Back to Journals » Journal of Inflammation Research » Volume 14
The Differential Expression of the Inflammasomes in Adipose Tissue and Colon Influences the Development of Colon Cancer in a Context of Obesity by Regulating Intestinal Inflammation
Authors Frühbeck G ![]() , Mentxaka A, Ahechu P, Gómez-Ambrosi J
, Mentxaka A, Ahechu P, Gómez-Ambrosi J ![]() , Ramírez B, Becerril S, Rodríguez A
, Ramírez B, Becerril S, Rodríguez A ![]() , Unamuno X, Cienfuegos JA, Casado M, Burrell MA, Martín M, Baixauli J
, Unamuno X, Cienfuegos JA, Casado M, Burrell MA, Martín M, Baixauli J ![]() , Valentí V, Moncada R, Reina G, Silva C, Catalán V
, Valentí V, Moncada R, Reina G, Silva C, Catalán V ![]()
Received 3 September 2021
Accepted for publication 17 November 2021
Published 1 December 2021 Volume 2021:14 Pages 6431—6446
DOI https://doi.org/10.2147/JIR.S335882
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Ning Quan
Gema Frühbeck,1– 4,* Amaia Mentxaka,1,2,* Patricia Ahechu,5 Javier Gómez-Ambrosi,1– 3 Beatriz Ramírez,1– 3 Sara Becerril,1– 3 Amaia Rodríguez,1– 3 Xabier Unamuno,1,2,6 Javier A Cienfuegos,5 Marcos Casado,1 María A Burrell,7 Marina Martín,7 Jorge Baixauli,5 Victor Valentí,5 Rafael Moncada,8 Gabriel Reina,9 Camilo Silva,4 Victoria Catalán1– 3
1Metabolic Research Laboratory, Clínica Universidad de Navarra, Pamplona, Spain; 2CIBER Fisiopatología de la Obesidad y Nutrición (CIBEROBN), Instituto de Salud Carlos III, Pamplona, Spain; 3Obesity and Adipobiology Group, Instituto de Investigación Sanitaria de Navarra (IdiSNA), Pamplona, Spain; 4Department of Endocrinology & Nutrition, Clínica Universidad de Navarra, Pamplona, Spain; 5Department of Surgery, Clínica Universidad de Navarra, Pamplona, Spain; 6Medical Engineering Laboratory, Clínica Universidad de Navarra, Pamplona, Spain; 7Department of Histology and Pathology, University of Navarra, Pamplona, Spain; 8Department of Anesthesia, Clínica Universidad de Navarra, Pamplona, Spain; 9Microbiology Laboratory, Clínica Universidad de Navarra, Pamplona, Spain
*These authors contributed equally to this work
Correspondence: Gema Frühbeck
Department of Endocrinology, Clínica Universidad de Navarra, Avda. Pío XII, 36, Pamplona, 31008, Spain
Tel +34 948 25 54 00 (ext. 4484)
Email [email protected]
Victoria Catalán
Metabolic Research Laboratory, Clínica Universidad de Navarra, Avda. Pío XII, 36, Pamplona, 31008, Spain
Tel +34 948 25 54 00 (ext. 5133)
Email [email protected]
Background: Inflammasomes maintain tissue homeostasis and their altered regulation in the colon, and the adipose tissue (AT) leads to chronic activation of inflammatory pathways promoting colon cancer (CC) development. We aimed to analyze the potential involvement of inflammasomes in obesity-associated CC.
Methods: Ninety-nine volunteers [61 with obesity (OB) and 38 normoponderal (NP)] further subclassified according to the approved protocol for the diagnosis of CC (58 without CC and 41 with CC) were included in the case–control study.
Results: CC (P< 0.01) and obesity (P< 0.01) were accompanied by increased mRNA levels of NLRP3, NLRP6, ASC, IL1B and NOD2 in VAT. Contrarily, patients with CC exhibited a downregulation of NLRP6 and IL18 in their colon. Additionally, we revealed that the decreased Nlrp1 (P< 0.05), Nlrp3 (P< 0.01) and Nlrp6 (P< 0.01) mRNA levels in the colon from obese rats significantly increase (P< 0.05) after caloric restriction. Adipocyte-conditioned media obtained from subjects with obesity reduced (P< 0.01) the mRNA of NLRP3 as well as molecules involved in maintaining the intestinal integrity (MUC2, CLDN1 and TJP1) and the anti-inflammatory factors FGF21, KLF4, and IL33 and in HT-29 cells. We also found that the knockdown of NLRP6 in HT-29 cells significantly upregulated (P< 0.05) the mRNA of NLRP1 and NLRP3 and inhibited (P< 0.05) the expression levels of MUC2. Finally, we showed that the incubation of HT-29 with Akkermansia muciniphila influence (P< 0.05) the inflammasome expression profile as well as intestinal integrity-related genes and aberrant inflammation.
Conclusions: These findings provide evidence that the downregulated levels of NLRP6 and IL18 in the colon from patients with CC may be responsible for a reduced intestinal-barrier integrity, triggering local inflammation, which in turn acts on the dysfunctional AT in obesity, increasing the expression of different inflammasome components and flaring up a vicious cycle of uncontrollable inflammatory cascades that favours a pro-tumorigenic microenvironment.
Keywords: inflammasome, NLRP, inflammation, colon cancer, obesity, adipose tissue, Akkermansia muciniphila
Corrigendum for this paper has been published.
Graphical Abstract:

Introduction
Colon cancer (CC) is the third most common cancer worldwide and the second in terms of mortality, with more than one-half of all cases and deaths attributable to modifiable risk factors including an unhealthy diet, excess body weight and physical inactivity.1 Despite a huge body of epidemiological knowledge supporting the link between excess adiposity and cancer, cellular and molecular mechanisms insights explaining this association are scarce.2 Different hallmarks of colon carcinogenesis are causally linked with obesity, being of special relevance to chronic and uncontrolled inflammation as well as the altered intestinal permeability and gut dysbiosis.3 Specifically, the innate immune response required to impair early carcinogenesis is also dysregulated in the obese state.4 In this sense, we previously demonstrated that the inflammasome, a cytosolic innate immune multi-protein complex, is involved in a diversity of metabolic diseases including obesity, type 2 diabetes (T2D) and nonalcoholic fatty liver disease (NAFLD) by regulating inflammation and extracellular matrix remodelling.5 In the gastrointestinal tract, the inflammasomes are crucially implicated in maintaining the homeostasis of gut epithelial due to their capacity to recognize diverse pathogenic insults.6,7 Although the main function of inflammasomes is to restrict damage, when inadequately controlled, intestinal inflammasomes promote a constant activation of inflammatory pathways prompting immune pathology, tissue damage and cancer development.7,8
The impact of the nucleotide-binding oligomerization domain (NOD)-like receptors (NLRPs) family on tumorigenesis has been revealed over the last decade.9,10 Inflammasomes exhibit complex and diverse roles in all stages of colon carcinogenesis, playing either pro-tumoral (perpetuating inflammation) or anti-tumoral (maintaining intestinal barrier integrity) functions and, therefore, conflicting results regarding CC development have been reported.10 These apparently differing functions have been suggested to be context-dependent and tissue-specific.11,12
The colon is the main organ for host-microbiome crosstalk and a disturbance of the gut microbiota not only influences the development and progression of CC, but also promotes the progression of other major conditions including obesity, metabolic syndrome and T2D.13,14 In this context, the inflammasomes tightly regulate intestinal microbiota and mice deficient in the different inflammasome components Nlrp3, Nlrp6 or Asc exhibit intestinal dysbiosis promoting gut inflammation, metabolic syndrome and CC development.15–17 Of note, the supplementation with Akkermansia muciniphila improved different metabolic parameters in both mice18 and humans.19
Obesity-associated sustained inflammation is a recognized risk factor for colon carcinogenesis. Whether excess adiposity has an impact on CC via a direct mechanism is widely twilight and the function of the inflammasomes in cancer remains controversial. Therefore, we hypothesized that obesity and colon cancer have an impact on the expression of the main components of the inflammasomes as well as their different effectors in the colon and the visceral AT (VAT) from patients with obesity and CC. The potential changes in the expression of the inflammasome components after weight loss were evaluated in the colon of a diet-induced obese rat model. The regulatory role of A. muciniphila in the expression of the inflammasome components and key inflammation- and intestinal integrity-associated factors in HT-29 cells was evaluated.
