")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 15
The neuroprotective mechanism of 2-arachidonoylglycerol 2-AG against non-caspase-dependent apoptosis in mice hippocampal neurons following MCAO
Authors Zhong HY, Yang Z, Qiu Z, Lei SQ, Xia ZY
Received 9 March 2019
Accepted for publication 23 July 2019
Published 23 August 2019 Volume 2019:15 Pages 2417—2424
DOI https://doi.org/10.2147/NDT.S208094
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yuping Ning
He-Ying Zhong, Zhou Yang, Zhen Qiu, Shao-Qing Lei, Zhong-Yuan Xia
Department of Anesthesiology, Renmin Hospital of Wuhan University, Wuhan, Hubei Province, People’s Republic of China
Correspondence: Zhong-Yuan Xia
Department of Anesthesiology, Renmin Hospital of Wuhan University, No. 238 Jiefang Road, Wuhan 430060, Hubei Province, People’s Republic of China
Tel +86 1 380 862 8560
Email [email protected]
Objective: In this study, the neuroprotective mechanism of 2-arachidonoylglycerol 2-AG against non-caspase-dependent apoptosis in mice hippocampal neurons following MCAO was investigated.
Method: One hundred and fifty healthy clean male C57BL/6 mice were randomly divided into 3 groups: sham group, model group and 2-AG treatment group, 50 mice in each group. A modified Zea Longa method was used to establish a model of middle cerebral artery occlusion (MCAO) in mice. The apoptosis rate and mitochondrial membrane potential of hippocampal nerve cells were measured by flow cytometry. The mRNA expressions of AIF, Endo G and BNIP3 in hippocampal tissues were determined by qPCR. Western blot was used to determine the protein expressions of AIF, Endo G and BNIP3 in the mitochondria of hippocampal tissue.
Results: The apoptosis rate of hippocampal neurons in the group treated with 2-AG was significantly lower than that of the model (P<0.01), which indicated that 2-AG could inhibit the apoptosis of hippocampal neurons induced by MCAO. However, the mitochondrial membrane potential of hippocampal neurons in the group treated with 2-AG was significantly higher than that of the model (P<0.01), indicating that 2-AG could improve the mitochondrial membrane potential of hippocampal neurons in MCAO mice. Real-time quantitative PCR (qPCR) showed that 2-AG could inhibit the gene expressions of AIF, Endo G and BNIP3 in hippocampal tissues. Western blot results showed that 2-AG could inhibit the secretions of AIF, Endo G and BNIP3 into cytoplasm in mitochondria.
Conclusion: Endocannabinoids 2-AG had a protective effect on neurons injury, and the mechanism was possibly associated with the protection of the brain nerve cells in the hippocampus and the integrity of the mitochondrial function. Endocannabinoids 2-AG may inhibit the non-caspase-dependent apoptosis pathway, so as to exert its nerve protective effect.
Keywords: 2-arachidonoylglycerol, hippocampus, nerve cell, non-caspase-dependent pathway
Introduction
Acute cerebral ischemia is a common disease in clinical, which is seriously harmful to people’s health because of its high incidence, high disability rate and high death rate. The main pathogenesis of ischemic stroke is apoptosis of nerve cells.1 Studies have found that cerebral ischemia can cause significant apoptosis of nerve cells; cerebral ischemia was closely related to apoptosis of nerve cells.2 Mitochondria is the regulatory center of eukaryotic cell life activities, which is not only the site of cell function, but also involve in the regulation of cell apoptosis.3 Therefore, in the treatment of acute cerebral ischemia, the integrity of mitochondrial physiological function plays an important role in apoptosis.
