Back to Journals » Neuropsychiatric Disease and Treatment » Volume 19
The Interaction of LAMA2 and Duration of Illness Affects the Thickness of the Right Transverse Temporal Gyrus in Major Depressive Disorder
Authors Chen G, Li L, Sun T, Jiang C, Xu W, Chen S ![]() , Hu C
, Hu C ![]() , Yue Y, Wang T
, Yue Y, Wang T ![]() , Jiang W, Yuan Y
, Jiang W, Yuan Y ![]()
Received 11 August 2023
Accepted for publication 14 December 2023
Published 20 December 2023 Volume 2023:19 Pages 2807—2816
DOI https://doi.org/10.2147/NDT.S435025
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Richard J Porter
Gang Chen,1– 3 Lei Li,2,4 Taipeng Sun,1– 3 Chenguang Jiang,1,2 Wei Xu,1,2 Suzhen Chen,1,2 Changchun Hu,1 Yingying Yue,1,2 Tianyu Wang,1,2 Wenhao Jiang,1,2 Yonggui Yuan1,2
1Department of Psychosomatics and Psychiatry, ZhongDa Hospital, School of Medicine, Southeast University, Nanjing, Jiangsu, People’s Republic of China; 2Institute of Psychosomatics, School of Medicine, Southeast University, Nanjing, Jiangsu, People’s Republic of China; 3Department of Medical Psychology, Huai’an NO 3 People’s Hospital, Huaian, People’s Republic of China; 4Department of Sleep Medicine, The Fourth People’s Hospital of Lianyungang, Lianyungang, People’s Republic of China
Correspondence: Wenhao Jiang; Yonggui Yuan, Department of Psychosomatics and Psychiatry, ZhongDa Hospital, School of Medicine, Southeast University, No. 87 Dingjiaqiao, Gulou District, Nanjing, Jiangsu, 210009, People’s Republic of China, Tel +86-25-83285124, Email [email protected]; [email protected]
Background: Depression is a heritable brain disorder. Laminin genes were recently identified to affect the brain’s overall thickness through neurogenesis, differentiation, and migration in depression. This study aims to explore the effects of the LAMA2’s single nucleotide polymorphisms (SNP), a subunit gene of laminin, on the detected brain regions of patients with major depressive disorder (MDD).
Methods: The study included 89 patients with MDD and 60 healthy controls with T1-weighted structural magnetic resonance imaging and blood samples for genotyping. The interactions between LAMA2 gene SNPs and diagnosis as well as duration of illness (DOI) were explored on brain measures controlled for age, gender, and site.
Results: The right transverse temporal gyrus and right parahippocampal gyrus showed reduced thickness in MDD. Almost all seven LAMA2 SNPs showed significant interactions with diagnosis on both gyrus (corrected p < 0.05 or trending). In MDD, rs6569604, rs2229848, rs2229849, rs2229850, and rs2784895 interacted with DOI on the right transverse temporal gyrus (corrected p < 0.05), but not the right parahippocampal gyrus.
Conclusion: The thickness of the right transverse temporal gyrus in patients with MDD may be affected by LAMA2 gene and DOI.
Keywords: major depressive disorder, right transverse temporal gyrus, LAMA2, interaction
Introduction
Major depressive disorder (MDD), a heritable brain disorder, commonly begins in youth and continues throughout one’s life. According to World Health Organization, the lifetime prevalence of depression is around 3.63%,1 which has caused an enormous burden.2 The pathogenesis of MDD is still unclear, and several brain structure alterations. Two meta-analyses found that the hippocampal volume, especially the cortical thickness of the orbitofrontal cortex and anterior cingulate cortex in patients with depression, were smaller than those in normal people.3,4 In addition, the disease course of MDD might affect these brain deficits. Several longitudinal studies of depression have found that gray matter loss is associated with the disease course of depression.5,6
The link between genetic variation and brain structure is a complex and controversial topic.7,8 Generally, the primary genetic cause of brain structural changes is usually thought to be a massive amount of common variation with insignificant effects, accounting for 26% of average thickness variance and 34% of surface area variation. Common variations are also likely responsible for changes in the brain’s structure in MDD, accounting for about 21% of genetic variance.9 However, some research has revealed that some genes play a role in forming particular cortical regions, which affect particular brain regions in different people. A meta-analysis identified 237 significant loci affecting cortical thickness and cortical area,10 of which rs533577 encoding Lama receptor-binding protein is a key fetal cortical expression quantitative trait locus,11 significantly affecting brain thickness. Identifying specific risk-associated genes and providing key information on pathogenesis is critical for developing depression diagnosis and treatment strategies.
