cells are a subset of T cells that direct self-immune reactions and are recognized into characteristic sort administrative T cells and initiated administrative T cells, the former developing in the thymus and closely related to autoantibody induction, and the latter mainly a subset of T cells differentiated by initial CD4+ T cells after receiving antigenic stimulation and stimulated by different cytokines, of which are differentiated into Treg cells induced by β-transforming growth factor (TGF-β) alone, secreting TGF-β and participating in autoimmune regulation in vivo.133
The ability of Notch to promote Treg cells has been explored for a long time, and Notch signaling in melanoma cells increases FOXP3 expression through upregulation of TGF-β1, increasing the percentage of Treg cells, which appears to suppress anti-tumor immunity.134 As specified, prior Treg cells are primarily partitioned into two distinctive sorts of Treg cells, but the common highlight of both is that they both express the translation figure FOXP3, which is imperative for their development.135 Anastasi et al first observed that CD4+CD25+ Treg cells express more abundant Notch3 receptors than CD4+CD25-Treg cells and found that Notch 3 promotes Treg cell development by upregulating FOXP3 expression, furthermore, animal experiments showed that Notch signaling can also downregulate FOXP3 expression and inhibit Treg cell differentiation.136,137 Recent studies have shown that Th17 and Treg cells share a common developmental pathway, that they functionally regulate each other, and that the Th17/Treg balance is essential for maintaining immune homeostasis in vivo.138 He’s experiments found an increased Th17/Treg ratio in chronic asthmatic mice, which was positively reversed after aggressively expanding the expression of Notch ligands Jagged1 and DLL4.139 Tumor Syndecan-1 can apply immunomodulatory effects on Th17/Treg in the isolated tumor microenvironment of BC patients, and Syndecan-1 was shown to achieve such modulatory effects on breast cancer through upregulation of DLL4-mediated activation of Notch signaling.107 Recently, analytical methods capable of quantitatively measuring the activity of signal transduction pathways in tissue and blood samples have been developed, and the pathway analysis revealed that Treg cells in breast cancer have high Notch pathway activity scores, and Notch is expected to be a new target for the characterization of immune activity and immunosuppressive therapy in BC.107,140 In summary, Notch signaling has important implications for Th17/Treg in BC.
Notch Signaling Pathway Inhibitors and Treatment Prospects in Breast Cancer
Notch Signaling Pathway Inhibitors
The Notch signaling pathway, as a dominant pathway influencing cell destiny, is characterized by a promoting or inhibitory relationship with an assortment of tumors including BC. Treg acts as a double-edged sword in tumor cell resistance. Notch signaling can encourage fortification of tumor immune evasion mechanisms by advancing the activity and expression of Treg cells. Thus, Notch pathway inhibitors have become a hot spot for research in recent years and may not only become a treatment target but also is expected to bring a new powerful weapon to overcome tumor immune escape.
Based on Notch receptors and their activation mechanisms, several classes of Score pathway inhibitors have been developed [Table 2]. As mentioned above, after binding to the Notch ligand and receptor, these inhibitors will undergo proteolytic hydrolase cleavage before entering the cytoplasm, which is achieved by γ-secretase activity. Thus, γ-secretase inhibitors (GISs) block Notch intracellular region (NICD) production, thus inhibiting the occurrence of Notch downstream events.141 From DAPT with low activity in the early stage, LY411575, RO4929097, LY3039478, MK-0752, BMS906024, PF-03084014, and NMK-T-057 were synthesized successively with an almost 100-fold increase inactivity. Of these, LY411575 can bind to presenilin 1 (PS1) and induce apoptosis of HER2+ BC cells, inhibit the formation of N1ICD and N3ICD, and thus inhibit Notch signaling. In addition, LY411575 combined with death receptor-5 (DR-5) and the DLL4 monoclonal antibody can effectively target the treatment of TNBC.142,143
 |
Table 2 γ-Secretase Inhibitor and Monoclonal Antibody |
RO4929097 blocks Notch signaling by inhibiting Notch receptor catabolism, and its combination with exemestane can help ER+ metastatic BC overcome resistance to endocrine therapy.144 LY3039478 is a verbal Notch signaling inhibitor for progressed or metastatic BC that represses BC cell metastases by blocking the cleavage of the basic transmembrane space of Indent protein to diminish Notch signaling and its downstream activity.145 MK-0752-induced downregulation of Notch signaling inhibits breast tumor growth while increasing the levels of BC stem cell (BCSC) population in BC cells expressing Notch3, while IL6 induction is produced during MK-0752 inhibition of Notch3-Hey2 signaling. BC stem cells and breast tumor growth are significantly reduced on combined exposure to MK-0752 and IL6R antagonists.146 TNBC is sensitive to PF-03084014, and the Notch inhibitor PF-03084014 combined with the AKT inhibitor MK-2206 or NF-κB inhibitor can have a significant inhibitory impact on TNBC.147,148 NMK-T-057 triggers autophagic apoptosis in BC cells by restraining γ-secretase-mediated activation of Notch signaling.149
Nevertheless, GISs cannot target specific receptors; thus, they also bring about adverse effects such as nausea and vomiting, which appear to be very limiting as monotherapy drugs.140 The development of antibodies that specifically inhibit members of the Notch family indicates a step toward a solution to this problem. There are currently two main classes of such antibodies. The first blocks the binding of the Notch receptor ligand, such as OMP-21M18 and Dl1.72. OMP-21M18 is a humanized anti-DLL4 monoclonal counter-acting agent that restricts Notch signaling by blocking the interaction of DLL4 with Notch1 or Notch4 and is currently being used in patients with advanced tumors such as BC evaluated in clinical trials.150 Dl1.72 could be a novel anti-DLL1 counteracting agent that strikingly interferes with DLL1-Notch signaling in estrogen-positive BC, subsequently preventing BC cell development, multiplication, and metastasis.151 Another type of monoclonal antibody that blocks ADAM10-mediated hydrolytic cleavage of proteins, such as OMP-59R5, which inhibits ADAM-mediated cleavage in Notch 2 and Notch 3, is effective in suppressing mammary carcinoma cell growth and has also entered clinical trials.152
Natural compounds such as cimigenoside, paeoniflorin, shogaol, 3-O-(E)-p-coumaroyl betulinic, oridonin, resveratrol, and celastrol have also been described [Table 3]. Cimigenoside, a natural compound isolated and extracted from Cimicifuga dahurica, exerts a critical inhibitory impact on the expansion or metastasis of BC cells by restricting mitochondrial apoptosis mediated by the Notch signaling pathway and EMT as a γ-secretase inhibitor.153 Paeoniflorin can influence BC cell proliferation and invasion by inhibiting the Notch1 signaling pathway.154 Shogaol is an active ingredient belonging to the ginger family of spices. 6-shogaol inhibits the proliferative activity of BC cells by downregulating Notch signaling and inhibiting the expression of the Hes1 gene.155 3-O-(E)-p-Coumaroyl betulinic is a powerful inhibitor of the Notch pathway, and interferes with BC proliferation by blocking the G0/G1 stage cell cycle of BC cells.156 Oridonin, a compound derived from red terpene, has a viable anticancer activity and represses the migration and metastasis of BC by essentially inhibiting Indent receptor expression and in this manner hindering Notch signaling, thus accelerating the initiation of apoptosis in BC cells.157 Resveratrol has antitumor effects, and the expression of Notch1, Dll4 mRNA and protein in BC cells was found to be down-regulated in the cells treated with resveratrol, while the growth of breast cells was inhibited, which could suggest that resveratrol regulates the proliferation of BC by suppressing the Notch pathway.158 In TNBC stem cells, celastrol and triptolide avoid stemness by lessening Notch1 activation and expression of downstream targets Hes1 and Hey1.159
 |
Table 3 Natural Compounds That Inhibit Notch Pathway |
Notch pathway inhibitors in combination with other pathway inhibitors or in combination with chemotherapy have also played a key role in BC treatment. The combination of the Notch pathway inhibitor (IMR-1) and the Wnt pathway inhibitor (PRI-724) improves the IMR-1 and PRI-724-mediated effects on MDA-MB-231 cells by initiating apoptotic cell death, achieving dual targeting for BC treatment.160 The Notch signaling pathway often leads to antiestrogen resistance, and in clinical trials, Notch pathway inhibitors in combination with doxorubicin have been effective in reversing chemoresistance in BC and have shown promise in improving the health of patients with advanced BC.161 Currently, most Notch pathway inhibitors are not approved for clinical use, but evidence is growing rapidly supporting the treatment of BC by targeting the Notch signaling pathway.
