Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 17
The Interaction Between Body Mass Index Genetic Risk Score and Dietary Intake on Weight Status: A Systematic Review
Authors Sokary S, Almaghrbi H, Bawadi H ![]()
Received 12 December 2023
Accepted for publication 15 February 2024
Published 27 February 2024 Volume 2024:17 Pages 925—941
DOI https://doi.org/10.2147/DMSO.S452660
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Juei-Tang Cheng
Sara Sokary,1 Heba Almaghrbi,2 Hiba Bawadi1
1Department of Human Nutrition, College of Health Sciences, QU Health, Qatar University, Doha, Qatar; 2Department of Biomedical Science, College of Health Sciences, QU Health, Qatar University, Doha, Qatar
Correspondence: Hiba Bawadi, Email [email protected]
Background: The escalating global obesity epidemic and the emergence of personalized medicine strategies point to the pressing need to investigate the interplay between genetic risk scores (GRSs), dietary intake, and their combined impact on weight status. This systematic review synthesizes evidence from diverse studies to elucidate how dietary patterns and individual foods interact with genetic predisposition to obesity.
Methods: Literature searches were conducted in the PubMed, Embase, Science Direct, and Scopus databases until August 2023, following PRISMA guidelines. Out of 575 articles, 15 articles examining the interaction between genetic risk score for body mass index and dietary intake on weight outcomes met the inclusion criteria. All included studies were cross-sectional in design and were assessed for quality using the Newcastle‒Ottawa Scale.
Results: Unhealthy dietary intake exacerbated the genetic predisposition to obesity, evident in studies assessing Western diet, sulfur microbial diet, and individual macronutrients, including saturated fatty acids, sugar-sweetened beverages and fried foods. Conversely, adhering to healthier dietary intake mitigated the genetic predisposition to obesity, as observed in studies involving Alternative Healthy Eating Index, Alternative Mediterranean Diet, Dietary Approach to Stop Hypertension scores, healthy plant-based diets, and specific foods such as fruits, vegetables, and n-3 polyunsaturated fatty acids.
Conclusion: This is the first systematic review to explore the interaction between genetics and dietary intake in shaping obesity outcomes. The findings have implications for tailored interventions; however, more controlled clinical trials with robust designs are needed to be able to recommend personalized nutrition based on nutrition for obesity prevention and management.
Keywords: Body Mass Index, Genetic Risk Score, Dietary Intake, Weight Status, Systematic Review
Introduction
Obesity is a complex multifactorial health condition with a profound impact on global public health.1 The etiology of this morbid disease involves an interplay between genetic predisposition and environmental factors, of which dietary intake is an important modifiable risk factor.2 In recent years, advanced studies of genetics have given rise to genetic risk scores (GRSs), which represent a summary of an individual’s genetic susceptibility to various phenotypes, such as obesity.3 The GRS represents a predictive genetic model that can be calculated by including all identified risk alleles associated with the investigated phenotype and creating a score.4 The GRS can be calculated through two methods: a weighted method, which considers the reported magnitudes of effect for the chosen alleles and may be adjusted for the total quantity of risk alleles and their respective effect sizes assessed, or an unweighted approach, which simply involves the summation of the number of risks the person carries in their genetic makeup.4 Concurrently, research has increasingly recognized that the relationship between dietary intake and obesity is not straightforward, suggesting that genetic factors may modulate the effects of dietary intake on weight status. Genome-wide association studies (GWAS) have identified numerous genetic loci linked to body mass index (BMI). Nevertheless, these associations can account for only approximately 3–5% of the BMI variation observed in the general population.2,5–8
Dietary intake is a central determinant of energy balance and, consequently, weight status. The distinction between the impact of a comprehensive dietary pattern and the effects of individual foods or nutrients is a noteworthy topic among researchers. Whole diets include the holistic consumption of various foods and nutrients, which reflects the complex interplay of multiple dietary components.9 This, in turn, can demonstrate the synergistic effects that influence health outcomes in ways that single foods or nutrients may not fully elucidate.9 In contrast, investigating the effects of individual foods or nutrients refers to a more targeted examination into their specific effects on health. Nonetheless, this approach is valuable for understanding the effect of isolated single dietary components, which enables researchers to mechanisms of action and assess dietary sufficiency.9 Understanding whether specific dietary patterns and/or foods mitigate genetic predisposition to obesity is crucial for tailoring effective strategies for obesity prevention and management. This systematic review aims to explore the complex relationship between GRSs, dietary intake, and weight status, with a primary focus on understanding how different dietary interventions and/or intakes impact the weight status of individuals with varying GRSs.
Methods
Registration of Protocol and Reporting
The registration of the review protocol on PROSPERO (Record ID: CRD42023452734; https://www.crd.york.ac.uk/prospero/) was completed before finalizing data extraction. Essential details such as the review title, timeline, team composition, methodologies, and general particulars were uploaded onto the PROSPERO register.
Literature Searches
The Preferred Reporting of Systematic Reviews and Meta-Analyses (PRISMA) statement served as the basic guideline followed in this systematic review. Relevant articles were obtained by conducting an extensive search in online databases, including PubMed, Embase, Science Direct, and Scopus, from their respective index dates until August 2023. An additional free-hand search was conducted to ensure the collection of all relevant articles. All possible combinations of the following MeSH terms and test keywords were used to conduct the search: ”BMI” OR “body mass index” AND “GRS” OR “genetic risk score” AND “dietary intake” OR “dietary pattern” AND “obesity” OR ”weight change” OR “overweight” OR “body weight”. Retrieved articles were then imported into Endnote, where duplicates and articles written in non-English languages were excluded. An initial screening was then performed, and articles were evaluated based on the inclusion criteria to then be either included or excluded from this systematic review.
