Back to Journals » Nature and Science of Sleep » Volume 14
The Combination of Betahistine and Oxybutynin Increases Respiratory Control Sensitivity (Loop Gain) in People with Obstructive Sleep Apnea: A Randomized, Placebo-Controlled Trial
Authors Messineo L ![]() , Loffler K
, Loffler K ![]() , Chiang A
, Chiang A ![]() , Osman A, Taranto-Montemurro L, Eckert DJ
, Osman A, Taranto-Montemurro L, Eckert DJ ![]()
Received 22 February 2022
Accepted for publication 16 May 2022
Published 7 June 2022 Volume 2022:14 Pages 1063—1074
DOI https://doi.org/10.2147/NSS.S362205
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Ahmed BaHammam
Ludovico Messineo,1,2 Kelly Loffler,1 Alan Chiang,3 Amal Osman,1 Luigi Taranto-Montemurro,2 Danny J Eckert1
1Adelaide Institute for Sleep Health, Flinders University, Adelaide, SA, Australia; 2Division of Sleep and Circadian Disorders, Departments of Medicine and Neurology, Brigham and Women’s Hospital and Harvard Medical School, Boston, MA, USA; 3Neuroscience Research Australia, Randwick, NSW, Australia
Correspondence: Ludovico Messineo, Division of Sleep and Circadian Disorders, Departments of Medicine and Neurology, Brigham and Women’s Hospital and Harvard Medical School, 221 Longwood Avenue, Boston, MA, USA, Email [email protected]
Rationale: There are widespread histaminergic projections throughout the brain, including hypoglossal nuclei, that modulate pharyngeal muscle tone and respiratory control. Hence, histaminergic stimulation pharmacologically may increase pharyngeal muscle tone and stabilize respiratory control (loop gain) to reduce obstructive sleep apnea (OSA) severity. Antimuscarinics also increase REM pharyngeal muscle tone in rats. Thus, a combination of histaminergic and anti-muscarinic drugs may be a novel target for OSA pharmacotherapy. However, this has not been investigated. Accordingly, we aimed to test the effects of betahistine (Beta), an H3-autoreceptor antagonist which thereby increases histamine levels, in combination with the antimuscarinic oxybutynin (Oxy), on OSA severity, OSA endotypes, polysomnography parameters and next-day sleepiness and alertness.
Methods: Thirteen adults with OSA received either Beta-Oxy (96– 5mg) or placebo according to a randomized, crossover, double-blind design, prior to polysomnography. Participants completed the Karolinska Sleep Scale and Leeds Sleep Evaluation Questionnaire and a driving simulation task to quantify next-day sleepiness and alertness. OSA endotypes were estimated through validated algorithms using polysomnography.
Results: Compared to placebo, Beta-Oxy increased respiratory control sensitivity (loop gain) (0.52[0.24] vs 0.60[0.34], median [IQR], P = 0.021) without systematically changing OSA severity (34.4± 17.2 vs 40.3± 27.3 events/h, mean±SD, P = 0.124), sleep efficiency, arousal index or markers of hypoxemia. Beta-Oxy was well tolerated and did not worsen next-day sleepiness/alertness.
Conclusion: Rather than stabilize breathing during sleep, Beta-Oxy increases loop gain, which is likely to be deleterious for most people with OSA. However, in certain conditions characterized by blunted respiratory control (eg, obesity hypoventilation syndrome), interventions to increase loop gain may be beneficial.
Keywords: pharmacotherapy, respiratory control, histamine, sleep disordered breathing, upper airway, endotyping
Plain Language Summary
In light of observations that histaminergic receptors are highly expressed at the hypoglossal level and that antimuscarinics increase pharyngeal muscle tone in REM sleep, we tested the combination of betahistine (Beta), to increase histamine levels, with the antimuscarinic oxybutynin (Oxy) on obstructive sleep apnea (OSA) severity and endotypes. Beta-Oxy increased the sensitivity of the respiratory control system (loop gain) in the absence of an effect on AHI, which makes this combination unsuitable for most OSA patients, but may be promising for disorders characterized by reduced chemosensitivity.
Introduction
Obstructive sleep apnea (OSA) is a common breathing disorder characterized by repetitive narrowing or occlusion of the upper airway during sleep.1 Impairment in the anatomical components of the upper airway (eg, a narrow/collapsible pharynx) and other physiological traits/endotypes, such as respiratory control stability/loop gain, the respiratory arousal threshold, and upper airway muscle function, play a major role in mediating the propensity for OSA.2 Impaired upper airway muscle function or elevated loop gain, via destabilization of upper airway patency and breathing, are estimated to be key pathogenic contributors for more than half of all OSA patients.2 Current treatments however, such as continuous positive airway pressure (CPAP), mandibular advancement devices, and surgery, primarily target the anatomical endotype, with variable efficacy, compliance and patient outcomes.3
Recent research has shown that pharmacotherapy that targets one or more of the non-anatomical OSA endotypes can reduce OSA severity and thus may have a potential future role in personalized treatment for OSA.3–9 Key background findings stemmed from animal studies that demonstrated several neurotransmitters play a major role in upper airway stabilization during sleep.10,11 Hypoglossal nuclei express abundant concentrations of H1-receptors in rats12 and guinea pigs.13,14 H2-receptors have been identified in the medulla.15 H3-autoreceptors may also modulate hypoglossal motor-nuclei activity.16 Histamine administration at the hypoglossal motor nucleus significantly increased tonic (ie, expiratory) activity of the largest upper airway dilator muscle, genioglossus, in both non-REM and REM sleep via activating H1-receptors in rats17,18 and cats.19 However, histamine neurons become largely silent at sleep onset during natural sleep.20,21 In humans, knowledge on the role of histaminergic stimulation on upper airway stability is limited. Desipramine, which reduced upper airway collapsibility in healthy controls and OSA patients,22,23 had minimal effects on the AHI overall,22 likely due to a wide, non-specific spectrum of target activity, including antagonism of histaminergic receptors.24 Pitolisant, an H3-autoreceptor inverse agonist with wake promoting properties, increases daytime alertness in people with OSA.25,26 However, the effects of histaminergic mechanisms on upper airway stability and respiratory control in people with OSA have not been investigated.
