Back to Journals » Infection and Drug Resistance » Volume 16
The Drug-Specific Propensity Regarding the Acquisition of Fluoroquinolone Resistance in Escherichia coli: An in vitro Challenge and DNA Mutation Analysis
Authors Sato T ![]() , Ito R, Kawamura M
, Ito R, Kawamura M ![]() , Fujimura S
, Fujimura S
Received 24 July 2023
Accepted for publication 21 September 2023
Published 26 September 2023 Volume 2023:16 Pages 6357—6366
DOI https://doi.org/10.2147/IDR.S428383
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Takumi Sato,1 Ryota Ito,1,2 Masato Kawamura,1 Shigeru Fujimura1
1Division of Clinical Infectious Diseases & Chemotherapy, Tohoku Medical and Pharmaceutical University, Sendai, Japan; 2Department of Pharmacy, Japan Labor Health and Welfare Organization Tohoku Rosai Hospital, Sendai, Japan
Correspondence: Takumi Sato, Division of Clinical Infectious Diseases & Chemotherapy, Tohoku Medical and Pharmaceutical University, 4-4-1, Komatsushima, Aoba-ku, Sendai, 981-8558, Japan, Tel +81227270176, Fax +81227270177, Email [email protected]
Objective: Many fluoroquinolones, such as ciprofloxacin, are used clinically. We investigated the relationship between resistance acquisition and exposure duration in each drug through the exposure of fluoroquinolone to Escherichia coli clinical isolates in vitro.
Methods: Eleven E. coli clinical isolates were exposed to each fluoroquinolone, ie, ciprofloxacin, levofloxacin, sitafloxacin, garenoxacin, and lascufloxacin, with the concentration of the mutant selection window for 5 days; these procedures were repeated 5-times. In addition, the DNA sequence in the quinolone-resistance determining region (QRDR) and the expression level in the drug efflux pump acrA were analyzed to determine the resistance mechanism.
Results: Although resistant strains were not detected after 5 to 10 days of exposure to fluoroquinolone, after 25 days of exposure to ciprofloxacin and levofloxacin, 100% and 45% of isolates acquired resistance, respectively. Due to 25 days of exposure to sitafloxacin, garenoxacin, and lascufloxacin, MIC measurement was elevated 2- to 4096-fold for those of the parental strain, and the cross-resistance rate to levofloxacin was 72%, 54%, and 27%, respectively. In strains with high fluoroquinolone resistance, acrA overexpression was observed in addition to QRDR mutation.
Conclusion: In our findings, fluoroquinolone resistance was not observed in the E. coli strain after 5- to 10-days of exposure. However, resistance acquisition was detected frequently after 15- to 25-days of exposure. Among fluoroquinolones, lascufloxacn had the least impact on the resistance acquisition in E. coli.
Plain Language Summary: The need to combat quinolone-resistant E. coli is urgent. The resistance acquisition tendency of E. coli varies according to the drug used. After 25 days of exposure, 45% of E. coli showed levofloxacin resistance. Lascufloxacin may have the lowest influence on resistance acquisition to regular E. coli.
Keywords: Escherichia coli, respiratory quinolone, mutant selection window, quinolone-resistance determining region, efflux pump
Introduction
The recent increase in fluoroquinolone resistance in Escherichia coli is one of the most severe threats to public health.1 In the least surveillance, the frequency of fluoroquinolone-resistant E. coli has been reported at 22% in Europe,2 21% in the United States,3 and 38% in Japan.4 The increase of such quinolone-resistant E. coli has been attributed to the use of fluoroquinolones. Subsequently, the guidelines of the Infectious Diseases Society of America and European Society of Clinical Microbiology and Infectious Diseases were reviewed,5 and a policy to refrain from quinolone consumption was implemented. However, the epidemic of fluoroquinolone-resistance E. coli continues to increase.