Eligibility Criteria
Scholarly articles were included in the systematic review if they met the following inclusion criteria: adult individuals of any gender; available BMI GRS; studies investigating the impact of different dietary interventions on weight status; studies that compare the effects of various dietary interventions on weight status in individuals with varying GRSs; studies that report quantitative data on weight status outcomes; randomized controlled trials, prospective cohort studies, cross-sectional studies, and case‒control studies. On the other hand, articles were excluded if they included individuals with health conditions that may influence weight status independently of GRSs and dietary interventions; exclusively included pediatric or geriatric populations; solely focused on pharmacological or surgical interventions without a significant dietary component; did not assess dietary interventions or did not provide sufficient information on the dietary intervention; did not report weight-related outcomes or have inadequate reporting of these outcomes; did not test the interaction between GRS, dietary intervention and obesity outcomes, animal studies, reviews, editorials, commentaries, and opinion pieces; and studies not published in English.
Study Selection
Screening of the retrieved articles from the comprehensive search was conducted using Rayyan (https://rayyan.ai/). The filtration and screening process was completed by two independent researchers (SS and HA), and any disagreements were resolved by consensus. Following the removal of duplicates, titles and abstracts of studies were screened and classified as included, excluded or possibly through Rayyan. Studies classified as possible were accessed in full text to decide their inclusion or exclusion. Any study not following our inclusion criteria was excluded. At the end of the screening, the full text of the included articles was obtained to retrieve pertinent information. The PRISMA flow chart demonstrating the details of the screening and selection process is shown in Figure 1.
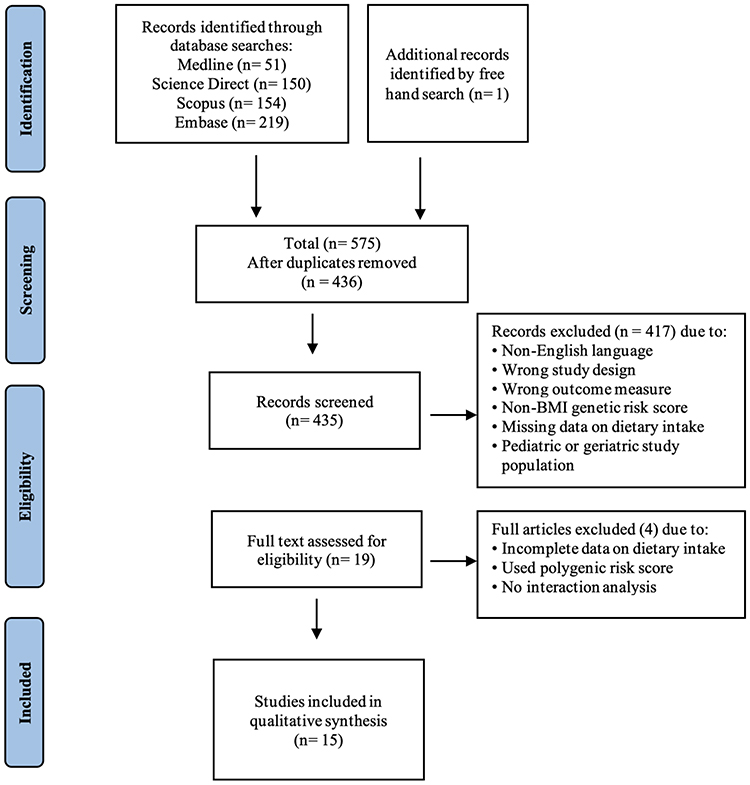 |
Figure 1 PRISMA flow chart for the study selection process. |
Data Extraction