Some experiments showed that caspase inhibitor had a dose-dependent effect on bnip3-induced apoptosis. There are three main apoptosis pathways, namely exogenous death receptor pathway, endogenous mitochondria and endoplasmic reticulum pathway.4 The endogenous mitochondrial pathways are mainly composed of caspase-dependent and non-caspase-dependent apoptosis pathways. The caspase-dependent apoptosis pathway is involved by the Bcl family and Caspase family. Non-caspase-dependent apoptosis pathway is mediated by apoptosis-inducing factor (AIF).5 Currently, non-caspase-dependent apoptosis pathway plays a major role in acute cerebral ischemia. It is commonly seen in apoptosis of nerve cells caused by ischemia and is the main regulatory mechanism of delayed neuron apoptosis.6 Studies have shown that AIF, Endo G and BNIP3 are the proteins which associate with apoptosis in non-caspase-dependent cells.7 Among them, AIF is the first non-caspase-dependent apoptotic molecule discovered, and also an important executive molecule. Studies have found that, when cells are activated by apoptosis factors, mitochondria released AIF and endonuclease G (Endo G) into the cytoplasm. Eventually, it is transposed into the nucleus, destroying DNA and eventually leading to apoptosis.
Endogenous cannabinoids are structural functional analogs of cannabinoids, which are synthesized by the body’s cells. They are involved in the physiological functions of neuron growth and differentiation, immune regulation, synaptic delivery and vasodilatation.8 At present, the research has found that two types of endocannabinoids representative compounds are 2-arachidonic acid glycerol (2-AG) and N-arachidonic acid ethanolamine.9 Endogenous cannabinoids can be synthesized by astrocytes, macrophages, neurons, endothelial cells and microglia cells and secreted out of the cells. It regulates the immunity, nerves and cardiovascular functions; in addition, it has anti-inflammatory and analgesic functions.10 The endocannabinoid system helps fine-tune the most important physiological functions. The endocannabinoid system promotes homeostasis, affecting sleep, appetite, pain, inflammation, memory, mood and even reproduction. Therefore, the endocannabinoid system helps the body balance all major body systems, ensuring that all systems work together.11,12
In this study, a model of middle cerebral artery occlusion (MCAO) was established to explore the protective mechanism of endogenous cannabinoids 2-AG on apoptosis of mouse neurons caused by non-caspase-dependent apoptosis pathway, which provided the experimental basis and theoretical basis for the application of endogenous cannabinoids 2-AG in the treatment of cerebral ischemia.
Materials and methods
Experimental materials and reagents
The study protocol was approved by the Ethics Committee of Renmin Hospital of Wuhan University. Procedures involving animals and their care were conducted in conformity with NIH guidelines (NIH Pub. No. 85-23, revised 1996) and was approved by Animal Care and Use Committee of Renmin Hospital of Wuhan University. Experimental animals: 30 healthy SPF male C57BL/6 mouse, weighing 20–25 g, were purchased from Beijing vitonghua laboratory animal technology co., Ltd. Growth temperature was 23.0±2.0°C, environment humidity: 45–50%, illumination time: 12 hrs a day; free food, water, adaptive feed for a week was provided.
Primary reagents: Endogenous cannabinoids 2-AG injection was purchased from Jiangsu Rui Heng pharmaceutical co., Ltd. 2-AG was purchased from Shanghai Wei Huan Biotech Co., Ltd. AV/PI cell apoptosis dual infection kit was purchased from BD company. The JC-1 probe was purchased from Sigma in the United States. BCA protein concentration determination kit and ECL luminescence kit were purchased from Biyuntian Institute of Biotechnology. Mitochondrial protein extraction kit was purchased from Beijing Kangwei Biotechnology co., Ltd. AIF antibody, Endo G antibody, BNIP3 antibody and trader-actin antibody were all purchased from ABcom company; others are domestic analytical pure reagents.
Methods
Experimental group and administration
One hundred and fifty healthy SPF male C57BL/6 mouse were randomly divided into 3 groups: Sham group, MCAO group and 2-AG + MCAO group, 50 in each group. A modified Zea Longa method was used to establish a model of MCAO in mice. 2-AG + MCAO group: intraperitoneal injection of 5 mg/kg endogenous cannabinoids 2-AG before the establishment of the MCAO model. The sham group and MCAO group were given the same amount of saline intraperitoneally. The main feature of Zea Longa method is that common caroti artery (CCA) is not ligated, and the bolting line is inserted from extemal carotid artery (ECA).