There are two main reasons for the expansion of drug-resistant E. coli. The first is the expansion of mobile genetic elements across bacterial species, mediated by a plasmid including an extended spectrum of beta-lactamase. The second is inappropriate use of antibacterial agents. Generally, a quinolone is selected as treatment for respiratory or urinary tract infections. In addition, the on-period for respiratory infection is longer than that of urinary tract infection.6 Respiratory quinolones such as levofloxacin, sitafloxacin, garenoxacin, and lascufloxacin are sometimes used to manage respiratory infections such as pneumonia or lung abscesses.7 Thus, respiratory quinolone brands have increased recently. It is known that the fluoroquinolone resistance mechanism is the mutation in the quinolone-resistance determining region (QRDR) of gyrA/parC and the high expression of efflux pumps such as AcrAB. In this study, to elucidate the impact of respiratory quinolone on the development of fluoroquinolone resistance in E. coli, we performed an in vitro challenge against E. coli clinical isolates and analyzed QRDR DNA mutation.
Materials and Methods
Bacterial Strains and Antimicrobial Susceptibility Testing
A total of 40 E. coli clinical isolates [vaginal discharges (n=30), urine (n=6), and other (n=4)] detected in a tertiary hospital in Japan were used in this study. The minimum inhibitory concentration (MIC) of each fluoroquinolone, ciprofloxacin (CPFX; LKT Laboratories Inc., MN), levofloxacin (LVFX; LKT Laboratories Inc.), sitafloxacin (STFX; Daiichi Sankyo Co., Ltd., Japan), garenoxacin (GRNX; Toyama Chemical Co., Ltd., Japan) and lascufloxacin (LSFX; Kyorin Pharmaceutical Co., Ltd., Japan), was determined by a broth microdilution method according to CLSI guidelines.8 The decision of susceptibility was conducted using the CLSI breakpoint table M100-S30.9 Namely, it was judged that CPFX: ≥2 μg/mL or LVFX: ≥4 μg/mL had resistance.
Mutant Prevention Concentration and Mutant Selection Window by in vitro for Each Fluoroquinolone Exposure
A total of 11 strains of 10 clinical isolates indicating susceptibility to all quinolones, and E. coli ATCC 25922 were selected for in vitro exposure to each fluoroquinolone. Initially, mutant prevention concentration (MPC) was determined for these 11 strains with agar dilution.10 In brief, 100 μL of bacterial suspensions with 1010 CFU were applied onto a Mueller-Hinton agar plate containing quinolone ranging from 0.5x to 32x against their MIC. After incubation for 120 hours (ie, 5 days) at 37°C, the MPC was determined to have the lowest quinolone concentration at which no colonies grew on the agar plate. The mutant selection window (MSW) was shown as the concentration zone from MIC to MPC. A strain that was exposed to fluoroquinolone with half the MPC for 5 days was determined as MIC and subsequently incubated for 5 days (total of 10 days) on the plate containing quinolone ranging from 0.5x to 32x against its MIC to determine a second-time MPC. To expose fluoroquinolone for 25 days, these procedures were repeated by 5 times.
DNA Sequencing of QRDR and Quantification of Efflux Pump Expression
To investigate the quinolone-resistant mechanisms in the strain exposed for 25 days, genome DNA was extracted using the InstaGene matrix (BIO-RAD). DNA sequences of gyrA, gyrB, and parC were determined by the dideoxy chain termination method.11 Furthermore, to compare the mRNA expression of the drug efflux pump gene acrA12,13 between the parental strain and the strain exposed for 25 days, total RNA was extracted using TRI Reagent LS (Molecular Research Center, Inc., Cincinnati, OH). The mRNA expressions were quantified by the iTaq Universal CYBR Green One-step kit (BIO-RAD) and CFX Connect Real-Time System (BIO-RAD). The target gene expression levels were standardized by the ΔΔCt method against the gapA gene and displayed relative expression levels as a multiple of that of the parental strain. The over-expression was defined as a more than 3-fold increase of expression levels.14
Spontaneous Mutation Frequency
Spontaneous mutation frequency was measured by the standard method ascribed by Kohanski et al.15 Bacteria were incubated for 24 hours in the MHB with sub-MIC of fluoroquinolone. Subsequently, rifampicin-resistant, and total bacterial counts were acquired by plating onto MHA with or without 100 μg/mL rifampicin. The mutation frequency was shown as a multiple of rifampicin-resistant bacterial counts against total bacterial counts. In addition, the frequency between a group control and a drug-exposed group was compared by an unpaired t-test.