The data extraction of the included articles was conducted independently by two authors (SS and HA), and a discussion took place to finalize the extracted data, including information on the study characteristics, which were author details, year of publication, study design, country, study population, cohort name, sample size, age range or mean, percentage of females, and main outcomes (Table 1). We also extracted information regarding the number of SNPs selected in the study; GRS computation method; GRS construction criteria; dietary intervention; dietary assessment method and duration; association analysis model used; nongenetic covariates the model was adjusted for; and main study findings with their respective p values (Table 2).
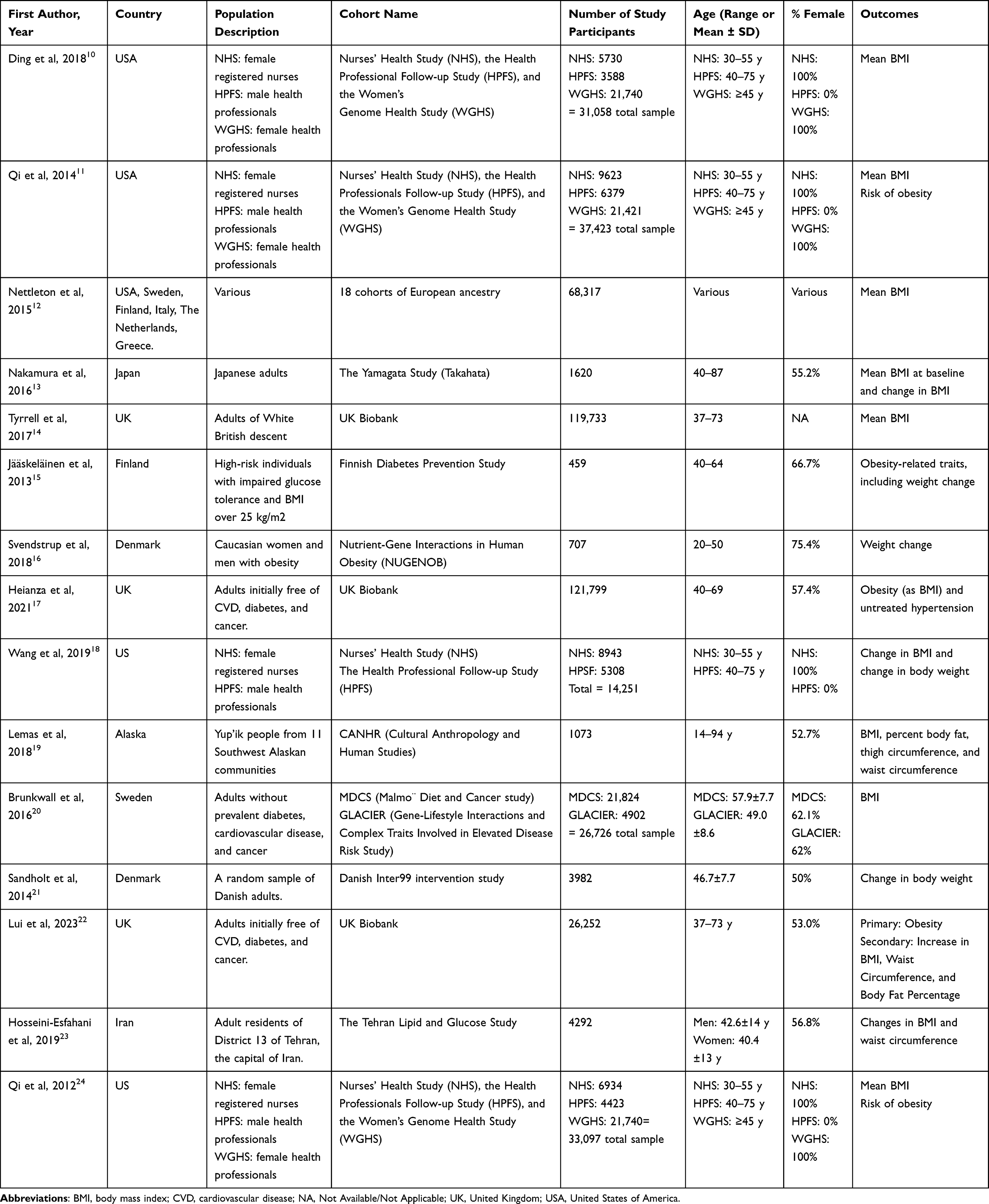 |
Table 1 Description of the Characteristics of Studies Included in the Systematic Review |
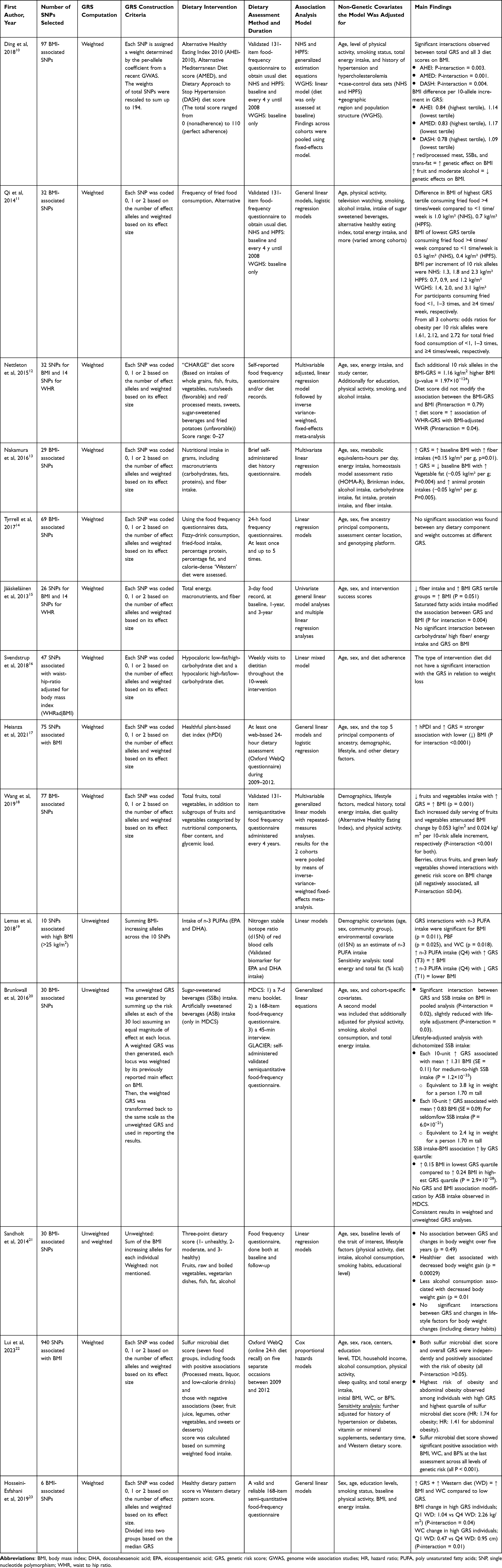 |
Table 2 Description of the Utilized Genetic Risk Score, Dietary Intakes, and Main Findings |
Quality Assessment
Two authors conducted the quality assessment of the included articles (SS and HA). Since all included studies were cross-sectional in design, the Newcastle‒Ottawa Scale (NOS) tool25 was used for quality assessment. The NOS tool includes three criteria for evaluation, which are selection, comparability, and outcome, each of which can be given a maximum of two points except selection, which can be given a maximum of three points. The quality of the study is indicated by the given number of stars in the NOS tool, where 6–7 stars indicate high quality, 3–5 stars indicate moderate quality, and 0–2 stars indicate poor quality. The details of the quality assessment are shown in Table 1.
Results
Search Outcome
The results of the initial screening of abstracts and of full texts with reasons for exclusion are shown in the PRISMA flow chart in Figure 1. The extensive search of multiple databases resulted in the identification of 575 records. An additional study was identified although a free-hand search. After removal of duplicates, 436 records remained and were evaluated against our inclusion criteria, which resulted in the exclusion of 417 records. The remaining 19 studies were assessed by their full texts, resulting in the exclusion of 3 articles for either using a polygenic risk score, having incomplete or missing data on dietary intake, or not conducting an interaction analysis. This resulted in a total of 15 studies being included in the qualitative synthesis of this review, all of which were cross-sectional studies.