Recent Promising Targets Related to Notch in Breast Cancer Treatment
In addition to Notch signaling pathway inhibitors, the emergence of many new potential targets for BC treatment have emerged favoring BC therapy [Table 4]. The oncogenic effect of the transcription factor CBFB on BC was recently reported to be achieved by downregulating the expression of the Notch3 receptor and inhibiting Notch signaling.162 Asparagine β-hydroxylase (ASPH), which is not expressed in normal tissues and only appears during tumorigenesis, is capable of producing and maintaining the malignant phenotype of tumor tissue. Recent studies have reported that ASPH can initiate BC progression by activating the Notch signaling cascade, which may represent a potential target to reverse the aggressive phenotype of BC.163 Histone methyltransferase (NSD3) can induce methylation of H3K36 to activate Notch signaling to drive BC progression, and NSD3 may also be a potential target for BC therapy.164 DTX1 is a single transmembrane protein with ubiquitin E3 ligase activity, and low levels of DTX1 can regulate BC growth and survival through the Notch signaling pathway and promote BC proliferation and metastasis, thus DTX1 is expected to be a marker for BC treatment.165 Knockdown of USP9x in a mouse TNBC model inhibited activation of Notch signaling and revealed an enhanced antitumor immune response and attenuation of tumor growth in TNBC, further suggesting that the USP9x/Notch pathway could become a new therapeutic goal for BC.166 Overexpression of NF-E2-related factor (Nrf2) can upregulate Notch1 expression through the G6PD/HIF-1α pathway, which in turn affects the Notch signaling pathway and the downstream gene HES1 to influence BC proliferation and can also regulate BC cell migration by affecting the expression of the EMT pathway.167
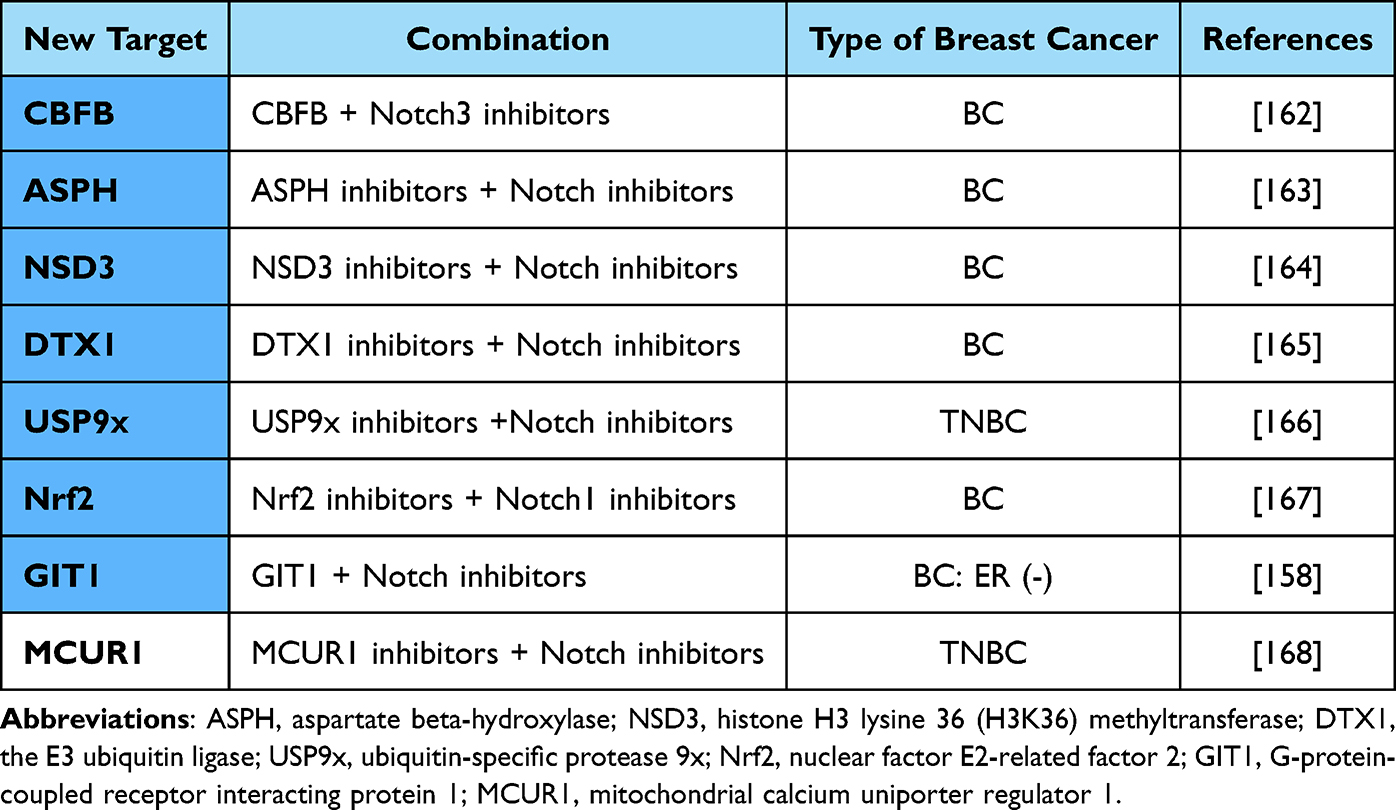 |
Table 4 Potential New Targets for Breast Cancer Treatment Linked to Notch |
GIT1 knockdown in estrogen receptor-negative breast tumor cells increases signaling and aldehyde dehydrogenase activity downstream of Notch, while overexpressed GIT1 interacts with the intracellular structural domain (ICD) of Notch and inhibits Notch signaling by inhibiting NICD transport from the cytoplasm to the nucleus, and thus, GIT1 regulates BC growth.158
Mitochondrial calcium single-exclusion regulator 1 (MCUR1) is highly expressed in TNBC, and silencing MCUR1 can inhibit the epithelial–mesenchymal transition in TNBC by suppressing Notch signaling, suggesting that MCUR1, which is involved in Notch signaling, may become a new target for TNBC therapy.168 As technology advances, additional new strategies for BC treatment will merge.