Laminin is a large heterotrimer composed of α, β and γ subunits that is widely expressed in the central nervous system during brain development,12 regulating neural stem cell proliferation, axon growth,13,14 synapse formation.15 Laminin mutations lead to many congenital disorders. For example, Laminin β2 mutations lead to Pierson syndrome, a genetic disorder with renal failure and vision loss as the main symptoms.16 Studies have shown that another inherited disease, herpetic junctional epidermolysis bullosa, is associated with LAMA332 (α3, β3, and γ2).17,18 It has also been found that LAMAα3 mutations can lead to laryngo-onycho-cutaneous syndrome a variant of junctional epidermolysis bullosa.19 Secondly, LAMA has also been associated with the development of tumours squamous cell carcinoma, blast tumor, pancreatic cancer, etc.20–22 LAMA2, which encodes the α2 chain, whose mutation disrupts the expression or binding ability of α2 laminin is the most important cause of muscular dystrophy type 1A (MDC1A).23 It was reported that the loss of LAMA2 could result in brain abnormalities such as neuronal migration problems, which can lead to anencephaly.24,25 In some cases, patients with muscle illnesses, such as α-dystroglycanopathies were shown to have clear cortical stratification anomalies, resulting in significant neurological dysfunction.26,27 In a study of Chinese Attention-deficit/hyperactivity disorder (ADHD) children, LAMA2 SNP rs2229848 was found to be one of the important SNV variants. ADHD shares a high genetic predisposition with depression,28 and individuals with ADHD are estimated to have a 6.5-fold increased risk of depression.29
Our team previously discovered that seven SNPs (rs2784895, rs2229848, rs2229849, rs2229850, rs2275214, rs6569604, and rs6938825) of the LAMA2 gene are associated with MDD in a next-generation sequencing30(Supplementary 1, Figure S1, Table S1). We aimed to explore the effects of LAMA2 variations on brain measures highlighted by MDD. Then, we tried to reveal the interactions between LAMA2 and DOI on brain measures.
Methods
Participants
The study was approved by the Ethics Committee of the Affiliated ZhongDa Hospital of Southeast University (Approval number: 2016ZDSYLL100-P01). The study was registered in the Chinese Clinical Trial Registry, registration number: ChiCTR-ROC-17013448. After receiving a thorough description of the study in accordance with the Declaration of Helsinki, all participants or their legal representatives provided written informed consent.
The data of this study come from the genetic imaging study of MDD in the ZhongDa Hospital Affiliated to Southeast University. The included patients must have both genetic and T1 weighted images data. Following screening, 60 sex-matched healthy controls (31 women and 29 men) without a history of mental or physical illnesses and 89 MDD patients (50 women and 39 men) were enrolled.
Two senior psychiatrists conducted MDD diagnosis according to DSM-IV. All patients were 18–60 years old. Participants were excluded if: (1) pregnant or lactating, (2) had another axis I diagnostic, (3) used a psychoactive substance such as tobacco, alcohol, or drugs, or (4) had a severe physical illness, they would be excluded from the study (tumor, organic brain diseases cardiovascular diseases, endocrine disease, and autoimmune diseases and so on). Matched healthy controls were recruited through advertising. Experienced psychiatrists determined that the controls had no prior or current psychiatric illnesses.
Clinical Assessments
A bespoke questionnaire was used to collect demographic characteristics of patients and controls at the start of the study (such as age, sex, education levels, age of onset, number of episodes, duration of illness, and others). At baseline and two weeks, expert psychiatrists used the HAMD-17 to assess the severity of depression symptoms. Meanwhile, HC required that these scales be normal.
Genotyping
5 mL peripheral blood samples were collected from each participant into EDTA tubes and stored in a refrigerator at −80°C Tianhao Biotechnology (Shanghai, China) using pre-designed Illumina next sequencing and array technology to execute the LAMA2 gene assay (Illumina Inc., San Diego, CA, USA). PLINK1.9 software (https://www.cog-genomics.org/plink/1.9/) was used to perform standard quality control procedures such as Hardy-Weinberg equilibrium (HWE) tests, linkage disequilibrium statistics, and allele and genotype frequencies. According to the findings, none of the above seven SNPs depart from HWE (P > 0.05). We looked through dbGAP (https://www.ncbi.nlm.nih.gov/gap/) and determined minor alleles according to East Asia samples and reports.