Quality of Selected Studies
The quality assessment of included studies, shown in Table 3, was completed using the NOS tool modified for cross-sectional design.25 Out of the 15 studies, only one study received the full 7 stars, indicating a high quality and low risk of bias.17 Additionally, ten studies received a 6-star rating, also indicating high quality and low risk of bias in these studies.12–16,19–23 The remaining four studies received an overall rating of 5 stars, indicating a moderate quality and moderate risk of bias.10,11,18,24 The quality of the included studies was high overall, and the risk of bias was low. Most studies lost stars due to not reporting the characteristics of nonrespondents, which is not crucial for our systematic review, as most studies included participants with available data from large cohorts to conduct analyses.
 |
Table 3 Quality Assessment of Included Studies Using Newcastle-Ottawa Quality Assessment Scale Criteria for Cross-Sectional Studies |
Characteristics of Included Studies
The key characteristics of the included studies are shown in Table 2. The studies are published over the period of 11 years, between 2012 and 2023, with the majority being published between 2013 and 2019 (80%). The studies were conducted on various study populations; 4 studies included cohorts from the US, while seven others included cohorts from European populations such as the UK, Sweden, Denmark, and Finland. The remaining studies were conducted in Japan (n=1), Alaska (n=1), Iran (n=1), and one large study including populations from the US, Sweden, Finland, Italy, The Netherlands, and Greece. Most studies included a large number of participants, ranging from 84 to 121,799 participants. The ages of the participants were mostly in their 30s and 40s, and only one study19 included participants aged 14 to 94 years. Participants were mainly healthy individuals free of disease, but one study included overweight people with impaired glucose tolerance.15 The main outcomes of the included studies were mostly mean BMI or other body fatness indicators, such as risk of obesity, weight change, BMI change, percent body fat (%BF), thigh circumference, waist circumference (WC), total abdominal fat, visceral fat, and subcutaneous fat.
Genetic Risk Score Characteristics
The number of SNPs used in the generation of the GRSs ranged from 6 to 940 SNPs. To include the genetic variation in the prediction models, the weighted method was used by the majority of the studies (n= 12), and the remaining studies used the unweighted method (n= 1) or both methods (n= 2). Most of the included studies selected the SNPs they included from previously published genome-wide association studies (GWAS) or from other published association studies that showed a high correlation between chosen SNPs and BMI.
The Influence of Comprehensive Dietary Interventions
The influence of dietary interventions was assessed in the included studies in two different approaches; 11 studies tested the effect of dietary patterns or whole diet interventions out of the 15 studies included in this systematic review. Ding et al10 obtained dietary information using a validated food frequency questionnaire (FFQ) from 31,058 participants living in the US. The authors generated scores for the Alternative Healthy Eating Index 2010 (AHEI), Alternative Mediterranean Diet (AMED), and Dietary Approach to Stop Hypertension (DASH). The weighted GRS used was computed based on 97 BMI-associated SNPs. The study found that the difference in BMI per 10-allele increment of the GRS was smaller among participants with higher diet scores, reflecting higher adherence, than among those with lower scores. This indicates that adhering to healthier diets may attenuate the genetic predisposition to body adiposity. This was consistent for the three cohorts included in the study (NHS, HPFS, and WGHS). Another study by Nettleton et al12 assessed the healthfulness of dietary intake of participants in 18 cohorts by the “CHARGE diet” score, where a higher score reflects a healthier eating pattern. The study included 68,317 participants of European ancestry and constructed a weighted GRS based on 32 SNPs associated with BMI and 14 SNPs associated with waist-to-hip ratio (WHR). The results showed that a higher GRS resulted in a higher mean BMI (1.16 kg/m2 increase for each additional 10-risk allele); however, CHARGE diet scores did not modify this association (Pinteraction = 0.79). In the Danish population, 707 Caucasian participants in the Nutrient-Gene Interactions in Human Obesity (NUGENOB) cohort were assigned to either a hypocaloric low-fat/high-carbohydrate diet or a hypocaloric high-fat/low-carbohydrate diet.16 The weighted GRS used was based on 47 SNPs associated with WHR adjusted for BMI. Adhering to either of the diets did not have a significant interaction with the GRS in relation to weight change/loss. Additionally, in the Danish population, Sandholt et al21 included a random sample of 3982 healthy adults from the Danish Inter99 intervention study and obtained their dietary information using an FFQ at baseline and at follow-up. The GRS used was computed as both weighted and unweighted and was based on 30 BMI-associated SNPs. A three-point dietary score (1- unhealthy, 2- moderate, and 3- healthy) was generated. No interaction was observed between GRS and any lifestyle change, including dietary intake, in relation to body weight change. A similar diet score was generated by Hosseini et al23 using semiqualitative FFQ data for 4292 Iranian adults. However, the participants’ intake was categorized into either a healthy dietary pattern or a Western dietary pattern score. The GRS used was based on 6 SNPs associated with BMI. The study found that individuals with a higher genetic predisposition to obesity and consuming a Western diet experienced higher BMI and waist circumference (WC) changes at follow-up than individuals with a lower GRS. Specifically, individuals with high GRS who were in the first quartile of the Western diet score had a higher BMI by 1.04 kg/m2, while those in the fourth quartile had a 2.26 kg/m2 higher BMI (Pinteraction = 0.04). Similarly, WC increased by 0.47 cm vs 0.95 cm in individuals in the Q1 and Q4 western diet scores, respectively, and these individuals are highly genetically predisposed to obesity (Pinteraction = 0.01).23 Thus far, the results are pointing at either the weight gain accelerating effect or the absence of an effect of adhering to an unhealthy eating pattern such as a Western diet while having high genetic susceptibility to obesity. To investigate the effect of following a healthy eating pattern such as a healthy plant-based diet, Heianza et al17 used the genetic, anthropometric and dietary information of 121,799 participants in the UK Biobank cohort to investigate this interaction. The weighted GRS computed in this study was based on 75 BMI-associated SNPs. The study found that a higher healthy plant-based diet index and having high GRS increased the strength of association with a lower BMI (Pinteraction < 0.0001). An interesting study on the same British cohort investigated the influence of adhering to a sulfur microbial diet on obesity, BMI, WC, and body fat percentage (BF%) at varying GRSs.22 The analysis revealed that the highest risk of obesity and high abdominal obesity was observed among individuals with high GRS and the highest quartile of sulfur microbial diet score (HR: 1.74 for obesity; HR: 1.41 for abdominal obesity). In addition, regardless of the GRS, a high sulfur microbial diet score was significantly associated with higher BMI, WC, and BF% (all p < 0.001).