Materials and Methods
Patient Selection
Ninety-nine subjects [38 normoponderal (NP) and 61 with obesity (OB)] were included in the case–control study. Patients were also subclassified according to the approved protocol for the diagnosis of colon cancer (58 without CC and 41 with CC) and were included in the case–control study (Supplemental Table 1). VAT samples were obtained from volunteers undergoing Nissen fundoplication (for normoponderal volunteers), Roux-en-Y gastric bypass (RYGB) (for morbid obesity treatment) and curative resection for primary colon carcinoma (for CC treatment). The experimental design was approved, from an ethical and scientific standpoint, by the Clínica Universidad de Navarra’s Ethics Committee responsible for research (2018.094). The written informed consent of all participants was obtained. This study was conducted in accordance with the Declaration of Helsinki. A cohort of human colon total RNA samples (5 from normal colon tissue and 9 from CC tissue) were purchased from OriGene (Rockville, MD, USA). All the clinical information associated with each of these samples is available from the OriGene website.
Experimental Models of Diet-Induced Obesity
Surgical Procedures
Eight-week-old male Wistar rats (n=40) were fed ad libitum either a normal chow diet (ND) (n=20) (Diet 2014S, Harlan Laboratories Inc., Barcelona, Spain) or a high-fat diet (HFD) (n=20) (Diet F3282, Bio-Serv, Frenchtown, NJ, USA) for 16 weeks as previously described.20 Body weight and food intake were weekly registered. Diet-induced obesity (DIO) rats were randomized to be submitted to sham surgery (n=10), sleeve gastrectomy (SG) (n=10) or pair-fed (n=10) to the amount of food eaten by animals undergoing SG in order to differentiate possible effects of the decrease in food intake after surgery. Anesthesia and surgical procedures were performed as previously described.20 After interventions, animals were fed a ND.
Caloric Restriction
Ten-week-old male Wistar rats (n=40) (breeding house of the University of Navarra) were used in the study. Experimental animals were housed in the same conditions as described above. Rats were randomized in four groups (n=10) to analyze the effect of the diet [ND (Diet 2014S) or HFD (Diet F3282)] and the effect of a 25% of caloric restriction (ND-R and HFD-R, respectively).
After all the interventions, rats were sacrificed by decapitation, and the colon was dissected out and immediately stored at −80°C until next experiments. All experimental protocols were approved by the Ethics Committee for Animal Experimentation of the University of Navarra (049/10) and conformed to the European Guidelines for the Care and Use of Laboratory Animals (directive 2010/63/EU).
Blood Processing and Assays
Blood samples were collected by venipuncture after an overnight fast and centrifuged at 12,000 g for 15 min at 4°C to obtain serum and plasma. Glucose and triglycerides, as well as carcinoembryonic antigen (CEA) and C-reactive protein (CRP), were determined as previously described.21,22 An automated cell counter (Beckman Coulter, Inc., Fullerton, CA) was used to determine the white blood cell (WBC) count. Lipopolysaccharide-binding protein (LBP) (R & D Systems, Minneapolis, MN, USA), lactoferrin (R & D Systems), calgranulin A (S100A8) (R & D Systems) and IL-18 (R & D Systems) were measured by ELISA according to the manufacturer’s instructions. The limulus amoebocyte lysate assay (Lonza, Basel, Switzerland) was used to detect and quantify circulating levels of bacterial lipopolysaccharide (LPS).
RNA Isolation and Real-Time PCR
Total RNA isolation and purification were performed by homogenization with an Ultra-Turrax® T25 basic (IKA® Werke GmbH, Staufen, Germany) using TRIzol® Reagent (Invitrogen, Carlsbad, CA, USA) for colon samples and HT-29 cultured cells and QIAzol® Reagent (Qiagen, Hilden, Germany) for AT and then treated with DNase I (RNase Free DNase set, Qiagen). Transcript levels were determined by Real-Time PCR (7300 Real Time PCR System, Applied Biosystems, Foster City, CA, USA) as previously reported.21 Primers and probes (Supplemental Table 2) were designed using the software Primer Express 2.0 (Applied Biosystems).
Western-Blot Studies
VAT was homogenized, and protein content was measured as previously described.23 Protein samples (20 μg) were separated by SDS-PAGE (Criterion™ TGX™ Precast Gels, Bio-Rad Laboratories, Inc., Hercules, CA, USA) under denaturing conditions and transferred to a nitrocellulose membrane (Bio-Rad) for immunoblotting. Blots were incubated overnight at 4°C with rabbit monoclonal anti-NLRP-3 antibody (#13158, Cell Signaling Technology, Danvers, MA, USA) diluted 1:2500. The antigen–antibody complexes were visualized using HRP-conjugated secondary antibodies (1:5000) and the enhanced chemiluminescence ECL Plus detection system (Amersham Biosciences, Buckinghamshire, UK). The intensity of the bands was determined using the ChemiDocTM MP imaging system and the Image Lab 4.0.1 software (Bio-Rad) and normalized to total protein values.
Cell Cultures
Human stromovascular fraction cells were isolated from VAT from patients with obesity and were grown to confluence in six-well plates and differentiated into mature adipocytes, as previously described.23 The adipocyte conditioned media (ACM) were prepared by collecting the supernatant from differentiated adipocytes. After that, the supernatant was centrifuged and diluted (20% and 40%) in adipocyte medium. Two colorectal adenocarcinoma cell lines, HT-29 (HTB-38TM) and Caco-2 (HTB-37TM) were obtained from the ATCC® (Middlesex, UK) and cultured following manufacturer’s instructions. Caco-2 cell line was a generous gift from Dr. Amaya Azqueta of the University of Navarra. HT-29 cells were serum-starved for 2 h and then treated with increasing concentrations of LPS (10, 100 and 1000 ng/mL) (Merck), TNF-α (1, 10 and 100 ng/mL) (Merck), IL-4 (1, 10 and 100 ng/mL) (R&D systems), IL-13 (1, 10 and 100 ng/mL) (R&D systems), leptin (1, 10 and 100 ng/mL) (Merck), insulin (10, 100 and 1000 ng/mL) (Merck), CoCl2 (100 and 200 nmol/L) (Merck), palmitic acid (100 and 200 nmol/L) (Merck) and ACM (20% and 40%). Caco-2 cells were stimulated with ACM (20% and 40%).
HT-29 Transfection with siNLRP6
HT-29 cells were serum-starved for 2 h, and then two pairs of small interfering RNAs (siRNAs) for blocking the expression of NLRP6 (s46905 and s46907, Ambion, ThermoFisher Scientific, Waltham, MA, USA) were annealed and transfected into HT-29 cells (100 pmol/L siRNA/2×105 cells/well) using 40 nmol of Lipofectamine 2000 (Invitrogen, ThermoFisher). A scrambled siRNA was used as a negative control. Knockdown effectiveness was determined after 24 h by real-time PCR. The treatment with the NLRP6-s46905 resulted in an average knockdown of 49%, whereas no decreased expression was found after the transfection with the siNLRP6-s46907 (Supplemental Figure 1). Thus, the NLRP6-siRNA s46905 was selected for NLRP6 knocking-down studies. HT-29 cells were also cultured in the presence of LPS (1000 ng/mL) for 24 h.