Betahistine, a drug commonly used in clinical practice for Ménière syndrome, is a mild H1-agonist and a potent H3-autoreceptor antagonist/inverse agonist.27 H3-autoreceptor blockage increases brain levels of histamine.28 Additionally, H3 antagonism can potentiate the activity of other neurotransmitters in the central nervous system, including norepinephrine,29 highly expressed at the hypoglossal motor nuclei, and acetylcholine.30
Recent studies indicate that noradrenergic agents, which, like histamine agonists, also have wake promoting properties, can reduce OSA severity when combined with an antimuscarinic.4,6,31,32 An antimuscarinic would be an ideal candidate to combine with histamine for several reasons: 1) animal data suggest that antimuscarinics directly increase pharyngeal muscle tone during REM sleep;10 2) antimuscarinics also have mild sleep promoting effects4,6,31–33 and this property may be beneficial to counteract, at least in part, the wake promoting properties of other agents (such as betahistine in the current work) when used in combination therapy for OSA; 3) an antimuscarinic may offset H3-mediated cholinergic stimulation effects.30
In this randomized, double-blinded, placebo controlled, crossover study we aimed to investigate the effects of betahistine combined with the antimuscarinic oxybutynin (Beta-Oxy) on OSA severity (primary outcome). This previously untested combination of theoretically synergic drugs may provide insight into new treatment targets for OSA. Secondary outcomes were to investigate the effects of the combination on OSA endotypes, other standard polysomnography parameters and next-day sleepiness and alertness.
Methods
Participants
Thirteen people with a diagnosis of OSA within the past year (apnea/hypopnea index [AHI] >15 events/hr) aged 18 to 75 years were recruited. Participants on CPAP therapy were asked to suspend treatment during the trial and for one week prior to the first study visit. Exclusion criteria included any acute or chronic condition other than controlled hypertension and hypercholesterolemia; hypersensitivity to the study drugs; class 3 obesity; any medication known to influence breathing, sleep/arousal, muscle physiology, or to interact with mono amino oxidases; and current treatment with tricyclic antidepressants.
The study was approved by the Southern Adelaide Clinical Research Ethics Committee (248.20), a joint committee of the Southern Adelaide Local Health Network and Flinders University, was prospectively registered (ACTRN12621000158864) and was performed in accordance with the principles of the Declaration of Helsinki. All participants provided written informed consent prior to enrolment. Participants studied at Adelaide Institute for Sleep Health, Flinders University.
Protocol
Participants were asked to come to the sleep laboratory twice, one week apart, to undertake two overnight sleep studies. Prior to lights-out (based on the participant’s usual bedtime and kept constant between study nights), participants received 96 mg betahistine plus 5mg oxybutynin or placebo according to a double-blind, randomized, crossover design (Figure 1). The study pharmacist, separate to the study site, provided the randomization code and maintained allocation concealment throughout the study. Study medications were placed in identical capsules that could not be identified by study personnel or participants. Prior to sleep, participants were instructed to sleep on their back as much as possible and were given a standardized 8-hour sleep opportunity during each study visit.
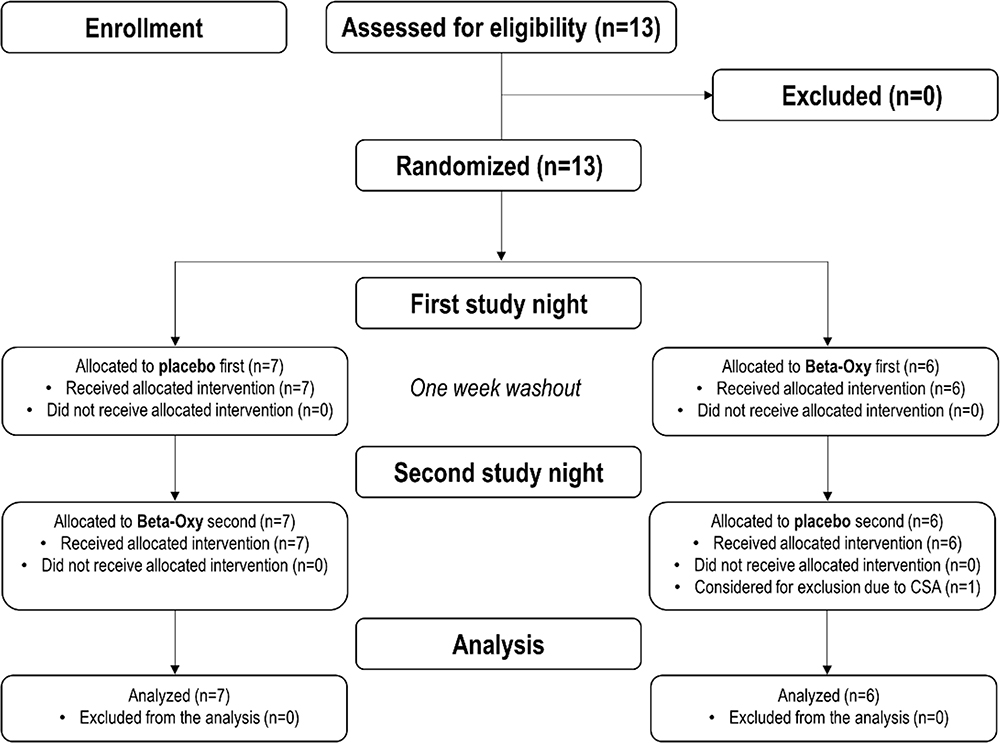 |
Figure 1 CONSORT diagram that shows recruitment, randomization, and analysis procedures for the trial. Notes: Adapted from: Schulz KF, Altman DG, Moher D, CONSORT Group. CONSORT 2010 Statement: Updated Guidelines for Reporting Parallel Group Randomised Trials. PLoS Med. 2010;7(3):e1000251.58 Copyright: © 2010 Schulz et al. Creative Commons Attribution License. |
Participants slept with standard clinical polysomnography equipment including a nasal cannula attached to a pressure transducer to estimate airflow.34
On the first night, ~30 mins after arrival, participants completed the Insomnia Severity Index and the Epworth Sleepiness Scale questionnaires. Systemic blood pressure and the Karolinska Sleepiness Scale (KSS) were recorded during both visits, ~30 min before bed and ~30 min after wake time. In addition, participants completed a 30-minute alertness test using the AusEd driving simulator35 and the Leeds Sleep Evaluation Questionnaire (LSEQ) during the next morning following each study night. Potential side effects (eg, dry mouth, dysuria, etc.) were also investigated in the morning after each sleep study and on arrival back to the laboratory after the one-week washout period.