MRI Data and Cortical Characteristics Calculation
Data collection for MRI images Utilizing a Siemens 3.0 T scanner and a 12-channel head coil, high-resolution three-dimensional T1-weighted images were produced using a magnetic fast gradient echo (mprage) program (Siemens, Erlangen, Germany). Our past investigations have a full description of the parameters31 (Supplementary 2). The whole automated “recon-all” standard technique from FreeSurfer 7.1.1 (Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts, USA, https://surfer.nmr.mgh.harvard.edu/) was used in a Linux desktop environment to handle raw structural pictures. Motion correction, non-uniform intensity correction, Talairach transform, skull stripping, subcortical and white matter segments, normalization and atlas registration, surface reconstruction, registration, and cortical parcellation were among the processes completed. The data were carefully validated for the accuracy of skull strip, white matter segmentation, intensity normalization, and topologic defect after all subjects’ images had been preprocessed. One image was manually modified and then reconstructed from the control points of modifications due to a major topologic fault. The DKT atlas32 was used to segment gray matter.
We extracted the cortex thickness and surface area and local gyrification index (LGI) for analysis and obtained the cortical thickness and surface area after the data was pre-processed by the recon-all standard. LGI is based on the soft meningeal surface image output from recon-all, plotting the outer surface immediately adjacent to the soft meningeal edge and calculating the ratio of local cortical area to outer surface area by vertex.
Statistical Analysis
All statistical analyses were performed in SPSS (Version 21.0, Chicago), with the exception of the false discovery rate (FDR) (Benjamini and Hochberg, 1995). Patient demographics and clinical characteristics were described as the mean ± standard deviation (SD). Independent samples t-tests were performed for continuous variables, and χ2 tests were performed for categorical variables.
Age, gender, and site effects were regressed out from all brain measures prior to analysis of brain structure measures. One-way ANOVA was used to explore differences in patient brain structure indices between HC and MDD groups and between different DOIs. Gene-diagnosis, gene-DOI interactions using general linear models. The corrected P < 0.05 was used to determine significance.
Results
Demographics and Clinical Characteristics
The demographic, genetic, and clinical characteristics of all 149 participants are shown in Table 1. There was no significant difference in gender and education level between the MDD group and HC group, but the age of the MDD group was higher than the HC group. There were significant differences in seven LAMA2 SNPs between the two groups. The HAMD-17 score decreased from 22.34 ± 3.64 to 9.79±4.75 (t = 18.707, P < 0.001) after treatment, indicating that the severity of depressive symptoms was significantly relieved.
 |
Table 1 Demographic and Clinical Assessments of Participants |
Group Differences in Cortical Characteristics
After controlling for age and gender factors, the brain structure indicators of the two groups were compared through a general linear model. We did not find a significant difference between the two groups in the surface area and LGI. However, there were differences in thickness between the two groups in the right parahippocampal gyrus and the right transverse temporal gyrus (p < 0.05 for all, corrected by FDR; Figure 1).
 |
Figure 1 Differences in thickness of right transverse temporal gyrus and right parahippocampal gyrus between HC and MDD. (a) Thickness of right parahippocampal gyrus in the MDD was significantly lower than HC (p < 0.001). (b) Thickness of right transverse temporal gyrus in the MDD was significantly lower than HC (p < 0.001). ***p < 0.001. Abbreviations: TH, cortical thickness; HC, health control; MDD, major depressive disorder. |
Difference of Cortical Thickness in Patients with Different DOI
The DOI was satisfied into two groups: less than or equal to 1 year and longer than 1 year. The right transverse temporal gyrus showed a significant difference in cortical thickness between the two groups (p = 0.016, Figure 2a), whereas thickness of the right parahippocampal gyrus did not show a significant difference between the two groups (Figure 2b).