Akkermansia muciniphila Cell Culture and HT-29 Treatment
A. muciniphila (ATCC® BAA-835TM) was cultured aseptically and anaerobically in 6 mL tubes of brain heart infusion broth (Becton Dickinson, Franklin Lakes, NJ, USA) at 37°C for 7 days. Cultures were washed and concentrated in anaerobic phosphate buffered saline (PBS) (Merck). Additionally, an identical quantity of A. muciniphila was heat-inactivated for 30 minutes at 70°C. The bacteria-conditioned medium (BCM) was obtained by collecting the supernatant. The BCM was centrifuged and diluted at 20% and 40% in McCoy’s 5A medium.
In the infection step, HT-29 cells were serum-starved for 2 h and incubated with alive and pasteurized A. muciniphila at a multiplicity of infection (MOI) of 100 as well as with the BCM (20% and 40%) for 4 hours. The brain heart infusion medium (diluted at 20% and 40% in McCoy’s 5A medium) was used as a control medium. The HT-29 media was collected after treatment with different stimuli, centrifuged at 200 g for 10 min and stored at −80°C. In order to assess the concentrations of mucin-2 in the media, a commercially available ELISA kit (MyBioSource, San Diego, CA, USA) was used according to the manufacturer’s instructions.
Statistical Analysis
Differences between groups were evaluated by Mann–Whitney U-test, two-way ANOVA and one-way ANOVA followed by Dunnett's or Tukey’s post hoc tests as appropriate. The analysis of covariance (ANCOVA) was used to analyze differences between groups adjusted for age. Pearson’s correlation coefficients (r) were used to evaluate the association between variables. The calculations were performed using the SPSS/Windows version 15.0 statistical package (SPSS, Chicago, IL), and the figures were created with GraphPad Prism version 8.3 (GraphPad Software, Inc., San Diego, CA). A P value <0.05 was considered statistically significant.
Results
VAT- and Colon-Specific Expression of the Inflammasome Components and Its Mediators in Obesity and CC
Clinical characteristics of the cohort are shown in Table 1. All markers of adiposity were increased (P<0.001) in patients with obesity compared to the normoponderal volunteers. Both obesity (P<0.001) and CC (P<0.001) were associated with increased levels of CRP. Although patients with CC showed a higher number of leucocytes (P<0.05) compared to those without CC, no differences in global WBC regarding obesity were observed.
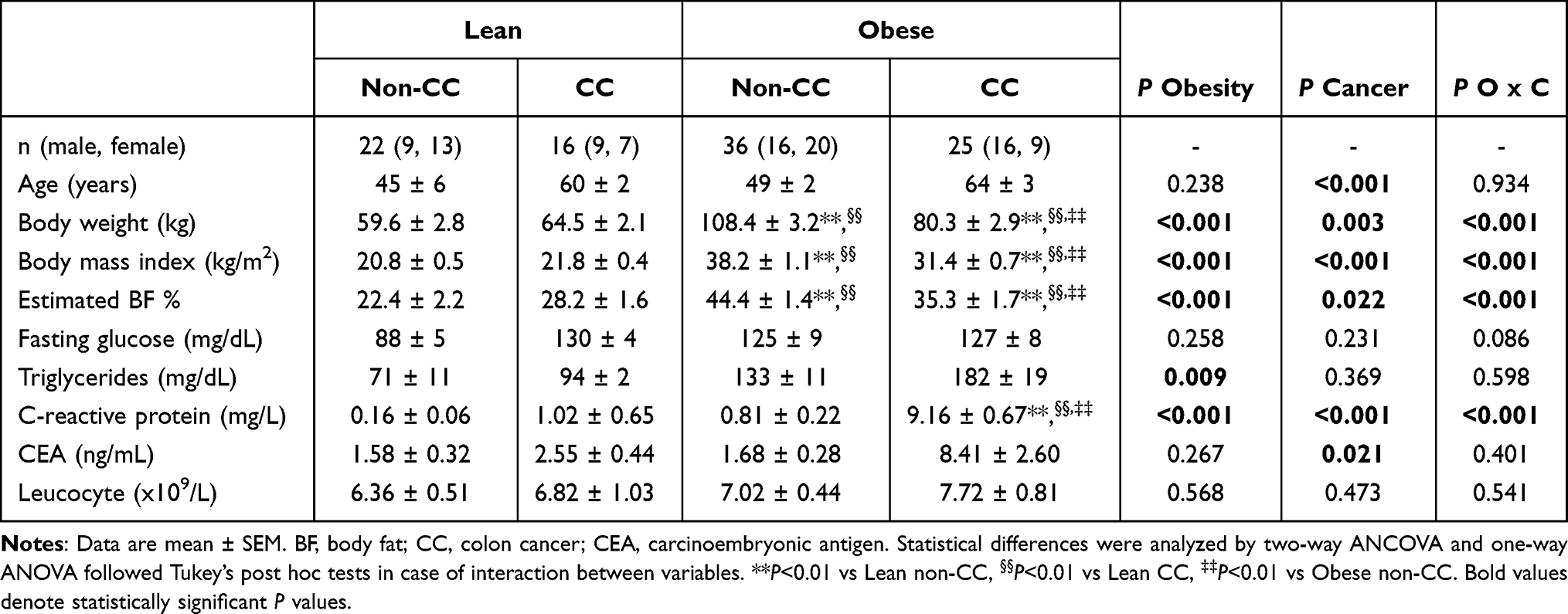 |
Table 1 Anthropometric and Biochemical Characteristics of Subjects Included in the Study |
First, we analyzed whether both obesity and CC influence the expression of the inflammasome pathway in VAT given its relevance in the development of obesity-associated CC. Although no changes were found regarding the expression of NLRP1, significant differences in mRNA levels of NLRP3, NLRP6 and ASC were observed, being significantly increased due to CC (P<0.05) and obesity (P<0.05) (Figure 1A-D). We also confirmed increased (P<0.05) protein expression levels of NLRP-3 in the VAT from patients with obesity (Supplemental Figure 2). Importantly, patients with CC showed an upregulation (P<0.05) of IL1B and IL18, the main inflammasome mediators, with the expression of IL1B being also increased (P<0.05) in patients with obesity (Figure 1E and F). Since NLRPs directly interact with NOD2, we also analyzed its mRNA levels, which were higher (P<0.05) in patients with obesity and CC (Figure 1G). Noteworthy, mRNA levels of ADIPOQ, an anti-inflammatory factor with proven decreased levels in obesity, were decreased in VAT from patients with obesity (P<0.01) and CC (P<0.05) (Figure 1H) and also negatively associated with BMI (r=−0.32; P=0.011). Gene expression levels of all inflammasome components were significantly associated between them (Supplemental Table 3). As major mediators of the innate immunity, NLRP3 and NOD2 were positively correlated with the percentage of monocytes (P<0.05) and negatively with the percentage of eosinophils (P<0.01). In addition, NLRP3 (r=0.44; P=0.012) and NOD2 (r=0.48; P=0.005) expression levels were positively correlated with the circulating concentrations of CRP.