Establishment of the MCAO model in mice
The MCAO model was established by modified Zea Longa method. After intraperitoneal injection of 10% chloral hydrate at 300 mg/kg, the mice were immobilized in supine position on a constant temperature electric heating plate at 37°C. The right common carotid artery, external carotid artery and internal carotid artery were separated through the median carotid incision, and the proximal end of the common carotid artery was ligated. Then the distal end of the common carotid artery and the external carotid artery were clamped with non-invasive blood vessels. Under the bifurcation of the common carotid artery, the incision was made, and the nylon thread passivated by the head was placed. The nylon thread was pushed forward slowly for 8–10 mm. When the resistance was felt, it stopped and fixed to the common carotid artery. After embolization for 1 hr, the thread plug was pulled out to restore blood flow. Two hours after reperfusion, the neurological function score was used to verify the success of the model. The sham-operated group was the same except that no thread bolt was inserted.
Determination of the apoptosis rate of hippocampal neurons by flow cytometry
After 6 hrs of modeling, the mice in each group were immediately executed with a severed head. Under stereomicroscope, the hippocampus tissue is carefully dissected (meninges and vascular membranes are carefully removed) and washed twice with Hank’s buffer. The hippocampal tissues were cut into small pieces of 1 mm3 and use of trypsin digestion. 1×105 cells were taken, centrifuged 1000 r/min−1 for 5 mins, and precipitate was washed by PBS for 2 times. The cells were stained with AV/PI cell apoptosis double infection kit. The apoptosis rates of hippocampal neurons were determined by flow cytometry.
Determination of the mitochondrial membrane potential of hippocampal neurons by flow cytometry
The hippocampal nerve monocyte suspension was prepared. 1×105 cells were taken, centrifuged 1000 r/min−1 for 5 mins, and precipitate was washed by PBS for 2 times. A 0.5 mL JC-1 dye solution was added and mixed well. The cells were treated at 37°C incubator incubation for 15 mins. After the incubation, cells were centrifuged 600×g for 5 mins. Cells were re-suspended with 500 μL PBS buffer. The changes of mitochondrial membrane potential in hippocampal neuron cells were measured by flow cytometry.
Determination of the gene expression in hippocampal tissue of mice in each group by qPCR
Total RNA in cells was extracted by Trizol extraction, and total RNA was transcribed according to the instructions of Micro RNA reverse transcription kit. The reverse transcriptase system was 20 μL. 1 μL total RNA, 4 μL 5×Reaction Mix, 2 μL 10×Super Script Enzyme Mix were added to the double steaming water and filled the system to 20 μL, mixed centrifugation. Reaction conditions were as follows: 37°C for 60 mins, 95°C for 5 mins, saved at 4°C. According to the instructions of q-PCR kit (MSK, Wuhan, China), the reaction system was 20 μL. The SYBR Green dye method was used for determination, and 4 compound holes were set for each sample. Amplification was performed according to the reaction conditions of the kit instructions. The relative expressions of the genes were calculated by the 2-△△CT method.
Determination of the protein expressions in the mitochondria of hippocampal tissue of mice in each group by Western blot
Under sterile conditions, hippocampal tissues of mice in each group were taken. Mitochondrial protein was extracted from the hippocampal tissue of mice using the mitochondrial protein extraction kit (Beibo, Shanghai, China). Western blot was used to detect the protein expression levels of AIF, Endo G and BNIP3 in hippocampal mitochondria. The BCA protein concentration kit was used for protein quantification (TransGen Biotech, Beijing, China). After electrophoresis, the protein was transferred to the PVDF membrane. After using 10% of the dried skim milk solution for close 2 hrs, primary antibodies (1:1000) was used to incubate for the night at 4°C. The membrane was washed with TBST for three times, second antibody (1:4000) was incubated at room temperature for 2 hrs and 3 times with TBST. The protein was colored by chemiluminescence.
Statistical treatment
The experimental results were expressed as mean value ± standard deviation (± s), and SPSS 19.0 statistical software was used for data analysis. Multiple comparisons between the groups were performed using S-N-K method. P<0.05 was a statistically significant difference.