Results
Fluoroquinolone Susceptibility of E. coli Clinical Isolates
The distribution of each quinolone MIC for 40 E. coli clinical isolates is shown in Figure 1. The MIC50 / MIC90 of each quinolone was as follows: CPFX, 0.031 / 64 μg/mL; LVFX, 0.031 / 32 μg/mL; STFX, 0.031 / 1 μg/mL; GRNX, 0.063 / 32 μg/mL; LSFX, 0.25 / 32 μg/mL. Among these 40 E. coli isolates, 6 strains had high resistance with CPFX MIC ≥32 μg/mL. These 6 strains had four mutation points in QRDR uniformly (Table 1).
 |
Figure 1 Distribution of MIC values of fluoroquinolone against Escherichia coli clinical isolates (n=40). The data shows the number of strains indicating each MIC value of fluoroquinolone. Abbreviations: CPFX, ciprofloxacin; LVFX, levofloxacin; STFX, sitafloxacin; GRNX, garenoxacin; LSFX, lascufloxacin. |
 |
Table 1 MIC Values and Resistance Mechanisms on High Ciprofloxacin-Resistant Clinical Isolates |
The Selection of CPFX- or LVFX-Resistant Strain Within MSW
Fluoroquinolone susceptible E. coli (n=11) had CPFX MSWs ranging from 0.016 to 0.18 μg/mL, which were displayed in the area vastly below the CLSI breakpoint (Figure 2). After 15 days of CPFX exposure, 6 CPFX-resistant strains (54%) were selected from MSW that expanded to 0.66–4 μg/mL. Furthermore, the CPFX-resistant rate reached 100% with expanded MSW ranging from 4 to 21 μg/mL through 25 days of exposure. In 15 days of LVFX exposure, although MSW was expanded from 0.04–0.44 μg/mL to 0.6–4.6 μg/mL, only 1 strain (9%) showed resistance; 5 strains (45%) had resistance after 25 days of exposure (Figure 3). Determining the QRDR mutation in each fluoroquinolone resistance acquired strain, GyrA mutation (S83L or D87G) was observed in all 11 strains that showed CPFX resistance after 25 days of exposure (Figure 4). Among GyrA mutants, 4 strains also had GyrB mutation (Q411R, E454D, E454H, or V467M), and 6 strains had ParC mutation (S80I or E84K). The high CPFX-resistant strains with MIC ≥32 μg/mL had an overexpression in the efflux pump gene acrA in addition to GyrA and ParC mutation. In 5 LVFX-resistant strains detected after 25 days of exposure, ParC mutation or acrA overexpression was not observed, although they had GyrA mutation (S83L, D87G, D87Y) or GyrB mutation (E454D, E454K, V467E).
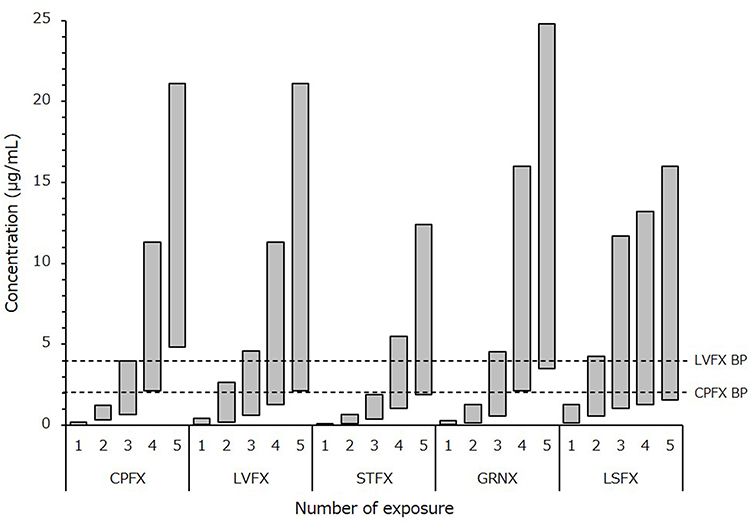 |
Figure 2 Changes of each quinolone mutant selection window after any number of exposures to the drug The mutant selection window (MSW) mean is indicated as a gray box, ranging from MIC to MPC. The vertical axis shows the concentration of each fluoroquinolone. A dashed line shows the CLSI breakpoint of ciprofloxacin and levofloxacin. Abbreviations: CPFX, ciprofloxacin; LVFX, levofloxacin; STFX, sitafloxacin; GRNX, garenoxacin; LSFX, lascufloxacin. |
 |
Figure 3 Cumulative resistance rate of each quinolone antibiotic after any days of exposure to the drug The vertical axis shows the cumulative resistance rate (%) and the abscissa axis shows the total number of days of exposure to ciprofloxacin (Open diamond) or levofloxacin (Open square). |
 |
Figure 4 The MIC changes of Escherichia coli isolates and their resistance mechanisms in the exposure to ciprofloxacin or levofloxacin. The data plots the ciprofloxacin and levofloxacin MIC measurement. In the fluoroquinolone-exposed strain, different icons are used to indicate the corresponding resistance mechanisms. Abbreviations: CPFX, ciprofloxacin; LVFX, levofloxacin. |