Data Analysis
All analyses were conducted blinded to the study interventions. Respiratory events and arousals were scored using standard American Academy of Sleep Medicine 2020 criteria.36 OSA endotypes (ie, loop gain, arousal threshold, upper airway collapsibility (Vpassive), markers of pharyngeal muscle compensation (Vactive and Vcomp) and the ventilatory response to arousal) were estimated from the polysomnography-derived flow signal using previously validated algorithms.37–39 In brief, loop gain was calculated as the response to disturbance of different frequencies: one cycle/minute (ie, loop gain1) and the frequency that would lead to periodic breathing onset (ie, loop gain at natural frequency [loop gainn]). Arousal threshold was calculated as the average estimated ventilation during sleep prior to arousals (ie, maximum ventilatory drive). Vpassive was defined as the ventilation during sleep at eupneic ventilatory drive when the pharyngeal muscles are relatively passive. Vactive was defined as the level of ventilation at maximum drive (ie, arousal threshold). Vcomp was taken as the difference between ventilation at maximal drive and as an estimate of pharyngeal muscle compensation. All traits, except for loop gain, which is dimensionless, were expressed as percent of the estimated eupneic ventilation (Veupnea).
Statistical Analysis
An a priori power calculation indicated that 12 participants were required to detect a minimally important change in AHI of 10 events/hour (SD = 10) with >80% power at an alpha level = 0.05 (two-tailed paired t-test), allowing for a 20% drop-out rate.
Continuous data were expressed as mean±SD, or median [interquartile range] for non-normally distributed data. Statistical significance was inferred if p<0.05. According to our statistical analysis plan, data were analyzed using two-tailed paired Student’s t-tests or a Wilcoxon signed-rank test as appropriate. A mixed model analysis was also carried out (random effect: participants; fixed effects: treatment and percent supine sleep) to explore potential effects of sleep positions on the AHI (effect size [confidence interval]). Exploratory linear regression assessed the association between baseline loop gain and change in AHI between the nights. Analyses were performed using Graph Pad Prism 6.0 (Graph Pad Software, La Jolla, CA) and SPSS 23.00 (IBM, Armonk, NY).
Results
Thirteen participants were recruited to allow for a potential drop out to reach our recruitment target of n = 12. However, all 13 participants successfully completed both nights and were included in the analyses (Figure 1). Baseline participant characteristics are shown in Table 1. Participants were recruited from October 2020 to May 2021.
