")
Back to Journals » OncoTargets and Therapy » Volume 13
The Effects of TRAF6 on Growth and Progression in Colorectal Cancer are Regulated by miRNA-140
Authors Zhu G , Cheng Z, Lin C, Wang Q, Huang Y, Zheng W, Yang S, Ye J
Received 16 April 2020
Accepted for publication 22 September 2020
Published 20 November 2020 Volume 2020:13 Pages 11991—12001
DOI https://doi.org/10.2147/OTT.S257733
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Takuya Aoki
Guangwei Zhu,1,* Zhibin Cheng,1,2,* Chunlin Lin,1 Qin Wang,1,2 Yongjian Huang,1 Wei Zheng,1 Shugang Yang,1 Jianxin Ye1
1Department of Gastrointestinal Surgery 2 Section, The First Hospital Affiliated to Fujian Medical University, Fuzhou 350005, People’s Republic of China; 2Key Laboratory of Ministry of Education for Gastrointestinal Cancer, Fujian Medical University, Fuzhou 350000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jianxin Ye Email [email protected]
Background and Aim: Some studies have confirmed that miRNA-140 exhibits a suppressive role in gastric cancer, Wilms’ tumor. However, the function of miRNA-140 in colorectal cancer has not been completely elucidated. The present study aims to verify TRAF6 as the targeted gene by miRNA-140 which was investigated in colorectal cancer tissues and cells, and its effects on the biological characteristics of colorectal cancer cells were determined, in order to provide an experimental and theoretical basis for the application of TRAF6 in the treatment of colorectal cancer.
Methods: qPCR analyzed miRNA-140 expression levels in colorectal cancer tissues, normal colorectal cancer tissues and colorectal cells including SW480 and HCT116 cancer cells and FHC normal colorectal epithetical cells. A serial biological experiment analyzed miRNA-140 effects on cell proliferation, migration and invasion capacities in SW480 and HCT116 cells. miRNA targeting gene prediction and a dual luciferase assay were used to analyze miRNA-140-targeted TRAF6. qPCR and Western blot analyzed miRNA-140 effects on the mRNA and protein expression of TRAF6. Western blot analyzed miRNA-140 effects on NF-κB/c-jun signaling pathways. Animal studies were performed to investigate the effects of miRNA-140 on colorectal cancer implantation tumor growth. Immunohistochemistry analyzed TRAF6 expression in animal experimentation tumors.
Results: miRNA-140 expression is lower in colorectal cancer tissues and colorectal cancer cells. Over-expression of miRNA-140 inhibited the proliferation, migration and invasion capacities of colorectal cancer cells. miRNA-140 targeted the TRAF6 mRNA 3ʹUTR area and decreased TRAF6 protein expression. miRNA-140 suppressed p-NF-κB/p-c-jun proteins expression. miRNA-140 inhibited colorectal cancer implantation tumor growth in the mice model.
Conclusion: miRNA-140 targeting TRAF6 affects the progression and growth of colorectal cancer, the mechanism could be miRNA-140 decreasing the TRAF6 expression effects on the NF-κB/c-jun signaling pathways.
Keywords: TRAF6, miRNA-140, NF-kB/c-jun signaling pathways, progression, colorectal cancer
Introduction
Colorectal caner (CRC) is the third most commonly diagnosed cancer-related disease and cause of death in the United States.1 The American Cancer Society evaluated that more than 140 thousands new CRC patients will be diagnosed and more than 50 thousands CRC patients will die because of this cancer in 2019.2 Surgical resection is still the most effective and most possible curative treatment for CRC.3,4 However, many CRC patients develop recurrencies. It is necessary to study the molecular mechanisms that effect the growth and progression of CRC, as these may be potential therapeutic targets for CRC treatment.
The tumor necrosis factor receptor-associated factor-6 (TRAF6) family includes a family of seven members TRAF1-7. The TRAFs family play key roles in regulating cell survival and cell death.5,6 TRAF6 has been proved to be involved in cancer progression.7,8 Luo et al7 reported that TRAF6 is involved in regulating melanoma invasion and metastasis. Han et al8 indicated that TRAF6 promotes the invasion and metastasis and predicts a poor prognosis in gastric cancer. TRAF6 can promote the metastasis of esophageal squamous cell carcinoma and tumorigenicity of pancreatic cancer cells.9,10 Our previously study also showed that TRAF6 promotes the progression and growth of colorectal cancer.11 However, the regulation mechanisms of TRAF6 in cancer cells is not fully understood, and why TRAF6 expression is upregulated in colorectal cancer cells remains unclear. Therefore, research regarding the regulation of TRAF6 expression may have vital theoretical significance and practical value.
In recent years, microRNA (miRNA) has been discovered to have the ability of regulating gene expression.12,13 miRNAs are small, single stranded, non-coding RNAs consisting of 20–22 nucleotides, which play significant roles in many biological processes by regulating gene expression at the post-translational level.14 The mechanisms by which miRNAs work is binding to the complementary sites in the 3ʹ untranslated region (3ʹUTR) of the targeting genes which induced degradation of the transcript or translational repression.15 miRNAs are involved in many physiological processes, including cell proliferation, cell apoptosis, cell cycle and cell differentiation.16,17
Studies revealed that miRNA-140 is a tumor suppressor in several types of human cancer.18–20 We carried out bioinformatics analysis using Targetscan and starBase related prediction software.21,22 The bioinformatics analysis showed that TRAF6 was a miRNA-140 targeted gene. Therefore, the present study aimed to determine the role of miRNA-140 regulating TRAF6 in colorectal cancer in vitro and in vivo.
Patients and Methods
Patients with CRC Tissues Specimens
5 pairs of fresh cancer specimens and normal colorectal tissues were collected from patients with CRC by surgical excision. The specimens and tissues were immediately cut into 1 mm3 fragments and then cultured in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) with 10% fetal bovine serum (FBS, Life Technologies), 1% penicillin (Sigma Millipore, Louis, USA) and incubated at 37 °C under 95% air and 5% CO2 for 48 h. All patients provided informed consent and the experiment was conducted in accordance with the Declaration of Helsinki. The present study was approved by the Ethics Committee of The First Hospital Affiliated to Fujian Medical University.