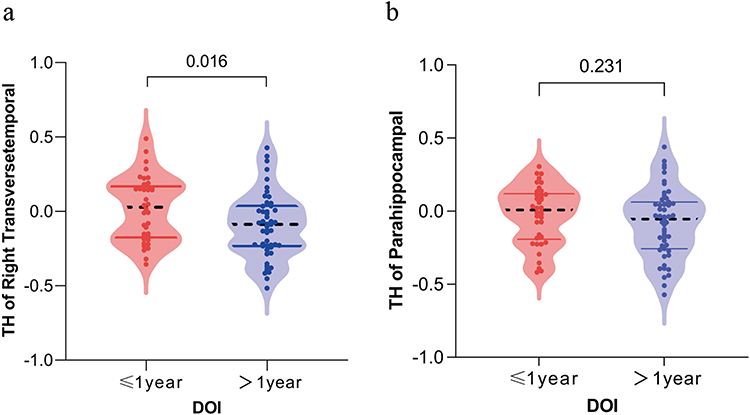 |
Figure 2 Scatter-box violin plot showing the difference of thickness in the right transverse temporal gyrus and right parahippocampal gyrus in MDD patients with different DOI. (a) Thickness of right transverse temporal gyrus in MDD patients with DOI ≤ 1 year was significantly lower than that in the group > 1 year (p = 0.016). (b) There was no significant difference in thickness of right parahippocampal gyrus between MDD patients with DOI ≤ 1 year and >1 year (p = 0.231). Abbreviations: TH, cortical thickness; DOI, duration of illness. |
LAMA2 SNPs-Diagnosis Interaction on Brain Structure
A general linear model revealed the effect of LAMA2 SNPs-diagnosis interaction on two distinct brain regions. Among the seven LAMA2 SNPs, rs2784895, rs2229848, rs2229849, and rs2229850 corrected significant affected cortical thickness of two brain regions, while rs2275214, rs6569604, and rs6938825 only had corrected significant effects on thickness of right parahippocampal gyrus (Table 2).
 |
Table 2 Interaction of LAMA2 SNPs-Diagnosis on Thickness of Right Transverse Temporal Gyrus and Right Parahippocampal Gyrus |
LAMA2 SNPs-DOI Interaction on Brain Structure
Further exploration of the interaction between LAMA2 SNPs-DOI on cortical thickness revealed that the interaction of rs6569604, rs2229848, rs2229849, rs2229850, and rs2784895 with DOI had corrected significant effects on thickness of right transverse temporal gyrus, but rs2229849 showed the opposite effect of other SNPs (Figure 3). No corrected significant LAMA2 SNPs-DOI interactions on thickness of the right parahippocampal gyrus (Table 3).
 |
Table 3 Interaction of LAMA2 SNPs-DOI on Thickness of Right Transverse Temporal Gyrus and Right Parahippocampal Gyrus |
 |
Figure 3 (a–e) The interaction effect of the five SNP of LAMA2 and MDD on thickness of right transverse temporal gyrus. rs2229850, rs2229848, rs2229849, rs6569604 and rs2784895 had significant interaction with DOI on thickness of right transverse temporal gyrus, but the minor allele of rs2229850 shows opposite effects to the other four SNPs on thickness of right transverse temporal gyrus (p < 0.05). Abbreviation: DOI, duration of illness. |
Discussion
This study explored the differences in brain structure between MDD and HC patients, as well as the effects of the interaction of seven LAMA2 gene SNPs with diagnosis and DOI on different brain regions. Our study found significant case vs control in the thickness of the right transverse temporal gyrus significantly lower than the right parahippocampal gyrus. Further exploration showed that the interaction of LAMA2 SNPs with diagnosis and DOI had a significant effect on these two brain regions, but rs2229849 showed the opposite effect of other SNPs.
Cortical thickness might reflect the number of neurogenic divisions of neural progenitor cells in the brain, and these changes may occur more rapidly in specific regions of the cortex, such as the hippocampus.33,34 Cortical thickness and cortical surface area together constitute gray matter volume, and changes in hippocampal volume are closely related to the occurrence of depression.35,36 The right parahippocampal was also smaller than the HC in elderly posttraumatic stress disorder (PTSD) patients and was negatively correlated with the severity,37 which suggests that the right parahippocampal may also be associated with abnormal emotional processing. This is consistent with our finding that the thickness of right parahippocampal gyrus was significantly lower in MDD patients. This study also found that the thickness of right transverse temporal gyrus was significantly lower than that of HC in MDD patients, and there was a slight decrease trend with age. The transverse temporal gyrus, also referred to as Heschl’s gyrus (HG), is connected to the primary sensory cortex. Its principal function is to integrate and process auditory information.38 Thus, it regulates sensory social processing, behaviors, and emotions. Previous studies have found a trend of thinning of the temporal cortex in MDD patients,3 especially in adult depression patients with a history of childhood neglect and abuse.39,40 Our research further demonstrated the right transverse temporal gyrus’ possible role in emotional disorders, and it is a significant brain region that requires future research.