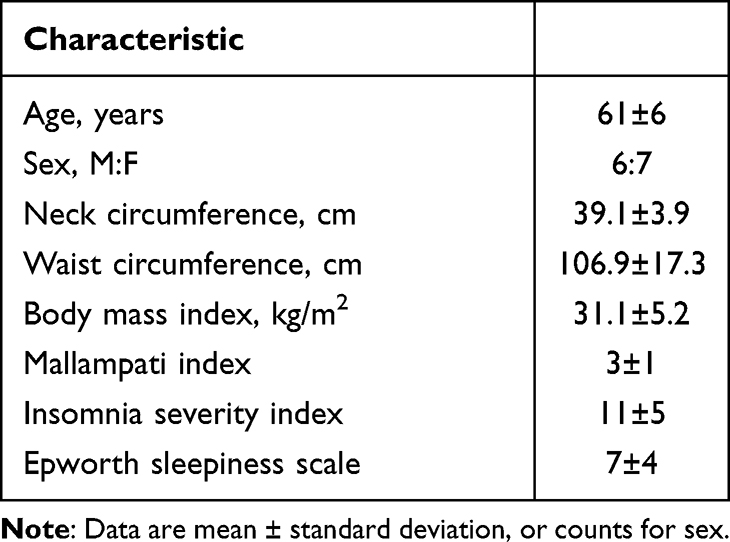 |
Table 1 Baseline Characteristics |
Effect of Beta-Oxy on OSA Severity
Beta-Oxy did not systematically alter OSA severity versus placebo (Figure 2), including when separated according to sleep stage (Table 2). The supine AHI also did not change between study nights (51.0±22.8 on placebo versus 51.0±31.1 events/h on Beta-Oxy, P = 0.37). When adjusting for sleeping body position (Table 2) and missing values in the supine position (in N = 2 nights there was no recorded supine sleep data), there was no effect of the combined drugs on OSA severity (AHI: +6.33 [–0.93, 13.60] events/h, mean [CI], P = 0.08). Overnight desaturation profiles were also not significantly different between the two treatment arms (Table 2).
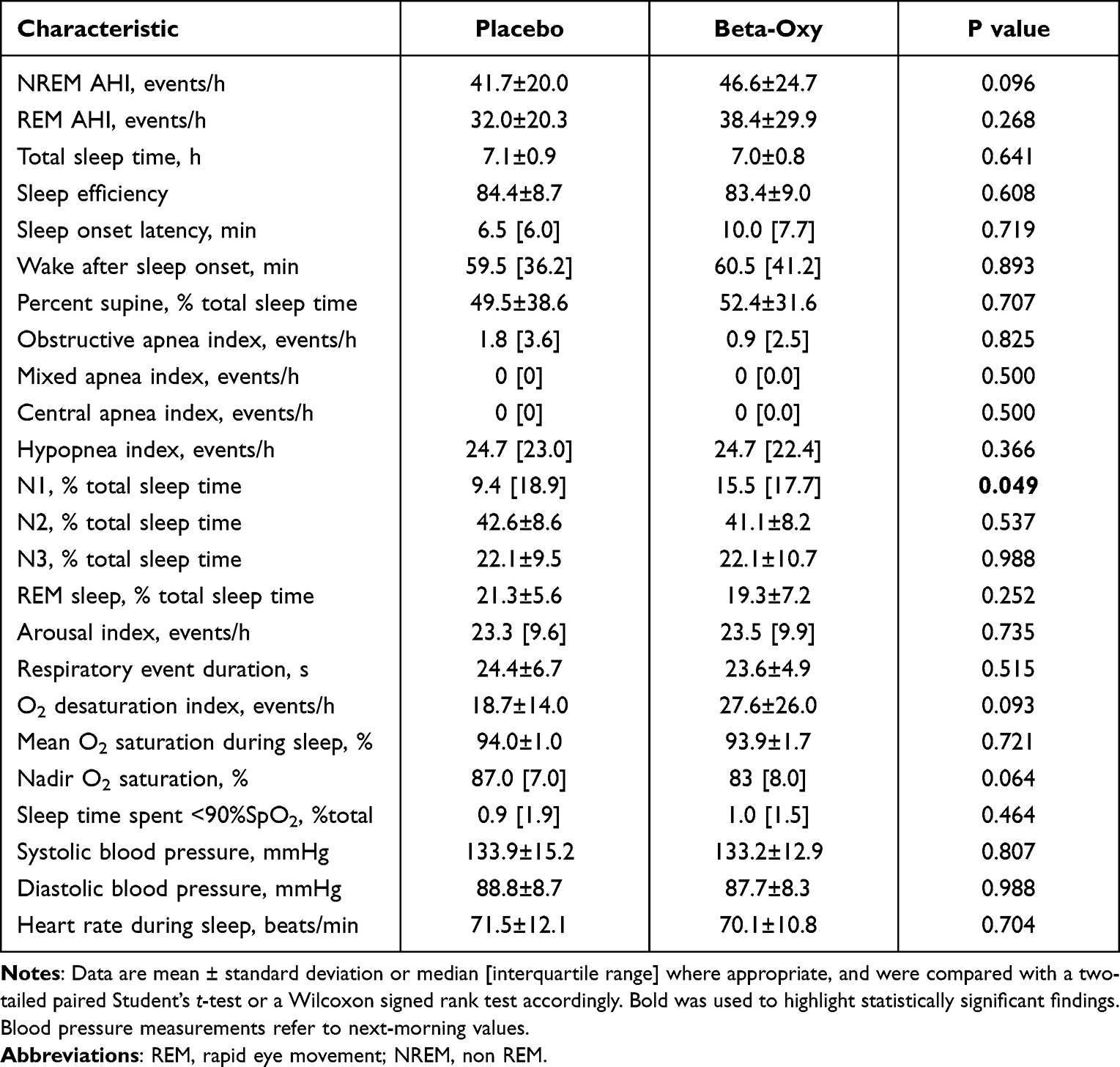 |
Table 2 Polysomnography Parameters |
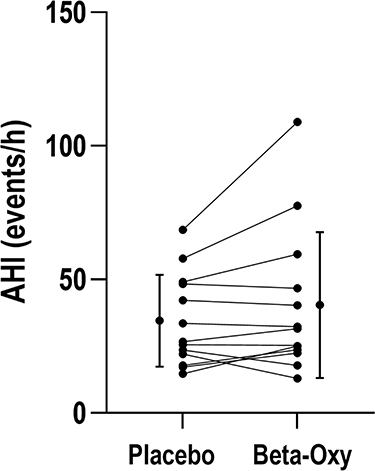 |
Figure 2 There was no systematic difference in apnea-hypopnea index (AHI) between study nights (34.4±17.2 vs 40.3±27.3 events/h sleep, P = 0.124). Bars represent mean ± standard deviation. |
Effect of Beta-Oxy on OSA Endotypes
Beta-Oxy significantly increased loop gain1 compared to placebo (Table 3). Loop gainn was also greater on Beta-Oxy versus placebo in all but one of the study participants (Figure 3). The other OSA endotypes did not change between nights (Table 3). Notably, the change in AHI between the nights was directly associated with loop gain on the placebo night such that OSA severity increased in those with higher loop gain values (Figure 4).
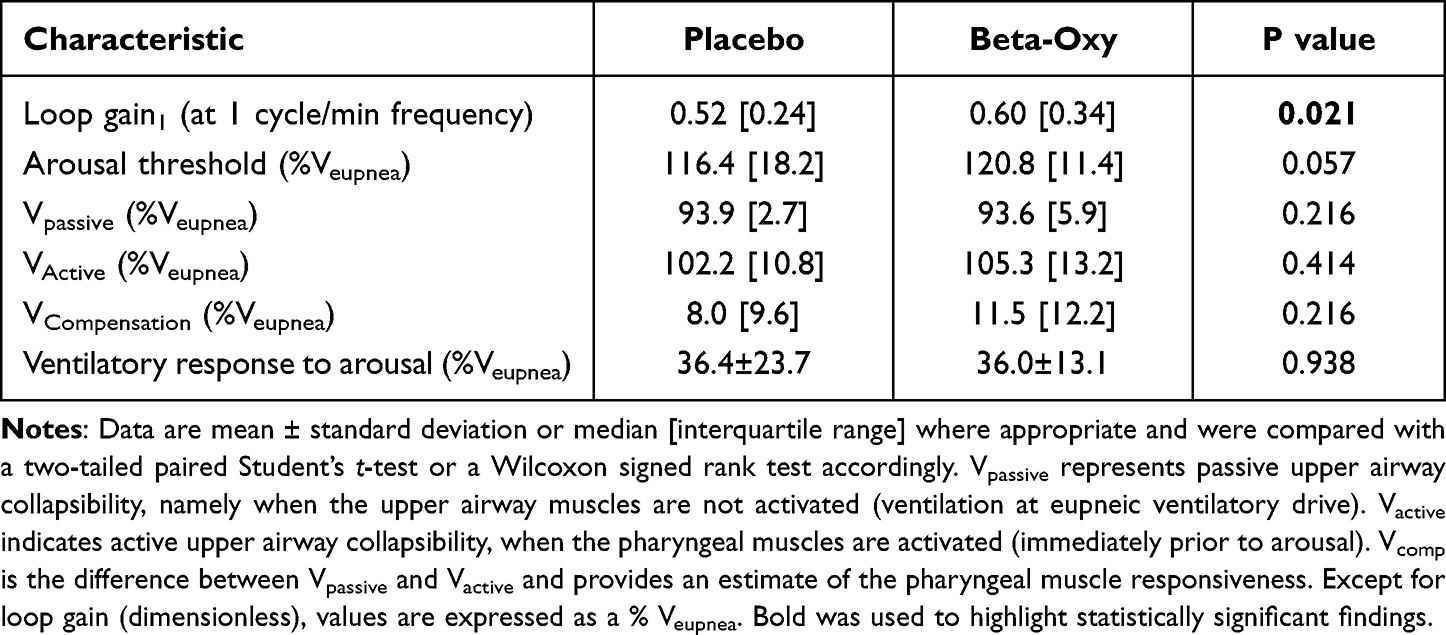 |
Table 3 Polysomnography-Estimated OSA Endotypes |
 |
Figure 3 Beta-oxy significantly increased loop gain at the natural frequency of resonance (loop gainn) by a physiologically meaningful margin (>10%; Δchange = 0.06±0.05 [mean±SD], P = 0.001, asterisk). |
 |
Figure 4 Relationship between change in apnea-hypopnea index (AHI) and loop gain (placebo visit). Change in AHI and loop gain1 (upper panel) and loop gainn (lower panel) is illustrated. Individual participants are indicated by dots and solid line indicates the calculated relationship from linear regression. |
Effect of Beta-Oxy on Other Polysomnography Parameters, Next-Day Sleepiness and Alertness, and Safety
Beta-Oxy changed sleep architecture as reflected via an increase in N1 sleep versus placebo (Table 2). No other significant differences were detected between nights for overall sleep efficiency, total sleep time, wake after sleep onset or arousal index (Table 2).