Conclusions and Perspectives
It is increasingly recognized that the Notch pathway plays a key role in the progression and metastasis of BC. In this review, the potential application value of the Notch pathway in BC is clarified by summarizing the role of Notch on BC angiogenesis as well as its impact on the immunological response. Overall, this review describes the significance of the Notch signaling pathway in the development of BC. We also provide a list of natural Notch inhibitors that may be used in the treatment of BC and further provide a rationale for the combination of Notch inhibitors in the treatment of BC.
The Notch pathway is of great significance to the angiogenesis of BC. FOXC2 is involved in the induction process of Notch signaling, and its overexpression can directly upregulate the expression levels of DLL4 and Notch1 mRNA in BC and together with the Notch pathway mediate the pro-vascular effect of BC.50 DLL4 is highly expressed in BC and may achieve inhibition of BC angiogenesis by inhibiting DLL4/Notch to promote non-functional tumor angiogenesis.65 Jagged1 also has an important role in vascular outgrowth, proliferation, and metastasis. As a pro-angiogenic factor, Jagged1 contributes to the maintenance of VEGF pathway expression by antagonizing the action of DLL4.61–64 In particular, Notch and the VEGF signaling pathways have important cross-linkages, and their combined inhibition may provide new ideas for antiangiogenic therapy in BC.53,56,169 The linkage between the Notch pathway and angiogenesis could be a potential target for the treatment of BC and deserves to be explored in depth.
There is a large infiltration of immune cells in the BC microenvironment, all of which are inextricably linked to Notch. Notch signaling plays a role in recruiting TAMs in BC.74–77 Blocking Notch signaling can block the M2 polarization process.89 Notch signaling can mediate IFN-γ secretion by NK cells to produce antitumor effects on BC, and its mediated T cell responses are likewise importantly linked to BC.97,98,102–104 DLL4 can upregulate the expansion of Th17 and exert an immunomodulatory effect on BC, the upregulated activation of Notch signaling can promote the differentiation of CD8+ T cells in BC.107,110,111 The Notch pathway can also be affected by CAFs to mediate BC metastasis, and it may have a synergistic effect with PD-1/PD-L1 in BC.115,126–128,131 Furthermore, Notch signaling can inhibit Treg cell differentiation and regulate Th17/Treg homeostasis in the BC microenvironment.137 The above results confirm that the Notch signaling pathway has important implications for BC immunity and deserves further investigation in this regard. Meanwhile, this paper summarizes the application of Notch inhibitors in BC, and most of these inhibitors have been confirmed in vitro experiments but need to be further explored in clinical trials, which also lays a good theoretical basis for clinical application.
In recent years, antitumor angiogenesis combined with immunotherapy has gradually become a promising strategy for antitumor therapy. This review reviews recent findings on Notch signaling pathways that are involved in tumor angiogenesis and immunity. Notch promises to be a prospective marker for the development of drugs related to antitumor angiogenesis and enhancement of tumor immunity or inhibition of immune escape. However, due to the diversity and complexity of the tumor microenvironment and the fact that the regulation of Notch signaling is not yet sufficiently understood, it is still necessary to investigate the role of the Notch pathway in other signaling levels with respect to BC angiogenesis and the immune response associated with carcinogenesis.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sung H, Ferlay J, Siegel RL., et al. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
2. Capaccione KM, Pine SR. The Notch signaling pathway as a mediator of tumor survival. Carcinogenesis. 2013;34(7):1420–1430.
3. Korkaya H, Wicha MSHER-2. Notch, and breast cancer stem cells: targeting an axis of evil. Clin Cancer Res. 2009;15(6):1845–1847.
4. Zhong Y, Shen S, Zhou Y, et al. NOTCH1 is a poor prognostic factor for breast cancer and is associated with breast cancer stem cells. Onco Targets Ther. 2016;9:6865–6871.
5. Giuli MV, Giuliani E, Screpanti I, Bellavia D, Checquolo S. Notch signaling activation as a hallmark for triple-negative breast cancer subtype. J Oncol. 2019;2019:8707053.
6. Yuan X, Zhang M, Wu H, et al. Expression of Notch1 correlates with breast cancer progression and prognosis. PLoS One. 2015;10(6):e0131689.
7. Ercan C, van Diest PJ, Vooijs M. Mammary development and breast cancer: the role of stem cells. Curr Mol Med. 2011;11(4):270–285.
8. Lee JH, Zhao XM, Yoon I, et al. Integrative analysis of mutational and transcriptional profiles reveals driver mutations of metastatic breast cancers. Cell Discov. 2016;2:16025.
9. Florena AM, Tripodo C, Guarnotta C, et al. Associations between Notch-2, Akt-1 and HER2/neu expression in invasive human breast cancer: a tissue microarray immunophenotypic analysis on 98 patients. Pathobiology. 2007;74(6):317–322.
10. Alanazi IO, Shaik JP, Parine NR, et al. NOTCH single nucleotide polymorphisms in the predisposition of breast and colorectal cancers in Saudi patients. Pathol Oncol Res. 2021;27:616204. doi:10.3389/pore.2021.616204
11. Hirose H, Ishii H, Mimori K, et al. Notch pathway as candidate therapeutic target in Her2/Neu/ErbB2 receptor-negative breast tumors. Oncol Rep. 2010;23(1):35–43.
12. Leontovich AA, Jalalirad M, Salisbury JL, et al. NOTCH3 expression is linked to breast cancer seeding and distant metastasis. Breast Cancer Res. 2018;20(1):105. doi:10.1186/s13058-018-1020-0
13. Mittal S, Subramanyam D, Dey D, Kumar RV, Rangarajan A. Cooperation of Notch and Ras/MAPK signaling pathways in human breast carcinogenesis. Mol Cancer. 2009;8(1):128. doi:10.1186/1476-4598-8-128
14. Wang J-W, Wei X-L, Dou X-W, Huang W-H, Du C-W, Zhang G-J. The association between Notch4 expression, and clinicopathological characteristics and clinical outcomes in patients with breast cancer. Oncol Lett. 2018;15(6):8749–8755. doi:10.3892/ol.2018.8442
15. Kim DC, Jin H, Lee JS, Son E, Lee GW, Kim HJ. P2Y(2)R has a significant correlation with Notch-4 in patients with breast cancer. Oncol Lett. 2020;20(1):647–654.