Cell Culture
The CRC cell lines SW480 and HCT116 were purchased from the Shanghai Institute for Biological Science (Shanghai, China). Both cell lines were cultured in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) with 10% fetal bovine serum (FBS, both Life Technologies; Thermo Fisher Scientific, Inc.). The FHC (normal colonic epithelial cells) were obtained from Biological Science and kept in Dulbecco’s modified Eagle’s media (DMEM)/F12 containing 10% FBS, 10 ng/mL cholera toxin, 10 mM HEPES, insulin and transferring both at 0.005 mg/mL plus 100 ng/mL hydrocortisone medium. The SW480, HCT116 and FHC cell lines were incubated at 37 °C under 95% air and 5% CO2.
RNA Extraction and Quantitative Real-Time PCR
Total RNA was extracted from cells and tissues using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s protocol and quantified by UV 260/280 nm, as described previously.23,24 Total RNA (2 μg) and concentrations of sample RNA were reverse transcribed to cDNA in a final volume of 20 μL using the AVM First Strand cDNA synthesis kit (Invitrogen) following the manufacture’s instructions. The primer sequences were as follows: miRNA-140 forward, 5ʹ-ACACTCCAGCTGGGTACCACAGGG-3ʹ and reverse, 5ʹ-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCCGTG GTT-3ʹ. U6 forward, 5ʹ-CTCGCTTCGGCAGCACA-3ʹ, and reverse, 5ʹ-AAACGCTTCACGAATTTGCGT-3ʹ. They were used as cDNA templates for qPCR. The qPCR primers used for amplification areas were as follows: miRNA-140, forward 5ʹ-TCGGCAATTCAGTTGAGCCGTGGT T-3ʹ, reverse 5ʹ-ACACTCCAGCTGGGTACCACAGGG-3ʹ and TRAF6, forward 5ʹ-ATGCGGCCATAGGTTCTGC-3ʹ, reverse 5ʹ-TCCTCAAG ATGTCTCAGTTCCAT-3ʹ. All qPCRs were performed with the Thermo Scientific SYBR Green qPCR kit on an Applied Biosystems StepOne Real-time PCR System. PCR conditions were: 95 °C for 2 min, 95 °C for 15 sec, and 60 °C for 30 sec for 40 cycles. All gene transcripts were quantified by the 2−ΔΔCt method.
miRNA-140 Oligonucleotides, Vectors
The miRNA oligonucleotides including the mimic of miRNA-140 as well as their corresponding negative controls (NC) were commercially obtained from Genechem Co. Ltd. (Shanghai, China). pGL4.1-wtTRAF6 (Wild type TRAF6 3ʹUTR sequence) or pGL4.1-mutTRAF6 (Mutant TRAF6 3ʹUTR sequence) were stored by the Key Laboratory of Ministry of Education for Gastrointestinal Cancer, Fujian Medical University.
miRNA-140 Oligonucleotides Transfection
The transfection with miRNA-140 NC or miRNA-140 mimic of SW480 and HCT116 cells was performed with Lipofectamine 2000 (Life Technologies). At 48 h following transfection, the transfected SW480 and HCT116 cells were used for other experimental assays.
Cell Proliferation and Invasion Assays
Colorectal cancer cells which were treated using miRNA-140 oligonucleotides are planted in 96-well plates at a density of 1500 each well, 3 replicate wells per experimental group. These cells were cultured in RPMI-1640 medium with 10% fetal bovine serum in a 5% CO2, 37 °C incubator for 1, 2, 3, 4 and 5 days, then add 10% CCK-8 (Cell Counting kit-8, dojindo, Kumamoto, Japan) serum-free medium, counted by microplate reader (BioTek, Winooski, VT, USA) after continuing to culture for 1 h, as described previously.11
For cell invasion assay, 5×104 CRC cells with miRNA-140 oligonucleotides transfection were seeded into the upper chamber of 8 μm pore transwells (Corning, NY 14831, USA) coated with Matrigel (BD Bioscience, USA). A medium including 10% FBS was added to the lower chamber. Experiments were performed as described previously.11
Cell Migration and Wound Healing Assays
For the migration assay, the Transwell migration chamber experiment was used without Matrigel, and experiment steps were the same as the invasion assay.
For wound healing assay, CRC cells with miRNA-140 oligonucleotide transfection were implanted into 6-well plates. The experiments were performed as previously described.11
Dual-Luciferase Reporter Assay
SW480 and HCT116 cells were seeded at a density of 2×105 cells per well in 12-well plates. miRNA-140 oligonucleotide transfection was performed by use of Lipofectamine 2000 (Life Technologies), according to the manufacturer’s protocol. SW480 and HCT116 cells were co-transfected with miRNA-140 oligonucleotides, pRL-TK (Promega Corporation) and pGL4.1-wtTRAF6 or pGL4.1-mutTRAF6. Cells were lysed 48 h after transfection. A total of 20 μg of cell lysate was used for the intracellular luciferase activity, following the manufacturer’s protocol of the Dual-Luciferase reporter Assay system (Promega Corporation). The Renilla luciferase expression vector pRL-TK was used for normalization. Luminescence measurements were performed on a luminometer (Orion II Microplate Luminometer; Berthold Detection Systems GmbH). Each transfection was performed in duplicate and data are expressed as the mean ± standard deviation of three separate experiments.
Western Blot Analysis
The experiments and results analysis were carried out as described previously.23,25 The following primary antibodies were used in this study under the listings, respectively: TRAF6 (1:1000, Affinity), GAPDH (1:1000, Abcam), NF-κB (1:1000, Affinity), p-NF-κB (1:1000, Abcam), c-jun (1:1000, Bioss) and p-c-jun (1:1000, Bioss).
Animal Experiment
Animal experiments were performed in the Animal Experimental Center of Fujian Medical University and approved by the Animal Protection and Use Committee of Fujian Medical University. The animal experiments followed the guidelines of the 5 freedoms-animal welfare standards. 4×106 HCT116 cells which were transfected with miRNA-140 NC or miRNA-140 mimic oligonucleotides using lipofectamine2000 were injected into the right armpit of 4- to 6-week old BALB/c nu/nu nude mice. Mice were housed in a barrier facility on a high efficiency particulate arrestance (HEPA)-filtered rack under standard conditions of 12-hour light/dark cycles. The animals were fed an autoclaved laboratory rodent diet. Tumor length, width and mouse body weight were measured twice in a week. Tumor volume was calculated by following formula: tumor volume (mm3)=length (mm)×width (mm)× width (mm)×1/2. Data was presented as mean±SD. After 4 weeks, mice were humanely sacrificed and the tumors were removed, then the tumors were weighed and photographed. The tumors were soaked in formalin for further experiments.