Results
Effects of 2-arachidonoylglycerol 2-AG on apoptosis rate of hippocampal neurons in mice
As shown in Table 1, compared with the sham group, the apoptosis rate of hippocampal neurons cells in the MCAO group was increased significantly (P<0.01). The apoptosis rate of hippocampal neurons cells of mice treated with 2-AG was significantly lower than that of the MCAO group (P<0.01). This indicated that 2-AG could effectively inhibit the apoptosis of hippocampal nerve cells induced by MCAO.
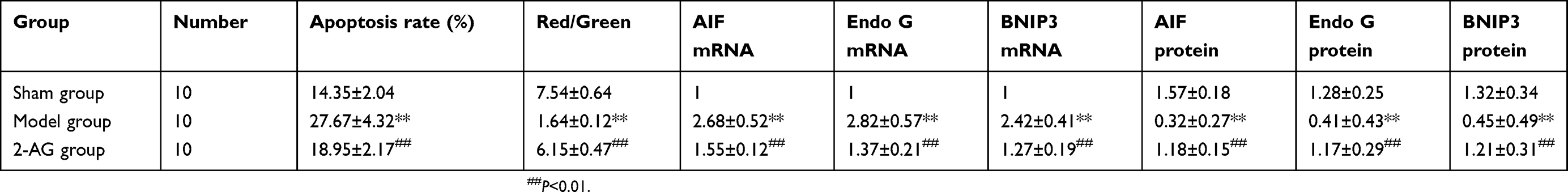 |
Table 1 Effects of 2-AG on apoptosis rate of hippocampal neurons, the expression of AIF, Endo G and BNIP3 in mice (± s) |
Effects of 2-arachidonoylglycerol 2-AG on mitochondrial membrane potential of hippocampal neurons in mice
As shown in Table 2, compared with the sham group, the mitochondrial membrane potential of hippocampal neuron cells in the MCAO group was significantly decreased (P<0.01). The mitochondrial membrane potential of hippocampal neurons was significantly higher in the 2-AG group than in the MCAO group (P<0.01), which indicated that 2-AG could improve the mitochondrial membrane potential of hippocampal neurons in MCAO mice.
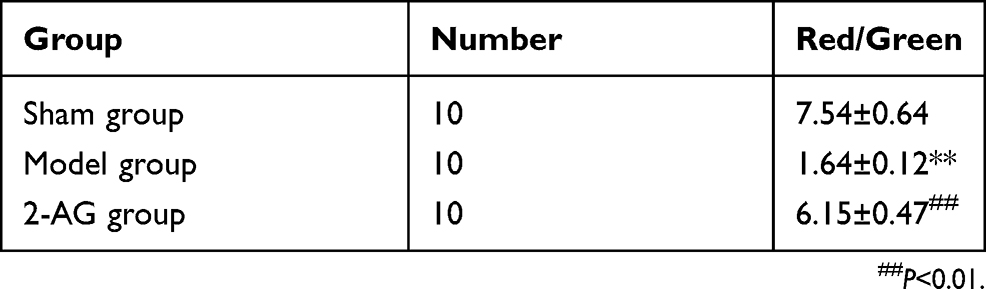 |
Table 2 Effects of endogenous cannabinoids 2-AG on mitochondrial membrane potential of hippocampal neurons in mice (± s) |
Effects of 2-arachidonoylglycerol 2-AG on the expression of AIF mRNA in hippocampal tissues of mice in each group
As shown in Table 3, real-time quantitative PCR (qPCR) results showed that the mRNA expression level of AIF in the hippocampal tissue of mice in the MCAO group was significantly higher than that of the sham group (P<0.01). However, the mRNA expression of AIF in hippocampal tissue of mice treated with 2-AG was significantly lower than that of MCAO group (P<0.01), which indicated that 2-AG could effectively inhibit the gene expression of AIF in the hippocampal tissue of MCAO mice.