The Influence of Individual Foods or Nutrients
Several studies have investigated the influence of single foods or specific macronutrients on obesity outcomes in individuals with varying genetic susceptibilities, such as sugar-sweetened beverages, fried foods, fruits and vegetables, fiber, and n-3 polyunsaturated fatty acids. In the Finnish population, Jääskeläinen el al. included 459 overweight participants with impaired glucose tolerance who had records of their dietary intake for 3 days at baseline and at the 1-year and 3-year follow-ups.15 The study computed a weighted GRS using 26 SNPs for BMI and 14 SNPs for WHR. The results showed that participants with low fiber intake and high GRS reported high BMI (p = 0.051), and saturated fatty acid intake significantly interacted with GRS and BMI (Pinteraction = 0.004). Meanwhile, consuming high or low carbohydrate, high fiber, and high or low energy intakes did not modify the association between GRS and BMI.15 Among the Japanese population, Nakamura et al13 found that a higher GRS was associated with a higher baseline BMI among participants with high fiber intake (0.15 kg/m² per g, p=0.01). On the other hand, a higher intake of vegetable fat and animal protein was associated with a lower baseline BMI and a higher genetic predisposition to obesity (−0.05 kg/m² per g; P=0.004 and −0.05 kg/m² per g; P=0.005, respectively). These results are controversial, as fiber intake is mainly associated with health-promoting effects and lower body weight through multiple mechanisms. Meanwhile, saturated fatty acids are considered harmful and cause weight gain and cardiovascular diseases when consumed in excess. Other results from this study were discrepant with established and previous knowledge, suggesting a potential uncontrolled confounder in the study. The authors computed a weighted GRS based on 29 BMI-associated SNPs. Qi et al24 conducted one of the oldest studies investigating the effect of dietary intake on obesity outcomes at varying BMI GRSs. The study included 33,097 individuals from three large US cohorts, the Nurses’ Health Study (NHS), the Health Professionals Follow-up Study (HPFS), and the Women’s Genome Health Study (WGHS), with available data on their sugar-sweetened and artificially sweetened beverage consumption levels (SSB and ASB). Notably, NHS and WGHS are cohorts exclusively composed of females, while HPFS is a cohort exclusively composed of males. The weighted GRS in this study was computed based on 32 SNPs associated with BMI. After pooling the results from the three cohorts, the study found that for each 10 risk alleles, the increment increase in BMI was higher by 1.00, 1.20, 1.37, and 1.85 kg/m2 for individuals consuming SSB at <1 serving/month and 1–4 serving/month, respectively. 2–6 servings/week, and >1 serving/day, respectively (Pinteraction <0.001). Since fat-mass and obesity (FTO)-related gene loci are well known to have a strong effect on BMI, the investigators performed a sensitivity analysis to ensure that the results were not derived only from the genetic effect of this gene and obtained similarly strong results (Pinteraction <0.001 in the pooled data). On the other hand, consumption of ASB did not influence the association between GRS and mean BMI. In addition, the pooled results from the 3 cohorts showed that consuming SSB <1 serving/month, 1–4 serving/month. 2–6 servings/week, and >1 serving/day increased the risk for obesity incidence by 1.35, 1.59, 1.56, and 3.35, respectively (Pinteraction < 0.001). The study also showed that one daily serving of SSB consistently increased the BMI across the quartiles of GRS in the three cohorts. The pooled results showed the strongest association in individuals in the highest GRS quartile (β coefficient = 0.44, P = 0.003) compared to the lowest GRS quartile (β coefficient = −0.07, P = 0.36) (Pinteraction <0.001). The same group of researchers investigated the effect of the frequency of fried food (FF) consumption on obesity outcomes using the same 32 BMI-associated SNPs to calculate a weighted GRS.11 The investigators found that among individuals with the highest GRS tertile, the difference in mean BMI between people consuming FF more than 4 times per week and people consuming less than once a week was 1.0 kg/m² in the NHS cohort and 0.7 kg/m² in the HPFS cohort. These results were also significant in the WGHS cohort. In addition, the association between FF and BMI became stronger with each additional 10-risk allele in the GRS, and BMI increased by 1.1, 1.6, and 2.2 kg/m2 for total FF consumption <1 time, 1–3 times, and ≥4 times/week, respectively, in the pooled cohorts. Finally, the researchers found an increased odds ratio (95% confidence interval) for obesity as the GRS increased in 10-risk allele increments. Total FF consumption of <1, 1–3, and ≥4 times/week was associated with 1.61 (1.40–1.87), 2.12 (1.73–2.59), and 2.72 (2.12–3.48) increased odds for obesity, respectively.11 Brunkwall et al20 sought to replicate the study by Qi et al among 26,726 individuals from two Swedish cohorts. The authors computed both a weighted and unweighted GRS based on 30 SNPs associated with BMI. Similarly, the study found a significant interaction between SSB intake and GRS on BMI outcomes in the pooled analysis (Pinteraction = 0.02). When SSB intake was categorized into either seldom to low or medium to high intake, each 10-risk allele increase in the GRS corresponded to a mean BMI increase of 1.31 kg/m2 for medium-to-high SSB intake (p = 1.2×10−33) and 0.83 kg/m2 for seldom/low intake (p = 6.0×10−21). This is equivalent to 3.8 kg and 2.4 kg in weight for a person 1.70 m tall, respectively. In addition, the association between SSB intake and BMI became stronger at higher GRS quartiles, where 1 increment of SSB intake caused a 0.15 kg/m2 (95% CI: 0.08, 0.24) increase in BMI for individuals in the lowest quartile of GRS compared to 0.24 kg/m2 (95% CI: 0.15, 0.32) for individuals in the highest quartile, according to the pooled analysis (all p < 0.01). Artificially sweetened beverage intake was assessed in one of the cohorts only (ie, Malmo¨ Diet and