 |
Figure 1 The increased gene expression levels of different components of the inflammasome in visceral adipose tissue (VAT) due to both, obesity and colon cancer (CC) suggest that obesity-associated VAT inflammation create a microenvironment favourable for CC development. Bar graphs show the mRNA levels of (A) NLRP1, (B) NLRP3, (C) NLRP6, (D) ASC, (E) IL1B and (F) IL18 (G) NOD2 and (H) ADIPOQ in VAT from lean subjects (LN) and volunteers with obesity (OB) classified according the presence or absence of CC. Bars represent the mean ± SEM. Differences between groups were analyzed by two-way ANOVA or one-way ANOVA followed by Tukey’s tests as appropriate. *P<0.05, **P<0.01 and ***P<0.01. (LN non CC: n=11; OB non CC: n= 34; LN CC: n=16; OB CC: n= 8). Bold values denote statistically significant P values. Abbreviations: ADIPOQ, adiponectin; ASC, apoptosis-associated speck-like protein containing a CARD; IL, interleukin; MUC2, mucin 2; NLRP, nucleotide-binding oligomerization domain, leucine rich repeat and pyrin; NOD2, nucleotide binding oligomerization domain containing 2. |
In light of the highly tissue-dependent expression of the inflammasomes, we analyzed the expression of the inflammasome components and their effectors in colon samples from patients with and without CC. Although we found a tendency towards an increased colonic gene expression of NLRP1, NLRP3 and IL1B in patients with CC, differences did not reach statistical significance. Importantly, the gene expression levels of NLRP6 and its mediator IL18 were downregulated (P<0.05) in the colon from patients with CC (Figure 2A). The results also showed reduced (P<0.01) MUC2 together with increased CLND1 gene expression levels in the colon from patients with CC compared with normal tissue colon samples (Figure 2B).
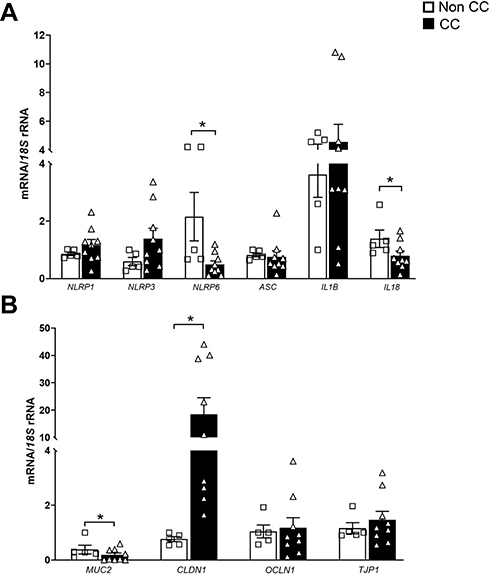 |
Figure 2 Impact of colon cancer (CC) on colonic inflammasome- and intestinal integrity-associated gene expression levels. Downregulation of gene expression levels of NLRP6, IL18 and MUC2 in the colon from patients with CC. mRNA levels of (A) NLRP1, NLRP3, NLRP6, ASC, IL1B and IL18 and (B) MUC2, CLDN, OCLN and TJP1 in the colon from patients with and without colon cancer. Bars represent the mean ± SEM. Differences between groups were analyzed by unpaired two-tailed Student’s t tests. *P<0.05. (non CC: n=5; CC: n= 9). Abbreviations: ASC, apoptosis-associated speck-like protein containing a CARD; CLDN1, claudin 1; IL, interleukin; OCLN, occludin; NLRP, nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin; NOD2, nucleotide-binding oligomerization domain containing 2; TJP1, tight junction protein 1. |
We further investigated the circulating levels of biomarkers of obesity and colon damage. In this line, subjects with obesity showed higher serum levels of S100A8 (P<0.001), lactoferrin (P<0.05), IL-18 (P<0.05) and the marker of LPS exposure LBP (P<0.001). However, no differences were found due to CC (Figure 3).
 |
Figure 3 Circulating concentrations of factors affecting intestinal barrier integrity in obesity and colon cancer (CC). Circulating concentrations of (A) limulus amebocyte lysate (LAL), (B) LPS-binding protein (LBP), (C) lactoferrin, (D) calgranulin A and (E) interleukin (IL)-18 of lean (LN) and obese (OB) volunteers classified according the presence or absence of CC. Differences between groups were analyzed by two-way ANOVA. Bold values denote statistically significant P values. LN non CC: n=13-15; OB non CC: n= 31; LN CC: n=16; OB CC: n= 18. |
Impact of Weight Loss Achieved by Surgery and Caloric Restriction on Colonic Expression of the Inflammasome Components in a DIO Rat Model
To gain further insight into the effect of obesity and weight loss on the regulation of the inflammasomes in the colon, we analyzed their expression levels in a DIO rat model submitted to SG and caloric restriction. Results showed decreased Nlrp1 (P<0.05), Nlrp3 (P<0.01) and Nlrp6 (P<0.01) mRNA levels in the colon from obese rats in both animal models (Figure 4A-C). Although an increase in the expression levels of the components of the inflammasome after surgery in rats under a ND was detected, differences were not statistically significant (Figure 4A-C). However, an increase (P<0.05) in the colonic mRNA levels of Nlrp1 after caloric restriction under both a ND and a HFD was observed together with an upregulation of Nlrp6 in rats under a restricted HFD (Figure 4C).
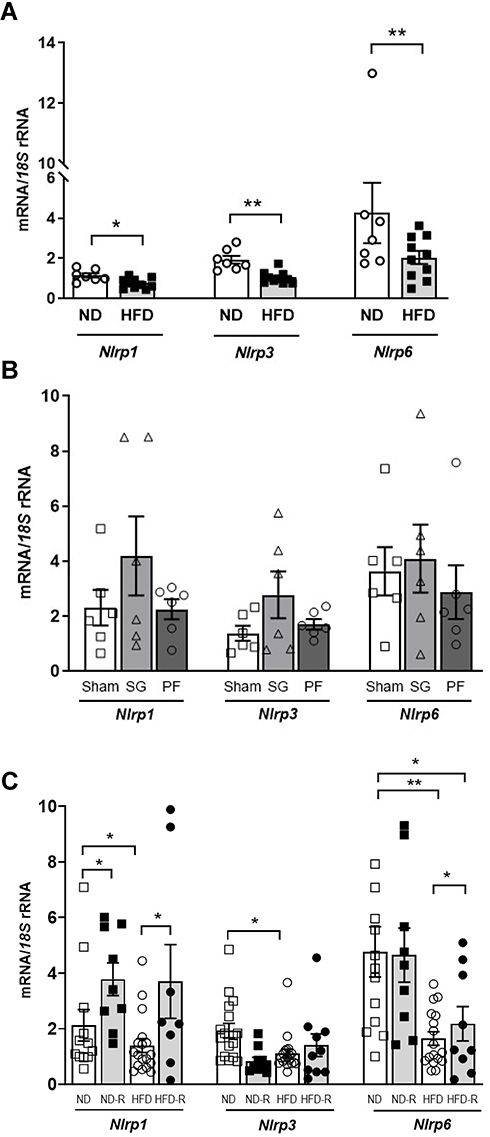 |
Figure 4 Impact of weight loss achieve by bariatric surgery and caloric restriction on the inflammasome components. (A) Gene expression levels of Nlrp1, Nlrp3 and Nlrp6 in the colon from rats fed a normal diet (ND) or a high-fat diet (HFD). (B) mRNA levels of Nlrp1, Nlrp3 and Nlrp6 in the colon from rats submitted to sleeve gastrectomy (SG) surgery and fed a ND. (C) Gene expression levels of Nlrp1, Nlrp3 and Nlrp6 in the colon from rats fed a normal diet (ND) or a high-fat diet (HFD) and a 25% of caloric restriction (ND-R and HFD-R). Bars represent the mean ± SEM. Unpaired Student’s t-test was used to observe differences between ND vs HFD control groups while the effect of the SG and diet was evaluated by a two-way ANOVA followed by Tukey’s tests. *P<0.05 and **P<0.01. Abbreviation: Nlrp, nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin. |
Effects of Inflammation-Related Factors and Hypoxia in the mRNA Levels of Inflammasome Components in Colon Cancer Cells
Since NLRPs recognize different inflammation-inducing stimuli, we further analyzed whether well-known inflammation-related markers that are increased in obesity influence the expression of the inflammasomes in HT-29 cells. LPS and TNF-α treatments induced opposite expression profiles of NLRP3 being increased (P<0.05) and decreased (P<0.05), respectively (Supplemental Figure 3A and B). Moreover, LPS enhanced (P<0.05) mRNA levels of NLRP6 and TNF-α upregulated the expression of NLRP1 (Supplemental Figure 3A and B). No differences on the expression of NLRPs in HT-29 cells after the treatment with the anti-inflammatory cytokine IL-13 were observed, but the expression of NLRP6 decreased after IL-4 stimulation (Supplemental Figure 3C and D). NLRP1 showed a robust upregulation (P<0.001) under hypoxic conditions mimicked by the divalent transition-metal ion cobalt in contrast with the downregulation observed for NLRP3 and NLRP6 (Supplemental Figure 3E). Whereas palmitic acid and leptin did not affect the expression of none of the components of the inflammasome, insulin promoted a decrease (P<0.05) of NLRP3 (Supplemental Figure 3F-H).