16. Pece S, Serresi M, Santolini E, et al. Loss of negative regulation by Numb over Notch is relevant to human breast carcinogenesis. J Cell Biol. 2004;167(2):215–221.
17. Rennstam K, McMichael N, Berglund P, et al. Numb protein expression correlates with a basal-like phenotype and cancer stem cell markers in primary breast cancer. Breast Cancer Res Treat. 2010;122(2):315–324.
18. Edwards A, Brennan K. Notch signalling in breast development and cancer. Front Cell Dev Biol. 2021;9:692173.
19. Kumar S, Nandi A, Singh S, et al. Dll1(+) quiescent tumor stem cells drive chemoresistance in breast cancer through NF-κB survival pathway. Nat Commun. 2021;12(1):432.
20. Yuan C, Chang K, Xu C, Li Q, Du Z. High expression of DLL3 is associated with a poor prognosis and immune infiltration in invasive breast cancer patients. Transl Oncol. 2021;14(7):101080.
21. Zohny SF, Zamzami MA, Al-Malki AL, Trabulsi NH. Highly Expressed DLL4 and JAG1: their Role in Incidence of Breast Cancer Metastasis. Arch Med Res. 2020;51(2):145–152.
22. Andrieu G, Tran AH, Strissel KJ, Denis GV. BRD4 regulates breast cancer dissemination through Jagged1/Notch1 signaling. Cancer Res. 2016;76(22):6555–6567.
23. Li CY, Miao KL, Chen Y, et al. Jagged2 promotes cancer stem cell properties of triple negative breast cancer cells and paclitaxel resistance via regulating microRNA-200. Eur Rev Med Pharmacol Sci. 2018;22(18):6008–6014.
24. Shen Q, Reedijk M. Notch signaling and the breast cancer microenvironment. Adv Exp Med Biol. 2021;1287:183–200.
25. Glittenberg M, Pitsouli C, Garvey C, Delidakis C, Bray S. Role of conserved intracellular motifs in Serrate signalling, cis-inhibition and endocytosis. EMBO j. 2006;25(20):4697–4706.
26. Kopan R, Ilagan MX. The canonical Notch signaling pathway: unfolding the activation mechanism. Cell. 2009;137(2):216–233.
27. Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: cell fate control and signal integration in development. Science. 1999;284(5415):770–776.
28. Weinmaster G. Notch signal transduction: a real rip and more. Curr Opin Genet Dev. 2000;10(4):363–369.
29. Ohashi S, Natsuizaka M, Yashiro-Ohtani Y, et al. NOTCH1 and NOTCH3 coordinate esophageal squamous differentiation through a CSL-dependent transcriptional network. Gastroenterology. 2010;139(6):2113–2123.
30. Jarriault S, Brou C, Logeat F, Schroeter EH, Kopan R, Israel A. Signalling downstream of activated mammalian Notch. Nature. 1995;377(6547):355–358.
31. Schroeter EH, Kisslinger JA, Kopan R. Notch-1 signalling requires ligand-induced proteolytic release of intracellular domain. Nature. 1998;393(6683):382–386.
32. Petcherski AG, Kimble J. Mastermind is a putative activator for Notch. Curr Biol. 2000;10(13):R471–473.
33. Nam Y, Sliz P, Song L, Aster JC, Blacklow SC. Structural basis for cooperativity in recruitment of MAML coactivators to Notch transcription complexes. Cell. 2006;124(5):973–983.
34. Fortini ME, Artavanis-Tsakonas S. The suppressor of hairless protein participates in notch receptor signaling. Cell. 1994;79(2):273–282.
35. Struhl G, Adachi A. Nuclear access and action of notch in vivo. Cell. 1998;93(4):649–660.
36. Yu MM, Zhou QM. 3,6-dihydroxyflavone suppresses the epithelial-mesenchymal transition, migration and invasion in endometrial stromal cells by inhibiting the Notch signaling pathway. Eur Rev Med Pharmacol Sci. 2018;22(12):4009–4017.
37. Reedijk M. Notch signaling and breast cancer. Adv Exp Med Biol. 2012;727:241–257.
38. Borggrefe T, Oswald F. The Notch signaling pathway: transcriptional regulation at Notch target genes. Cell Mol Life Sci. 2009;66(10):1631–1646.
39. Folkman J. Tumor angiogenesis: therapeutic implications. N Engl J Med. 1971;285(21):1182–1186.
40. Weis SM, Cheresh DA. Tumor angiogenesis: molecular pathways and therapeutic targets. Nat Med. 2011;17(11):1359–1370.
41. Hanahan D, Folkman J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell. 1996;86(3):353–364.
42. Lawson ND, Scheer N, Pham VN, et al. Notch signaling is required for arterial-venous differentiation during embryonic vascular development. Development. 2001;128(19):3675–3683.
43. Hofmann JJ, Iruela-Arispe ML. Notch signaling in blood vessels: who is talking to whom about what? Circ Res. 2007;100(11):1556–1568.
44. Limbourg FP, Takeshita K, Radtke F, Bronson RT, Chin MT, Liao JK. Essential role of endothelial Notch1 in angiogenesis. Circulation. 2005;111(14):1826–1832.
45. Villa N, Walker L, Lindsell CE, Gasson J, Iruela-Arispe ML, Weinmaster G. Vascular expression of Notch pathway receptors and ligands is restricted to arterial vessels. Mech Dev. 2001;108(1–2):161–164.
46. Hayashi H, Kume T. Foxc transcription factors directly regulate Dll4 and Hey2 expression by interacting with the VEGF-Notch signaling pathways in endothelial cells. PLoS One. 2008;3(6):e2401.
47. Surendran S, Suresh A. Arterialization and anomalous vein wall remodeling in varicose veins is associated with upregulated FoxC2-Dll4 pathway. Lab Invest. 2016;96(4):399–408.
48. Xia S, Menden HL, Korfhagen TR, Kume T, Sampath V. Endothelial immune activation programmes cell-fate decisions and angiogenesis by inducing angiogenesis regulator DLL4 through TLR4-ERK-FOXC2 signalling. J Physiol. 2018;596(8):1397–1417.
49. Li Y, Yang W, Yang Q, Zhou S. Nuclear localization of GLI1 and elevated expression of FOXC2 in breast cancer is associated with the basal-like phenotype. Histol Histopathol. 2012;27(4):475–484.
50. Liu HXJ, Zheng YY, et al. Influence of FOXC2 in angiogenesis of breast cancer MCF-7 cells by DLL4/Notch1 signal pathway. J Jilin Univ. 2014;40(3):488–492.
51. Zhu Q, Li J, Wu Q, et al. Linc-OIP5 in the breast cancer cells regulates angiogenesis of human umbilical vein endothelial cells through YAP1/Notch/NRP1 signaling circuit at a tumor microenvironment. Biol Res. 2020;53(1):5.
52. Hultgren NW, Fang JS, Ziegler ME, et al. Slug regulates the Dll4-Notch-VEGFR2 axis to control endothelial cell activation and angiogenesis. Nat Commun. 2020;11(1):5400.