Cancer Study), and it did not affect the association between GRS and BMI outcome.20 An interesting study by Lemas et al19 aimed to assess the influence of n-3 polyunsaturated fatty acids (PUFAs) on body fatness indicators at varying GRSs for BMI. The study included 1073 Yup’ik adults, one of the indigenous groups in southwestern Alaska, and n-3 PUFA intake was assessed using the nitrogen stable isotope ratio (δ15 N) of red blood cells, which is a validated biomarker for EPA (eicosapentaenoic acid) and DHA (docosahexaenoic acid). The fatty acids EPA and DHA are two specific types of n-3 PUFA involved in multiple anti-inflammatory and developmental functions. The analysis used an unweighted GRS based on 10 SNPs associated with high BMI (>25 kg/m2) and found a significant interaction between GRS, n-3 PUFAs and BMI (p = 0.011), percentage body fat (p = 0.025), and WC (p = 0.018). Statistical analysis also showed that individual intake of n-3 PUFAs significantly modified the association between GRS and BMI outcome. At high n-3 PUFA intake levels (Q4), individuals with high GRS (T3) had the highest BMI (~ 34.8 kg/m2), while high n-3 PUFA intake levels (Q4), while having low GRS levels, resulted in lower BMI outcomes (~ 31 kg/m2). These associations remained consistent even after adjusting for total fat intake or total energy intake in a sensitivity analysis. To assess the effect of healthy dietary components, Wang et al18 used two US cohorts (NHS and HPFS) and included 14,251 total participants to assess the effect of fruit and vegetable consumption on BMI change and body weight change. The weighted GRS was computed based on 77 BMI-associated SNPs derived from a genome-wide association study (GWAS). The results from the combined cohorts showed that individuals with the highest genetic risk and who consumed low fruits and vegetables showed the highest increase in BMI every 4 years over a 20-year follow-up period. In addition, individuals with high GRS were most sensitive to the influence of fruit and vegetable intake; as a result, people with the highest fruit and vegetable intake experienced a decrease in BMI or no weight gain over the follow-up period. Specifically, per 10-risk allele increment, each additional daily serving of fruits attenuated 0.053 kg/m2 of BMI change, while an extra daily serving of vegetables attenuated 0.024 kg/m2 of BMI change, corresponding to 0.13 kg and 0.05 kg weight loss. (Pinteraction <0.001 for both). Analysis of certain groups of fruits and vegetables revealed that berries, citrus fruits, and green leafy vegetables significantly interacted with genetic risk score on BMI change, all of which were negatively associated with BMI change (all Pinteraction ≤0.005). Last, Tyrell et al14 assessed the influence of 12 measures of an obesogenic environment, including frizzy drink consumption, fried food intake, percentage protein, percentage fat, and a calorie-dense Western diet score, on weight outcomes at different GRSs. The study used a weighted GRS computed based on 69 BMI-associated SNPs. The analysis included 119,733 British adults from the UK Biobank and revealed no significant association between any dietary component and mean BMI at various GRSs.
Discussion
Given the global obesity epidemic and the growing interest in personalized medicine approaches, there is a pressing need to comprehensively evaluate the interplay between GRSs and dietary intake in shaping weight status. This systematic review demonstrated the influence of dietary patterns and individual foods or macronutrients. The genetic predisposition to higher BMI and obesity was exacerbated by following certain unhealthy eating patterns and individual foods. The differential effect was significant in multiple studies, and individuals genetically predisposed to obesity were affected more severely by these unhealthy eating patterns. This effect was observed after consuming a western diet23 and a sulfur microbial diet.22 The Western diet is characterized by a high intake of processed and fast foods rich in sugars, unhealthy fats, and red meats. It typically includes low consumption of fruits, vegetables, whole grains, and dietary fiber.26 The link between this dietary pattern and health issues such as obesity, cardiovascular diseases, and diabetes is well established.26 A sulfur microbial diet is composed of foods that promote the production of sulfur by sulfur-reducing bacteria present in the human gut microbiome. Examples of these foods are red meats, seafood, garlic, onions, leeks, and cruciferous vegetables, which are rich in sulfur-containing amino acids.27 This diet is biologically known to potentially induce obesity due to high production of sulfur, which disrupts gut permeability, allows more nutrients to pass into the bloodstream and therefore more energy uptake, and promotes absorption of lipopolysaccharides into the bloodstream, which further worsens body inflammation.28,29 On the other hand, following a healthy eating pattern attenuated the impact of genetic predisposition in multiple studies, such as following AHEI, AMED and DASH diets10 and consuming a healthy plant-based diet.17
Individual foods were also found to have a particular weight-gaining effect on highly genetically predisposed individuals compared to those less genetically predisposed. This includes the consumption of sugar-sweetened beverages20,24 and fried foods,11 both of which increased BMI in a dose-dependent manner. In addition, fiber,13,15 saturated fatty acids,15 animal protein and vegetable fat13 showed conflicting results in the two studies. In addition, n-3 PUFAs were found to have an anti-obesogenic effect in individuals with a low genetic predisposition to obesity, and this association was determined to be derived mainly from n-3 PUFA intake rather than the consumption of a traditional dietary pattern high in fat and/or calories.19 Consuming fruits and vegetables was also shown to attenuate weight gain in individuals with high GRS.18 In contrast, no influence was observed for the consumption of artificially sweetened beverages on body weight status at varying GRSs in the American population24 or in the Swedish population.20 Additionally, energy and carbohydrate intake did not affect the association between GRS and BMI.15 Multiple studies testing the influence of dietary patterns found no interaction between genetic predisposition and body weight outcomes.12,16,21
