Back to Journals » OncoTargets and Therapy » Volume 11
The association between HOTAIR polymorphisms and cancer susceptibility: an updated systemic review and meta-analysis
Authors Min L, Mu X, Tong A, Qian Y, Ling C, Yi T, Zhao X
Received 12 September 2017
Accepted for publication 3 December 2017
Published 14 February 2018 Volume 2018:11 Pages 791—800
DOI https://doi.org/10.2147/OTT.S151454
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Geoffrey Pietersz
Ling Min, Xiyan Mu, An Tong, Yanping Qian, Chen Ling, Tao Yi, Xia Zhao
Department of Gynecology and Obstetrics, Key laboratory of Obstetrics and Gynecologic and Pediatric Diseases and Birth Defects of Ministry of Education, West China Second Hospital, Sichuan University, Chengdu, People’s Republic of China
Objectives: This work aims to explore whether HOX transcript antisense intergenic RNA (HOTAIR) polymorphisms are associated with cancer susceptibility.
Materials and methods: A comprehensive search was conducted for literature published from January 2007 to July 2017. The pooled odds ratios (ORs) and the corresponding 95% CIs were calculated using the Revman 5.2 software. Eighteen articles of 36 case–control studies were enrolled including six HOTAIR polymorphisms and 10 cancer types.
Results: The results showed that cancer risk was elevated in recessive mutation of rs12826786 (TT vs CC+CT: OR =1.55, 95% CI =1.19, 2.03; TT+CT vs CC: OR =1.23, 95% CI =1.04, 1.46; TT vs CC: OR =1.67, 95% CI =1.24, 2.24; T vs C: OR =1.24, 95% CI =1.09, 1.40) and rs920778 (TT vs CC+CT: OR =1.73, 95% CI =1.30, 2.30; TT+CT vs CC: OR =1.40, 95% CI =1.16, 1.70; TT vs CC: OR =1.83, 95% CI =1.25, 2.68; T vs C: OR =1.37, 95% CI =1.18, 1.59), while the results for polymorphisms of rs7958904, rs4759314, rs874945, and rs1899663 were insignificant. The stratified results for Chinese population were consistent with the overall group analysis.
Conclusion: Our meta-analysis showed that HOTAIR polymorphisms of rs12826786 and rs920778 were correlated with increased cancer risk, while rs7958904, rs4759314, rs874945, and rs1899663 were not. More studies with different types of cancer are needed to confirm the findings.
Keywords: HOTAIR, polymorphism, cancer, susceptibility, meta-analysis
Introduction
The recent sequencing technologies and genome-wide analysis have indicated that only 2% of the genome is in protein-encoded regions and that the majority of the genome is the so-called dark matter that is transcribed into noncoding RNAs (ncRNAs).1–3 These ncRNAs are classified as short and long ncRNAs depending on the nucleotide size. Long ncRNAs (lncRNAs) are commonly defined as non-protein-coding transcripts longer than 200 nucleotides.4,5 They are crucial players in a wide range of biologic processes on the epigenetic, transcriptional, or posttranscriptional level, and are the important regulators of pathophysiological activities such as cell growth, invasion, apoptosis, and metastasis.6–8 So far, more than 3000 lncRNAs have been found, among which the HOX transcript antisense intergenic RNA (HOTAIR) is the mostly widely studied. HOTAIR is a 2.2 kb lncRNA that is transcribed in antisense orientation from the homebox C (HOXC) gene on chromosome 12q13.13. HOTAIR 5′-domain recruits the Polycomb Repressive Complex 2, leading to histone H3 lysine 27 trimethylation (H3K27me3) in the HOXD locus, and HOTAIR 3′-domain interacts with LSD1/REST/CoREST complex, to regulate the metastasis suppressor genes silence.9,10 Clinical and biochemical studies have indicated that deregulation of HOTAIR is a powerful indicator of poor prognosis and malignant progression for several cancers such as ovarian cancer, gastric cancer, and lung cancer.11–13
Genetic variants, mainly composed of single-nucleotide polymorphisms (SNPs), have long been confirmed in various loci of the genome. These variants may exert various influences on the expression or function of a particular gene.14,15 Even with the potential importance of HOTAIR in carcinogenesis, only a few studies have investigated the effects of HOTAIR SNPs on cancer susceptibility. For example, Guo et al reported that the mutated T allele of rs12826786 in HOTAIR could increase the risk of developing gastric cancer and was associated with TNM stage. In addition, higher expression levels of HOTAIR were found in tumor tissues, and rs12826786 SNP had a genotype-specific effect on HOTAIR expression.16 However, in another case–control study conducted by Ulger et al, HOTAIR rs12826786 (C/T) polymorphism was not playing any major role in genetic susceptibility to gastric carcinogenesis in Turkish population.17 As for HOTAIR rs7958904 (G/C), Jin et al found that the rs7958904 CC genotype was related to an increased risk of cervical cancer compared with the GG/GC genotypes. Their MTT assay demonstrated a growth-promoting role of rs7958904 C allele on cervical cancer cells.18 On the contrary, Xue et al revealed that individuals with rs7958904 CC genotype had a significantly decreased risk of colorectal cancer in both stages 1 and 2, compared with those carrying GG genotype.19
To address the inconsistency among different case–control studies, some meta-analyses have been performed to draw a conclusion between HOTAIR polymorphisms and cancer susceptibility. Chu et al pooled eight articles on three HOTAIR polymorphisms and concluded that HOTAIR rs920778 increased the cancer risk in the recessive model.20 Meanwhile, Lv et al summarized five HOTAIR polymorphisms from 16 studies, showing that the rs920778 (C/T) polymorphism was associated with increased risk of overall cancer in the recessive model, while the rs7958904 (G/C) polymorphism was associated with decreased overall risk of cancer in all genetic models.21 Since then, several new case–control studies have been published, some of which reported controversial results compared with previous publications. Moreover, more types of polymorphisms have been explored, providing a perspective to a further systemic review. In this study, we comprehensively collected and assessed all the available articles using meta-analysis with the aim to better clarify the association between currently reported HOTAIR polymorphisms and cancer susceptibility.