Immunohistochemistry (IHC) and Evaluation
The IHC and assessment were carried out as previously described.25 The following primary antibody used was TRAF6 antibody (1:250 affinity). The expression of TRAF6 was evaluated using Image-Pro Plus 6.0 software (Media Cybernetics, Inc. USA). Semiquantitatively analyzed the expression of TRAF6 in each group using Mean Optical Density (MOD) = the Integral Optical Density (IOD)/the positive area.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism 5 software (GraphPad Software, Inc. La Jolla, CA, USA). Data were analyzed by Student’s t-test. The data were expressed as the mean ± standard deviation. A P-value of <0.05 was considered to indicate a statistically significant difference.
Results
The Expression Levels of miRNA-140 in Colorectal Cancer Tissues and Cells
We determined the miRNA-140 expression in CRC tissues and normal paired colorectal tissues which were cultured in a medium for 48 h. miRNA-140 is down-regulated in the CRC tissues compared with normal colorectal tissues (Figure 1A). And miRNA-140 expression is lower in SW480 and HCT116 cells than FHC cells which is normal for colorectal epithelial cells (Figure 1B).
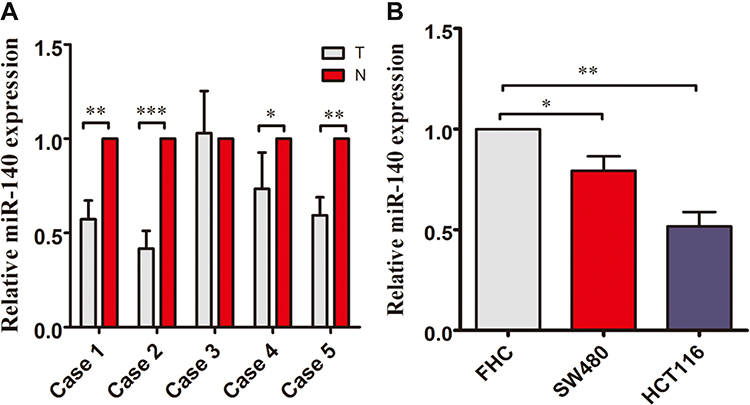 |
Figure 1 The expression of miRNA-140 is down-regulated in the colorectal cancer and cells. (A) The miRNA-140 expressions were decreased in colorectal cancer tissues (T) compared with normal colorectal tissues (N). (B) The miRNA-140 expression was significantly lower in SW480 and HCT116 cells than that in FHC cells which is the normal colorectal epithetical cell.*p<0.05,**p<0.01,***p<0.001. |
Over-Expression of miRNA-140 Suppressed the Proliferation, Migration and Invasion of SW480 and HCT116 Cells
To determine the influence of miRNA-140 on the colorectal cancer cells, we evaluated the proliferation, migration and invasion capacities using a series of cell biology assays in vitro. As shown in Figure 2A and B, miRNA-140 mimic transfection markedly enhanced the miRNA-140 expression in SW480 and HCT116 cells, respectively. Over-expression of miRNA-140 significantly decreased cell proliferation capacity in SW480 and HCT116 cells (Figure 2C and D). Wound healing and cell migration assays showed that over-expression of miRNA-140 significantly inhibited SW480 and HCT116 cells migration, respectively, compared with miRNA-140 NC controls (Figure 2E–H and I–K). The invasion transwell assays showed that over-expression of miRNA-140 could significantly weaken the invasive abilities of SW480 and HCT116 cells (Figure 2L–N). These data indicated that over-expression miRNA-140 could inhibit CRC growth and progression in vitro.
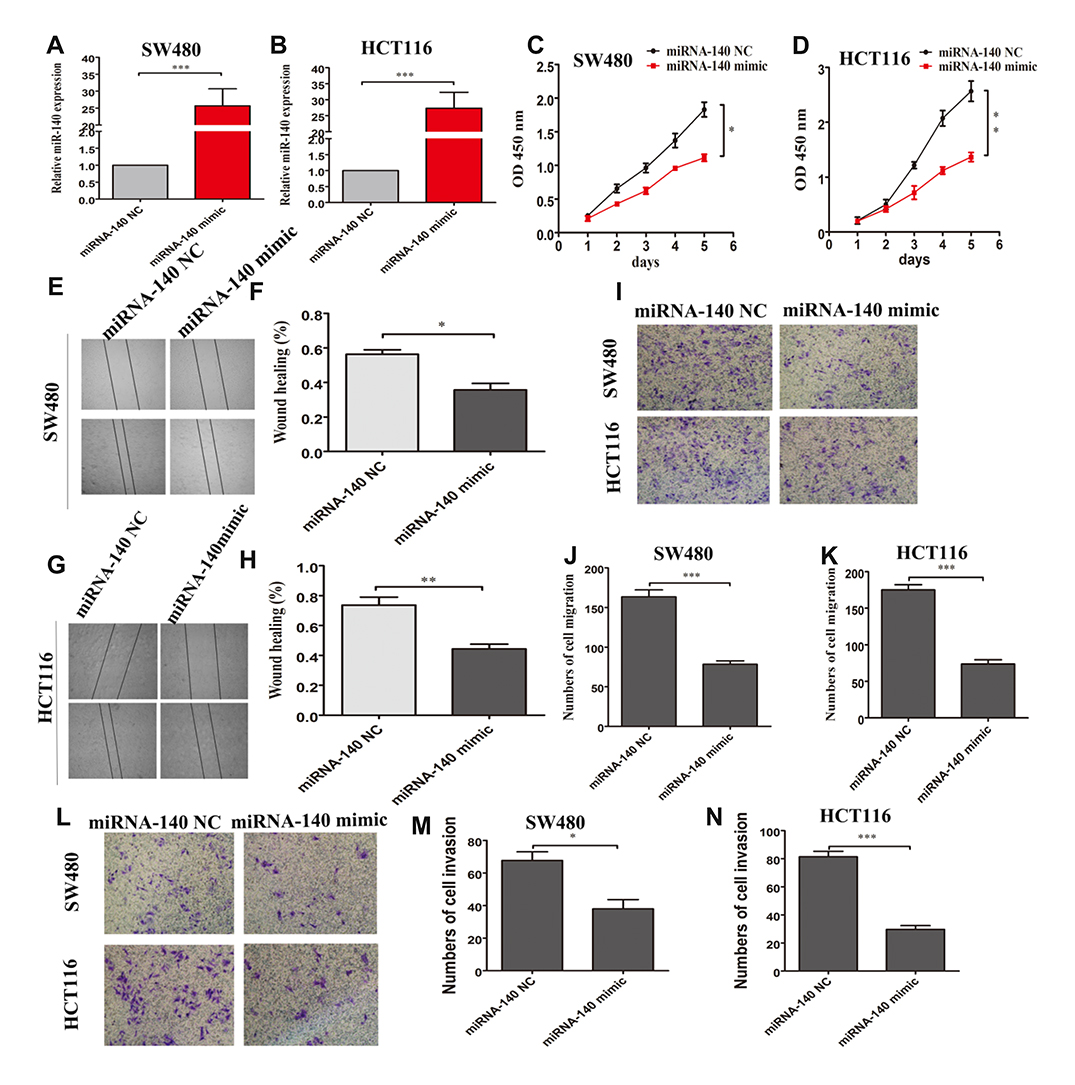 |