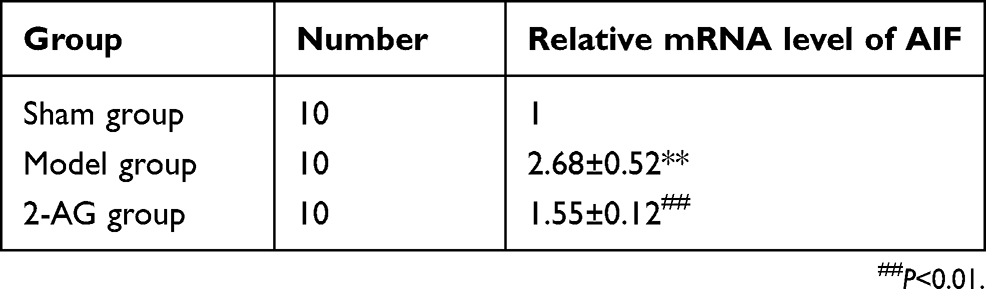 |
Table 3 Effects of endogenous cannabinoids 2-AG on the expression of AIF mRNA in hippocampal tissues of mice in each group (± s) |
Effects of 2-arachidonoylglycerol 2-AG on the expression of Endo G mRNA in hippocampal tissues of mice in each group
As shown in Table 4, real-time quantitative PCR (qPCR) results showed that the mRNA expression level of Endo G in the hippocampal tissue of mice in the MCAO group was significantly higher than that of the sham group (P<0.01). However, the mRNA expression of Endo G in hippocampal tissue of mice treated with 2-AG was significantly lower than that of MCAO group (P<0.01), which indicated that 2-AG could effectively inhibit the gene expression of Endo G in the hippocampal tissue of MCAO mice.
 |
Table 4 Effects of endogenous cannabinoids 2-AG on the expression of Endo G mRNA in hippocampal tissues of mice in each group (± s) |
Effects of 2-arachidonoylglycerol 2-AG on the expression of BNIP3 mRNA in hippocampal tissues of mice in each group
As shown in Table 5, real-time quantitative PCR (qPCR) results showed that the mRNA expression level of BNIP3 in the hippocampal tissue of mice in the MCAO group was significantly higher than that of the sham group (P<0.01). However, the mRNA expression of BNIP3 in hippocampal tissue of mice treated with 2-AG was significantly lower than that of MCAO group (P<0.01), which indicated that 2-AG could effectively inhibit the gene expression of BNIP3 in the hippocampal tissue of MCAO mice.
 |
Table 5 Effects of endogenous cannabinoids 2-AG on the expression of BNIP3 mRNA in hippocampal tissues of mice in each group (± s) |
Effects of 2-arachidonoylglycerol 2-AG on the expression of mitochondrial AIF protein in hippocampal tissue of mice in each group
As shown in Figure 1, Western Blot results showed that compared with the sham operation group, the protein expression of AIF in the mitochondria of the hippocampal tissue of mice in the MCAO group was significantly decreased. The protein expression of AIF in the hippocampal mitochondria of mice treated with 2-AG was significantly higher than that of the MCAO group, which indicated that 2-AG could effectively inhibit the secretion of AIF protein in mitochondria into cytoplasm.
 |
Figure 1 Expression of mitochondrial AIF protein in hippocampal tissue of mice in each group. |
Effects of 2-arachidonoylglycerol 2-AG on the expression of mitochondrial Endo G protein in hippocampal tissue of mice in each group
As shown in Figure 2, Western Blot results showed that compared with the sham operation group the protein expression of Endo G in the mitochondria of the hippocampal tissue of mice in the MCAO group was significantly decreased. The protein expression of AIF in the hippocampal mitochondria of mice treated with Endo G was significantly higher than that of the MCAO group, which indicated that 2-AG could effectively inhibit the secretion of Endo G protein in mitochondria into cytoplasm.