Materials and methods
Search for eligible literature
A comprehensive electronic search was performed using PubMed, Embase, Medline (Ovid), Weipu, Wanfang, and CNKI for studies published from January 2007 to July 2017. The following keywords were variably combined: “cancer”, “malignancy”, “HOTAIR”, “lncRNA”, “polymorphism”, “variant”, and “mutation”. The search was updated every week until July 15, 2017.
Inclusion and exclusion criteria
Articles fulfilling the following criteria were included: 1) analyzed HOTAIR polymorphisms in cancer; 2) provided sufficient data in both case and control groups to calculate the odds ratios (ORs) and the corresponding 95% CIs; 3) studied the polymorphisms that appeared in at least two publications; and 4) case–control studies. When duplicate data were present in different articles, only the latest one would be included. Meanwhile, articles that did not fulfill the criteria mentioned above were excluded.
Data extraction
Two investigators independently reviewed all potential studies. The following items were extracted: first author, year of publication, ethnicity, SNPs, cancer type, source of control, genotyping method, adjusted risk factors, and genotype distributions in cases and controls. Any discrepancies were resolved by discussion with a third investigator until a consensus was reached.
The Newcastle–Ottawa Scale (NOS) was used to investigate the quality of included studies. Three aspects of selection, comparability, and exposure (nine scores in total) were carefully evaluated. Studies with scores higher than 5 were included (http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp).49
Statistical analysis
Pooled ORs and corresponding 95% CIs were calculated to estimate the strength of the association between different HOTAIR SNPs and cancer risk. All SNPs were considered as binary variables, and five comparative models were used as follows: recessive genotype vs heterozygous genotype + dominant wild type, recessive genotype + heterozygous genotype vs dominant wild type, heterozygous genotype vs dominant wild type + recessive genotype, recessive genotype vs dominant wild type, and mutant allele vs wild-type allele. The Z-test was conducted to determine the significance of the pooled ORs where P<0.05 was interpreted as statistically significant. Higgins I2 test was used to assess heterogeneity among studies. When I2 was <50%, a fixed-effects model was used, indicating the lack of heterogeneity; otherwise, a random-effects model was applied. The presence of publication bias was evaluated by the inspection of funnel plots. When the funnel plots showed visible asymmetry, Egger’s test was performed to further measure the bias, which was considered as existing when P<0.05. All analyses were undertaken using Revman 5.2 software (Cochrane Collaboration, Copenhagen, Denmark) with the exception of the Egger’s test, which was performed using STATA 14.0 (StataCorp LP, College Station, TX, USA).
Results
Search results
The initial search yielded 337 publications, 262 of which were excluded for being irrelevant to HOTAIR polymorphisms, by reading titles and abstracts. On further evaluation, 37 articles were either biochemical studies or reviews and were therefore ruled out; 18 articles focused on non-cancer diseases such as rheumatoid arthritis and hearing loss; 1 article explored the relationship between cervical cancer risk and HOTAIR rs2366152 polymorphisms, which were not repeated in other published studies, resulting in the impossibility of data pooling;22 and 1 article focused on the HOTAIR rs7958904 polymorphisms in lung cancer, but failed to offer detailed genotype information data.23 Therefore, we enrolled 18 articles of 36 studies in this meta-analysis (Figure 1).16–19,24–37
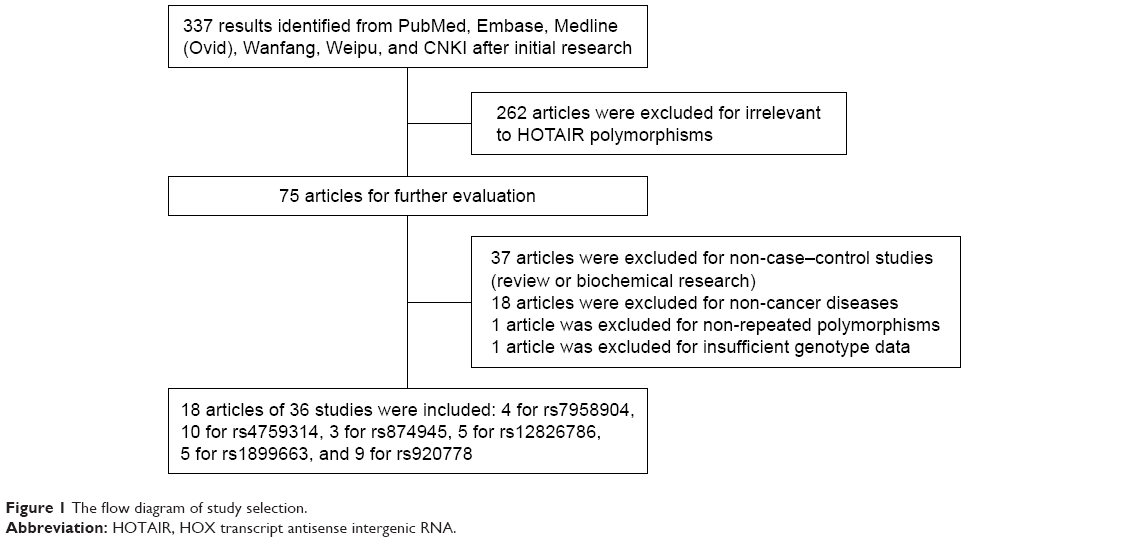 | Figure 1 The flow diagram of study selection. |
Study characteristics
Among the 36 enrolled case–control studies, six HOTAIR polymorphisms were analyzed (rs7958904, rs4759314, rs874945, rs12826786, rs1899663, and rs920778), while 10 cancer types were reported (breast cancer, cervical cancer, colorectal cancer, esophageal cancer, gastric cancer, glioma, lung cancer, ovarian cancer, prostate cancer, and thyroid cancer). Twelve articles were about Chinese population, four were about Turkish, one was about Iranian, and one was about Portuguese. The source of control was also retrieved. Despite the fact that there were 4 articles that failed to mention the detailed control source, 11 articles were hospital based and 3 were population based. The NOS showed that 14 articles were of moderate quality (NOS score of 6 or 7) and 4 were of high quality (NOS score of 8 or 9). All studies reported the numbers of corresponding genotypes as to recessive mutants, heterogeneous mutants, and dominant wild types for both case and control groups. Adjusted variables that might affect the ORs were also summarized for each publication (Table 1).