The Changes in MICs with STFX, GRNX, or LSFX Exposure
The MSWs of STFX and GRNX were 0.016–0.11 and 0.04–0.3 μg/mL, respectively. After 25 days of exposure to each drug, these MSWs expanded to 1.9–12.4 μg/mL (113-fold) and 3.3–25 μg/mL (83-fold) (Figure 2). Conclusively, the MIC measurement of STFX became 1–64 μg/mL which increased 64–4096-fold from its parental strains; similarly, the MIC of GRNX increased 16–2048-fold, and its measurement showed 0.5–64 μg/mL (Figure 5). Furthermore, all STFX and GRNX exposed strains with MIC ≥8 μg/mL had acrA over-expression (ranging from 3.13- to 6.11-fold) in addition to QRDR mutation.
 |
Figure 5 The MIC changes of Escherichia coli isolates and their resistance mechanisms in the exposure to sitafloxacin, garenoxacin, or lascufloxacin The plotted data shows sitafloxacin, garenoxacin, and lascufloxacin MIC measurement in wildtype and fluoroquinolone-exposed strains. Different icons are used to indicate the corresponding resistance mechanisms. Abbreviations: STFX, sitafloxacin; GRNX, garenoxacin; LSFX, lascufloxacin. |
On the other hand, the changes of MSW in LSFX were most gradual among the fluoroquinolones tested; it was only 12-fold (from 0.17–1.3 μg/mL to 1.9–16 μg/mL) (Figure 2).
LSFX MICs were elevated to 1- to 32-fold from those of parental strains after 25 days of exposure; these strains had a mutation in GyrA or GyrB, and ParC mutation and acrA over-expression was not detected (Figure 5).
Mutation Frequency of E. coli
Spontaneous QRDR mutation frequency, as incubated in an antibiotic-free medium, was the mean of 10−8.4 (Figure 6). Due to the exposure to half the MIC of CPFX, STFX, and GRNX, this frequency was increased by 3.2-, 7.9-, and 6.3-fold, respectively (P <0.05). In contrast, no increased mutation frequency was observed in the exposure to LVFX and LSFX.
 |
Figure 6 Spontaneous mutation frequency of Escherichia coli strains exposed to each quinolone. The box-plot data shows comparisons of the spontaneous mutation frequency between a strain with control or exposed to half the MIC fluoroquinolone for 24 hours. *P <0.05. |
Discussion
Fluoroquinolones are used for the treatment of systemic bacterial infections because they exhibit excellent bactericidal efficacy and tissue penetration, and their dosing period varies according to each infection. For example, although a dosing period from 3 to 5 days is recommended for uncomplicated cystitis,5 bacteremia and pneumonia require 7 to 14 days.16–18 Furthermore, the dosing period for lung abscesses is extremely long, and the median in this dosing period was reported as 26 days (maximum 180 days).19 These prolonged administrations of fluoroquinolone are one of the risk factors for the global increase in fluoroquinolone-resistant E. coli.20,21 In the previous studies that investigated the risk for the emergence of fluoroquinolone-resistant E. coli,22,23 cancer and respiratory tract infection, not uncomplicated-urinary tract infection, were suggested. These results are considered to be due to the difference in the dosing period of the fluoroquinolone; however, our findings also showed the difference in each fluoroquinolone.