ACM from Patients with Obesity Influences the Expression Levels of NLRP3 as Well as Inflammation- and Intestinal-Related Factors in HT-29 and Caco-2 Cells
A dynamic interaction between adipocytes and tumor cells in an obesity context has been proposed. To address this question, we first examined how the ACM from subjects with obesity affects the expression of NLRPs and intestinal barrier integrity markers in HT-29 cells. Interestingly, we found a downregulation (P<0.01) in the mRNA of NLRP3 (Figure 5A). No differences were observed in other inflammasome components neither IL1B nor IL18. To gain further insight on the potential effects of ACM in tumor cells, we analyzed changes in intestinal integrity-related genes, finding a significant reduction of MUC2 (P<0.01), CLDN1 (P<0.05) and TJP1 (P<0.05) (Figure 5B). Metals constitute vital nutrients for bacterial growth. Importantly, the expression of antimicrobial proteins that chelate iron, zinc or manganese including TNFAIP9 (P<0.01), NGAL (P<0.05) and S100A9 (P<0.01) were increased after the ACM treatment (Figure 5C). The expression of the widely accepted inflammatory gene SPP1 (P<0.05) was also increased after the stimulation with the ACM (Figure 5C). No changes were found in TGFB and CCL2 expression levels after exposure to adipocyte-derived factors. Next, we analyzed the effect of ACM from subjects with obesity on the mRNA levels of anti-inflammatory factors, finding a downregulation of IL33 (P<0.01), FGF21 (P<0.01) and KLF4 (P<0.05) (Figure 5D). Changes in the mRNA levels of inflammasome components after ACM treatment were confirmed in Caco-2 cells, another common cell line used to reproduce the features of the tumoral bowel epithelium. We also found a downregulation (P<0.01) in the mRNA of NLRP3 (Supplemental Figure 4). Interestingly, the expressions of NLRP6 and MUC2 were not detected. After the ACM treatment, an upregulation (P<0.05) of IL1B and CLDN1 was also observed.
 |
Figure 5 Effect of adipocyte-conditioned media (ACM) on the gene expression levels of the inflammasome as well as on intestinal integrity- and inflammation-associated markers in HT-29 cells. Bar graphs show the effect of ACM (20 and 40%) from obese subjects incubated for 24 h on the transcript levels of (A) inflammasome-, (B) intestinal integrity-, (C) inflammation- and (D) anti-inflammatory-associated genes in HT-29 cells. Values are the mean ± SEM (n=6 per group). Differences between groups were analyzed by one-way ANOVA followed by Tukey’s tests. *P<0.05 and **P<0.01. (n=6 per group). Abbreviations: ADIPOQ, adiponectin; ASC, apoptosis-associated speck-like protein containing a CARD; CCL2, monocyte chemoattractant protein-1; CLDN1, claudin 1; FGF21, fibroblast growth factor 21; GLP1, glucagon-like peptide-1; GSDMD, gasdermin D; HMGB1, high mobility group box 1; IL, interleukin; KLF4, kruppel-like factor 4; MUC2, mucin 2; NGAL, lipocalin 2; NLRP, nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin; NOD2, nucleotide-binding oligomerization domain containing 2; OCLN, occludin; S100A9, S100 calcium-binding A9; SPP1, osteopontin; TGFB1, transforming growth factor-β; TLR4, toll-like receptor-4; TJP1, tight junction protein 1; TNFAIP9, tumor necrosis factor-α-induced protein 9. |
NLRP6 Silencing Results in Increased Expression of NLRP1 and NLRP3 in HT-29 Cells
On the basis that NLRP6 is downregulated in the colon from patients with CC, and given its relevance in maintaining intestinal homeostasis, we reduced its constitutive expression in HT-29 cells using a specific siRNA. As shown in Figure 6, siNLRP6 significantly increased (P<0.05) the expression of NLRP1 and NLRP3 compared to the control group, whereas the stimulation with LPS upregulated the expression of NLRP3 and IL1B. No differences were observed regarding the expression levels of IL18. Importantly, the inhibition of NLRP6 significantly reduced (P<0.05) the expression levels of MUC2 whereas no changes in other intestinal-integrity markers were observed.
 |
Figure 6 Effect of blocking NLRP6 expression in the gene expression levels of (A) inflammasome- and (B) intestinal integrity-related factors in colon cancer cells. The downregulation of NLRP6 increased the mRNA levels of NLRP1 and NLRP3. HT-29 cells were incubated with LPS (1000 ng/mL) and also were transfected with or without 100 pmol/L NLRP6 siRNA/2x105 cells/well for 24 h. Values are the mean ± SEM (n=6 per group). Differences between groups were analyzed by one-way ANOVA followed by Tukey’s tests. *P<0.05 vs scrambled siRNA. (n=6 per group). Abbreviations: ASC, apoptosis-associated speck-like protein containing a CARD; CLDN1, claudin 1; IL, interleukin; OCLN, occludin; NLRP, nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin; NOD2, nucleotide-binding oligomerization domain containing 2; TJP1, tight junction protein 1. |
Akkermansia muciniphila Improved the Inflammasome Expression Profile as Well as Intestinal Integrity-Related Genes and Aberrant Inflammation in HT-29 Cells
To explore the proposed antitumoral properties of A. muciniphila,24 HT-29 cells were cocultured with the bacteria alive, heat-inactivated and also with the bacterial-conditioned medium (BCM). As shown in Supplemental Table 4, gene expression levels of NLRP3 (P<0.001), ASC (P<0.001), IL1B (P<0.01) and HMGB1 (P<0.01) were increased in HT-29 cells treated with A. muciniphila alive. Importantly, we revealed an upregulation (P<0.05) in the mRNA of the intestinal integrity-related genes CLDN1 and MUC2. Heat-inactivated bacteria did not exert significant changes but the BCM upregulated NLRP1 (P<0.001), NLRP6 (P<0.001), CLDN1 (P<0.01) and MUC2 (P<0.01) together with a downregulation of NLRP3 (P<0.01) and HMGB1 (P<0.001). We also measured the secretion levels of MUC-2 into the culture medium after A. muciniphila treatment, finding an increased (P<0.01) secretion into the medium (Supplemental Figure 5).
Discussion
A huge body of knowledge supports the multifaceted roles of inflammasomes in obesity and its associated risk for CC development displaying both tumor-suppressive and pro-tumorigenic functions.9,25 The main finding of our investigation is that patients with CC show increased expression of different inflammasome components in VAT in the context of obesity together with downregulated levels of NLRP6 and IL18 in the colon.