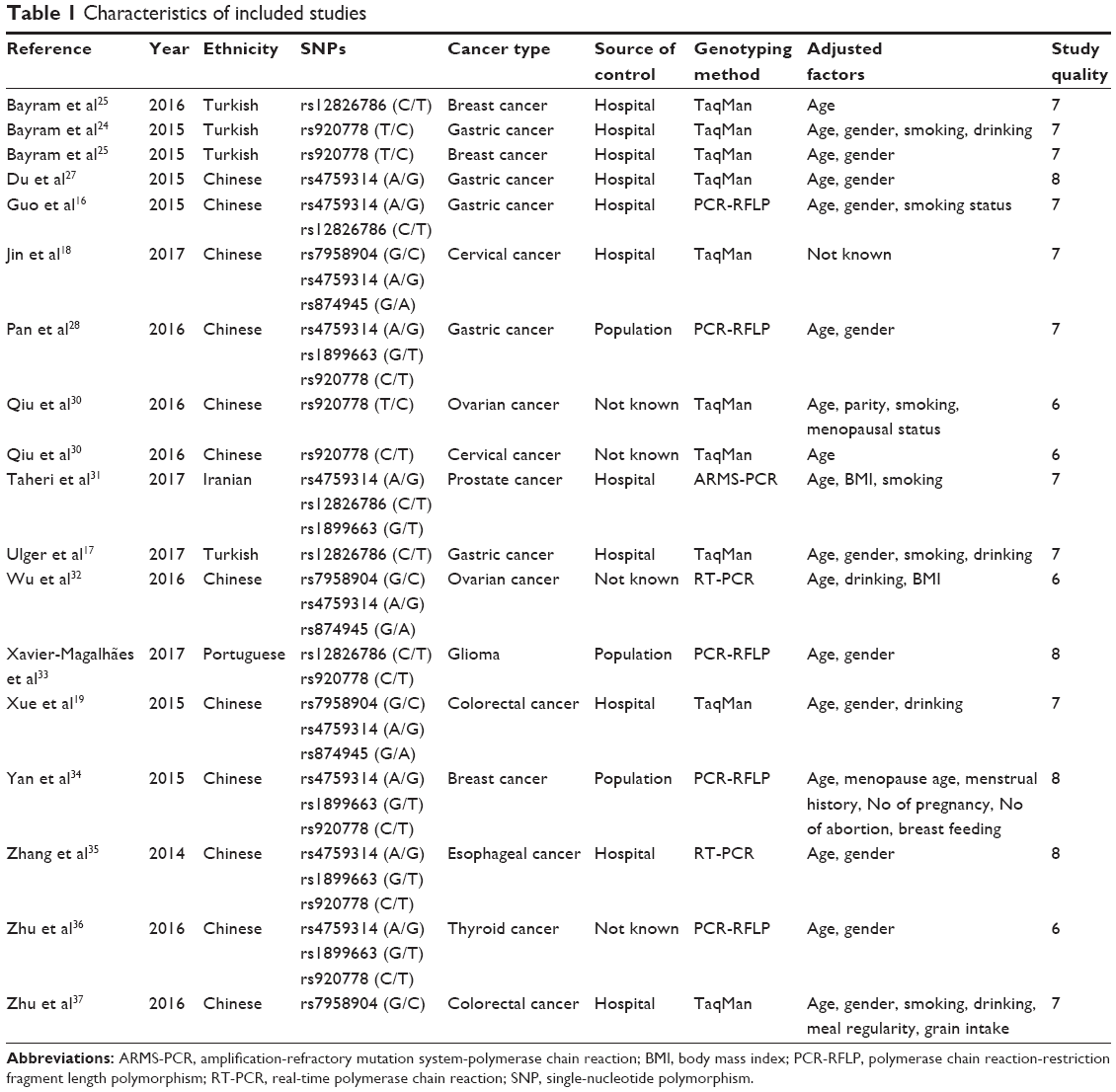 | Table 1 Characteristics of included studies |
Quantitative data analysis
As shown in Tables 2 and 3, six HOTAIR polymorphisms were analyzed in this meta-analysis. For rs12826786 (C/T), five studies including 1,048 cases and 1,432 controls were evaluated. The fixed-effects models proposed a significant association between C-to-T mutation and cancer risk (TT vs CC+CT: OR =1.55, 95% CI =1.19, 2.03; TT+CT vs CC: OR =1.23, 95% CI =1.04, 1.46; TT vs CC: OR =1.67, 95% CI =1.24, 2.24; T vs C: OR =1.24, 95% CI =1.09, 1.40), while heterozygous mutants alone failed to display statistically significant OR (Figure 2A and B). The results for rs920778 (C/T) were similar. Nine studies with 11,442 participants and seven cancer types were pooled. Recessive mutants presented significantly higher cancer risk when compared with either remaining genotypes or homozygous wild types (TT vs CC+CT: OR =1.73, 95% CI =1.30, 2.30; TT vs CC: OR =1.83, 95% CI =1.25, 2.68). The mutant containing genotypes and mutant allele T also showed statistical significance in elevated cancer risk (TT+CT vs CC: OR =1.40, 95% CI =1.16, 1.70; T vs C: OR =1.37, 95% CI =1.18, 1.59), as shown in Figure 2C and D. The stratified analysis of 10,508 Chinese was consistent with the overall group results. Thus, it can be concluded that rs12826786 (C/T) and rs920778 (C/T) were correlated with increased cancer risk.