We found that multiple LAMA2 gene SNPs interacted with diagnosis to affect the thickness of the right transverse temporal gyrus and the right parahippocampal gyrus. The LAMA2 gene was reported in hippocampal neurons’ cortex and dendritic spines. The expression of this gene was seen to decrease in case of neuron damage and increase during the recovery of neuronal synapses.41 In addition to LAMA, the parahippocampal gyrus and the transverse temporal gyrus are also affected by BDNF,42 catechol O-methyltransferase (COMT),43 dopamine D2 receptor (DRD2), NRG1, Disrupted-In-Schizophrenia-1 (DISC1), microcephaly associated (ASPM),44 and various other genes that influence them throughout different stages of life. BDNF encodes brain-derived neurotrophic factor, which plays a significant role in the neurogenesis and development of the parahippocampal and transverse temporal gyrus as well as other brain regions during early life.45 Additionally, COMT and DRD2 influence dopamine metabolism and cognitive function during adulthood.46 Furthermore, NRG1 is linked to the neuronal development and synaptic plasticity of the parahippocampal gyrus.47
The brain structure of normal people also changes with age. Studies have shown the morphology and microstructure of brain regions during normal aging, including the transverse temporal gyrus.48 However, the structural brain changes in MDD patients differ from those in normal subjects of the same age. Children with MDD were found to have a significant reduction in bilateral cortical gray matter thickness and right hemisphere volume;49 The cortical thickness of the right parahippocampal gyrus in patients with bipolar disorder was significantly thinner than that in HCs,50 while the cortical thickness of the right transverse temporal gyrus in patients with schizophrenia was significantly thinner than that in HCs.51,52
The study also found rs2229850, rs2229848, rs2229849, rs6569604 and rs2784895 had significant interaction with DOI on thickness of right transverse temporal gyrus. However, the minor alleles of five SNPs had inconsistent impacts on cortical thickness when interacted with DOI. Thickness of right transverse temporal gyrus was thinner in MDD patients with rs6569604, rs2229848, rs2229849 and rs2784895 minor alleles, but the opposite was true in patients with the rs2229850 minor allele. In contrast to the other four SNPs, rs2229850 may be a protective factor for thickness in the right temporal transverse gyrus. LAMA2 gene is the most common cause of MDC1A,53 Nevertheless, the condition has a widespread impact and affects not just the muscles but can also cause brain malformations, mental retardation, and other alterations.54 Moreover, it is frequently accompanied by seizures. Electroencephalogram (EEG) showed that temporal and occipital epilepsy was the most common, further proving that the right transverse temporal might be the primary brain region affected by the LAMA2 gene.
Limitations
There are inevitable limitations with this study. Firstly, the study had a small sample size, which may have led to false positives and false negatives. Additionally, this is a cross-sectional study and it is not possible to establish a causal link between genetics and the brain. Secondly, the region of interest was derived only through case vs control difference, and we only included seven LAMA2 gene SNPs highlighted in previous results. This may have limited the findings of the study. Thirdly, it is the first time that we have found that the cortex of the right transverse temporal gyrus is thinner in MDD patients, and further studies with larger samples are needed to verify this finding, especially longitudinal cohort studies with independent samples are critical to illustrate the links between detected brains features and genetic variations.
Conclusion
In conclusion, the LAMA2 genotype modulates right transverse temporal gyrus thickness in MDD patients. The LAMA2 gene is widely expressed in the brain and plays an important role in neural development and differentiation, suggesting that it may have a role in the pathophysiology of depression. Research on the function of LAMA2 gene will help to explore the pathogenesis of genetic and environmental factors in depression.
Data Sharing Statement
The datasets presented in this article are not readily available because of the sensitive nature of the questions asked in our study. Requests to access the datasets should be directed to [email protected].
Ethical Approval
The study was approved by the Ethics Committee of the Affiliated ZhongDa Hospital of Southeast University (Approval number: 2016ZDSYLL100-P01). All procedures performed in this study were in accordance with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Consent to Participate
Written informed consent was obtained from all individual participants included in the study.
Acknowledgments
We are doubly grateful to all of the participating colleagues and students from Zhongda Hospital for taking their time to assist with disease control and data collection for this study. This work was supported by the National Key Research and Development Program of China (2016YFC1306702), the National Natural Science Foundation of China (81971277). The funders had no additional involvement in the study’s design, data collecting, analysis, and interpretation, report writing, or the decision to publish the research.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by grants from the National Natural Science Foundation of China [81971277].
Disclosure
The authors declare that there were no commercial or financial relationships that might be considered as a potential conflict of interest during the research.