53. Noguera-Troise I, Daly C, Papadopoulos NJ, et al. Blockade of Dll4 inhibits tumour growth by promoting non-productive angiogenesis. Novartis Found Symp. 2007;283:106–120.
54. Williams CK, Li JL, Murga M, Harris AL, Tosato G. Up-regulation of the Notch ligand Delta-like 4 inhibits VEGF-induced endothelial cell function. Blood. 2006;107(3):931–939.
55. Erkasap N, Ozyurt R, Ozkurt M, et al. Role of Notch, IL-1 and leptin expression in colorectal cancer. Exp Ther Med. 2021;21(6):600.
56. Cao W, Zhao F, Zhao Y, Jing Z. Effect and correlation of VEGF and Notch1 in invasion and metastasis of breast cancer. J Basic Clin Oncol. 2012;25(6):480–483.
57. Patel NS, Li JL, Generali D, Poulsom R, Cranston DW, Harris AL. Up-regulation of delta-like 4 ligand in human tumor vasculature and the role of basal expression in endothelial cell function. Cancer Res. 2005;65(19):8690–8697.
58. Oon CE, Bridges E, Sheldon H, et al. Role of Delta-like 4 in Jagged1-induced tumour angiogenesis and tumour growth. Oncotarget. 2017;8(25):40115–40131.
59. Pedrosa AR, Trindade A, Fernandes AC, et al. Endothelial Jagged1 antagonizes Dll4 regulation of endothelial branching and promotes vascular maturation downstream of Dll4/Notch1. Arterioscler Thromb Vasc Biol. 2015;35(5):1134–1146.
60. Mailhos C, Modlich U, Lewis J, Harris A, Delta4 I-HD. an endothelial specific notch ligand expressed at sites of physiological and tumor angiogenesis. Differentiation. 2001;69(2–3):135–144.
61. Gale NW, Dominguez MG, Noguera I, et al. Haploinsufficiency of delta-like 4 ligand results in embryonic lethality due to major defects in arterial and vascular development. Proc Natl Acad Sci U S A. 2004;101(45):15949–15954.
62. Benedito R, Trindade A, Hirashima M, et al. Loss of Notch signalling induced by Dll4 causes arterial calibre reduction by increasing endothelial cell response to angiogenic stimuli. BMC Dev Biol. 2008;8:117.
63. Benedito R, Roca C, Sörensen I, et al. The notch ligands Dll4 and Jagged1 have opposing effects on angiogenesis. Cell. 2009;137(6):1124–1135.
64. Xue Y, Gao X, Lindsell CE, et al. Embryonic lethality and vascular defects in mice lacking the Notch ligand Jagged1. Hum Mol Genet. 1999;8(5):723–730.
65. Pitulescu ME, Schmidt I, Giaimo BD, et al. Dll4 and Notch signalling couples sprouting angiogenesis and artery formation. Nat Cell Biol. 2017;19(8):915–927.
66. Zhou R, Wang S, Wen H, Wang M, Wu M. The bispecific antibody HB-32, blockade of both VEGF and DLL4 shows potent anti-angiogenic activity in vitro and anti-tumor activity in breast cancer xenograft models. Exp Cell Res. 2019;380(2):141–148.
67. Wang S, Zhou R, Sun F, Li R, Wang M, Wu M. The two novel DLL4-targeting antibody-drug conjugates MvM03 and MGD03 show potent anti-tumour activity in breast cancer xenograft models. Cancer Lett. 2017;409:125–136.
68. High FA, Lu MM, Pear WS, Loomes KM, Kaestner KH, Epstein JA. Endothelial expression of the Notch ligand Jagged1 is required for vascular smooth muscle development. Proc Natl Acad Sci U S A. 2008;105(6):1955–1959.
69. Zeng Q, Li S, Chepeha DB, et al. Crosstalk between tumor and endothelial cells promotes tumor angiogenesis by MAPK activation of Notch signaling. Cancer Cell. 2005;8(1):13–23.
70. Soares R, Balogh G, Guo S, Gärtner F, Russo J, Schmitt F. Evidence for the notch signaling pathway on the role of estrogen in angiogenesis. Mol Endocrinol. 2004;18(9):2333–2343.
71. Sainson RC, Johnston DA, Chu HC, et al. TNF primes endothelial cells for angiogenic sprouting by inducing a tip cell phenotype. Blood. 2008;111(10):4997–5007.
72. Azizi E, Carr AJ, Plitas G, et al. Single-cell map of diverse immune phenotypes in the breast tumor microenvironment. Cell. 2018;174(5):1293–1308.e1236.
73. Mantovani A, Marchesi F, Malesci A, Laghi L, Allavena P. Tumour-associated macrophages as treatment targets in oncology. Nat Rev Clin Oncol. 2017;14(7):399–416.
74. Mahmoud SM, Lee AH, Paish EC, Macmillan RD, Ellis IO, Green AR. Tumour-infiltrating macrophages and clinical outcome in breast cancer. J Clin Pathol. 2012;65(2):159–163.
75. Kolb R, Liu GH, Janowski AM, Sutterwala FS, Zhang W. Inflammasomes in cancer: a double-edged sword. Protein Cell. 2014;5(1):12–20.
76. Qian BZ, Li J, Zhang H, et al. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature. 2011;475(7355):222–225.
77. Valković T, Lucin K, Krstulja M, Dobi-Babić R, Jonjić N. Expression of monocyte chemotactic protein-1 in human invasive ductal breast cancer. Pathol Res Pract. 1998;194(5):335–340.
78. Shen Q, Cohen B, Zheng W, et al. Notch Shapes the Innate Immunophenotype in Breast Cancer. Cancer Discov. 2017;7(11):1320–1335.
79. Lyu X. Innate and adaptive immune cells in tumor microenvironment. Gulf J Oncolog. 2021;1(35):77–81.
80. Coussens LM, Zitvogel L, Palucka AK. Neutralizing tumor-promoting chronic inflammation: a magic bullet? Science. 2013;339(6117):286–291.
81. Lin S, Sun L, Lyu X, et al. Lactate-activated macrophages induced aerobic glycolysis and epithelial-mesenchymal transition in breast cancer by regulation of CCL5-CCR5 axis: a positive metabolic feedback loop. Oncotarget. 2017;8(66):110426–110443.
82. Ahirwar DK, Charan M, Mishra S, et al. Slit2 inhibits breast cancer metastasis by activating M1-like phagocytic and antifibrotic macrophages. Cancer Res. 2021;81(20):5255–5267.
83. Zhao W, Hu X, Li W, et al. M2-like TAMs function reversal contributes to breast cancer eradication by combination dual immune checkpoint blockade and photothermal therapy. Small. 2021;17(13):e2007051.
84. Wang YC, He F, Feng F, et al. Notch signaling determines the M1 versus M2 polarization of macrophages in antitumor immune responses. Cancer Res. 2010;70(12):4840–4849.