 |
Figure 2 Expression of mitochondrial Endo G protein in hippocampal tissue of mice in each group. |
Effects of endogenous cannabinoids 2-AG on the expression of mitochondrial BNIP3 protein in hippocampal tissue of mice in each group
As shown in Figure 3, Western Blot results showed that compared with the sham operation group, the protein expression of BNIP3 in the mitochondria of the hippocampal tissue of mice in the MCAO group was significantly decreased. The protein expression of AIF in the hippocampal mitochondria of mice treated with BNIP3 was significantly higher than that of the MCAO group, which indicated that 2-AG could effectively inhibit the secretion of BNIP3 protein in mitochondria into cytoplasm.
 |
Figure 3 Expression of mitochondrial BNIP3 protein in hippocampal tissue of mice in each group. |
CB1 receptor antagonist can inhibit the neuroprotective effect by 2-AG
We used 2-AG to specifically block CB1 function. The results showed that CB1 blocker 2-AG could reverse the effect of 2-AG on the expression of AIF, Endo G and BNIP3. These results suggest that CB1 is involved in the regulation of AIF, Endo G and BNIP3 expression by 2-AG.
Discussion
In recent years, the role of 2-arachidonoylglycerol 2-AG in neurological diseases has attracted wide attention. Neuroprotective effect is one of the hotspots of 2-arachidonoylglycerol 2-AG.13 The whole brain ischemic susceptibility is species-specific. Studies have shown that the Willis ring of C57BL/6 mice is innately underdeveloped, and the posterior traffic branch is underdeveloped, making it difficult for the basilar artery to replenish blood to the carotid system. Therefore, C57BL/6 mice were very sensitive to cerebral ischemia caused by carotid artery ligation.14 In this study, C57BL/6 mice were selected to establish a mouse model of total cerebral ischemia to explore the neuroprotective mechanism of 2-arachidonoylglycerol 2-AG on non-caspase-dependent neuron injury. Studies have confirmed that the main mechanism of nerve injury in cerebral ischemia is apoptosis of nerve cells.15 Therefore, how to inhibit the apoptosis of nerve cells is the focus of research on the treatment of cerebral ischemia. In this study, the apoptosis rate of hippocampal neurons in mice was detected by AV/PI double staining and flow cytometry to study the protective effect of endogenous cannabinoids 2-AG on hippocampal nerve cells in mice. The results of this study suggested that the anticerebral ischemia of 2-arachidonoylglycerol 2-AG might be related to its protective effect on nerve cells.
When stimulated by hypoxia or ischemia, cells undergo a series of lesions, and the main change is mitochondrial membrane depolarization.16 When the membrane potential of mitochondria decreases, the membrane permeability will change. Thus, apoptosis-related proteins are released into the cytoplasm, leading to apoptosis.17 Therefore, the changes of mitochondrial membrane potential are closely related to the study of cerebral ischemic diseases. In this study, the changes of mitochondrial membrane potential of hippocampal neurons in mice were determined by JC-1 probe labeling. The results of this study showed that 2-arachidonoylglycerol 2-AG could increase the mitochondrial membrane potential of hippocampal nerve cells in mice, which showed that 2-arachidonoylglycerol 2-AG could alleviate the decrease of mitochondrial membrane potential caused by cerebral ischemia to some extent.
When cells are stimulated by certain factors, mitochondria will release AIF, Endo G and BNIP3 related molecules of non-caspase-dependent apoptosis pathways into the cytoplasm, and eventually transposition into the nucleus leading to apoptosis.18 AIF is not only the apoptotic molecule of nerve cells, but also an important protein to maintain cell survival and mitochondrial activity. Studies have shown that in animal models of cerebral arterial embolism (MCAO), the apoptosis rate of nerve cells can be significantly reduced by down-regulating the expression of AIF.19 Endo G, as an endonuclease located between the mitochondrial intima, is an important mitochondrial protein in the non-caspase-dependent apoptosis pathway.20 When cells are stimulated by certain apoptosis signals, the mitochondria will release Endo G to the cytoplasm. When Endo G is transposed into nucleus, it will destroy DNA and lead to apoptosis.21 BNIP3 is a pro-apoptotic protein in the BH3-only subfamily, which is involved in apoptosis of glial cells, cardiac cells, epithelial cells and neurons.22 The increase of BNIP3 expression will lead to mitochondrial dysfunction and apoptosis. The results of this study confirmed that 2-arachidonoylglycerol 2-AG could decrease the mRNA expression levels of AIF, Endo G and BNIP3 in hippocampal tissues of mice, which suggested that 2-arachidonoylglycerol 2-AG could reduce the gene expressions of non-caspase-dependent apoptosis factors in the hippocampal tissue of mice. That was the inhibition of hippocampal tissue neuron apoptosis. Meanwhile, this study also found that 2-arachidonoylglycerol 2-AG could effectively up-regulate the protein expressions of AIF, Endo G and BNIP3 in the mitochondria of hippocampal tissue of mice, which indicated that 2-arachidonoylglycerol 2-AG could inhibit the secretions of AIF, Endo G and BNIP3 proteins in mitochondria into cytoplasm and protect neurons. The above results were consistent with the literature reports.