References
1. Rehm J, Shield K. Global burden of disease and the impact of mental and addictive disorders. Curr Psychiatry Rep. 2019;21(2):10. doi:10.1007/s11920-019-0997-0
2. Malhi G, Mann J. Depression. Lancet. 2018;392(10161):2299–2312. doi:10.1016/S0140-6736(18)31948-2
3. Schmaal L, Hibar D, Sämann P, et al. Cortical abnormalities in adults and adolescents with major depression based on brain scans from 20 cohorts worldwide in the ENIGMA Major Depressive Disorder Working Group. Mol Psychiatry. 2017;22(6):900–909. doi:10.1038/mp.2016.60
4. Gray J, Müller V, Eickhoff S, Fox P. Multimodal abnormalities of brain structure and function in major depressive disorder: a meta-analysis of neuroimaging studies. Am J Psychiatry. 2020;177(5):422–434. doi:10.1176/appi.ajp.2019.19050560
5. Phillips J, Batten L, Tremblay P, Aldosary F, Blier P. A prospective, longitudinal study of the effect of remission on cortical thickness and hippocampal volume in patients with treatment-resistant depression. Int J Neuropsychopharmacol. 2015;18(8). doi:10.1093/ijnp/pyv037
6. Lebedeva A, Sundström A, Lindgren L, et al. Longitudinal relationships among depressive symptoms, cortisol, and brain atrophy in the neocortex and the hippocampus. Acta Psychiatr Scand. 2018;137(6):491–502. doi:10.1111/acps.12860
7. Evans P, Gilbert S, Mekel-Bobrov N, et al. Microcephalin, a gene regulating brain size, continues to evolve adaptively in humans. Science. 2005;309(5741):1717–1720. doi:10.1126/science.1113722
8. Marques J, Duarte S, Costa S, et al. Atypical phenotype in two patients with LAMA2 mutations. Neuromuscul Disord. 2014;24(5):419–424. doi:10.1016/j.nmd.2014.01.004
9. Amin N, Jovanova O, Adams H, et al. Exome-sequencing in a large population-based study reveals a rare Asn396Ser variant in the LIPG gene associated with depressive symptoms. Mol Psychiatry. 2017;22(4):537–543. doi:10.1038/mp.2016.101
10. Grasby K, Jahanshad N, Painter J, et al. The genetic architecture of the human cerebral cortex. Science. 2020;367(6484). doi:10.1126/science.aay6690
11. Digiacomo V, Meruelo D. Looking into laminin receptor: critical discussion regarding the non-integrin 37/67-kDa laminin receptor/RPSA protein. Biol Rev Camb Philos Soc. 2016;91(2):288–310. doi:10.1111/brv.12170
12. Gawlik K, Harandi V, Cheong R, Petersén Å, Durbeej M. Laminin Α1 reduces muscular dystrophy in dy(2J) mice. Matrix Biol. 2018;70:36–49. doi:10.1016/j.matbio.2018.02.024
13. Morissette N, Carbonetto S. Laminin alpha 2 chain (M chain) is found within the pathway of avian and murine retinal projections. J Neurosci. 1995;15(12):8067–8082. doi:10.1523/JNEUROSCI.15-12-08067.1995
14. Ahmed M, Marziali L, Arenas E, Feltri M, Ffrench-Constant C. Laminin α2 controls mouse and human stem cell behaviour during midbrain dopaminergic neuron development. Development. 2019;146(16). doi:10.1242/dev.172668
15. Powell S, Williams C, Nomizu M, Yamada Y, Kleinman H. Laminin-like proteins are differentially regulated during cerebellar development and stimulate granule cell neurite outgrowth in vitro. J Neurosci Res. 1998;54(2):233–247. doi:10.1002/(SICI)1097-4547(19981015)54:2<233::AID-JNR11>3.0.CO;2-5
16. Zenker M, Aigner T, Wendler O, et al. Human laminin beta2 deficiency causes congenital nephrosis with mesangial sclerosis and distinct eye abnormalities. Hum Mol Genet. 2004;13(21):2625–2632. doi:10.1093/hmg/ddh284
17. Pulkkinen L, Christiano A, Airenne T, Haakana H, Tryggvason K, Uitto J. Mutations in the gamma 2 chain gene (LAMC2) of kalinin/laminin 5 in the junctional forms of epidermolysis bullosa. Nat Genet. 1994;6(3):293–297. doi:10.1038/ng0394-293
18. Mühle C, Jiang Q, Charlesworth A, Bruckner-Tuderman L, Meneguzzi G, Schneider H. Novel and recurrent mutations in the laminin-5 genes causing lethal junctional epidermolysis bullosa: molecular basis and clinical course of herlitz disease. Hum Genet. 2005;116(1–2):33–42. doi:10.1007/s00439-004-1210-y