85. Yin D, Wang W, Han W, Targeting Notch-activated FC. M1 macrophages attenuate lung tissue damage in a rat model of ventilator induced lung injury. Int J Mol Med. 2019;44(4):1388–1398.
86. Fukuda D, Aikawa E, Swirski FK, et al. Notch ligand delta-like 4 blockade attenuates atherosclerosis and metabolic disorders. Proc Natl Acad Sci U S A. 2012;109(27):E1868–1877.
87. Pagie S, Gérard N, Charreau B. Notch signaling triggered via the ligand DLL4 impedes M2 macrophage differentiation and promotes their apoptosis. Cell Commun Signal. 2018;16(1):4.
88. Singla DK, Wang J, Singla R. Primary human monocytes differentiate into M2 macrophages and involve Notch-1 pathway. Can J Physiol Pharmacol. 2017;95(3):288–294.
89. Tao S, Chen Q, Lin C, Dong H. Linc00514 promotes breast cancer metastasis and M2 polarization of tumor-associated macrophages via Jagged1-mediated notch signaling pathway. J Exp Clin Cancer Res. 2020;39(1):191.
90. Wu SY, Fu T, Jiang YZ, Shao ZM. Natural killer cells in cancer biology and therapy. Mol Cancer. 2020;19(1):120.
91. Rezaeifard S, Talei A, Shariat M, Erfani N. Tumor infiltrating NK cell (TINK) subsets and functional molecules in patients with breast cancer. Mol Immunol. 2021;136:161–167.
92. Chaves P, Zriwil A, Wittmann L, et al. Loss of canonical Notch signaling affects multiple steps in NK cell development in mice. J Immunol. 2018;201(11):3307–3319.
93. Beck RC, Padival M, Yeh D, Ralston J, Cooke KR, Lowe JB. The Notch ligands Jagged2, Delta1, and Delta4 induce differentiation and expansion of functional human NK cells from CD34+ cord blood hematopoietic progenitor cells. Biol Blood Marrow Transplant. 2009;15(9):1026–1037.
94. Nalin AP, Kowalski JJ, Sprague AC, et al. Notch regulates innate lymphoid cell plasticity during human NK cell development. J Immunol. 2020;205(10):2679–2693.
95. Wang K, Li L, Fu L, et al. Integrated bioinformatics analysis the function of RNA binding proteins (RBPs) and their prognostic value in breast cancer. Front Pharmacol. 2019;10:140.
96. Tao Z, Suo H, Zhang L, et al. MRPL13 is a prognostic cancer biomarker and correlates with immune infiltrates in breast cancer. Onco Targets Ther. 2020;13:12255–12268.
97. Manaster I, Gazit R, Goldman-Wohl D, et al. Notch activation enhances IFNgamma secretion by human peripheral blood and decidual NK cells. J Reprod Immunol. 2010;84(1):1–7.
98. Wenbo R, Jing H, Liancheng L. Progress in the study of the relationship between IFN-γ and the development of breast cancer. Shandong Med J. 2017;57(13):101–103.
99. Brandstadter JD, Maillard I. Notch signalling in T cell homeostasis and differentiation. Open Biol. 2019;9(11):190187.
100. Van de Walle I, Dolens AC, Durinck K, et al. GATA3 induces human T-cell commitment by restraining Notch activity and repressing NK-cell fate. Nat Commun. 2016;7:11171. doi:10.1038/ncomms11171
101. Hozumi K, Negishi N, Tsuchiya I, et al. Notch signaling is necessary for GATA3 function in the initiation of T cell development. Eur J Immunol. 2008;38(4):977–985. doi:10.1002/eji.200737688
102. Li X, Cao Y, Jin F. HES1 expression and its correlation with prognosis in breast cancer. J Modern Oncol. 2018;26(06):0872–0875.
103. De Obaldia ME, Bell JJ, Wang X, et al. T cell development requires constraint of the myeloid regulator C/EBP-α by the Notch target and transcriptional repressor Hes1. Nat Immunol. 2013;14(12):1277–1284. doi:10.1038/ni.2760
104. De Decker M, Lavaert M, Roels J, et al. HES1 and HES4 have non-redundant roles downstream of Notch during early human T-cell development. Haematologica. 2021;106(1):130–141. doi:10.3324/haematol.2019.226126
105. Rutz S, Mordmüller B, Sakano S, Scheffold A. Notch ligands Delta-like1, Delta-like4 and Jagged1 differentially regulate activation of peripheral T helper cells. Eur J Immunol. 2005;35(8):2443–2451. doi:10.1002/eji.200526294
106. Niu XL, Mao LQ, Ye L, Wang Y. Effects of IFN-γ and IL-4 on the growth and the estrogen receptor subtypes of breast cancer cell MCF-7. Chin J Cancer Prevention Treatment. 2011;18(20):1581–1584.
107. Saleh ME, Gadalla R, Hassan H, et al. The immunomodulatory role of tumor Syndecan-1 (CD138) on ex vivo tumor microenvironmental CD4+ T cell polarization in inflammatory and non-inflammatory breast cancer patients. PLoS One. 2019;14(5):e0217550. doi:10.1371/journal.pone.0217550
108. Ruffell B, Au A, Rugo HS, Esserman LJ, Hwang ES, Coussens LM. Leukocyte composition of human breast cancer. Proc Natl Acad Sci U S A. 2012;109(8):2796–2801. doi:10.1073/pnas.1104303108
109. Mathieu M, Duval F, Daudelin JF, Labrecque N. The Notch signaling pathway controls short-lived effector CD8+ T cell differentiation but is dispensable for memory generation. J Immunol. 2015;194(12):5654–5662. doi:10.4049/jimmunol.1402837
110. De Sousa DM, Duval F, Daudelin JF, Boulet S, Labrecque N. The Notch signaling pathway controls CD8+ T cell differentiation independently of the classical effector HES1. PLoS One. 2019;14(4):e0215012. doi:10.1371/journal.pone.0215012
111. Oshi M, Asaoka M, Tokumaru Y, et al. CD8 T cell score as a prognostic biomarker for triple negative breast cancer. Int J Mol Sci. 2020;21:18.