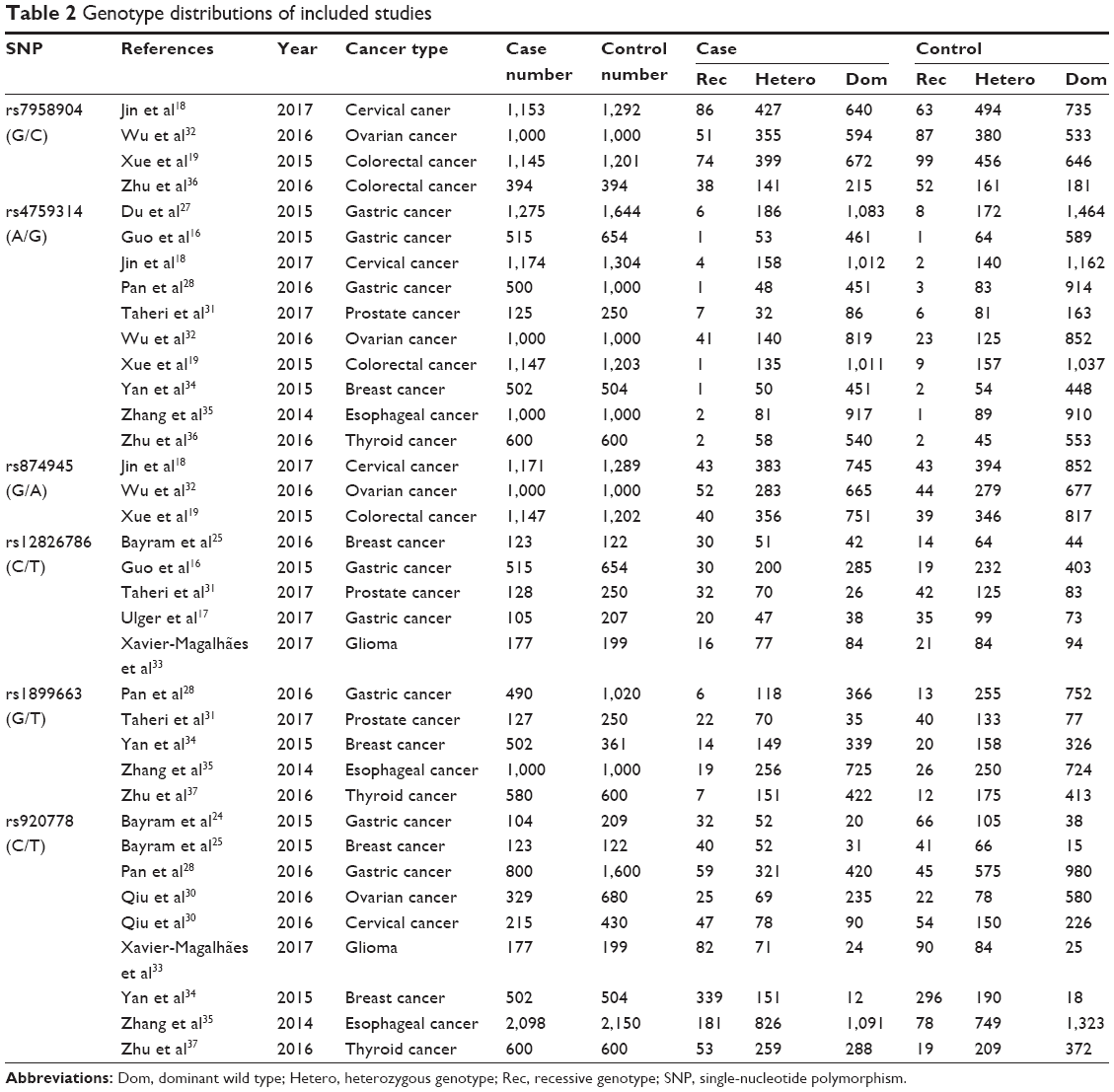 | Table 2 Genotype distributions of included studies |
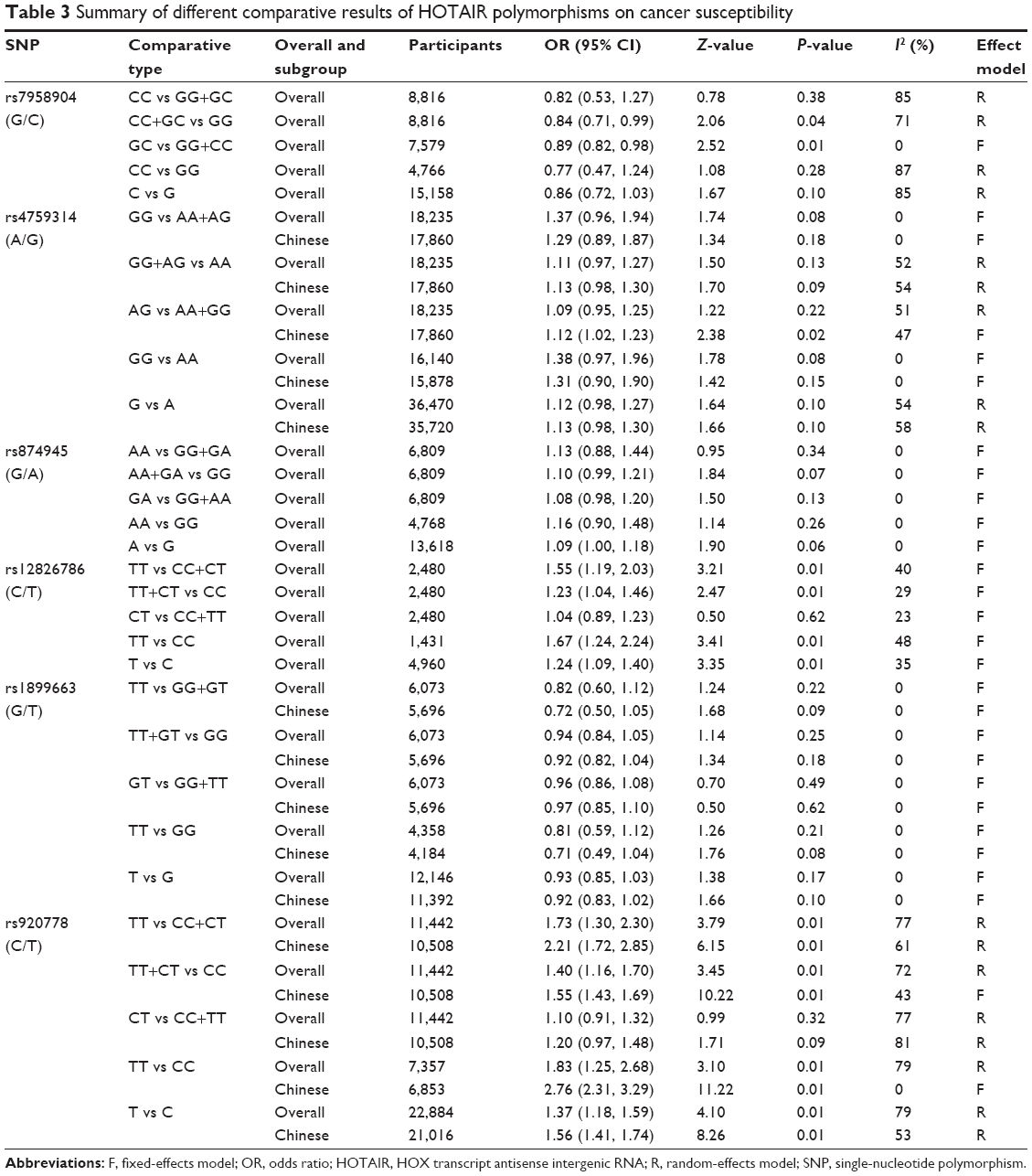 | Table 3 Summary of different comparative results of HOTAIR polymorphisms on cancer susceptibility |
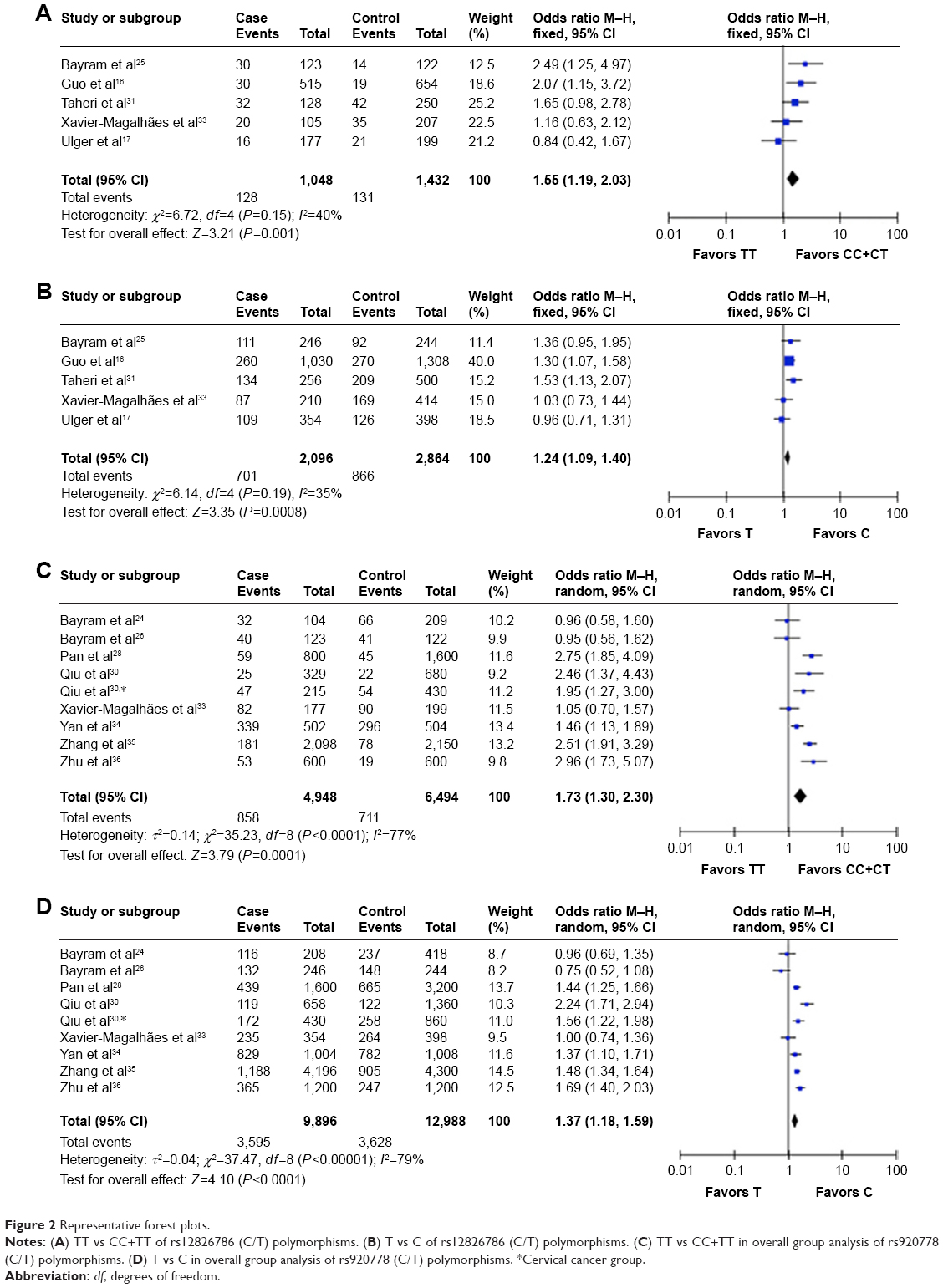 | Figure 2 Representative forest plots. |
The results for rs7958904 (G/C) polymorphisms were less direct. Four studies including 8,816 Chinese participants were analyzed. The meta-analysis showed that the heterozygous mutants alone and combined with recessive mutants posed lower cancer risks (GC vs GG+CC: OR =0.89, 95% CI =0.82–0.98; CC+GC vs GG: OR =0.84, 95% CI =0.71–0.99). Notably, three out of four studies reported that G-to-C mutation could decrease colorectal and ovarian cancer risks, while one study pointed that CC genotype was related to increased cervical cancer risk. Therefore, it is hard to conclude that rs7958904 (G/C) polymorphisms are related to overall cancer susceptibility. However, this inconsistency indicated that rs7958904 (G/C) polymorphisms might play different roles in different types of cancer.