Important studies have proposed that AT expansion in obesity is a key first step in promoting an enhanced state of inflammation that confers an additional risk for cancer development.26 Our experiments confirmed increased expression of NLRP3, NLRP6, ASC and NOD2 in VAT due to both, obesity and CC suggesting that obesity-associated VAT inflammation creates a microenvironment favourable for CC development. We also showed upregulated levels of IL1B and IL18 in the VAT from patients with obesity and CC, strengthening that the inflammasome-dependent production of these cytokines may influence colon tumorigenesis. In this line, we previously reported that the blocking of the expression of NLRP3 reduces the inflammation of the AT together with an important attenuation in fibrosis controlling the development of obesity-associated comorbidities, including T2D or NAFLD.5 Moreover, a pathogenic role of IL-1β as an important player in colon carcinogenesis has been previously proposed.26 Low levels of adiponectin constitute a risk factor for obesity-associated cancers including CC and, accordingly, we found decreased levels of adiponectin in the VAT from subjects with obesity and CC. Results obtained regarding the expression levels of inflammasomes in the colon indicate that NLRP6 and IL18 were downregulated in patients with CC, identifying opposed functions for both molecules in VAT and colon, probably due to their highly tissue-specific expression. Certainly, the characteristics and functions of the inflammasome complexes depend on the simultaneous expression of their components in the same cell type of inflamed tissues at the same time.12,27 In this line, ASC, an inflammasome component, acts as a tumor promoter in the myeloid cells infiltrating the tumor being required for the synthesis of IL-1β and also functions as a tumor suppressor in the epithelial skin tumor cells.28 The NLRP-6 protein is mainly expressed in the intestine29 and thus, the homeostasis of the intestinal epithelium is extremely dependent on NLRP-6 with its uncontrolled expression critically affecting the susceptibility to intestinal bowel disease (IBD). Importantly, the protein expression levels of NLRP-1, −3 and −6 are reduced in patients with CC and the loss of NLRP-6 and IL-18 have been associated with an increased mortality.30 The expression of NLRP6 and its effector IL18 are closely linked. Nlrp6-knockout mice exhibit reduced circulating levels of IL-18 in physiological conditions and after chemical-induced colitis and colitis-associated tumorigenesis.15,31 NLRP-6 signaling protects against colitis-associated tumor development by decreasing the epithelial barrier permeability and by inducing immune responses against different microbes.15,29,31 In this regard, the outer mucosal layer of the colonic epithelium that prevents the direct exposure to invading commensals is covered by mucin and, NLRP-6 is critical for an appropriate mucin secretion.32 Muc2-knockout mice showed impaired secretion of mucus promoting bacterial colonization of the crypts, inflammation and carcinogenesis.33 Accordingly, we found decreased expression of MUC2 in the colon from subjects with CC accompanied with increased expression of CLDN1. The complex role of CLDN1 expression in CC has been exhaustively reviewed,34 with some studies showing upregulated and others decreased expression levels. The higher levels of CLDN1 found in our study may reflect an invasive and metastatic behavior of tumor cells as well as a poor prognosis.35
A pioneering study by Elinav et al15 revealed a significant alteration in the microbiota composition from Nlrp6-, Asc- and Il18-knockout mice compared to that of wild-type mice. The lack of Nlrp6 together with environmental microbial diversity have been shown to promote not only dysbiosis but also a diminished intestinal-barrier integrity that triggered inflammation, intimately linked to tumor growth.36 Indeed, metabolic endotoxemia is strongly related with the development of metabolic alterations including obesity, insulin resistance and liver steatosis.37,38 In this line, although no effects of CC were observed in our cohort, patients with obesity exhibited increased circulating levels of factors of loss of the intestinal integrity barrier, including LPB, S100A8 and lactoferrin. These gut-derived endotoxins may function as potential priming and activating danger signals for the inflammasomes in VAT, driving a vicious cycle and the perpetuation of inflammasome-mediated chronic inflammation and insulin resistance and prompting CC development.
We observed that the expression of Nlrp1, 3 and 6 was decreased in rats fed a HFD. Although weight loss achieved by SG tended to increase the expression of the inflammasome components, differences were not statistically significant. Previous studies had demonstrated that the reduced intestinal Nlrp6 levels in obesity may be efficiently reversed by RYGB.39 The lack of differences found in our study may be explained by the inherent variations between both bariatric surgery techniques as well as by the specific changes in gut microbiota after surgery since without microbiota changes, the induction of Nlrp6 was diminished.39 In addition, bariatric surgery has been described to enhance glucose tolerance via decreasing Nlrp3 inflammasome signaling in the AT from obese rats.40 Similar phenotypes have been reported in caloric restriction studies41 and, in our study, an increased expression of Nlrp1 and Nlrp6 in the colon after food intake restriction was also observed. An increased gastrointestinal permeability together with higher circulating concentrations of LPS have been previously reported in rats with HFD-induced obesity.42,43 Furthermore, rats undergoing Roux-en-Y gastric bypass exhibited lower LPS and inflammatory cytokine concentrations than rats in the control group, suggesting that RYGB decrease the degree of endotoxemia, being also associated with strong intestinal tight junctions and an improved intestinal barrier integrity.44
To decipher the inflammasome activation pathways in CC, HT-29 cells were treated with different endogenous and exogenous pathogenic insults. While NLRP3 and NLRP6 showed the same trend in their expression levels after the treatment with LPS, TNF-α, CoCl2 and insulin, NLRP1 displayed an opposite tendency. The complex and differential regulatory pathways to promote not only a balanced but also an efficient immune response mediated by the inflammasomes have been extensively reviewed by Latz et al.45 The identification of upstream mechanisms is a very interesting field to develop novel strategies for the activation or inhibition of individual inflammasomes.