Figure 2 Over-expression of miRNA-140 suppressed the proliferation, migration and invasion of SW480 and HCT116 cells. (A and B) miRNA-140 mimic transfection were markedly increased the expression of miRNA-140 compared with miRNA-140 NC transfection in SW480 and HCT116 cells, respectively. (C and D) CCK-8 assays showed that over-expression of miRNA-140 significantly inhibited the proliferation ability of SW480 and HCT116 cells. (E, F, G and H) Wound healing experiments showed that over-expression of miRNA-140 significantly inhibited the migration of SW480 and HCT116 cells. (I, J and K) Transwell migration assays reported that over-expression of miRNA-140 significantly suppressed the migration of SW480 and HCT116 cells. (L, M and N) Transwell invasion assays reported that over-expression of miRNA-140 significantly suppressed the invasion of SW480 and HCT116 cells. *p<0.05,**p<0.01,***p<0.001. |
miRNA-140 Post-Transcriptionally Regulates TRAF6 Expression
To further investigate the reason of miRNA-140 regulating the expression of TRAF6, we carried out bioinformatics analysis using related prediction software. As illustrated in Figure 3A, a putative binding site for miRNA-140 was identified in the 3ʹUTR of TRAF6 mRNA. miRNA-140 over-expression inhibited wild-type TRAF6 3ʹUTR-luciferase pGL4.1-wtTRAF6 reporter vector in SW480 and HCT116 cells (Figure 3B and D). miRNA-140 over-expression did not affect the luciferase activity of pGL4.1-mutTRAF6 reporter vector in SW480 and HCT116 cells (Figure 3C and E).
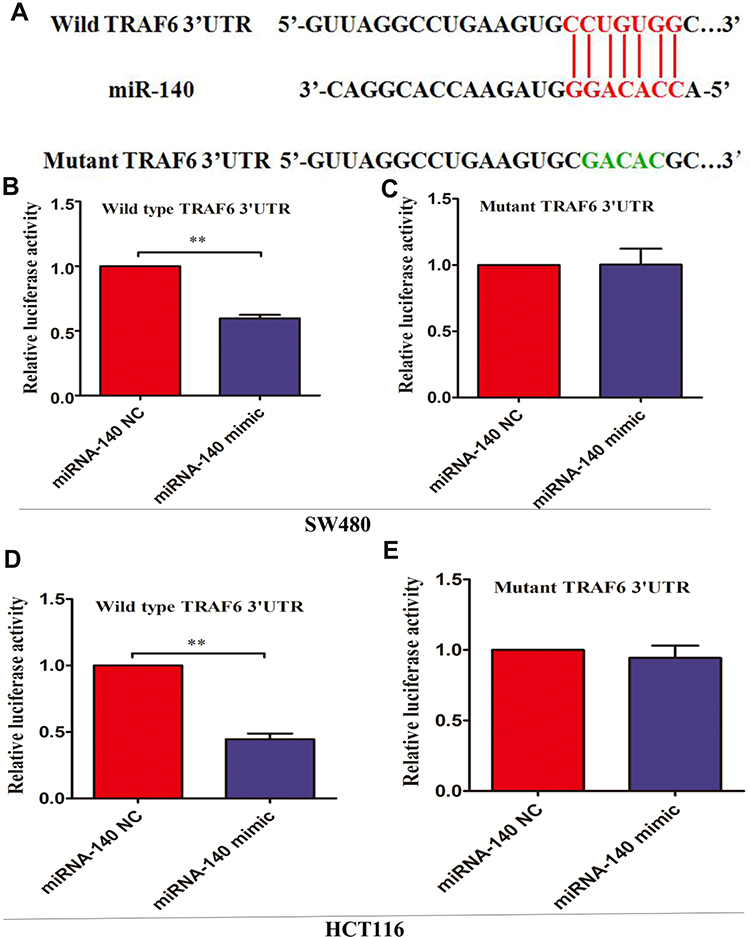 |
Figure 3 miRNA-140 inhibited TRAF6 expression though targeting the 3ʹ UTR area. (A) The wild-type 3ʹUTR of TRAF6 mRNA includes a putative miRNA-140 binding site. The mutant sequence was indicated with green letters. (B) Relative luciferase activity in SW480 cells co-transfected with miRNA-140 NC, miRNA-140 mimic and pGL4.1-wtTRAF6 reporter vectors. (C) Relative luciferase activity in SW480 cells co-transfected with miRNA-140 NC, miRNA-140 mimic and pGL4.1-mutTRAF6 reporter vectors. (D) Relative luciferase activity in HCT116 cells co-transfected with miRNA-140 NC, miRNA-140 mimic and pGL4.1-wtTRAF6 reporter vectors. (E) Relative luciferase activity in HCT116 cells co-transfected with miRNA-140 NC, miRNA-140 mimic and pGL4.1-mutTRAF6 reporter vectors. **p<0.01. |
Over-Expression of miRNA-140 Inhibited TRAF6 Expression
To determine the effects of miRNA-140 over-expression, we tested the mRNA and protein expression levels of TRAF6, after transfection miRNA-140 NC and miRNA-140 mimic for 48 h. The TRAF6 mRNA was significantly inhibited by miRNA-140 mimic transfection compared with miRNA-140 NC transfection in SW480 and HCT116 cells (Figure 4A and C). Consistent with the TRAF6 mRNA situation, TRAF6 protein expression was also markedly suppressed by miRNA-140 mimic transfection compared with miRNA-140 NC transfection in SW480 and HCT116 cells (Figure 4B and D).