112. Elwakeel E, Weigert A. Breast cancer CAFs: spectrum of phenotypes and promising targeting avenues. Int J Mol Sci. 2021;22(21):21. doi:10.3390/ijms222111636
113. Shao H, Moller M, Cai L, et al. Converting melanoma-associated fibroblasts into a tumor-suppressive phenotype by increasing intracellular Notch1 pathway activity. PLoS One. 2021;16(3):e0248260. doi:10.1371/journal.pone.0248260
114. Du Y, Shao H, Moller M, Prokupets R, Tse YT, Liu ZJ. Intracellular Notch1 signaling in cancer-associated fibroblasts dictates the plasticity and stemness of melanoma stem/initiating cells. Stem Cells. 2019;37(7):865–875. doi:10.1002/stem.3013
115. Pupo M, Pisano A, Abonante S, Maggiolini M, Musti AM. GPER activates Notch signaling in breast cancer cells and cancer-associated fibroblasts (CAFs). Int J Biochem Cell Biol. 2014;46:56–67. doi:10.1016/j.biocel.2013.11.011
116. Pelon F, Bourachot B, Kieffer Y, et al. Cancer-associated fibroblast heterogeneity in axillary lymph nodes drives metastases in breast cancer through complementary mechanisms. Nat Commun. 2020;11(1):404. doi:10.1038/s41467-019-14134-w
117. Simon S, Labarriere NPD-1. Expression on tumor-specific T cells: friend or foe for immunotherapy? Oncoimmunology. 2017;7(1):e1364828. doi:10.1080/2162402X.2017.1364828
118. Zak KM, Grudnik P, Magiera K, Dömling A, Dubin G, Holak TA. Structural biology of the immune checkpoint receptor PD-1 and its ligands PD-L1/PD-L2. Structure. 2017;25(8):1163–1174. doi:10.1016/j.str.2017.06.011
119. Patsoukis N, Wang Q, Strauss L, Boussiotis VA. Revisiting the PD-1 pathway. Sci Adv. 2020;6(38):38. doi:10.1126/sciadv.abd2712
120. Han Y, Liu D, Li L. PD-1/PD-L1 pathway: current researches in cancer. Am J Cancer Res. 2020;10(3):727–742.
121. Ai L, Xu A, Xu J. Roles of PD-1/PD-L1 pathway: signaling, cancer, and beyond. Adv Exp Med Biol. 2020;1248:33–59.
122. Jiang X, Wang J, Deng X, et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol Cancer. 2019;18(1):10. doi:10.1186/s12943-018-0928-4
123. Adams S, Gatti-Mays ME, Kalinsky K, et al. Current landscape of immunotherapy in breast cancer: a review. JAMA Oncol. 2019;5(8):1205–1214. doi:10.1001/jamaoncol.2018.7147
124. Polk A, Svane I-M, Andersson M, Nielsen D. Checkpoint inhibitors in breast cancer – current status. Cancer Treat Rev. 2018;63:122–134. doi:10.1016/j.ctrv.2017.12.008
125. Vranic S, Cyprian FS, Gatalica Z, Palazzo J. PD-L1 status in breast cancer: current view and perspectives. Semin Cancer Biol. 2021;72:146–154. doi:10.1016/j.semcancer.2019.12.003
126. Muenst S, Soysal SD, Gao F, Obermann EC, Oertli D, Gillanders WE. The presence of programmed death 1 (PD-1)-positive tumor-infiltrating lymphocytes is associated with poor prognosis in human breast cancer. Breast Cancer Res Treat. 2013;139(3):667–676. doi:10.1007/s10549-013-2581-3
127. Mittendorf EA, Philips AV, Meric-Bernstam F, et al. PD-L1 expression in triple-negative breast cancer. Cancer Immunol Res. 2014;2(4):361–370. doi:10.1158/2326-6066.CIR-13-0127
128. Huang W, Ran R, Shao B, Li H. Prognostic and clinicopathological value of PD-L1 expression in primary breast cancer: a meta-analysis. Breast Cancer Res Treat. 2019;178(1):17–33. doi:10.1007/s10549-019-05371-0
129. Chen W, Liu J, Ge F, et al. Long noncoding RNA HOTAIRM1 promotes immunosuppression in sepsis by inducing T cell exhaustion. J Immunol. 2022;208(3):618–632. doi:10.4049/jimmunol.2100709
130. Pan T, Liu Z, Yin J, Zhou T, Liu J, Qu H. Notch signaling pathway was involved in regulating programmed cell death 1 expression during sepsis-induced immunosuppression. Mediators Inflamm. 2015;2015:539841. doi:10.1155/2015/539841
131. Meng J, Jiang Y-Z, Zhao S, et al. Tumor-derived Jagged1 promotes cancer progression through immune evasion. Cell Rep. 2022;38(10):110492. doi:10.1016/j.celrep.2022.110492
132. Mansour FA, Al-Mazrou A, Al-Mohanna F, Al-Alwan M, Ghebeh H. PD-L1 is overexpressed on breast cancer stem cells through notch3/mTOR axis. Oncoimmunology. 2020;9(1):1729299. doi:10.1080/2162402X.2020.1729299
133. Savage PA, Klawon DEJ, Miller CH. Regulatory T cell development. Annu Rev Immunol. 2020;38(1):421–453. doi:10.1146/annurev-immunol-100219-020937
134. Yang Z, Qi Y, Lai N, et al. Notch1 signaling in melanoma cells promoted tumor-induced immunosuppression via upregulation of TGF-β1. J Exp Clin Cancer Res. 2018;37(1):1. doi:10.1186/s13046-017-0664-4
135. Ohkura N, Hamaguchi M, Morikawa H, et al. T cell receptor stimulation-induced epigenetic changes and Foxp3 expression are independent and complementary events required for Treg cell development. Immunity. 2012;37(5):785–799. doi:10.1016/j.immuni.2012.09.010
136. Anastasi E, Campese AF, Bellavia D, et al. Expression of activated Notch3 in transgenic mice enhances generation of T regulatory cells and protects against experimental autoimmune diabetes. J Immunol. 2003;171(9):4504–4511. doi:10.4049/jimmunol.171.9.4504
137. Jiao W-E, Wei J-F, Kong Y-G, Xu Y, Tao -Z-Z, Chen S-M. Notch signaling promotes development of allergic rhinitis by suppressing Foxp3 expression and Treg cell differentiation. Int Arch Allergy Immunol. 2019;178(1):33–44. doi:10.1159/000493328
138. Knochelmann HM, Dwyer CJ, Bailey SR, et al. When worlds collide: th17 and Treg cells in cancer and autoimmunity. Cell Mol Immunol. 2018;15(5):458–469. doi:10.1038/s41423-018-0004-4
139. He Z, Wu J, Zeng X, Bao H, Liu X. Role of the Notch ligands Jagged1 and Delta4 in Th17/Treg immune imbalance in a mouse model of chronic asthma. Exp Lung Res. 2021;47(6):289–299. doi:10.1080/01902148.2021.1933653
140. Wesseling-Rozendaal Y, van Doorn A, Willard-Gallo K, van de Stolpe A. Characterization of immunoactive and immunotolerant CD4+ T cells in breast cancer by measuring activity of signaling pathways that determine immune cell function. Cancers. 2022;14(3):490. doi:10.3390/cancers14030490
141. Kaushik B, Pal D, Saha S. Gamma secretase inhibitor: therapeutic target via NOTCH signaling in T cell acute lymphoblastic leukemia. Curr Drug Targets. 2021;22(15):1789–1798. doi:10.2174/1389450122666210203192752
142. Kumari M, Krishnamurthy PT, Pinduprolu S, Sola PDR-5. DR-5 and DLL-4 mAb functionalized SLNs of gamma-secretase inhibitors – an approach for TNBC treatment. Adv Pharm Bull. 2020;11(4):618–623. doi:10.34172/apb.2021.070
143. Chen X, Chen X, Zhou Z, et al. LY411575, a potent γ-secretase inhibitor, suppresses osteoclastogenesis in vitro and LPS-induced calvarial osteolysis in vivo. J Cell Physiol. 2019;234(11):20944–20956. doi:10.1002/jcp.28699
144. Means-Powell JA, Mayer IA, Ismail-Khan R, et al. A phase Ib dose escalation trial of RO4929097 (a γ-secretase inhibitor) in combination with exemestane in patients with ER +metastatic breast cancer (MBC). Clin Breast Cancer. 2022;22(2):103–114. doi:10.1016/j.clbc.2021.10.013
145. Massard C, Azaro A, Soria J-C, et al. First-in-human study of LY3039478, an oral Notch signaling inhibitor in advanced or metastatic cancer. Ann Oncol. 2018;29(9):1911–1917. doi:10.1093/annonc/mdy244
146. Wang D, Xu J, Liu B, et al. IL6 blockade potentiates the anti-tumor effects of γ-secretase inhibitors in Notch3-expressing breast cancer. Cell Death Differ. 2018;25(2):330–339. doi:10.1038/cdd.2017.162
147. Hossain F, Sorrentino C, Ucar DA, et al. Notch signaling regulates mitochondrial metabolism and NF-κB activity in triple-negative breast cancer cells via IKKα-dependent non-canonical pathways. Front Oncol. 2018;8:575.