As to the remaining three types of polymorphisms (rs4759314, rs874945, and rs1899663), no significant association was found between mutant genotypes (or alleles) and cancer susceptibility in the corresponding effect models, either for overall population or for Chinese subgroups. Even though allele A implied a cancer-prone tendency (A vs G: OR =1.09; 95% CI =1.00–1.18) in rs874945 (G/A) polymorphisms, it is impossible to draw any significant conclusion.
Publication bias
The publication bias was first assessed by visually examining the funnel plots (Figure 3). Studies on rs4759314, rs874945, rs12826786, and rs1899663 were symmetric, while the existence of bias was indicated in rs7958904 and rs920778. Egger’s test was then performed in the two polymorphisms. The results demonstrated no significant bias in the two polymorphisms (P>0.05, data not shown).
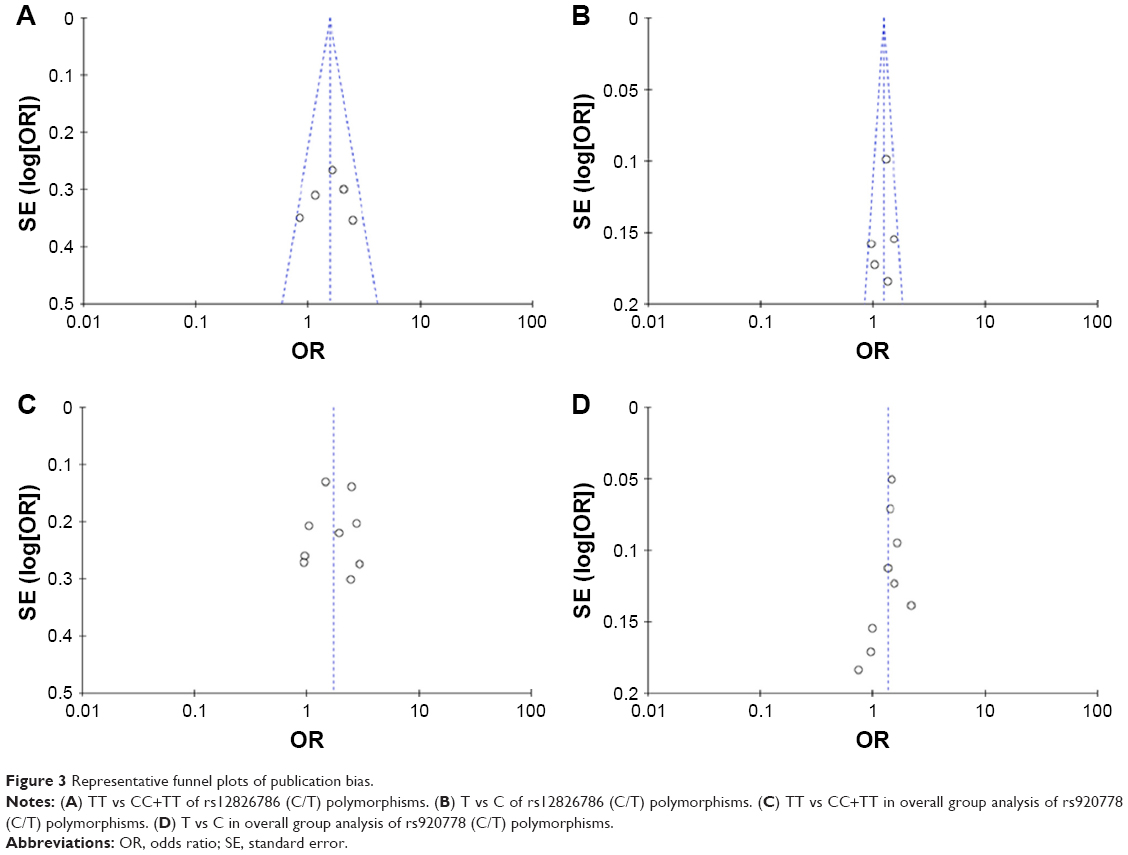 | Figure 3 Representative funnel plots of publication bias. |
Discussion
lncRNAs are a crucial class of RNAs involved in multiple biologic processes such as proliferation and progression of cancer, despite so often being branded as transcriptional noise.38,39 Especially, lncRNA HOTAIR, which is coded from the HOXC locus, has been identified to participate in the development and metastasis of malignancies.40,41 Several biochemical studies suggested that HOTAIR could not only increase Polycomb Repressive Complex 2 recruitment to the genomic positions of target genes to promote malignant transformation but also sponge miR-331-3p to regulate HER2 expression.42–44 Clinically, unregulated expression of HOTAIR was found to be a powerful indicator of poor prognosis for several cancers.45,46 Since the Homo sapiens HOTAIR gene contains many SNPs, recent molecular epidemiologic studies have focused on the association between HOTAIR polymorphisms and cancer susceptibility. Although multiple SNPs and cancer types were explored, no consensus was reached, possibly due to limited sample sizes and variant participant characteristics. In order to draw a more concrete conclusion, we comprehensively searched the existing publications and performed a meta-analysis for six HOTAIR polymorphisms and 10 cancer types by enrolling 36 studies from 18 articles.
Our results showed that polymorphisms of rs12826786 (C/T) and rs920778 (C/T) were correlated with increased cancer risk. Both T-containing genotypes and T alleles were correlated with cancer susceptibility, especially in Chinese population. HOTAIR rs12826786 (C/T) and rs920778 polymorphisms are respectively located within an intronic promoter region and enhancer region, where specific mutations may exert a genotype-specific transcriptional effect on HOTAIR expression.33,35 Previous luciferase assay showed that the substitution of cytosine (C) by thymine (T) in either of the two loci could result in a higher HOTAIR expression, which was pervasively detected in both primary and metastasized tumors of breast cancer, colorectal cancer, lung cancer, and others.19,23,24 Moreover, recent case–control studies revealed that high expression of HOTAIR was correlated with lower survival rates.29,33,37 All these findings are consistent with the results of this meta-analysis, highlighting the roles of SNPs in cancer risk and prognosis.