Beta-Oxy did not change next-day sleepiness or alertness according to KSS, LSEQ scores or to the AusEd driving simulation task versus placebo (Table 4). Overall, the drug combination was well tolerated. Three participants had minor complaints following study visits. One participant reported vertigo and visual aura in the morning following the Beta-Oxy night, another reported a longer than usual menstrual phase on return to the laboratory during the washout week following the placebo night.
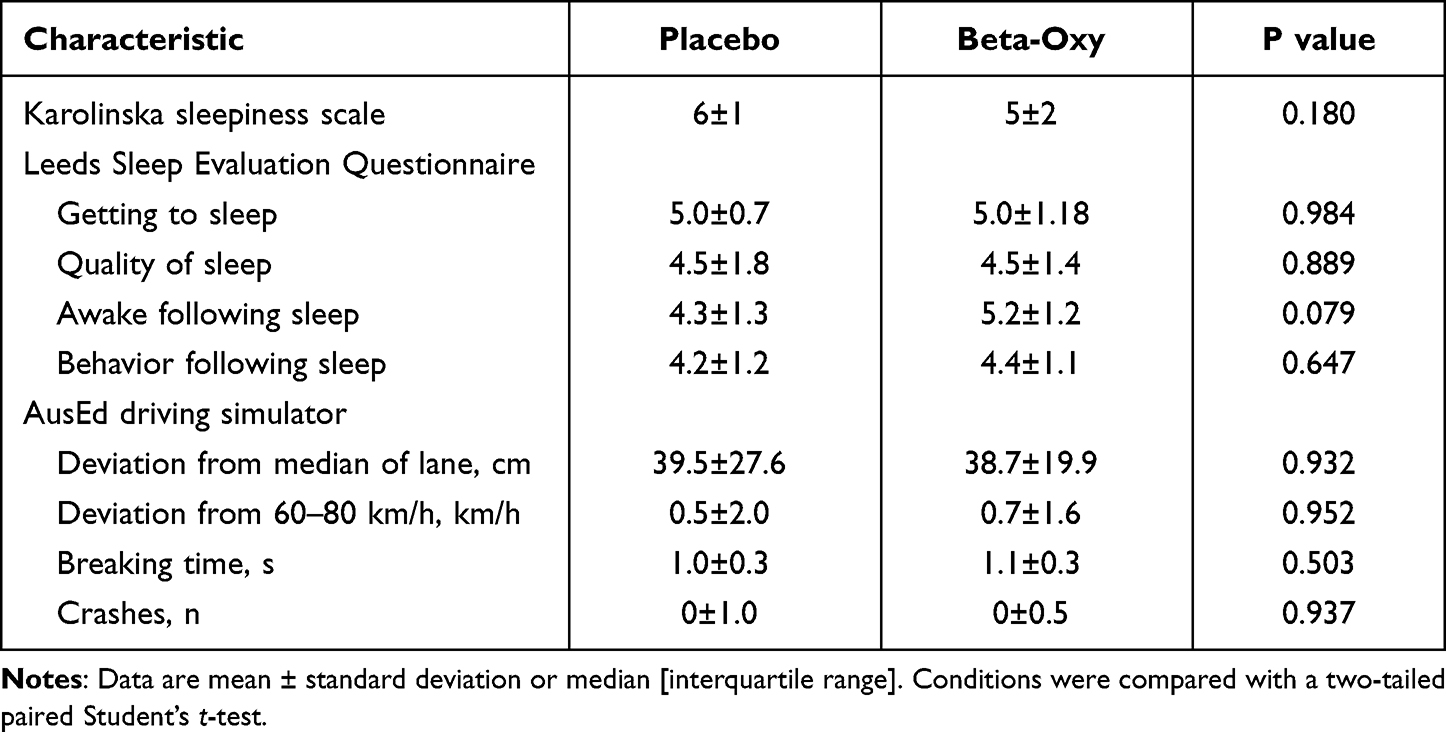 |
Table 4 Next-Day Perceived Sleepiness and Objective Alertness |
Three participants reported feeling rested/wide awake after Beta-Oxy single-night treatment versus one after placebo (P = 0.194). Similarly, only one participant reported feeling very tired after Beta-Oxy versus three participants after the placebo night (P = 0.139). Four participants on Beta-Oxy versus six on placebo felt that the corresponding laboratory night was worse/much worse than sleeping at home (P = 0.619). Perceived sleep latency was comparable between placebo and Beta-Oxy (29±18 min v. 44±60 respectively, P = 0.397).
Discussion
This is the first proof-of-concept, mechanistic study to investigate the effects of a histaminergic agent, together with an anti-muscarinic, on OSA severity in humans. Beta-Oxy led to a physiologically important increase in loop gain, without changing the AHI, other sleep parameters or next-day sleepiness/alertness. These findings provide novel physiological insight and, if confirmed in larger follow-up studies, may have implications for certain respiratory diseases where blunted respiratory control is a feature.
Novel Physiological Insights
The response to a respiratory disturbance during sleep (reduced ventilation) consists of an accumulation of ventilatory drive that generally matches increased hypoxic—and hypercapnic—demand to generate a subsequent ventilatory compensation response.40 The main effectors of this process, a constituent component of loop gain,41 are the chemoreceptors, that project to the nucleus tractus solitarii in the brainstem and are influenced via a wide supply of neurotransmitters, including histamine.42 In rats, histaminergic modulation through H1-receptor stimulation42 or H3-receptor blockade in the brain,43 augments chemoreflex control. A similar effect was observed in goats44 and cats.45 However, its human translatability was only putative. This study shows that loop gain is almost invariably increased with Beta-Oxy in people with OSA, indicating that this potent chemosensitivity excitatory modulation is also present in humans during sleep.
Conversely, it is not clear why Beta-Oxy did not have an effect on estimated pharyngeal muscle compensation despite the strong neurobiological rationale behind our study hypothesis. One explanation could be that the dose of betahistine was not high enough to produce a detectable effect on this endotype. 96 mg was selected in this study as it is twice as high as a typical dose administered clinically, although up to 200 mg was well tolerated in other studies with no significant complications.46 A second explanation is that betahistine did lead to an increase in genioglossus muscle activity (not directly measured in this study) but its potential beneficial effect on OSA was offset by the more pronounced increase in loop gain and thus was not detectable via our indirect measurement technique. Indeed, Vcomp is the most challenging endotype to accurately quantify using polysomnography-based estimates.37 Accordingly, it will be important in future studies to directly measure pharyngeal muscle activity, including different doses, to investigate the effects of betahistine definitively.