19. Mclean W, Irvine A, Hamill K, et al. An unusual N-terminal deletion of the laminin alpha3a isoform leads to the chronic granulation tissue disorder laryngo-onycho-cutaneous syndrome. Hum Mol Genet. 2003;12(18):2395–2409. doi:10.1093/hmg/ddg234
20. Marinkovich M. Tumour microenvironment: laminin 332 in squamous-cell carcinoma. Nat Rev Cancer. 2007;7(5):370–380. doi:10.1038/nrc2089
21. Dai J, Cimino P, Gouin K 3rd, et al. Astrocytic laminin-211 drives disseminated breast tumor cell dormancy in brain. Nat Cancer. 2022;3(1):25–42. doi:10.1038/s43018-021-00297-3
22. Erice O, Narayanan S, Feliu I, et al. LAMC2 regulates key transcriptional and targetable effectors to support pancreatic cancer growth. Clin Cancer Res. 2023;29(6):1137–1154. doi:10.1158/1078-0432.CCR-22-0794
23. Allamand V, Sunada Y, Salih M, et al. Mild congenital muscular dystrophy in two patients with an internally deleted laminin alpha2-chain. Hum Mol Genet. 1997;6(5):747–752. doi:10.1093/hmg/6.5.747
24. Saredi S, Gibertini S, Matalonga L, et al. Exome sequencing detects compound heterozygous nonsense LAMA2 mutations in two siblings with atypical phenotype and nearly normal brain MRI. Neuromuscul Disord. 2019;29(5):376–380. doi:10.1016/j.nmd.2019.04.001
25. Previtali S, Zambon A. Lama2 neuropathies: human findings and pathomechanisms from mouse models. Front Mol Neurosci. 2020;13:60. doi:10.3389/fnmol.2020.00060
26. Raphael A, Couthouis J, Sakamuri S, et al. Congenital muscular dystrophy and generalized epilepsy caused by GMPPB mutations. Brain Res. 2014;1575:66–71. doi:10.1016/j.brainres.2014.04.028
27. Zaum A, Kolokotronis K, Kress W, Goebel H, Rost S, Seeger J. A new case expanding the mutation and phenotype spectrum of TMEM5-related alpha-dystroglycanopathy. Neuromuscul Disord. 2018;28(8):671–674. doi:10.1016/j.nmd.2018.06.006
28. Faraone S, Larsson H. Genetics of attention deficit hyperactivity disorder. Mol Psychiatry. 2019;24(4):562–575. doi:10.1038/s41380-018-0070-0
29. Gundel L, Pedersen C, Munk-Olsen T, Dalsgaard S. Longitudinal association between mental disorders in childhood and subsequent depression - a nationwide prospective cohort study. J Affect Disord. 2018;227:56–64. doi:10.1016/j.jad.2017.10.023
30. Jang W. The multivariable data-driven imaging genetic study on anhedonia in major depressive disorder; 2018.
31. Liu X, Hou Z, Yin Y, et al. Dopamine multilocus genetic profile, spontaneous activity of left superior temporal gyrus, and early therapeutic effect in major depressive disorder. Front Psychiatry. 2020;11:591407. doi:10.3389/fpsyt.2020.591407
32. Fischl B. Freesurfer. Neuroimage. 2012;62(2):774–781. doi:10.1016/j.neuroimage.2012.01.021
33. Fotuhi M, Do D, Jack C. Modifiable factors that alter the size of the hippocampus with ageing. Nat Rev Neurol. 2012;8(4):189–202. doi:10.1038/nrneurol.2012.27
34. Moreno-Jiménez E, Terreros-Roncal J, Flor-García M, Rábano A, Llorens-Martín M. Evidences for adult hippocampal neurogenesis in humans. J Neurosci. 2021;41(12):2541–2553. doi:10.1523/JNEUROSCI.0675-20.2020
35. Sahay A, Hen R. Adult hippocampal neurogenesis in depression. Nat Neurosci. 2007;10(9):1110–1115. doi:10.1038/nn1969
36. Eisch A, Petrik D. Depression and hippocampal neurogenesis: a road to remission. Science. 2012;338(6103):72–75. doi:10.1126/science.1222941
37. Stark E, Parsons C, Van Hartevelt T, et al. Post-traumatic stress influences the brain even in the absence of symptoms: a systematic, quantitative meta-analysis of neuroimaging studies. Neurosci Biobehav Rev. 2015;56:207–221. doi:10.1016/j.neubiorev.2015.07.007
38. Levy F. Pharmacological and therapeutic directions in ADHD: specificity in the PFC. Behav Brain Funct. 2008;4:12. doi:10.1186/1744-9081-4-12