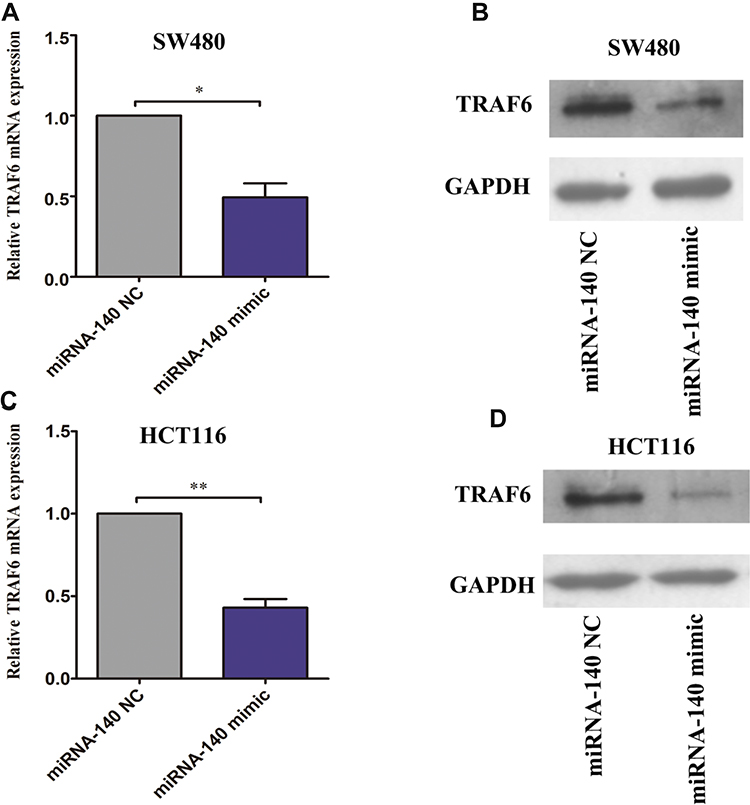 |
Figure 4 Over-expression of miRNA-140 inhibited TRAF6 expression in colorectal cancer cells. (A) qPCR analysis of the expression after transfection miRNA-140 NC and miRNA-140 mimic in SW480 cells, the TRAF6 mRNA expression levels were significantly inhibited by over-expression of miRNA-140. (B) Western blot analysis of the expression after transfection miRNA-140 NC and miRNA-140 mimic in SW480 cells, the TRAF6 protein expression levels were significantly inhibited by over-expression of miRNA-140. (C) qPCR analysis of the expression after transfection miRNA-140 NC and miRNA-140 mimic in HCT116 cells, the TRAF6 mRNA levels were significantly inhibited by over-expression of miRNA-140. (D) Western blot analysis of the expression after transfection miRNA-140 NC and miRNA-140 mimic in HCT116 cells, the TRAF6 protein levels were significantly inhibited by over-expression of miRNA-140. *p<0.05,**p<0.01. |
Over-Expression of miRNA-140 Suppressed the NF-κB/c-Jun Signaling Pathway Proteins Expression
Because over-expression of miRNA-140 inhibited the TRAF6 expression, we next determined the expression of NF-κB/c-jun signaling pathway which was regulated by TRAF6. After transfection of HCT116 cells with miRNA-140 NC and miRNA-140 mimic for 48 h, Western blot analysis showed that p-NF-κB and p-c-jun proteins were significantly inhibited by the miRNA-140 over-expression compared with miRNA-140 NC controls (Figure 5A and B). However, the proteins expression of NF-κB and c-jun were not changed in both miRNA-140 NC and miRNA-140 mimic over-expression (Figure 5A and B).
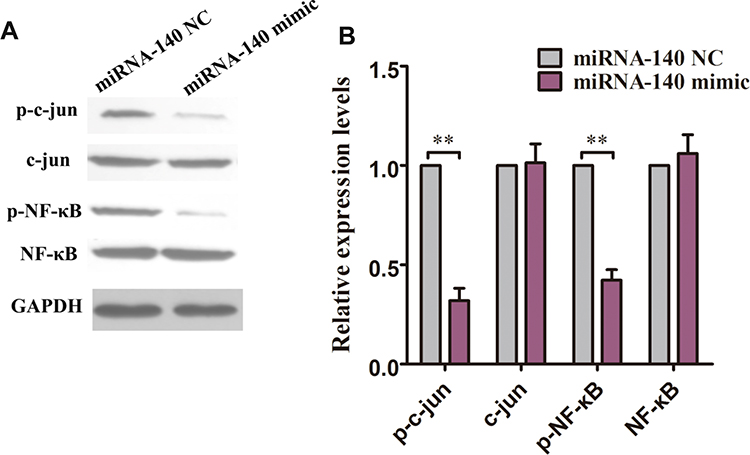 |
Figure 5 Over-expression of miRNA-140 decreased the activity of NF-κB/c-jun signaling pathways in HCT116 cells. (A) Western blot analysis found that over-expression of miRNA-140 markedly inhibited the p-NF-κB/p-c-jun proteins expression. (B) The histogram of NF-κB/c-jun signaling pathway is derived from A. **p<0.01. |
Over-Expression of miRNA-140 Inhibited the Growth of Colorectal Cancer in vivo
To determine the effects of over-expression of miRNA-140 on colorectal cancer in vivo, 4×106 miRNA-140 NC or miRNA-140 mimic HCT116 cells were injected into the right armpit of nude mice, respectively. After 4 weeks, we sacrificed the mice and found that the tumor growths were significantly inhibited in the miRNA-140 mimic group compared with miRNA-140 NC group (Figure 6A). Similarly, the weight of tumors in the miRNA-140 mimic group were lighter than that in the miRNA-140 NC group (Figure 6B). We made a tumor volume growth curve, and found that the speed of growth of tumors in the miRNA-140 mimic group were significantly suppressed compared with the miRNA-140 NC group (Figure 6C). We further carried out immunohistochemistry of tumors and found that the expression of TRAF6 was significantly suppressed in the miRNA-140 mimic group compared with miRNA-140 NC group (Figure 6D and E).