Clinical and Physiological Implications
A drug that increases loop gain without altering pharyngeal pathophysiology could have disparate effects in clinical practice. The prokinetic domperidone has been shown to increase chemosensitivity and loop gain in animal models47,48 and early reports in healthy humans.49 However, this effect is presumably only mediated at the peripheral chemoreceptors as domperidone poorly penetrates the blood brain barrier.50 Betahistine could exert its effects on either central or peripheral chemoreceptors, and this may unveil therapeutic implications for conditions in which central chemosensitivity is impaired or depressed, such as obesity hypoventilation syndrome,51 congenital central hypoventilation52 and opioid-induced respiratory depression.53–56 Thus, the use of betahistine in these conditions is worthy of further investigation in light of the current novel findings on respiratory control.
Conversely, although there was no overall increase in AHI in the current study, any agent that increases loop gain in people with OSA where blunted respiratory control is not a feature, especially in the more than one-third of patients who already have high loop gain,2 is likely to be deleterious. Our finding that Beta-Oxy was associated with increased OSA severity in participants with high loop gain on placebo and vice versa provides initial support for this concept. Thus, the use of betahistine in most people with OSA should be cautioned until further endotype-specific studies are performed to separate out the characteristics of those who may experience beneficial versus deleterious effects.
Despite well-known histamine-related arousal facilitation,57 Beta-Oxy did not worsen sleep efficiency or increase the arousal index or the arousal threshold. However, there was a small increase in lighter N1 stage sleep. While these findings may provide support that betahistine (widely used worldwide at any time of the day) is unlikely to disrupt sleep, it may also be that potential sleep disruption effects were alleviated by oxybutynin which can serve as a mild sleep promotion aid.4,6,31–33 This will require further investigation with betahistine studied in isolation rather than in combination with oxybutynin.
Methodological Considerations
This study has several limitations. 1) As highlighted, betahistine and oxybutynin were not tested separately, and we cannot, therefore, confidently discriminate the effects of the single drugs. However, when studied in a single drug trial, oxybutynin alone did not increase loop gain.4 Thus, the effect on loop gain observed in this study is likely to be solely attributable to the increase in histaminergic tone. 2) Due to the clinical setting of the polysomnography studies, we did not record end-tidal CO2 and the different components of loop gain (eg, plant gain, controller gain) were not calculated. Also, tidal volume was not recorded. Yet, based on the neurobiological signaling attributed to H3-receptor blockade from animal data, betahistine effects on loop gain are likely to be predominantly mediated by increased chemosensitivity (controller gain). 3) We also did not assess whether the histaminergic mediated changes took place in the central, peripheral chemoreceptors or both. 4) All endotypes were collected using polysomnography-derived signals that provide estimates of gold-standard measurements. As highlighted, the current findings provide initial physiological insight to guide future more detailed physiological research investigations.
Conclusions
The combination of betahistine, a histaminergic drug, and the antimuscarinic oxybutynin increases loop gain in people with OSA, without major accompanying systematic effects on OSA severity, sleep architecture or next-day alertness and sleepiness.
Data Sharing Statement
Trial protocol and data will be stored for up to 15 years and will be available after de-identification upon request and subject to ethical approval. Data sharing requests should be directed to Danny Eckert: [email protected].
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was funded by a National Health and Medical Research Council (NHMRC) of Australia Investigator Grant (1196261). DJE is also supported by a NHMRC Senior Research Fellowship (1116942).
Disclosure
Alan Chiang reports grants from Cooperative Research Centre Project Grant (Australian Government, Academia and Industry collaboration, Industry partner: Oventus Medical), outside the submitted work. Luigi Taranto-Montemurro is Chief Scientific Officer of Apnimed and has a financial interest in Apnimed, a company developing pharmacologic therapies for sleep apnea. His interests were reviewed and are managed by Brigham and Women’s Hospital and Partners HealthCare in accordance with their conflict of interest policies; and reports personal fees from Apnimed, outside the submitted work. Danny J Eckert reports grants from National Health and Medical Research Council of Australia, during the conduct of the study; research grants and personal fees from Bayer, Apnimed, Takeda, and Invicta Medical, and a Collaborative Research Centre (CRC-P) Consortium Grant between the Australian Government, Academia and Industry (Industry partner: Oventus Medical) and serves on the Scientific Advisory Boards for Invicta Medical and Apnimed, outside the submitted work. The authors report no other potential conflicts of interest in relation to this work.
References
1. Young T, Palta M, Dempsey J, Skatrud J, Weber S, Badr S. The occurrence of sleep-disordered breathing among middle-aged adults. N Engl J Med. 1993;328:1230–1235. doi:10.1056/NEJM199304293281704
2. Eckert DJ, White DP, Jordan AS, Malhotra A, Wellman A. Defining phenotypic causes of obstructive sleep apnea. Identification of novel therapeutic targets. Am J Respir Crit Care Med. 2013;188:996–1004. doi:10.1164/rccm.201303-0448OC
3. Pepin JL, Eastwood P, Eckert DJ. Novel avenues to approach non-CPAP therapy and implement comprehensive OSA care. Eur Respir J. 2021;2101788. doi:10.1183/13993003.01788-2021
4. Taranto-Montemurro L, Messineo L, Sands SA, et al. The combination of atomoxetine and oxybutynin greatly reduces obstructive sleep apnea severity. A randomized, placebo-controlled, double-blind crossover trial. Am J Respir Crit Care Med. 2019;199:1267–1276. doi:10.1164/rccm.201808-1493OC
5. Messineo L, Carter SG, Taranto-Montemurro L, et al. Addition of zolpidem to combination therapy with atomoxetine-oxybutynin increases sleep efficiency and the respiratory arousal threshold in obstructive sleep apnoea: a randomized trial. Respirology. 2021;26:878–886. doi:10.1111/resp.14110
6. Lim R, Messineo L, Grunstein RR, Carberry JC, Eckert DJ. The noradrenergic agent reboxetine plus the antimuscarinic hyoscine butylbromide reduces sleep apnoea severity: a double-blind, placebo-controlled, randomised crossover trial. J Physiol. 2021;599:4183–4195. doi:10.1113/JP281912
7. Taranto-Montemurro T-M, Messineo M, Wellman W. Targeting endotypic traits with medications for the pharmacological treatment of obstructive sleep apnea. a review of the current literature. J Clin Med. 2019;8:1846. doi:10.3390/jcm8111846
8. Messineo L, Eckert DJ, Lim R, et al. Zolpidem increases sleep efficiency and the respiratory arousal threshold without changing sleep apnoea severity and pharyngeal muscle activity. J Physiol. 2020;598:4681–4692. doi:10.1113/JP280173
9. Dempsey JA, Veasey SC, Morgan BJ, O’Donnell CP. Pathophysiology of sleep apnea. Physiol Rev. 2010;90:47–112. doi:10.1152/physrev.00043.2008
10. Grace KP, Hughes SW, Horner RL. Identification of the mechanism mediating genioglossus muscle suppression in REM sleep. Am J Respir Crit Care Med. 2013;187:311–319.