Our meta-analysis also indicated that the remaining four polymorphisms (rs7958904, rs4759314, rs874945, and rs1899663) were not associated with cancer risk. We noticed that the results for rs7958904 (G/C) polymorphisms were different from a previous meta-analysis, which indicated a decreased cancer risk for G-to-C mutation. The reason lies in the inclusion of the cervical cancer study.18 The authors discovered that the rs7958904 (G/C) polymorphisms conferred an increased risk of cervical cancer. By performing functional assay and MTT assay, they found a higher HOTAIR expression in cervical cancer tissues with rs7958904 CC genotype and a growth-promoting role of rs7958904 C allele on cervical cancer cell line. However, the other three studies reported opposite results in both case–control studies and biochemical assays.19,32,37 As a result, our meta-analysis showed an insignificant result. This reflects the complex function of HOTAIR gene and its variants.18 It also suggests that HOTAIR polymorphisms might play different biologic roles in different types of cancer.
Despite our efforts to include all the eligible publications, several limitations to our meta-analysis should be noticed. First, the populations of included studies were Chinese, Turkish, Iranian, and Portuguese. It is epidemiologically known that other ethnicities such as blacks and Hispanics are also caner susceptible;47,48 thus, the lack of data for these populations might affect the overall results. However, it is worth noting that our stratified analysis may draw a more convincing conclusion for Chinese population. Second, our evaluation mainly focused on unadjusted results due to the insufficient data of several risk factors, while the majority of studies only matched age and gender between cases and controls; the possible imbalance among risk factors may cause distorted results. Moreover, although the number of pooled participants was so far the largest, the included cancer types were still limited. Thus, caution must be preserved when explaining the results to other cancers, especially when we noticed that HOTAIR polymorphisms might function differently in different cancers. Therefore, more studies on various cancer types are needed to help reach a consensus.
In conclusion, our meta-analysis showed that HOTAIR polymorphisms of rs12826786 and rs920778 were correlated with increased cancer risk, while rs7958904, rs4759314, rs874945, and rs1899663 were not. Our meta-analysis was the first to explore the relationship between rs12826786 polymorphisms and cancer susceptibility. It also raised the statistic evidence of discrepant HOTAIR behaviors in different cancer types. Clarifying the environmental and lifestyle risk factors and exploring wider types of cancer are required for future studies to help us draw a concrete conclusion.
Acknowledgment
This work was supported by the National Natural Science Foundation of China (No 81372246) and the National High Technology Research and Development Program of China (No 2014AA020708). The funders had no role in the study design, data extraction and analysis, decision to publish, or preparation of the manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
Nagano T, Fraser P. No-nonsense functions for long noncoding RNAs. Cell. 2011;145(2):178–181. | ||
Birney E, Stamatoyannopoulos JA, Dutta A, et al. Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Nature. 2007;447(7146):799–816. | ||
Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10(3):155–159. | ||
Carninci P, Hayashizaki Y. Noncoding RNA transcription beyond annotated genes. Curr Opin Genet Dev. 2007;17(2):139–144. | ||
Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):629–641. | ||
Zhang H, Chen Z, Wang X, Huang Z, He Z, Chen Y. Long non-coding RNA: a new player in cancer. J Hematol Oncol. 2013;6:37. | ||
Gupta RA, Shah N, Wang KC, et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010;464(7291):1071–1076. | ||
Gutschner T, Diederichs S. The hallmarks of cancer: a long non-coding RNA point of view. RNA Biol. 2012;9(6):703–719. | ||
Tsai MC, Manor O, Wan Y, et al. Long noncoding RNA as modular scaffold of histone modification complexes. Science. 2010;329(5992):689–693. | ||
Rinn JL, Kertesz M, Wang JK, et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell. 2007;129(7):1311–1323. | ||
Qiu JJ, Lin YY, Ye LC, et al. Overexpression of long non-coding RNA HOTAIR predicts poor patient prognosis and promotes tumor metastasis in epithelial ovarian cancer. Gynecol Oncol. 2014;134(1):121–128. | ||
Sun W, Yang Y, Xu C, Xie Y, Guo J. Roles of long noncoding RNAs in gastric cancer and their clinical applications. J Cancer Res Clin Oncol. 2016;142(11):2231–2237. | ||
Liu XH, Liu ZL, Sun M, Liu J, Wang ZX, De W. The long non-coding RNA HOTAIR indicates a poor prognosis and promotes metastasis in non-small cell lung cancer. BMC Cancer. 2013;13:464. | ||
Mu X, Zhao J, Yuan X, et al. Gene Polymorphisms of Toll-Like Receptor 9–1486T/C and 2848G/A in Cervical Cancer Risk. Int J Gynecol Cancer. 2015;25(7):1173–1178. | ||
Mu X, Du X, Yao K, et al. Association between HSD17B1 rs6,05,059 polymorphisms and the risk of uterine diseases: a systemic review and meta-analysis. Int J Clin Exp Pathol. 2015;8(6):6012–6018. | ||
Guo W, Dong Z, Bai Y, et al. Associations between polymorphisms of HOTAIR and risk of gastric cardia adenocarcinoma in a population of north China. Tumor Biol. 2015;36(4):2845–2854. | ||
Ulger Y, Dadas E, Yalinbas Kaya B, Sumbul AT, Genc A, Bayram S. The analysis of lncRNA HOTAIR rs12826786 C>T polymorphism and gastric cancer susceptibility in a Turkish population: lack of any association in a hospital-based case-control study. Ir J Med Sci. 2017;186(4):859–865. | ||