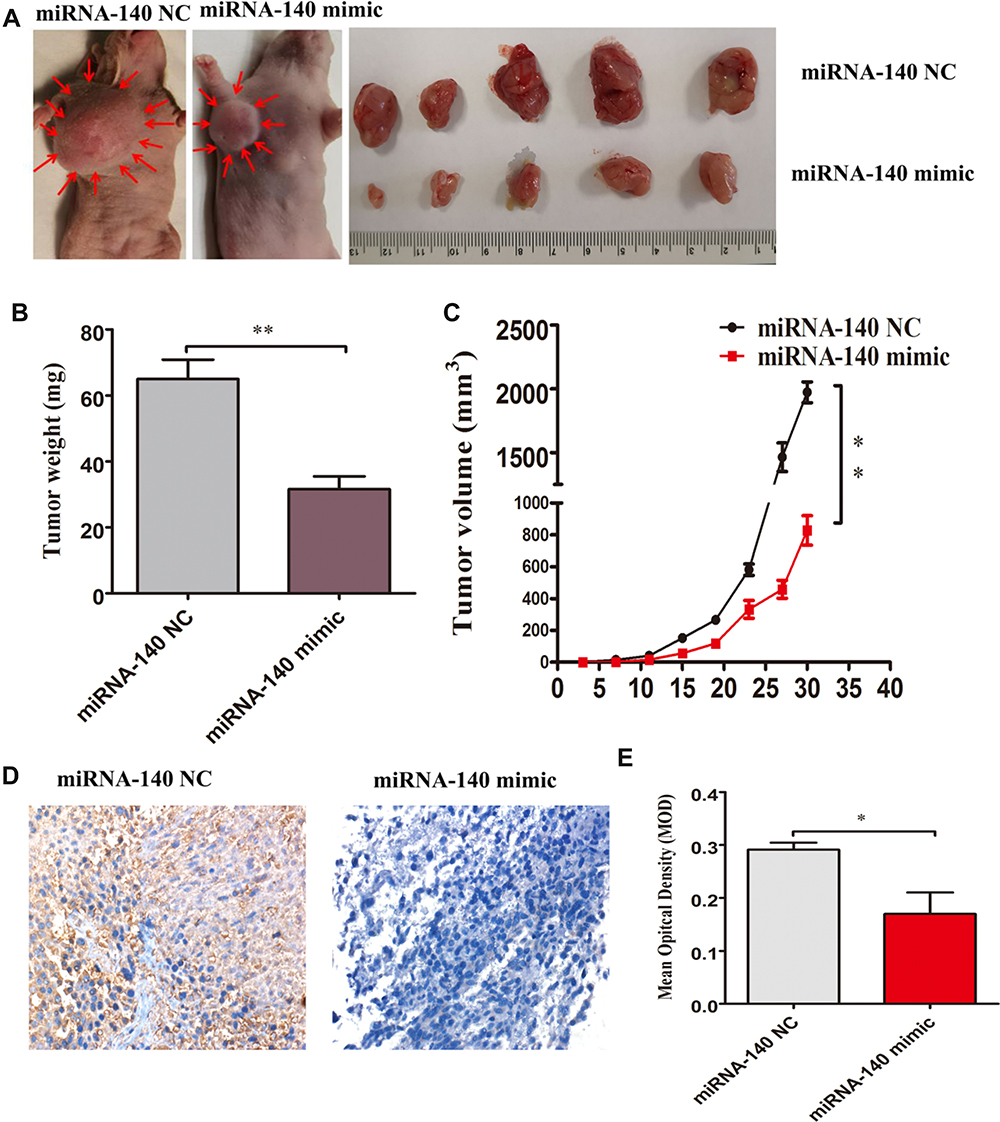 |
Figure 6 Over-expression of miRNA-140 suppressed growth of colorectal cancer in vivo. (A) Tumors in nude mice were harvested 4 weeks after initiation. The miRNA-140 mimic group tumor sizes were smaller than those in the miRNA-140 NC group. (B) After tumors were harvested, the weights of tumors were weighed, the miRNA-140 mimic group tumor weights were significantly reduced compared with miRNA-140 NC group. (C) The growth curves of tumors showed that the miRNA-140 mimic group tumors volume were markedly inhibited compared with the miRNA-140 NC group. (D) Immunohistochemistry analysis of expression of TRAF6 in subcutaneous xenograft tumors showed TRAF6 expression in miRNA-140 mimic group were decreased compared with miRNA-140 NC group. (E) The histogram of MOD of TRAF6 expression originated from (D). *p<0.05,**p<0.01. |
Discussion
TRAF6 is an important signal transduction molecule of the toll-like receptor (TLR) pathway, which plays a key role in innate and adaptive immunity.26,27 Recently studies have showed that TRAF6 functions as a cancer gene and has involvement in tumor malignancy.7,8,28,29 Our previous study also showed that TRAF6 promotes the progression and growth of colorectal cancer.11 In our previous study, we found that if we tested the miRNA-140 expression directly its expression levels were higher in colorectal cancer tissues than that in normal colorectal tissue. So in this study, we cultured the colorectal cancer tissues and normal colorectal tissues in the medium for 48 h. Then we determined that the expression levels of miRNA-140 in colorectal cancer tissue was lower than colorectal normal tissues. The reason may be LPS in the colorectal cancer tissues increased the expression of miRNA-140, because we previously found that LPS concentration in the colorectal cancer tissues was higher than that in normal cancer tissues.30 Consistent with cell lines miRNA-140 expression, we found miRNA-140 expression was highest in FHC cells than in SW480 and HCT116 cells.