11. Chan E, Steenland HW, Liu H, Horner RL. Endogenous excitatory drive modulating respiratory muscle activity across sleep-wake states. Am J Respir Crit Care Med. 2006;174:1264–1273. doi:10.1164/rccm.200605-597OC
12. Kivipelto L, Majane EA, Yang HY, Panula P. Immunohistochemical distribution and partial characterization of FLFQPQRFamidelike peptides in the central nervous system of rats. J Comp Neurol. 1989;286:269–287. doi:10.1002/cne.902860211
13. Traiffort E, Leurs R, Arrang JM, et al. Guinea pig histamine H1 receptor. I. Gene cloning, characterization, and tissue expression revealed by in situ hybridization. J Neurochem. 1994;62:507–518. doi:10.1046/j.1471-4159.1994.62020507.x
14. Bouthenet ML, Ruat M, Sales N, Garbarg M, Schwartz JC. A detailed mapping of histamine H1-receptors in Guinea-pig central nervous system established by autoradiography with [125I]iodobolpyramine. Neuroscience. 1988;26:553–600. doi:10.1016/0306-4522(88)90167-4
15. Traiffort E, Pollard H, Moreau J, et al. Pharmacological characterization and autoradiographic localization of histamine H2 receptors in human brain identified with [125I]iodoaminopotentidine. J Neurochem. 1992;59:290–299. doi:10.1111/j.1471-4159.1992.tb08903.x
16. Xie L, Wu Q, Hu W, et al. Impact of histaminergic H3 receptor antagonist on hypoglossal nucleus in chronic intermittent hypoxia conditions. Psychopharmacology. 2021;238:121–131. doi:10.1007/s00213-020-05663-0
17. Bastedo T, Chan E, Park E, Liu H, Horner RL. Modulation of genioglossus muscle activity across sleep-wake states by histamine at the hypoglossal motor pool. Sleep. 2009;32:1313–1324. doi:10.1093/sleep/32.10.1313
18. Liu ZL, Wu X, Luo YJ, et al. Signaling mechanism underlying the histamine-modulated action of hypoglossal motoneurons. J Neurochem. 2016;137:277–286. doi:10.1111/jnc.13548
19. Neuzeret PC, Sakai K, Gormand F, et al. Application of histamine or serotonin to the hypoglossal nucleus increases genioglossus muscle activity across the wake-sleep cycle. J Sleep Res. 2009;18:113–121. doi:10.1111/j.1365-2869.2008.00708.x
20. Vanni-Mercier G, Gigout S, Debilly G, Lin JS. Waking selective neurons in the posterior hypothalamus and their response to histamine H3-receptor ligands: an electrophysiological study in freely moving cats. Behav Brain Res. 2003;144:227–241. doi:10.1016/S0166-4328(0300091-3
21. Haas HL, Sergeeva OA, Selbach O. Histamine in the nervous system. Physiol Rev. 2008;88:1183–1241.
22. Taranto-Montemurro L, Sands SA, Edwards BA, et al. Desipramine improves upper airway collapsibility and reduces OSA severity in patients with minimal muscle compensation. Eur Respir J. 2016;48:1340–1350. doi:10.1183/13993003.00823-2016
23. Taranto-Montemurro L, Edwards BA, Sands SA, et al. Desipramine increases genioglossus activity and reduces upper airway collapsibility during non-REM sleep in healthy subjects. Am J Respir Crit Care Med. 2016;194:878–885. doi:10.1164/rccm.201511-2172OC
24. Kang JA, Lee K, Lee KM, et al. Desipramine inhibits histamine H1 receptor-induced Ca2+ signaling in rat hypothalamic cells. PLoS One. 2012;7:e36185. doi:10.1371/journal.pone.0036185
25. Dauvilliers Y, Verbraecken J, Partinen M, et al. Pitolisant for daytime sleepiness in patients with obstructive sleep apnea who refuse continuous positive airway pressure treatment. a randomized trial. Am J Respir Crit Care Med. 2020;201:1135–1145. doi:10.1164/rccm.201907-1284OC
26. Pepin JL, Georgiev O, Tiholov R, et al. Pitolisant for residual excessive daytime sleepiness in OSA patients adhering to CPAP: a randomized trial. Chest. 2021;159:1598–1609. doi:10.1016/j.chest.2020.09.281
27. Lacour M, Sterkers O. Histamine and betahistine in the treatment of vertigo: elucidation of mechanisms of action. CNS Drugs. 2001;15:853–870. doi:10.2165/00023210-200115110-00004
28. Arrang JM, Garbarg M, Quach TT. Actions of betahistine at histamine receptors in the brain. Eur J Pharmacol. 1985;111:73–84. doi:10.1016/0014-2999(85)90115-3
29. Schlicker E, Fink K, Hinterthaner M, Gothert M. Inhibition of noradrenaline release in the rat brain cortex via presynaptic H3 receptors. Naunyn Schmiedebergs Arch Pharmacol. 1989;340:633–638. doi:10.1007/BF00717738
30. Clapham J, Kilpatrick GJ. Histamine H3 receptors modulate the release of [3H]-acetylcholine from slices of rat entorhinal cortex: evidence for the possible existence of H3 receptor subtypes. Br J Pharmacol. 1992;107:919–923. doi:10.1111/j.1476-5381.1992.tb13386.x
31. Perger E, Taranto Montemurro L, Rosa D, et al. Reboxetine plus oxybutynin for OSA treatment: a 1-week, randomized, placebo-controlled, double-blind crossover trial. Chest. 2021;161:237–247. doi:10.1016/j.chest.2021.08.080
32. Taranto-Montemurro L, Messineo L, Azarbarzin A, et al. Effects of the combination of atomoxetine and oxybutynin on OSA endotypic traits. Chest. 2020;157:1626–1636. doi:10.1016/j.chest.2020.01.012
33. Lim R, Carberry JC, Wellman A, Grunstein R, Eckert DJ. Reboxetine and hyoscine butylbromide improve upper airway function during nonrapid eye movement and suppress rapid eye movement sleep in healthy individuals. Sleep. 2019;42:zsy261. doi:10.1093/sleep/zsy261
34. The Report of an American Academy of Sleep Medicine Task Force. Sleep-related breathing disorders in adults: recommendations for syndrome definition and measurement techniques in clinical research. Sleep. 1999;22:667–689. doi:10.1093/sleep/22.5.667
35. Vakulin A, Baulk SD, Catcheside PG, et al. Effects of alcohol and sleep restriction on simulated driving performance in untreated patients with obstructive sleep apnea. Ann Intern Med. 2009;151:447–455. doi:10.7326/0003-4819-151-7-200910060-00005
36. Berry RB, Brooks R, Gamaldo C, et al. AASM scoring manual updates for 2017 (Version 2.4). J Clin Sleep Med. 2017;13:665–666. doi:10.5664/jcsm.6576
37. Sands SA, Edwards BA, Terrill PI, et al. Phenotyping pharyngeal pathophysiology using polysomnography in patients with obstructive sleep apnea. Am J Respir Crit Care Med. 2018;197:1187–1197. doi:10.1164/rccm.201707-1435OC
38. Terrill PI, Edwards BA, Nemati S, et al. Quantifying the ventilatory control contribution to sleep apnoea using polysomnography. Eur Respir J. 2015;45:408–418. doi:10.1183/09031936.00062914
39. Sands SA, Terrill PI, Edwards BA, et al. Quantifying the arousal threshold using polysomnography in obstructive sleep apnea. Sleep. 2018;41:zsx183.