148. Paroni G, Zanetti A, Barzago MM, et al. Retinoic acid sensitivity of triple-negative breast cancer cells characterized by constitutive activation of the Notch1 pathway: the role of Rarβ. Cancers. 2020;12:10.
149. Das A, Narayanam MK, Paul S, et al. A novel triazole, NMK-T-057, induces autophagic cell death in breast cancer cells by inhibiting γ-secretase-mediated activation of Notch signaling. J Biol Chem. 2019;294(17):6733–6750.
150. Smith DC, Eisenberg PD, Manikhas G, et al. A Phase I dose escalation and expansion study of the anticancer stem cell agent demcizumab (anti-DLL4) in patients with previously treated solid tumors. Clin Cancer Res. 2014;20(24):6295–6303.
151. Silva G, Sales-Dias J, Casal D, et al. Development of Dl1.72, a novel anti-DLL1 antibody with anti-tumor efficacy against estrogen receptor-positive breast cancer. Cancers. 2021;13(16):48.
152. Yen WC, Fischer MM, Axelrod F, et al. Targeting Notch signaling with a Notch2/Notch3 antagonist (tarextumab) inhibits tumor growth and decreases tumor-initiating cell frequency. Clin Cancer Res. 2015;21(9):2084–2095.
153. Jia H, Liu M, Wang X, et al. Cimigenoside functions as a novel γ-secretase inhibitor and inhibits the proliferation or metastasis of human breast cancer cells by γ-secretase/Notch axis. Pharmacol Res. 2021;169:105686.
154. Zhang J, Yu K, Han X, et al. Paeoniflorin influences breast cancer cell proliferation and invasion via inhibition of the Notch‑1 signaling pathway. Mol Med Rep. 2018;17(1):1321–1325.
155. Bawadood AS, Al-Abbasi FA, Anwar F, El-Halawany AM, Al-Abd AM. 6-Shogaol suppresses the growth of breast cancer cells by inducing apoptosis and suppressing autophagy via targeting notch signaling pathway. Biomed Pharmacother. 2020;128:110302.
156. Kushwaha PP, Singh AK, Shuaib M, et al. 3-O-(E)-p-Coumaroyl betulinic acid possess anticancer activity and inhibit Notch signaling pathway in breast cancer cells and mammosphere. Chem Biol Interact. 2020;328:109200.
157. Xia S, Zhang X, Li C, Guan H. Oridonin inhibits breast cancer growth and metastasis through blocking the Notch signaling. Saudi Pharm J. 2017;25(4):638–643.
158. Dong J, Yang W, Han J, Cheng R, Li L. Effects of Notch signaling components from breast cancer cells treated in culture with resveratrol. Res Vet Sci. 2020;132:369–378.
159. Ramamoorthy P, Dandawate P, Jensen RA, Anant S. Celastrol and triptolide suppress stemness in triple negative breast cancer: Notch as a therapeutic target for stem cells. Biomedicines. 2021;9(5):164.
160. Nasser F, Moussa N, Helmy MW, Haroun M. Dual targeting of Notch and Wnt/β-catenin pathways: potential approach in triple-negative breast cancer treatment. Naunyn Schmiedebergs Arch Pharmacol. 2021;394(3):481–490.
161. Bai JW, Wei M, Li JW, Zhang GJ. Notch signaling pathway and endocrine resistance in breast cancer. Front Pharmacol. 2020;11:924.
162. Malik N, Yan H, Moshkovich N, et al. The transcription factor CBFB suppresses breast cancer through orchestrating translation and transcription. Nat Commun. 2019;10(1):2071.
163. Lin Q, Chen X, Meng F, et al. ASPH-Notch axis guided exosomal delivery of prometastatic secretome renders breast cancer multi-organ metastasis. Mol Cancer. 2019;18(1):156.
164. Jeong GY, Park MK, Choi HJ, et al. NSD3-induced methylation of H3K36 activates NOTCH signaling to drive breast tumor initiation and metastatic progression. Cancer Res. 2021;81(1):77–90.
165. Liu X, Xian Y, Xu H, et al. The associations between Deltex1 and clinical characteristics of breast cancer. Gland Surg. 2021;10(11):3116–3127.
166. Jaiswal A, Murakami K, Elia A, et al. Therapeutic inhibition of USP9x-mediated Notch signaling in triple-negative breast cancer. Proc Natl Acad Sci U S A. 2021;118(38):45.
167. Zhang HS, Zhang ZG, Du GY, et al. Nrf2 promotes breast cancer cell migration via up-regulation of G6PD/HIF-1α/Notch1 axis. J Cell Mol Med. 2019;23(5):3451–3463.
168. Gao P, Peng T, Lin S, et al. Key role of MCUR1 in malignant progression of breast cancer. Onco Targets Ther. 2021;14:4163–4175.
169. Fournier P, Viallard C, Dejda A, Sapieha P, Larrivée B, Royal I. The protein tyrosine phosphatase PTPRJ/DEP-1 contributes to the regulation of the Notch-signaling pathway and sprouting angiogenesis. Angiogenesis. 2020;23(2):145–157.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Progress of Platelets in Breast Cancer
Wang L, Zhang K, Feng J, Wang D, Liu J
Cancer Management and Research 2023, 15:811-821
Published Date: 11 August 2023
Anlotinib or Bevacizumab Combined with Taxane/Capecitabine for the Second-Line or Subsequent Treatment of HER-2 Negative Metastatic Breast Cancer: A Retrospective Cohort Study
Xiang J, Sun P, Liu Z, Zhang Q, Song L, Liu J
Drug Design, Development and Therapy 2025, 19:10871-10883
Published Date: 8 December 2025