In conclusion, in this study, various indicators were used to evaluate the mechanism of 2-arachidonoylglycerol 2-AG against acute cerebral ischemia. The results of this study showed that 2-arachidonoylglycerol 2-AG had protective effects on neuron damage and its mechanism is to inhibit mitochondria-mediated apoptosis of non-caspase-dependent pathways by protecting the hippocampal nerve cells and their mitochondrial membrane potentials, so as to play a neuroprotective role.
Ethics approval and consent to participate
All applicable international, national and/or institutional guidelines for the care and use of animals were followed.
Availability of data and material
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.
Author contributions
He-Ying Zhong and Zhong-Yuan Xia contributed to the conception and design of the study; Zhou Yang contributed to the acquisition of data; Zhen Qiu and Shao-Qing Lei performed the experiments; Zhen Qiu contributed to the analysis of data; He-Ying Zhong wrote the manuscript. All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Wang J, Li C, Chen T, et al. Nafamostat mesilate protects against acute cerebral ischemia via blood-brain barrier protection. Neuropharmacology. 2016;105:398–410. doi:10.1016/j.neuropharm.2016.02.002
2. Qi Z, Liang J, Pan R, et al. Zinc contributes to acute cerebral ischemia-induced blood-brain barrier disruption. Neurobiol Dis. 2016;95:12–21. doi:10.1016/j.nbd.2016.07.003
3. Schwammenthal Y, Tsabari R, Orion D, et al. Shifting perceptions of risk and reward: use of anticoagulation in patients with acute brain ischemia and atrial fibrillation: nine-year data from a National Acute Stroke Registry (National Acute Stroke Israeli Survey [NASIS]). Stroke. 2017;48(4):1092–1094. doi:10.1161/STROKEAHA.116.015776
4. Brose SA, Golovko SA, Golovko MY. Brain 2-arachidonoylglycerol levels are dramatically and rapidly increased under acute ischemia-injury which is prevented by microwave irradiation. Lipids. 2016;51(4):487–495. doi:10.1007/s11745-016-4144-y
5. Stanne TM, Aberg ND, Nilsson S, et al. Low circulating acute brain-derived neurotrophic factor levels are associated with poor long-term functional outcome after ischemic stroke. Stroke. 2016;47(7):1943–1945. doi:10.1161/STROKEAHA.115.012383
6. Smith ML, Murphy K, Doucette CD, Greenshields AL, Hoskin DW. The dietary flavonoid fisetin causes cell cycle arrest, caspase-dependent apoptosis, and enhanced cytotoxicity of chemotherapeutic drugs in triple-negative breast cancer cells. J Cell Biochem. 2016;117(8):1913–1925. doi:10.1002/jcb.25490
7. Wolff S, Groseth A, Meyer B, et al. The New World arenavirus Tacaribe virus induces caspase-dependent apoptosis in infected cells. J Gen Virol. 2016;97(4):855–866. doi:10.1099/jgv.0.000403
8. Mahajan SD, Tutino VM, Redae Y, et al. C5a induces caspase-dependent apoptosis in brain vascular endothelial cells in experimental lupus. Immunology. 2016;148(4):407–419. doi:10.1111/imm.12619
9. De Leo A, Chen HS, Hu CCA, Lieberman PM, Flemington EK. Deregulation of KSHV latency conformation by ER-stress and caspase-dependent RAD21-cleavage. PLoS Pathog. 2017;13(8):e1006596. doi:10.1371/journal.ppat.1006596