Jin H, Lu X, Ni J, et al. HOTAIR rs7958904 polymorphism is associated with increased cervical cancer risk in a Chinese population. Sci Rep. 2017;7(1):3144. | ||
Xue Y, Gu D, Ma G, et al. Genetic variants in lncRNA HOTAIR are associated with risk of colorectal cancer. Mutagenesis. 2015;30(2):303–310. | ||
Chu H, Chen Y, Yuan Q, et al. The HOTAIR, PRNCR1 and POLR2E polymorphisms are associated with cancer risk: a meta-analysis. Oncotarget. 2017;8(26):43271–43283. | ||
Lv Z, Xu Q, Yuan Y. A systematic review and meta-analysis of the association between long non-coding RNA polymorphisms and cancer risk. Mutat Res. 2017;771:1–14. | ||
Sharma Saha S, Roy Chowdhury R, Mondal NR, et al. Identification of genetic variation in the lncRNA HOTAIR associated with HPV16-related cervical cancer pathogenesis. Cell Oncol (Dordr). 2016;39(6):559–572. | ||
Gong WJ, Yin JY, Li XP, et al. Association of well-characterized lung cancer lncRNA polymorphisms with lung cancer susceptibility and platinum-based chemotherapy response. Tumor Biol. 2016;37(6):8349–8358. | ||
Bayram S, Sumbul AT, Batmaci CY, Genc A. Effect of HOTAIR rs920778 polymorphism on breast cancer susceptibility and clinicopathologic features in a Turkish population. Tumor Biol. 2015;36(5):3863–3870. | ||
Bayram S, Sumbul AT, Dadas E. A functional HOTAIR rs12826786 C>T polymorphism is associated with breast cancer susceptibility and poor clinicopathological characteristics in a Turkish population: a hospital-based case-control study. Tumor Biol. 2016;37(4):5577–5584. | ||
Bayram S, Ulger Y, Sumbul AT, et al. A functional HOTAIR rs920778 polymorphism does not contributes to gastric cancer in a Turkish population: a case-control study. Fam Cancer. 2015;14(4):561–567. | ||
Du M, Wang W, Jin H, et al. The association analysis of lncRNA HOTAIR genetic variants and gastric cancer risk in a Chinese population. Oncotarget. 2015;6(31):31255–31262. | ||
Pan W, Liu L, Wei J, et al. A functional lncRNA HOTAIR genetic variant contributes to gastric cancer susceptibility. Mol Carcinog. 2016;55(1):90–96. | ||
Qiu H, Wang X, Guo R, et al. HOTAIR rs920778 polymorphism is associated with ovarian cancer susceptibility and poor prognosis in a Chinese population. Future Oncol. 2017;13(4):347–355. | ||
Qiu H, Liu Q, Li J, et al. Analysis of the association of HOTAIR single nucleotide polymorphism (rs920778) and risk of cervical cancer. Apmis. 2016;124(7):567–573. | ||
Taheri M, Habibi M, Noroozi R, et al. HOTAIR genetic variants are associated with prostate cancer and benign prostate hyperplasia in an Iranian population. Gene. 2017;613:20–24. | ||
Wu H, Shang X, Shi Y. Genetic variants of lncRNA HOTAIR and risk of epithelial ovarian cancer among Chinese women. Oncotarget. 2016;7(27):41047–41052. | ||
Xavier-Magãlhaes A, Oliveira AI, de Castro JV, et al. Effects of the functional HOTAIR rs920778 and rs12826786 genetic variants in glioma susceptibility and patient prognosis. J Neurooncol. 2017;132(1):27–34. | ||
Yan R, Cao J, Song C, Chen Y, Wu Z, Wang K, Dai L. Polymorphisms in lncRNA HOTAIR and susceptibility to breast cancer in a Chinese population. Cancer Epidemiol. 2015;39(6):978–985. | ||
Zhang X, Zhou L, Fu G, et al. The identification of an ESCC susceptibility SNP rs920778 that regulates the expression of lncRNA HOTAIR via a novel intronic enhancer. Carcinogenesis. 2014;35(9):2062–2067. | ||
Zhu H, Lv Z, An C, et al. Onco-lncRNA HOTAIR and its functional genetic variants in papillary thyroid carcinoma. Sci Rep. 2016;6:31969. | ||
Zhu Z, Ni B, Zhou R, Gan T. lncRNA HOTAIR polymorphisms and the sucesptability and prognosis of colorectal cancer. Guangdong Med J. 2016;37(19):2916–2919. | ||
Bhan A, Mandal SS. LncRNA HOTAIR: a master regulator of chromatin dynamics and cancer. Biochim Biophys Acta. 2015;1856(1):151–164. | ||
Cai B, Song XQ, Cai JP, Zhang S. HOTAIR: a cancer-related long non-coding RNA. Neoplasma. 2014;61(4):379–391. | ||
Wang YL, Overstreet AM, Chen MS, et al. Combined inhibition of EGFR and c-ABL suppresses the growth of triple-negative breast cancer growth through inhibition of HOTAIR. Oncotarget. 2015;6(13):11150–11161. | ||
Lee NK, Lee JH, Park CH, et al. Long non-coding RNA HOTAIR promotes carcinogenesis and invasion of gastric adenocarcinoma. Biochem Biophys Res Commun. 2014;451(2):171–178. | ||
Wu L, Murat P, Matak-Vinkovic D, Murrell A, Balasubramanian S. Binding interactions between long noncoding RNA HOTAIR and PRC2 proteins. Biochemistry. 2013;52(52):9519–9527. | ||
Liu XH, Sun M, Nie FQ, et al. Lnc RNA HOTAIR functions as a competing endogenous RNA to regulate HER2 expression by sponging miR-331–3p in gastric cancer. Mol Cancer. 2014;13:92. | ||
Portoso M, Ragazzini R, Brencic Z, et al. PRC2 is dispensable for HOTAIR-mediated transcriptional repression. EMBO J. 2017;36(8):981–994. | ||
Kogo R, Shimamura T, Mimori K, et al. Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin modification and is associated with poor prognosis in colorectal cancers. Cancer Res. 2011;71(20):6320–6326. | ||
Zhang YY, Huang SH, Zhou HR, Chen CJ, Tian LH, Shen JZ. Role of HOTAIR in the diagnosis and prognosis of acute leukemia. Oncol Rep. 2016;36(6):3113–3122. | ||
Bandera EV, Maskarinec G, Romieu I, John EM. Racial and ethnic disparities in the impact of obesity on breast cancer risk and survival: a global perspective. Adv Nutr. 2015;6(6):803–819. | ||
Fiscella K, Winters P, Farah S, Sanders M, Mohile SG. Do lung cancer eligibility criteria align with risk among blacks and hispanics? PLoS One. 2015;10(11):e0143789. | ||
Wells GA, Shea B, O’Connell D, Peterson J, Welch V, Losos M, Tugwell P. The Newcastle-Ottawa Scale. http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp. Accessed July 15, 2017. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.