40. Feldman JL, Mitchell GS, Nattie EE. Breathing: rhythmicity, plasticity, chemosensitivity. Annu Rev Neurosci. 2003;26:239–266. doi:10.1146/annurev.neuro.26.041002.131103
41. Messineo L, Taranto-Montemurro L, Azarbarzin A, et al. Breath-holding as a means to estimate the loop gain contribution to obstructive sleep apnoea. J Physiol. 2018;596:4043–4056. doi:10.1113/JP276206
42. Dutschmann M, Bischoff AM, Busselberg D, Richter DW. Histaminergic modulation of the intact respiratory network of adult mice. Pflugers Arch. 2003;445:570–576. doi:10.1007/s00424-002-0904-z
43. Lazarov N, Rozloznik M, Reindl S, Rey-Ares V, Dutschmann M, Gratzl M. Expression of histamine receptors and effect of histamine in the rat carotid body chemoafferent pathway. Eur J Neurosci. 2006;24:3431–3444. doi:10.1111/j.1460-9568.2006.05241.x
44. Haxhiu MA, Cherniack NS, Altose MD, Kelsen SG. Effect of histamine on respiratory chemosensitivity in conscious goats. Respiration. 1983;44:411–418. doi:10.1159/000194578
45. Del Rio R, Moya EA, Koenig CS, Fujiwara K, Alcayaga J, Iturriaga R. Modulatory effects of histamine on cat carotid body chemoreception. Respir Physiol Neurobiol. 2008;164:401–410. doi:10.1016/j.resp.2008.09.005
46. Moorthy G, Sallee F, Gabbita P, Zemlan F, Sallans L, Desai PB. Safety, tolerability and pharmacokinetics of 2-pyridylacetic acid, a major metabolite of betahistine, in a Phase 1 dose escalation study in subjects with ADHD. Biopharm Drug Dispos. 2015;36:429–439. doi:10.1002/bdd.1955
47. Nemati S, Edwards BA, Sands SA, et al. Model-based characterization of ventilatory stability using spontaneous breathing. J Appl Physiol. 2011;111:55–67. doi:10.1152/japplphysiol.01358.2010
48. Edwards BA, Sands SA, Skuza EM, et al. Increased peripheral chemosensitivity via dopaminergic manipulation promotes respiratory instability in lambs. Respir Physiol Neurobiol. 2008;164:419–428. doi:10.1016/j.resp.2008.09.003
49. Delpierre S, Fornaris M, Guillot C, Grimaud C. Increased ventilatory chemosensitivity induced by domperidone, a dopamine antagonist, in healthy humans. Bull Eur Physiopathol Respir. 1987;23:31–35.
50. Schinkel AH, Wagenaar E, Mol CA, van Deemter L. P-glycoprotein in the blood-brain barrier of mice influences the brain penetration and pharmacological activity of many drugs. J Clin Invest. 1996;97:2517–2524. doi:10.1172/JCI118699
51. Sampson MG, Grassino K. Neuromechanical properties in obese patients during carbon dioxide rebreathing. Am J Med. 1983;75:81–90. doi:10.1016/0002-9343(83)91171-3
52. Carroll MS, Patwari PP, Kenny AS, Brogadir CD, Stewart TM, Weese-Mayer DE. Residual chemosensitivity to ventilatory challenges in genotyped congenital central hypoventilation syndrome. J Appl Physiol. 2014;116:439–450. doi:10.1152/japplphysiol.01310.2013
53. Weil JV, McCullough RE, Kline JS, Sodal IE. Diminished ventilatory response to hypoxia and hypercapnia after morphine in normal man. N Engl J Med. 1975;292:1103–1106. doi:10.1056/NEJM197505222922106
54. Robinson RW, Zwillich CW, Bixler EO, Cadieux RJ, Kales A, White DP. Effects of oral narcotics on sleep-disordered breathing in healthy adults. Chest. 1987;91:197–203. doi:10.1378/chest.91.2.197
55. Altree TJ, Chung F, Chan MTV, Eckert DJ. Vulnerability to postoperative complications in obstructive sleep apnea: importance of phenotypes. Anesth Analg. 2021;132:1328–1337. doi:10.1213/ANE.0000000000005390
56. Martins RT, Carberry JC, Wang D, Rowsell L, Grunstein RR, Eckert DJ. Morphine alters respiratory control but not other key obstructive sleep apnoea phenotypes: a randomised trial. Eur Respir J. 2020;55:45.
57. Lacour M, Chabbert C. Betahistine treatment in managing vertigo and improving vestibular compensation: clarification. J Vestib Res. 2013;23:139–151. doi:10.3233/VES-130496
58. Schulz KF, Altman DG, Moher D, CONSORT Group. CONSORT 2010 Statement: Updated Guidelines for Reporting Parallel Group Randomised Trials. PLoS Med. 2010;7(3):e1000251.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.