Liu et al18 reported that miRNA-140 suppressed the proliferation and progression of Wilms’ tumor. Takata et al19 showed that miRNA-140 inhibited liver tumor progression. In the present study, we found that miRNA-140 inhibited the proliferation, migration and invasion in colorectal cancer. As we expected, over-expression of miRNA-140 inhibited the growth of colorectal cancer in vivo.
miRNAs are a class of evolutionarily conserved, non-protein coding RNAs, which are found in numerous organisms and tissues.31,32 miRNAs has been thought a novel method of gene expression regulation and through the transcriptional regulation regulated gene expression levels.12,13 To further investigate the mechanisms of how miRNA-140 inhibited the progression of colorectal cancer, we found miRNA-140 targeted the 3ʹUTR of TRAF6, then made the TRAF6 mRNA degradation.
Recently studies have revealed that TRAF6 could activate the NF-κB/c-jun signaling pathway. Song et al33 reported that TRAF6 activates NF-κB/c-jun, because TRAF6 and NF-κB-inducing kinase (NIK) bind indirectly. Some studies showed TRAF6 could be recruited to other signaling pathways (IL-1, IL-6, IL-8 and LPS) as a very important adapter protein to activate the NF-κB/c-jun signaling pathway.34–36 In our previous study, we also found that TRAF6 could inhibit the NF-κB/c-jun signaling pathway.11 So, we determined the change of the NF-κB/c-jun signaling pathway after transfection with miRNA-140 NC and mimic for 48 h. Consistent with our expectations, we found that the NF-κB/c-jun signaling pathway was inhibited.
Conclusion
In the present study, we found that miRNA-140 inhibited the proliferation, migration and invasion in colorectal cancer. Over-expression of miRNA-140 inhibited the growth of colorectal cancer in vivo. The mechanism studies found that miRNA-140 binds to TRAF6 mRNA 3ʹUTR, then caused TRAF6 mRNA degradation and TRAF6 protein expression decreased which reduced the activities of NF-κB/c-jun signaling pathways, influencing the expression of related genes. The relevant schematic is shown in Figure 7.
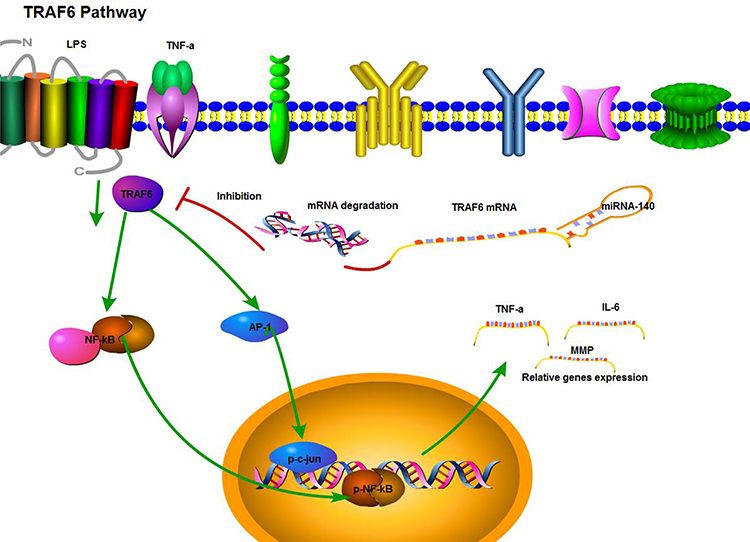 |
Figure 7 Schematic diagram of miRNA-140 regulation mechanism of TRAF6. Briefly, miRNA-140 binds with the TRAF6 mRNA 3ʹUTR area, then makes the TRAF6 mRNA degrade. Therefore, TRAF6 protein expression is reduced which reduces the activity of NF-κB/c-jun signaling pathways and decreases NF-κB/c-jun regulated related genes expression. |
Acknowledgments
This study was supported by the National Natural Science Foundation of China (No.81702424 and 81872364), the Joint Funds for the Innovation of Science and Technology, Fujian Province (No.2017Y9092), the Fujian Provincial Health Department Young and Middle-aged Talents Training Project (No.2018-ZQN-46), and the Project of Science and Technology Research Program in Fujian Province (No. BPB-2016 and 2018J01173).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics. CA cancer J clin. 2018;68(1):7–30.
2. Wang Z, Yip LY, Lee JHJ, et al. Methionine is a metabolic dependency of tumor-initiating cells. Nat med. 2019;25(5):825–837.
3. Andre T, Boni C, Mounedji-Boudiaf L, et al. Oxaliplatin, fluorouracil, and leucovorin as adjuvant treatment for colon cancer. N Engl J Med. 2004;350(23):2343–2351.
4. Yang AD, Fan F, Camp ER, et al. Chronic oxaliplatin resistance induces epithelial-to-mesenchymal transition in colorectal cancer cell lines. Clin Cancer Res. 2006;12(14 Pt 1):4147–4153.
5. Lin FT, Lin VY, Lin VT, Lin WC. TRIP6 antagonizes the recruitment of A20 and CYLD to TRAF6 to promote the LPA2 receptor-mediated TRAF6 activation. Cell Discovery. 2016;2.
6. Panda S, Nilsson JA, Gekara NO. Deubiquitinase MYSM1 Regulates Innate Immunity through Inactivation of TRAF3 and TRAF6 Complexes. Immunity. 2015;43(4):647–659.
7. Luo Z, Zhang X, Zeng W, et al. TRAF6 regulates melanoma invasion and metastasis through ubiquitination of Basigin. Oncotarget. 2016;7(6):7179–7192.
8. Han F, Zhang L, Qiu W, Yi X. TRAF6 promotes the invasion and metastasis and predicts a poor prognosis in gastric cancer. Pathol Res Pract. 2016;212(1):31–37.
9. Han Q, Yao F, Zhong C, Zhao H. TRAF6 promoted the metastasis of esophageal squamous cell carcinoma. Tumour Biology. 2014;35(1):715–721.