In our findings, both the values of MIC and MPC were elevated at every repeat of the MSW exposure. Although the Cmax (0.56 μg/mL) when healthy adults took once an approved dose of CPFX24 was higher than the initial MPC level, it was included in MSW after the second load experiment. However, in LVFX (7.6 μg/mL),25 STFX (1.0 μg/mL),26 and GRNX (11.1 μg/mL)27 with a relatively high C max, the period before the inclusion of each C max in MSW was long. Because E. coli is exposed to each quinolone in one MSW load experiment for 5 days, it is difficult to consider that those strains acquire quinolone resistance in approximately 5 days. However, by the treatment that assumes a remedy of 10 to 15 days, the resistant strain may emerge. Our results support the findings of Fantin et al23 that reported quinolone treatment for 14 days or more increases the risk of resistance.
After comparing the acquisition of the resistance rate of CPFX with LVFX, there was no difference by 2 times the exposure (10 days). Whereas that of LVFX was low by 5 times the exposure (25 days). The period of the fluoroquinolone recommended for the treatment of acute uncomplicated cystitis was 3 to 5 days, and it was considered that both drugs did not contribute to the acquisition of resistance by this regimen. Whereas, in the duration of treatment (more than 14 days) assumed for severe pyelonephritis and sepsis, it may be easy for CPFX to give quinolone resistance to E. coli. In this study, the resistant strain which emerged by CPFX exposure had multiple resistant mechanisms compared to that of LVFX where the MIC level was also high. Particularly, ParC mutation that greatly elevated the MIC value of both drugs was found only in resistant strains by CPFX. E. coli GyrA and ParC mutation increase CPFX IC50 by 280-fold but increase LVFX IC50 only 55-fold.28 In other words, this might delay resistance acquisition as LVFX maintained bactericidal activity even if the mutation of QRDR occurred. However, the ParC variant by the CPFX exposure greatly elevated the MIC of LVFX. Although the ParC variant did not emerge by 25-day LVFX exposure in this study, further long-term exposure may allow its emergence.
On the other hand, in most of the STFX or GRNX exposure strains, AcrA had overexpression. AcrA is one of the components of the trimeric ABC transporter AcrAB-TolC.29 The overexpression of this transporter plays a role in the efflux of fatty acid or biliary acid within the intestine and produces fluoroquinolone resistance. AcrAB-TolC expression is induced by exposure to organic anions such as the carboxylic acid group.30 Considering the carboxylic acid pKa value of the drug used in this study, the proportion of anion types of STFX and GRNX at a pH of 7.2, such as infection site and medium, is 10-fold higher than other drugs; these may induce acrA expression. Given highly resistant strains with GyrA+ParC mutation that did not observe acrA over-expression had low STFX MICs, acrA overexpression may be crucial in an increase of STFX MICs. GRNX and LSFX MICs in these strains increased 8- to 64-fold from parental strains. These results indicate that the rise of GRNX and LSFX MICs requires both GyrA and ParC mutation; acrA overexpression negligibly contributed to the elevation of both drugs’ MICs. On the other hand, 5 times LSFX-exposed strains had few elevated MICs, showing the lowest cross-resistance rate at 27% against CPFX and LVFX. This may result from the fact that GyrA mutation was observed in LSFX-exposed strains but was not observed in ParC mutation and acrA over-expression. LSFX is developed in order to enable strong binding to both GyrA and ParC;31 therefore, it was considered that strains with QRDR mutation were hardly selected with LSFX exposure.
Respiratory quinolones such as LVFX, STFX, GRNX, and LSFX are used for bacterial respiratory infection. In these cases, E. coli was rarely detected as a causative organism (approximately 1 to 4%);32 conversely, E. coli colonizes in intestinal microbiota. If fluoroquinolones are administered orally, about 10% of the dosage is distributed and eliminated in the intestine, and drug concentration within the intestine almost equals serum drug concentration.33 In other words, the fluoroquinolone exposure to E. coli occurs in not only the infection location, but also the intestinal tract. Each MIC of STFX, GRNX, and LSFX after the 5 times exposure that assumed a prolonged administration was 64- to 4096-fold, 16- to 2048-fold, and 2- to 32-fold elevated, respectively. And all strains in which these MICs were elevated acquired cross resistance to CPFX and LVFX. When respiratory quinolone is administered for a long term, it is necessary to be vigilant about fluoroquinolone resistance of E. coli that habitually resides in the intestinal tract. Our results suggest that LSFX has the lowest influence on fluoroquinolone-resistance acquisition to regular E. coli.