It is important to point out that systematic errors are specifically relevant biases in obesity-related research. These biases can occur when individuals either overreport or underreport their dietary intake, especially in overweight or underweight participants. This bias may also arise due to participants often being well informed about the connections between lifestyle choices and body weight, which could influence how they respond to questions about their habits. All studies included in the systematic review used data from large cohorts, and evaluating lifestyle accurately in large study populations is challenging, but differentiating the effect of perceived lifestyle factors from the unmeasured factors associated with genetic susceptibility is another important challenge yet to be overcome. These aspects warrant using more precise tools to assess diet, especially in gene-diet interaction studies. In addition, possible reverse causation is that food-related behavioral responses such as appetite, energy intake levels and macronutrient preferences may be influenced by adiposity-associated genetic variants.30–32 Accordingly, food preferences have been recently shown to be partially genetically determined, such as a high consumption of sugar and carbohydrates.33,34 The findings of this research suggest the potential application of the GRS in preventive medicine. Integrating the domain of gene‒environment interactions in preventive medicine involves assessing how genetic factors alter the impact of lifestyle elements on obesity within intervention strategies.35,36
Strengths and Limitations
There are multiple strengths in this systematic review. To our knowledge, this is the first systematic review to evaluate the interaction between GRS and dietary intake on weight status and summarize almost all the available evidence regarding this association. In addition, a rigorous systematic approach was followed to develop this review, such as the PRISMA guidelines and the Rayyan application, which was used to perform initial screening and enabled the research team to conduct blind review that would enhance the credibility of the findings. Additionally, the studies included were conducted on a large number of participants, which were relatively homogenous for their age (30s and 40s) and their health status (all were healthy at baseline except one study). Additionally, the studies included in this systematic review were conducted in various countries, which allows us to see the influence of dietary intake on body weight in individuals with various genetic backgrounds. Furthermore, the quality of the included studies was mainly high, and only four out of fifteen studies were of moderate quality, which indicates that the evidence drawn from these studies is unlikely to be biased or the result of other uncontrolled confounding factors. On the other hand, this systematic review has a few limitations, such as including only cross-sectional studies, which are observational in design. This gives rise to the need for clinical controlled trials with strong methodologies to be able to extract robust conclusions about the association between GRS and dietary intake and their effect on weight outcomes. Moreover, selected studies were heterogeneous for the number of selected SNPs and method of GRS calculation. Finally, our review exclusively incorporated English-language articles while excluding all papers written in foreign languages, thus limiting the available evidence to English literature only.
Conclusion
This systematic review explored the interplay between GRSs, dietary intake, and weight outcomes, and the synthesis of evidence from diverse studies revealed that dietary patterns and individual foods exert differential effects on individuals with varying genetic predispositions to higher BMI and obesity. Unhealthy dietary patterns such as Western diet and sulfur microbial diet exacerbated genetic predisposition to obesity and accelerated weight gain, while healthy diets and individual foods known to promote health mitigate the impact of genetics, including fruits, vegetables and n-3 PUFA. Multiple limitations related to study designs, collection of dietary data and possible systematic error warrant more controlled clinical trials with large sample sizes to be able to draw robust conclusions. Therefore, personalized nutrition and nutrigenetics can be incorporated in the design of interventions for obesity prevention and management.
Acknowledgement
We would like to thank Qatar University as the APC fund was provided with the Qatar University grant: QUST-1-CHS-2024-1676.
Disclosure
All authors declare no conflicts of interest in this work.
References
1. Lin X, Li H. Obesity: epidemiology, pathophysiology, and therapeutics. Front Endocrinol. 2021;12:706978. doi:10.3389/fendo.2021.706978
2. Locke AE, Kahali B, Berndt SI, et al. Genetic studies of body mass index yield new insights for obesity biology. Nature. 2015;518(7538):197–206. doi:10.1038/nature14177
3. Seral-Cortes M, Sabroso-Lasa S, De Miguel-Etayo P, et al. Development of a genetic risk score to predict the risk of overweight and obesity in European adolescents from the HELENA study. Sci Rep. 2021;11(1):3067. doi:10.1038/s41598-021-82712-4
4. Igo RP Jr, Kinzy TG, Cooke Bailey JN. Genetic risk scores. Curr Protoc Hum Genet. 2019;104(1):e95. doi:10.1002/cphg.95
5. Srivastava A, Srivastava N, Mittal B. Genetics of obesity. Indian J Clin Biochem. 2016;31(4):361–371. doi:10.1007/s12291-015-0541-x
6. Wang K, Li WD, Zhang CK, et al. A genome-wide association study on obesity and obesity-related traits. PLoS One. 2011;6(4):e18939. doi:10.1371/journal.pone.0018939
7. Yengo L, Sidorenko J, Kemper KE, et al. Meta-analysis of genome-wide association studies for height and body mass index in ∼700000 individuals of European ancestry. Hum Mol Genet. 2018;27(20):3641–3649. doi:10.1093/hmg/ddy271