10. Li J-A, Kuang T, Pu N, et al. TRAF6 regulates YAP signaling by promoting the ubiquitination and degradation of MST1 in pancreatic cancer. Clinical and Experimental Medicine. 2019;19(2):211–218. doi:10.1007/s10238-018-00543-6
11. Zhu G, Cheng Z, Huang Y, et al. TRAF6 promotes the progression and growth of colorectal cancer through nuclear shuttle regulation NF-kB/c-jun signaling pathway. Life Sci. 2019;235:116831. doi:10.1016/j.lfs.2019.116831
12. Yeh CH, Moles R, Nicot C. Clinical significance of microRNAs in chronic and acute human leukemia. Mol Cancer. 2016;15(1):37.
13. Peng S, Gao D, Gao C, Wei P, Niu M, Shuai C. MicroRNAs regulate signaling pathways in osteogenic differentiation of mesenchymal stem cells (Review). Mol Med Rep. 2016;14(1):623–629.
14. Fadaka AO, Pretorius A, Klein A. Biomarkers for Stratification in Colorectal Cancer: microRNAs. Cancer Control. 2019;26(1):1073274819862784.
15. O’Connell RM, Rao DS, Chaudhuri AA, Baltimore D. Physiological and pathological roles for microRNAs in the immune system. Nat Rev Immunol. 2010;10(2):111–122.
16. Gammell P. MicroRNAs: recently discovered key regulators of proliferation and apoptosis in animal cells: identification of miRNAs regulating growth and survival. Cytotechnology. 2007;53(1–3):55–63.
17. He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet. 2004;5(7):522–531.
18. Liu Z, He F, OuYang S, et al. miR-140-5p could suppress tumor proliferation and progression by targeting TGFBRI/SMAD2/3 and IGF-1R/AKT signaling pathways in Wilms’ tumor. BMC Cancer. 2019;19(1):405.
19. Takata A, Otsuka M, Yoshikawa T, et al. MicroRNA-140 acts as a liver tumor suppressor by controlling NF-kappaB activity by directly targeting DNA methyltransferase 1 (Dnmt1) expression. Hepatology. 2013;57(1):162–170.
20. Wolfson B, Eades G, Zhou Q. Roles of microRNA-140 in stem cell-associated early stage breast cancer. BMC Cancer. 2014;6(5):591–597.
21. Chen B, Liu B. MiRNA-381 inhibits the invasion of renal carcinoma and the underlying mechanisms. J Central South Univ Med Sci. 2015;40(10):1053–1059.
22. Li D, Bao P, Yin Z, et al. Exploration of the involvement of LncRNA in HIV-associated encephalitis using bioinformatics. PeerJ. 2018;6:e5721.
23. Zhu G, Ye J, Huang Y, et al. Receptor-interacting protein-1 promotes the growth and invasion in gastric cancer. Int J Oncol. 2016;48(6):2387–2398.
24. Zhu G, Du Q, Wang X, Tang N, She F, Chen Y. TNF-alpha promotes gallbladder cancer cell growth and invasion through autocrine mechanisms. Int J Mol Med. 2014;33(6):1431–1440.
25. Zhu G, Chen X, Wang X, et al. Expression of the RIP-1 gene and its role in growth and invasion of human gallbladder carcinoma. Cellular Physiol Biochem. 2014;34(4):1152–1165.
26. Zhang J, Zhu Y, Chen Z, Li C, Zhao X, Kong X. Molecular cloning and expression analysis of MyD88 and TRAF6 in Qihe crucian carp Carassius auratus. Fish Shellfish Immunol. 2019;87:829–838.
27. Wang P, Zhang Z, Xu Z, Guo B, Liao Z, Qi P. A novel invertebrate toll-like receptor with broad recognition spectrum from thick shell mussel Mytilus coruscus. Fish Shellfish Immunol. 2019;89:132–140.
28. Yang WL, Wang J, Chan CH, et al. The E3 ligase TRAF6 regulates Akt ubiquitination and activation. Science. 2009;325(5944):1134–1138.
29. Zhang X, Wu L, Xiao T, et al. TRAF6 regulates EGF-induced cell transformation and cSCC malignant phenotype through CD147/EGFR. Oncogenesis. 2018;7(2):17.
30. Zhu G, Huang Q, Huang Y, et al. Lipopolysaccharide increases the release of VEGF-C that enhances cell motility and promotes lymphangiogenesis and lymphatic metastasis through the TLR4- NF-kappaB/JNK pathways in colorectal cancer. Oncotarget. 2016;7(45):73711–73724.
31. Zhu Z, Tang J, Wang J, Duan G, Zhou L, Zhou X. MiR-138 Acts as a Tumor Suppressor by Targeting EZH2 and Enhances Cisplatin-Induced Apoptosis in Osteosarcoma Cells. PLoS One. 2016;11(3):e0150026.
32. Wu S, Du X, Wu M, Du H, Shi X, Zhang T. MicroRNA-409-3p inhibits osteosarcoma cell migration and invasion by targeting catenin-delta1. Gene. 2016;584(1):83–89.
33. Song HY, Regnier CH, Kirschning CJ, Goeddel DV, Rothe M. Tumor necrosis factor (TNF)-mediated kinase cascades: bifurcation of nuclear factor-kappaB and c-jun N-terminal kinase (JNK/SAPK) pathways at TNF receptor-associated factor 2. Proc Natl Acad Sci U S A. 1997;94(18):9792–9796.
34. Schwandner R, Yamaguchi K, Cao Z. Requirement of tumor necrosis factor receptor-associated factor (TRAF)6 in interleukin 17 signal transduction. J Exp Med. 2000;191(7):1233–1240.
35. Ye X, Wu H, Sheng L, et al. Oncogenic potential of truncated RXRalpha during colitis-associated colorectal tumorigenesis by promoting IL-6-STAT3 signaling. Nat Commun. 2019;10(1):1463.
36. Li ZW, Sun B, Gong T, et al. GNAI1 and GNAI3 Reduce Colitis-Associated Tumorigenesis in Mice by Blocking IL6 Signaling and Down-regulating Expression of GNAI2. Gastroenterology. 2019;156(8):2297–2312.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.