AT dynamically participates in cancer development not only as an endocrine organ releasing adipokines, proinflammatory cytokines, and extracellular matrix factors but also acting as a reservoir of energy.2 We found an imbalance in inflammation and intestinal barrier integrity markers after the treatment of HT-29 cells with the adipocyte-derived factors from patients with obesity. The decreased levels of NLRP3 found in the in vitro study suggest a dysregulation of immunosurveillance in HT-29 cells favouring immune escape and limiting the recruitment of antitumor cells.30 Additionally, we found a reduction in the expression of crucial genes for the maintenance of epithelial barrier integrity, including MUC2, CLDN1 and TJP1. An imbalance in the levels of tight junction proteins has been proposed as an initial event in the pathogenesis of IBD.46 In line, ACM stimulates a proinflammatory profile increasing the levels of the inflammatory molecules TNFAIP9 and SPP1 as well as a reduction of the anti-inflammatory markers IL33, FGF21 and KLF4. Iron and zinc constitute nutrients for bacterial growth and the increased levels of NGAL and S100A9 observed in our study may reflect a strategy to prevent bacterial growth by limiting the availability of iron and zinc, respectively as a response to inflammation.47 Defining the mechanisms that underline this active interaction, especially in the context of obesity, may help to identify novel therapeutic strategies. The downregulation of NLRP3 after the treatment with ACM was confirmed in Caco-2, another colon cancer cell line. On the contrary, gene expression levels of NLRP6 and MUC2 were not detected and IL1B and CLDN1 were increased. Differences exist between Caco-2 and HT-29, the most widely human intestinal cell lines used for in vitro attachment and mechanistic studies. While Caco-2 cells form polarized cells with high homology to enterocytes, HT-29 cells are more heterogeneous, containing a small proportion of mucus-secreting cells and columnar absorptive cells.48 The development of co-cultures comprising both Caco-2 and HT29 cells has been proposed as an optimal strategy to find an in vitro model to mimic the intestinal epithelium.49
Although the maturation and activation of IL-1β and IL-18 via the NLRP6 inflammasome has been described in macrophages,50 gene expression levels of both interleukins remain unaltered after the inhibition of NLRP6 in our in vitro experiments. Similarly, Nlrp6-deficient mice did not exhibit changes in their circulating levels of IL-1β.31 Of note, we found that NLRP1 and NLRP3 were upregulated after the treatment with siNLRP6 in HT-29 cells. In this line, the expression of Nlrp3 in the liver from Nlrp6-knockout mice was upregulated, suggesting a compensatory mechanism of Nlrp3 activation upon Nlrp6 deficiency.29,51 The finding of increased levels of NLRP1 and NLRP3 potentially explains the unaltered levels of IL1B and IL18 found in HT-29 cells. Importantly, a downregulation of MUC2 was observed after siNLRP6 treatment reinforcing the role of NLRP6 in mucin release. The exocytosis of mucin 2 from sentinel goblet cells by NLRP6 activation, and therefore, the protection of the colonic crypt from bacterial intruders has been elegantly described by Birchenough et al.52 In accordance, Nlrp6−/− mice exhibited a failure of exocytose mucin granules due to impaired autophagy and hyperplasia of goblet cells.32
The drastic shift in the gut microbiome of patients with obesity and T2D influences the impaired maintenance of the intestinal barrier integrity.53 Among these, A. muciniphila is a beneficial bacterial species with reduced levels of obesity and its related comorbidities.54 The administration of A. muciniphila and its derived components has demonstrated to improve the metabolic profile and diet-induced obesity in terms of body weight gain and body composition in both animal models and humans,18,19 as well as the symptoms of DSS-induced colitis in mice.55,56 Lee et al demonstrated that the administration of Lactobacillus paracasei induced changes in the gut microbiota composition in HFD-fed rats being associated with improvements in gut barrier functions as well as in local and systemic inflammation.57 Bariatric surgery-induced weight loss has also been associated with an enrichment in the abundance of A. muciniphila.43 Our results showed that the stimulation of HT-29 with alive A. muciniphila had a stronger effect compared with the heat-inactivated bacteria, increasing the expression levels of NLRP3 and ASC. On the other hand, the bacteria-derived medium induced a strong upregulation of the expression of NLRP1, NLRP6 and MUC2 together with a decrease of the alarmin HMGB1. In addition, increased levels of secreted mucin-2 were detected after the treatment with the bacteria-derived medium. In this regard, Amuc_1100, a protein isolated from A. muciniphila enhanced the intestinal barrier.18 The increase in NLRP expression after A. muciniphila may reflect a better mechanism to recognize derived PAMPs and DAMPs to inhibit persistent and undesirable inflammatory responses.
This study has some limitations. Although the number of subjects in the groups may appear somewhat limited, the detailed clinical, biochemical and metabolic characterization of our subjects has to be stressed. Patients were carefully selected to avoid confounding factors and the groups encompassed highly homogeneous patients that allowed them to perform a robust analysis. Further prospective studies to assess the involvement of inflammasomes in the obesity-associated colon cancer may contribute to better elucidate the role of this protein complex in the context of obesity.
The downregulated levels of NLRP6 and IL18 in the colon from patients with CC may be responsible for a diminished intestinal-barrier integrity, triggering local inflammation, which in turn acts on dysfunctional adipocytes in obesity, increasing the expression of different inflammasome components and flaring up a vicious cycle of uncontrollable inflammatory cascades that favour a pro-tumorigenic microenvironment (Figure 7). Since the colon is the predominant site for host–microbiome interactions, a reduction in metabolic endotoxemia through the improvement of the gut barrier permeability combined with strategies to restore the functions of immunosurveillance of inflammasome components could represent an interesting target to identify and treat patients with obesity at increased risk for CC development.
 |
Figure 7 Proposed mechanism in the crosstalk between visceral adipose tissue (VAT) and colon in the development of colon cancer (CC) in an obesity context. The downregulated levels of NLRP6 and IL18 in colon cancer (CC) may be responsible of a diminished intestinal-barrier integrity, triggering local inflammation which in turn acts on dysfunctional adipocytes increasing the expression of NLRP3, NLRP6 and its pro-inflammatory mediators IL1B and IL18 promoting a vicious cycle of uncontrolled inflammatory cascades that favours a pro-tumorigenic microenvironment for CC development. Abbreviations: IL, interleukin; MUC2, mucin 2; NLRP, nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin. |
Ethics Approval
The experimental design was approved, from an ethical and scientific standpoint, by the Clínica Universidad de Navarra’s Ethics Committee responsible for research (2018.094). This study was conducted in accordance with the Declaration of Helsinki.
All experimental protocols were approved by the Ethics Committee for Animal Experimentation of the University of Navarra (049/10) and conformed to the European Guidelines for the Care and Use of Laboratory Animals (directive 2010/63/EU).
Acknowledgments
The authors gratefully acknowledge the valuable collaboration of all the members of the Multidisciplinary Obesity Team, Clínica Universidad de Navarra, Pamplona, Spain and Dr. Amaya Azqueta, University of Navarra, Pamplona, Spain.
Funding
Funded by Instituto de Salud Carlos III (project PI19/73000785, PI20/00080 and PI20/00927), Gobierno de Navarra (10/2018), and CIBEROBN. Funding sources had no role in manuscript writing or the decision to submit it for publication.
Disclosure
Prof. Dr. Gema Frühbeck reports grants from Spanish Health Institute ISCIII, during the conduct of the study. The authors report no other potential conflicts of interest in this work. The results were partially presented at the 28th European Congress on Obesity.
References
1. Siegel RL, Miller KD, Goding Sauer A, et al. Colorectal cancer statistics, 2020. CA Cancer J Clin. 2020;70:145–164.
2. Park J, Morley TS, Kim M, Clegg DJ, Scherer PE. Obesity and cancer–mechanisms underlying tumour progression and recurrence. Nat Rev Endocrinol. 2014;10:455–465.
3. Terzic J, Grivennikov S, Karin E, Karin M. Inflammation and colon cancer. Gastroenterology. 2010;138(2101–14):e5.
4. Catalán V, Gómez-Ambrosi J, Rodríguez A, Frühbeck G. Adipose tissue immunity and cancer. Front Physiol. 2013;4:275.
5. Unamuno X, Gómez-Ambrosi J, Ramírez B, et al. NLRP3 inflammasome blockade reduces adipose tissue inflammation and extracellular matrix remodeling. Cell Mol Immunol. 2021;18:1045–1057.
6. Lamkanfi M, Dixit VM. Mechanisms and functions of inflammasomes. Cell. 2014;157:1013–1022.
7. Zmora N, Levy M, Pevsner-Fishcer M, Elinav E. Inflammasomes and intestinal inflammation. Mucosal Immunol. 2017;10:865–883.
8. Palazon-Riquelme P, Lopez-Castejon G. The inflammasomes, immune guardians at defence barriers. Immunology. 2018;155:320–330.
9. Karki R, Kanneganti TD. Diverging inflammasome signals in tumorigenesis and potential targeting. Nat Rev Cancer. 2019;19:197–214.
10. Karki R, Man SM, Kanneganti TD. Inflammasomes and cancer. Cancer Immunol Res. 2017;5:94–99.
11. Drexler SK, Yazdi AS. Complex roles of inflammasomes in carcinogenesis. Cancer J. 2013;19:468–472.
12. Petrilli V. The multifaceted roles of inflammasome proteins in cancer. Curr Opin Oncol. 2017;29:35–40.
13. Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444:1022–1023.
14. Tilg H, Zmora N, Adolph TE, Elinav E. The intestinal microbiota fuelling metabolic inflammation. Nat Rev Immunol. 2020;20:40–54.
15. Elinav E, Strowig T, Kau AL, et al. NLRP6 inflammasome regulates colonic microbial ecology and risk for colitis. Cell. 2011;145:745–757.
16. Hirota SA, Ng J, Lueng A, et al. NLRP3 inflammasome plays a key role in the regulation of intestinal homeostasis. Inflamm Bowel Dis. 2011;17:1359–1372.