39. Jaworska N, Macmaster F, Gaxiola I, Cortese F, Goodyear B, Ramasubbu R. A preliminary study of the influence of age of onset and childhood trauma on cortical thickness in major depressive disorder. Biomed Res Int. 2014;2014:410472. doi:10.1155/2014/410472
40. Tozzi L, Garczarek L, Janowitz D, et al. Interactive impact of childhood maltreatment, depression, and age on cortical brain structure: mega-analytic findings from a large multi-site cohort. Psychol Med. 2020;50(6):1020–1031. doi:10.1017/S003329171900093X
41. Tian M, Hagg T, Denisova N, Knusel B, Engvall E, Jucker M. Laminin-alpha2 chain-like antigens in CNS dendritic spines. Brain Res. 1997;764(1–2):28–38. doi:10.1016/S0006-8993(97)00420-4
42. Montag C, Weber B, Fliessbach K, Elger C, Reuter M. The BDNF Val66Met polymorphism impacts parahippocampal and amygdala volume in healthy humans: incremental support for a genetic risk factor for depression. Psychol Med. 2009;39(11):1831–1839. doi:10.1017/S0033291709005509
43. Di Giorgio A, Caforio G, Blasi G, et al. Catechol-O-methyltransferase Val(158)Met association with parahippocampal physiology during memory encoding in schizophrenia. Psychol Med. 2011;41(8):1721–1731. doi:10.1017/S0033291710002278
44. Passemard S, Verloes A, Billette De Villemeur T, et al. Abnormal spindle-like microcephaly-associated (ASPM) mutations strongly disrupt neocortical structure but spare the hippocampus and long-term memory. Cortex. 2016;74:158–176. doi:10.1016/j.cortex.2015.10.010
45. Miranda M, Morici J, Zanoni M, Bekinschtein P. Brain-derived neurotrophic factor: a key molecule for memory in the healthy and the pathological brain. Front Cell Neurosci. 2019;13:363. doi:10.3389/fncel.2019.00363
46. Furman D, Pappas I, White R 3rd, Kayser A, D’esposito M. Enhancing dopamine tone modulates global and local cortical perfusion as a function of COMT val158met genotype. Neuroimage. 2021;242:118472. doi:10.1016/j.neuroimage.2021.118472
47. Cannon D, Walshe M, Dempster E, et al. The association of white matter volume in psychotic disorders with genotypic variation in NRG1, MOG and CNP: a voxel-based analysis in affected individuals and their unaffected relatives. Transl Psychiatry. 2012;2(10):E167. doi:10.1038/tp.2012.82
48. Zhao X, Wu Q, Chen Y, Song X, Ni H, Ming D. The conjoint analysis of microstructural and morphological changes of gray matter during aging. Front Neurol. 2019;10:184. doi:10.3389/fneur.2019.00184
49. Luby J, Belden A, Jackson J, et al. Early childhood depression and alterations in the trajectory of gray matter maturation in middle childhood and early adolescence. JAMA Psychiatry. 2016;73(1):31–38. doi:10.1001/jamapsychiatry.2015.2356
50. Rimol L, Hartberg C, Nesvåg R, et al. Cortical thickness and subcortical volumes in schizophrenia and bipolar disorder. Biol Psychiatry. 2010;68(1):41–50. doi:10.1016/j.biopsych.2010.03.036
51. Alkan E, Davies G, Evans S. Cognitive impairment in schizophrenia: relationships with cortical thickness in fronto-temporal regions, and dissociability from symptom severity. NPJ Schizophr. 2021;7(1):20. doi:10.1038/s41537-021-00149-0
52. Wang C, Tishler T, Nuechterlein K, Ellingson B. Cortical thickness, gray-white matter contrast, and intracortical myelin in first-episode schizophrenia patients treated with long-acting paliperidone palmitate versus oral antipsychotics. Psychiatry Res. 2023;326:115364. doi:10.1016/j.psychres.2023.115364
53. Herrmann R, Straub V, Meyer K, Kahn T, Wagner M, Voit T. Congenital muscular dystrophy with laminin alpha 2 chain deficiency: identification of a new intermediate phenotype and correlation of clinical findings to muscle immunohistochemistry. Eur J Pediatr. 1996;155(11):968–976. doi:10.1007/BF02282889
54. Camelo C, Artilheiro M, Martins Moreno C, et al. Brain MRI abnormalities, epilepsy and intellectual disability in LAMA2 related dystrophy - a genotype/phenotype correlation. J Neuromuscul Dis. 2023;10(4):483–492. doi:10.3233/JND-221638
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.