Bacteria, such as E. coli, can mutate in DNA in every cell division; its natural frequency was reported at 10−9 to 10−8.14 This spontaneous mutation frequency sometimes increases when exposed to drugs harboring the N-cyclopropyl groups, such as cyclophosphamide.34 Three fluoroquinolones (CPFX, STFX, and GRNX) which showed over 50% of cross-resistance to LVFX, also located an N-cyclopropyl group binding to the quinoline structure. This structure plays an essential role in antibacterial activity. However, it was reported that DNA mutation was induced by the high chemical reactivity.34 Indeed, mutation frequency was increased when exposed to these three drugs (P <0.05); in contrast, that of LSFX and LVFX did not exceed baseline.
In other drug resistance, the extended-spectrum beta-lactamase (ESBL) -producing strains tend to increase. Recently, SHV-producing strains were determined in not only Klebsiella pneumonia but also E. coli.35 Most ESBL-producing E. coli show fluoroquinolone resistance; thus, the prevalence of these strains should be noted.
In conclusion, this study showed that fluoroquinolone resistance acquisition considerably depends on drug selection and exposure period. Our data shows that fluoroquinolone-resistant E. coli was not detected within ten days of a CPFX/LVFX challenge, so these drugs may be used for about seven days without developing resistance. On the other hand, E. coli was resistant at a high rate by 25-day exposure, and there was a higher tendency in CPFX; therefore, prolonged use of fluoroquinolones for longer than 25 days should be avoided. Although this tendency was similar for respiratory quinolone, it was difficult for LSFX to acquire cross-resistance for LVFX by exposure for 25 days.
Ethical Approval
Not required, according to the Research Ethics Statement of the Ministry of Health, Labour and Welfare of Japan, because this study is basic research that used bacterial isolates only. Following this statement, this study is exempt from ethical approval. Clinical isolates that were used in this study were collected and stocked by the Tohoku Infectious Diseases Society; therefore, we cannot access patient information.
Acknowledgments
We thank Dr. Akira Watanabe of Tohoku Infectious Disease Society, for he gifted E. coli clinical isolates. We also thank M. Yagi for technical assistance.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis, and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Aliabadi S, Anyanwu P, Beech E, et al. Effect of antibiotic stewardship interventions in primary care on antimicrobial resistance of Escherichia coli bacteraemia in England (2013–18): a quasi-experimental, ecological, data linkage study. Lancet Infect Dis. 2021;21(12):1689–1700. doi:10.1016/S1473-3099(21)00069-4
2. Critchley IA, Cotroneo N, Pucci MJ, Jain A, Mendes RE. Resistance among urinary tract pathogens collected in Europe during 2018. J Glob Antimicrob Resist. 2020;23:439–444. doi:10.1016/j.jgar.2020.10.020
3. Thompson DK, Muradyan AG, Miller ASF, Ahiawodzi PD. Antibiotic resistance of Escherichia coli urinary tract infections at a North Carolina community hospital: comparison of rural and urban community type. Am J Infect Control. 2022;50(1):86–91. doi:10.1016/j.ajic.2021.08.032
4. Kobayashi K, Yamamoto S, Takahashi S, et al. The third national Japanese antimicrobial susceptibility pattern surveillance program: bacterial isolates from complicated urinary tract infection patients. J Infect Chemother. 2020;26(5):418–428. doi:10.1016/j.jiac.2020.01.004
5. Gupta K, Hooton TM, Naber KG, et al. International clinical practice guidelines for the treatment of acute uncomplicated cystitis and pyelonephritis in women: a 2010 update by the infectious diseases society of America and the European society for microbiology and infectious diseases. Clin Infect Dis. 2011;52(5):e103–e120. doi:10.1093/cid/ciq257
6. Pédeboscq S, Issa N, Lahouati M, Labadie A, Pereyre S, Camou F. Impact of a 5-year antimicrobial stewardship program focusing on fluoroquinolone prescriptions. Int J Pharm Pract. 2021;29(6):616–621. doi:10.1093/ijpp/riab047
7. Ott SR, Allewelt M, Lorenz J, Reimnitz P, Lode H. German Lung Abscess Study Group. Moxifloxacin vs ampicillin/sulbactam in aspiration pneumonia and primary lung abscess. Infection. 2008;36(1):23–30. doi:10.1007/s15010-007-7043-6
8. Clinical and Laboratory Standards Institute. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria. CLSI Guideline M45.
9. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. CLSI Supplement M100.
10. Li X, Mariano N, Rahal JJ, Urban CM, Drlica K. Quinolone-resistant Haemophilus influenzae: determination of mutant selection window for ciprofloxacin, garenoxacin, levofloxacin, and moxifloxacin. Antimicrob Agents Chemother. 2004;48(11):4460–4462. doi:10.1128/AAC.48.11.4460-4462.2004
11. Pitondo-Silva A, Martins VV, Silva CFD, Stehling,EG. Conjugation between quinolone-susceptible bacteria can generate mutations in the quinolone resistance-determining region, inducing quinolone resistance. Int J Antimicrob Agents. 2015;45(2):119–123. doi:10.1016/j.ijantimicag.2014.07.018
12. Tamma PD, Aitken SL, Bonomo RA, Mathers AJ, van Duin D, Clancy CJ. Infectious Diseases Society of America Guidance on the Treatment of Extended-Spectrum β-lactamase Producing Enterobacterales (ESBL-E), Carbapenem-Resistant Enterobacterales (CRE), and Pseudomonas aeruginosa with Difficult-to-Treat Resistance (DTR-P. aeruginosa). Clin Infect Dis. 2021;72(7):e169–e183. doi:10.1093/cid/ciaa1478
13. Jwair NA, Al-Ouqaili MTS, Al-Marzooq F. Inverse Association between the Existence of CRISPR/Cas Systems with antibiotic resistance, extended spectrum β-lactamase and carbapenemase production in multidrug, extensive drug and pandrug-resistant Klebsiella pneumoniae. Antibiotics (Basel). 2023;12(6):980. doi:10.3390/antibiotics12060980
14. Doi Y, Park YS, Rivera JI, et al. Community-associated extended-spectrum β-lactamase-producing Escherichia coli infection in the United States. Clin Infect Dis. 2013;56(5):641–648. doi:10.1093/cid/cis942
15. Kohanski MA, DePristo MA, Collins JJ. Sublethal antibiotic treatment leads to multidrug resistance via radical-induced mutagenesis. Mol Cell. 2010;37(3):311–320. doi:10.1016/j.molcel.2010.01.003
16. Bergallo C, Jasovich A, Teglia O, et al. Safety and efficacy of intravenous tigecycline in treatment of community-acquired pneumonia: results from a double-blind randomized Phase 3 comparison study with levofloxacin. Diagn Microbiol Infect Dis. 2009;63(1):52–61. doi:10.1016/j.diagmicrobio.2008.09.001
17. Li Y, Zhu D, Peng Y, et al. A randomized, controlled, multicenter clinical trial to evaluate the efficacy and safety of oral sitafloxacin versus moxifloxacin in adult patients with community-acquired pneumonia. Curr Med Res Opin. 2021;37(4):693–701. doi:10.1080/03007995.2021.1885362
18. Thurber KM, Arnold JR, Narayanan PP, Dierkhising RA, Sampathkumar P. Comparison of intravenous and oral definitive antibiotic regimens in hospitalised patients with Gram-negative bacteraemia from a urinary tract infection. J Glob Antimicrob Resist. 2019;18:243–248. doi:10.1016/j.jgar.2019.03.013
19. Takayanagi N, Kagiyama N, Ishiguro T, Tokunaga D, Sugita Y. Etiology and outcome of community-acquired lung abscess. Respiration. 2010;80(2):98–105. doi:10.1159/000312404
20. Yagci D, Yoruk F, Azap A, Memikoglu O. Prevalence and risk factors for selection of quinolone-resistant Escherichia coli strains in fecal flora of patients receiving quinolone therapy. Antimicrob Agents Chemother. 2009;53(3):1287–1289. doi:10.1128/AAC.01228-08