10. Mwanza C, Chen Z, Zhang Q, Chen S, Wang W, Deng H. Simultaneous HPLC-APCI-MS/MS quantification of endogenous cannabinoids and glucocorticoids in hair. J Chromatogr B Analyt Technol Biomed Life Sci. 2016;1028:1–10. doi:10.1016/j.jchromb.2016.06.002
11. Papadogkonaki S, Theodorakis K, Thermos K. Endogenous and synthetic cannabinoids induce the downregulation of cannabinoid CB1 receptor in retina. Exp Eye Res. 2019;185:107694. doi:10.1016/j.exer.2019.107694
12. Pacheco DDF, Romero TRL, Duarte IDG. Ketamine induces central antinociception mediated by endogenous cannabinoids and activation of CB1 receptors. Neurosci Lett. 2019;699:140–144. doi:10.1016/j.neulet.2019.01.059
13. Leishman E, Cornett B, Spork K, Straiker A, Mackie K, Bradshaw HB. Broad impact of deleting endogenous cannabinoid hydrolyzing enzymes and the CB1 cannabinoid receptor on the endogenous cannabinoid-related lipidome in eight regions of the mouse brain. Pharmacol Res. 2016;110:159–172. doi:10.1016/j.phrs.2016.04.020
14. Gachet MS, Gertsch J. Quantitative analysis of arachidonic acid, endocannabinoids, N-acylethanolamines and steroids in biological samples by LCMS/MS: fit to purpose. J Chromatogr B Analyt Technol Biomed Life Sci. 2016;1012–1013:215–221. doi:10.1016/j.jchromb.2015.11.013
15. Song HL, Zhang X, Wang WZ, et al. Neuroprotective mechanisms of rutin for spinal cord injury through anti-oxidation and anti-inflammation and inhibition of p38 mitogen activated protein kinase pathway. Neural Regen Res. 2018;13(1):128–134. doi:10.4103/1673-5374.217349
16. Wang Y, Ren Q, Zhang X, Lu H, Chen J. Neuroprotective mechanisms of calycosin against focal cerebral ischemia and reperfusion injury in rats. Cell Physiol Biochem. 2018;45(2):537–546. doi:10.1159/000487031
17. Chen SQ, Wang ZS, Ma YX, et al. Neuroprotective effects and mechanisms of tea bioactive components in neurodegenerative diseases. Molecules. 2018;23(3):512.
18. Moriyama M, Moriyama H, Uda J, Matsuyama A, Osawa M, Hayakawa T. BNIP3 plays crucial roles in the differentiation and maintenance of epidermal keratinocytes. J Invest Dermatol. 2014;134(6):1627–1635. doi:10.1038/jid.2014.11
19. Liu J, Yuan C, Pu L, Wang J. Nutrient deprivation induces apoptosis of nucleus pulposus cells via activation of the BNIP3/AIF signalling pathway. Mol Med Rep. 2017;16(5):7253–7260. doi:10.3892/mmr.2017.7550
20. Xu W, Guo G, Li J, et al. Activation of Bcl-2-Caspase-9 apoptosis pathway in the testis of asthmatic mice. PLoS One. 2016;11(3):e0149353. doi:10.1371/journal.pone.0149353
21. Lee HJ, Jung YH, Choi GE, et al. BNIP3 induction by hypoxia stimulates FASN-dependent free fatty acid production enhancing therapeutic potential of umbilical cord blood-derived human mesenchymal stem cells. Redox Biol. 2017;13:426–443. doi:10.1016/j.redox.2017.07.004
22. Kapoor R, Rizvi F, Kakkar P. Naringenin prevents high glucose-induced mitochondria-mediated apoptosis involving AIF, Endo-G and caspases. Apoptosis. 2013;18(1):9–27. doi:10.1007/s10495-012-0781-7
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.