17. Levy M, Thaiss CA, Katz MN, Suez J, Elinav E. Inflammasomes and the microbiota-partners in the preservation of mucosal homeostasis. Semin Immunopathol. 2015;37:39–46.
18. Plovier H, Everard A, Druart C, et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat Med. 2017;23:107–113.
19. Depommier C, Everard A, Druart C, et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat Med. 2019;25:1096–1103.
20. Rodríguez A, Becerril S, Valentí V, et al. Short-term effects of sleeve gastrectomy and caloric restriction on blood pressure in diet-induced obese rats. Obes Surg. 2012;22:1481–1490.
21. Catalán V, Gómez-Ambrosi J, Rodríguez A, et al. IL-32α-induced inflammation constitutes a link between obesity and colon cancer. Oncoimmunology. 2017;6:e1328338.
22. Gómez-Ambrosi J, Salvador J, Rotellar F, et al. Increased serum amyloid A concentrations in morbid obesity decrease after gastric bypass. Obes Surg. 2006;16:262–269.
23. Rodríguez A, Catalán V, Gómez-Ambrosi J, et al. Insulin- and leptin-mediated control of aquaglyceroporins in human adipocytes and hepatocytes is mediated via the PI3K/Akt/mTOR signaling cascade. J Clin Endocrinol Metab. 2011;96:E586–97.
24. Wang L, Tang L, Feng Y, et al. A purified membrane protein from Akkermansia muciniphila or the pasteurised bacterium blunts colitis associated tumourigenesis by modulation of CD8(+) T cells in mice. Gut. 2020;69:1988–1997.
25. Stienstra R, Tack CJ, Kanneganti TD, Joosten LA, Netea MG. The inflammasome puts obesity in the danger zone. Cell Metab. 2012;15:10–18.
26. Deng T, Lyon CJ, Bergin S, Caligiuri MA, Hsueh WA. Obesity, inflammation, and cancer. Annu Rev Pathol. 2016;11:421–449.
27. Schroder K, Tschopp J. The inflammasomes. Cell. 2010;140:821–832.
28. Drexler SK, Bonsignore L, Masin M, et al. Tissue-specific opposing functions of the inflammasome adaptor ASC in the regulation of epithelial skin carcinogenesis. Proc Natl Acad Sci U S A. 2012;109:18384–18389.
29. Zheng D, Kern L, Elinav E. The NLRP6 inflammasome. Immunology. 2021;162:281–289.
30. Domblides C, Soubeyran I, Lartigue L, et al. Prognostic role of inflammasome components in human colorectal cancer. Cancers. 2020;12:3500.
31. Chen GY, Liu M, Wang F, Bertin J, Nunez G. A functional role for Nlrp6 in intestinal inflammation and tumorigenesis. J Immunol. 2011;186:7187–7194.
32. Wlodarska M, Thaiss CA, Nowarski R, et al. NLRP6 inflammasome orchestrates the colonic host-microbial interface by regulating goblet cell mucus secretion. Cell. 2014;156:1045–1059.
33. Velcich A, Yang W, Heyer J, et al. Colorectal cancer in mice genetically deficient in the mucin Muc2. Science. 2002;295:1726–1729.
34. Ouban A. Claudin-1 role in colon cancer: an update and a review. Histol Histopathol. 2018;33:1013–1019.
35. Singh AB, Sharma A, Smith JJ, et al. Claudin-1 up-regulates the repressor ZEB-1 to inhibit E-cadherin expression in colon cancer cells. Gastroenterology. 2011;141:2140–2153.
36. Gagliani N, Hu B, Huber S, Elinav E, Flavell RA. The fire within: microbes inflame tumors. Cell. 2014;157:776–783.
37. Cani PD, Amar J, Iglesias MA, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761–1772.
38. Damms-Machado A, Louis S, Schnitzer A, et al. Gut permeability is related to body weight, fatty liver disease, and insulin resistance in obese individuals undergoing weight reduction. Am J Clin Nutr. 2017;105:127–135.
39. Wang G, Wang Q, Bai J, et al. Upregulation of intestinal NLRP6 inflammasomes after Roux-en-Y gastric bypass promotes gut immune homeostasis. Obes Surg. 2020;30:327–335.
40. Mocanu AO, Mulya A, Huang H, et al. Effect of Roux-en-Y gastric bypass on the NLRP3 inflammasome in adipose tissue from obese rats. PLoS One. 2015;10:e0139764.
41. Vandanmagsar B, Youm YH, Ravussin A, et al. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat Med. 2011;17:179–188.
42. de La Serre CB, Ellis CL, Lee J, Hartman AL, Rutledge JC, Raybould HE. Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am J Physiol Gastrointest Liver Physiol. 2010;299:G440–8.
43. Lu C, Li Y, Li L, et al. Alterations of serum uric acid level and gut microbiota after Roux-en-Y gastric bypass and sleeve gastrectomy in a hyperuricemic rat model. Obes Surg. 2020;30:1799–1807.
44. Guo Y, Liu CQ, Liu GP, Huang ZP, Zou DJ. Roux-en-Y gastric bypass decreases endotoxemia and inflammatory stress in association with improvements in gut permeability in obese diabetic rats. J Diabetes. 2019;11:786–793.
45. Latz E, Xiao TS, Stutz A. Activation and regulation of the inflammasomes. Nat Rev Immunol. 2013;13:397–411.
46. Berkes J, Viswanathan VK, Savkovic SD, Hecht G. Intestinal epithelial responses to enteric pathogens: effects on the tight junction barrier, ion transport, and inflammation. Gut. 2003;52:439–451.
47. Zeng MY, Inohara N, Nunez G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017;10:18–26.
48. Gagnon M, Zihler Berner A, Chervet N, Chassard C, Lacroix C. Comparison of the Caco-2, HT-29 and the mucus-secreting HT29-MTX intestinal cell models to investigate Salmonella adhesion and invasion. J Microbiol Methods. 2013;94:274–279.
49. Ferraretto A, Bottani M, De Luca P, et al. Morphofunctional properties of a differentiated Caco2/HT-29 co-culture as an in vitro model of human intestinal epithelium. Biosci Rep. 2018;38:2.
50. Hara H, Seregin SS, Yang D, et al. NLRP6 inflammasome recognizes lipoteichoic acid and regulates gram-positive pathogen infection. Cell. 2018;175(1651–64):e14.
51. Li M, Chen Y, Shi J, et al. NLRP6 deficiency aggravates liver injury after allogeneic hematopoietic stem cell transplantation. Int Immunopharmacol. 2019;74:105740.
52. Birchenough GM, Nystrom EE, Johansson ME, Hansson GC. A sentinel goblet cell guards the colonic crypt by triggering Nlrp6-dependent Muc2 secretion. Science. 2016;352:1535–1542.
53. Cani PD, Osto M, Geurts L, Everard A. Involvement of gut microbiota in the development of low-grade inflammation and type 2 diabetes associated with obesity. Gut Microbes. 2012;3:279–288.
54. Everard A, Belzer C, Geurts L, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci USA. 2013;110:9066–9071.
55. Chelakkot C, Ghim J, Ryu SH. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp Mol Med. 2018;50:103.
56. Kang CS, Ban M, Choi EJ, et al. Extracellular vesicles derived from gut microbiota, especially Akkermansia muciniphila, protect the progression of dextran sulfate sodium-induced colitis. PLoS One. 2013;8:e76520.
57. Lee S, Kirkland R, Grunewald ZI, Sun Q, Wicker L, de La Serre CB. Beneficial effects of non-encapsulated or encapsulated probiotic supplementation on microbiota composition, intestinal barrier functions, inflammatory profiles, and glucose tolerance in high fat fed rats. Nutrients. 2019;11.
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.