21. Richard P, Delangle MH, Raffi F, Espaze E, Richet H. Impact of fluoroquinolone administration on the emergence of fluoroquinolone-resistant gram-negative bacilli from gastrointestinal flora. Clin Infect Dis. 2001;32(1):162–166. doi:10.1086/317551
22. de Lastours V, Chau F, Roy C, Larroque B, Fantin B. Emergence of quinolone resistance in the microbiota of hospitalized patients treated or not with a fluoroquinolone. J Antimicrob Chemother. 2014;69(12):3393–3400. doi:10.1093/jac/dku283
23. Fantin B, Duval X, Massias L, et al. Ciprofloxacin dosage and emergence of resistance in human commensal bacteria. J Infect Dis. 2009;200(3):390–398. doi:10.1086/600122
24. Wise R, Lockley RM, Webberly M, Dent J. Pharmacokinetics of intravenously administered ciprofloxacin. Antimicrob Agents Chemother. 1984;26(2):208–210. doi:10.1128/AAC.26.2.208
25. Fish DN, Chow AT. The clinical pharmacokinetics of levofloxacin. Clin Pharmacokinet. 1997;32(2):101–119. doi:10.2165/00003088-199732020-00002
26. Nakashima M, Uematsu T, Kosuge K, Umemura K, Hakusui H, Tanaka M. Pharmacokinetics and tolerance of DU-6859a, a new fluoroquinolone, after single and multiple oral doses in healthy volunteers. Antimicrob Agents Chemother. 1995;39(1):170–174. doi:10.1128/AAC.39.1.170
27. Wise R, Gee T, Marshall G, Andrews JM. Single-dose pharmacokinetics and penetration of BMS 284756 into an inflammatory exudate. Antimicrob Agents Chemother. 2002;46(1):242–244. doi:10.1128/AAC.46.1.242-244.2002
28. Lawrence LE, Wu P, Fan L, et al. The inhibition and selectivity of bacterial topoisomerases by BMS-284756 and its analogues. J Antimicrob Chemother. 2001;48(2):195–201. doi:10.1093/jac/48.2.195
29. Atac N, Kurt-Azap O, Dolapci I, et al. The Role of AcrAB–TolC efflux pumps on quinolone resistance of E. coli ST131. Curr Microbiol. 2018;75(12):1661–1666. doi:10.1007/s00284-018-1577-y
30. Thanassi DG, Cheng LW, Nikaido H. Active efflux of bile salts by Escherichia coli. J Bacteriol. 1997;179(8):2512–2518. doi:10.1128/jb.179.8.2512-2518.1997
31. Totsuka K, Sesoko S, Fukase H, Ikushima I, Odajima M, Niwayama Y. Pharmacokinetic study of lascufloxacin in non-elderly healthy men and elderly men. J Infect Chemother. 2020;26(3):231–239. doi:10.1016/j.jiac.2019.09.010
32. John TM, Deshpande A, Brizendine K, Yu PC, Rothberg MB. Epidemiology and outcomes of community-acquired Escherichia coli pneumonia. Open Forum Infect Dis. 2021;9(1):ofab597. doi:10.1093/ofid/ofab597
33. Neuman M. Comparative pharmacokinetic parameters of new systemic fluoroquinolones: a review. Chemioterapia. 1987;6(2):105–112.
34. Wiessler M, Pool BL. Mutagenic properties of N-cyclopropyl and N-allyl-N-nitroso compounds. Studies on the nature of alkylating species. Carcinogenesis. 1984;5(5):635–639. doi:10.1093/carcin/5.5.635
35. Al-Ouqaili MTS. Molecular detection and sequencing of SHV gene encoding for extended-spectrum β-lactamases produced by multidrug resistance some of the Gram-negative bacteria. Int J Green Pharm. 2018;12(4):S910–S918. doi:10.22377/ijgp.v12i04.2274
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.