8. Speliotes EK, Willer CJ, Berndt SI, et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat Genet. 2010;42(11):937–948. doi:10.1038/ng.686
9. Tapsell LC, Neale EP, Satija A, et al. Foods, nutrients, and dietary patterns: interconnections and implications for dietary guidelines. Adv Nutr. 2016;7(3):445–454. doi:10.3945/an.115.011718
10. Ding M, Ellervik C, Huang T, et al. Diet quality and genetic association with body mass index: results from 3 observational studies. Am J Clin Nut. 2018;108(6):1291–1300. doi:10.1093/ajcn/nqy203
11. Qi Q, Chu AY, Kang JH, et al. Fried food consumption, genetic risk, and body mass index: gene-diet interaction analysis in three US cohort studies. BMJ. 2014;348(mar19 1):g1610–g1610. doi:10.1136/bmj.g1610
12. Nettleton JA, Follis JL, Ngwa JS, et al. Gene × dietary pattern interactions in obesity: analysis of up to 68 317 adults of European ancestry. Human Molecular Genetics. 2015;24(16):4728–4738. doi:10.1093/hmg/ddv186
13. Nakamura S, Narimatsu H, Sato H, et al. Gene–environment interactions in obesity: implication for future applications in preventive medicine. J Hum Genet. 2016;61(4):317–322. doi:10.1038/jhg.2015.148
14. Tyrrell J, Wood AR, Ames RM, et al. Gene-obesogenic environment interactions in the UK Biobank study. Int J Epidemiol. 2017;46(2):559–575. doi:10.1093/ije/dyw337
15. Jääskeläinen T, Paananen J, Lindström J, et al. Genetic predisposition to obesity and lifestyle factors – the combined analyses of twenty-six known BMI- and fourteen known Waist: Hip Ratio (WHR)-associated variants in the Finnish Diabetes Prevention Study. Br J Nutr. 2013;110(10):1856–1865. doi:10.1017/S0007114513001116
16. Svendstrup M, Allin KH, Sørensen TIA, et al. Genetic risk scores for body fat distribution attenuate weight loss in women during dietary intervention. Int J Obesity. 2018;42(3):370–375. doi:10.1038/ijo.2017.279
17. Heianza Y, Zhou T, Sun D, et al. Healthful plant-based dietary patterns, genetic risk of obesity, and cardiovascular risk in the UK biobank study. Clin Nutr. 2021;40(7):4694–4701. doi:10.1016/j.clnu.2021.06.018
18. Wang T, Heianza Y, Sun D, et al. Improving fruit and vegetable intake attenuates the genetic association with long-term weight gain. Am J Clin Nut. 2019;110(3):759–768. doi:10.1093/ajcn/nqz136
19. Lemas DJ, Klimentidis YC, Wiener HH, et al. Obesity polymorphisms identified in genome-wide association studies interact with n-3 polyunsaturated fatty acid intake and modify the genetic association with adiposity phenotypes in Yup’ik people. Genes Nutr. 2013;8(5):495–505. doi:10.1007/s12263-013-0340-z
20. Brunkwall L, Chen Y, Hindy G, et al. Sugar-sweetened beverage consumption and genetic predisposition to obesity in 2 Swedish cohorts. Am J Clin Nut. 2016;104(3):809–815. doi:10.3945/ajcn.115.126052
21. Sandholt CH, Allin KH, Toft U, et al. The effect of GWAS identified BMI loci on changes in body weight among middle-aged danes during a five-year period. Obesity. 2014;22(3):901–908. doi:10.1002/oby.20540
22. Liu X, Wan X, Zhang L, et al. The sulfur microbial diet and increased risk of obesity: findings from a population-based prospective cohort study. Clin Nutr. 2023;42(5):764–772. doi:10.1016/j.clnu.2023.03.011
23. Hosseini-Esfahani F, Koochakpoor G, Mirmiran P, et al. Dietary patterns modify the association between fat mass and obesity-associated genetic variants and changes in obesity phenotypes. Br J Nutr. 2019;121(11):1247–1254. doi:10.1017/S0007114519000643
24. Qi Q, Chu AY, Kang JH, et al. Sugar-sweetened beverages and genetic risk of obesity. N Engl J Med. 2012;367(15):1387–1396. doi:10.1056/NEJMoa1203039
25. Stang A. Critical evaluation of the Newcastle-Ottawa scale for the assessment of the quality of nonrandomized studies in meta-analyses. Eur J Epidemiol. 2010;25(9):603–605. doi:10.1007/s10654-010-9491-z
26. Clemente-Suárez VJ, Beltrán-Velasco AI, Redondo-Flórez L, et al. Global impacts of western diet and its effects on metabolism and health: a narrative review. Nutrients. 2023;15(12):1.
27. Wang Y, Nguyen LH, Mehta RS, et al. Association between the sulfur microbial diet and risk of colorectal cancer. JAMA Network Open. 2021;4(11):e2134308. doi:10.1001/jamanetworkopen.2021.34308
28. Blachier F, Andriamihaja M, Larraufie P, et al. Production of hydrogen sulfide by the intestinal microbiota and epithelial cells and consequences for the colonic and rectal mucosa. Am J Physiol Gastrointest Liver Physiol. 2021;320(2):G125–G135. doi:10.1152/ajpgi.00261.2020
29. Ijssennagger N, Belzer C, Hooiveld GJ, et al. Gut microbiota facilitates dietary heme-induced epithelial hyperproliferation by opening the mucus barrier in colon. Proc Natl Acad Sci U S A. 2015;112(32):10038–10043. doi:10.1073/pnas.1507645112
30. Chu AY, Workalemahu T, Paynter NP, et al. Novel locus including FGF21 is associated with dietary macronutrient intake. Hum Mol Genet. 2013;22(9):1895–1902. doi:10.1093/hmg/ddt032
31. Kilpeläinen TO, Carli JF, Skowronski AA, et al. Genome-wide meta-analysis uncovers novel loci influencing circulating leptin levels. Nat Commun. 2016;7:10494.
32. Tanaka T, Ngwa JS, van Rooij FJ, et al. Genome-wide meta-analysis of observational studies shows common genetic variants associated with macronutrient intake. Am J Clin Nutr. 2013;97(6):1395–1402. doi:10.3945/ajcn.112.052183
33. Treur JL, Boomsma DI, Ligthart L, et al. Heritability of high sugar consumption through drinks and the genetic correlation with substance use. Am J Clin Nut. 2016;104(4):1144–1150. doi:10.3945/ajcn.115.127324
34. Pallister T, Sharafi M, Lachance G, et al. Food preference patterns in a UK twin cohort. Twin Res Hum Genet. 2015;18(6):793–805. doi:10.1017/thg.2015.69
35. Marti A, Martinez-González MA, Martinez JA. Interaction between genes and lifestyle factors on obesity. Proc Nutr Soc. 2008;67(1):1–8. doi:10.1017/S002966510800596X
36. Cooper RS. Gene–environment interactions and the etiology of common complex disease. Ann Intern Med. 2003;139(5_Part_2):437–440. doi:10.7326/0003-4819-139-5_Part_2-200309021-00011
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.