")
Back to Journals » OncoTargets and Therapy » Volume 13
Tetrandrine Reverses Paclitaxel Resistance in Human Ovarian Cancer via Inducing Apoptosis, Cell Cycle Arrest Through β-Catenin Pathway
Received 21 October 2019
Accepted for publication 28 March 2020
Published 30 April 2020 Volume 2020:13 Pages 3631—3639
DOI https://doi.org/10.2147/OTT.S235533
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Takuya Aoki
Luo Jiang,1 Rui Hou2
1Department of Ultrasound, Shengjing Hospital of China Medical University, Shenyang, Liaoning 110004, People’s Republic of China; 2Department of Obstetrics and Gynecology, Shengjing Hospital of China Medical University, Shenyang, Liaoning 110004, People’s Republic of China
Correspondence: Rui Hou
Department of Obstetrics and Gynecology, Shengjing Hospital of China Medical University, 36 Sanhao Street, Heping District, Shenyang 110004, People’s Republic of China
Email [email protected]
Background: Paclitaxel (PTX) resistance is a great obstacle for the treatment of ovarian cancer. A previous study indicated that tetrandrine (TET) could induce the apoptosis of ovarian cancer cells. This study aimed to explore the effect of TET in combination with PTX on PTX resistance in ovarian cancer cells.
Materials and Methods: CCK-8 assay, flow cytometry and wound healing assays were used to detect the proliferation, apoptosis and migration of PTX-resistant SKOV3 cells (SKOV3/PTX). The expressions of Bax, Bcl-2, cleaved caspase 3, β-catenin, c-Myc, cyclin D1 and p21 in SKOV3/PTX cells were detected with Western blot. In vivo animal study was performed finally.
Results: In this study, the inhibitory effects of PTX on the proliferation and migration of SKOV3/PTX cells were markedly enhanced by TET. In addition, PTX-induced apoptosis in SKOV3/PTX cells was significantly enhanced by the treatment of TET via upregulating the levels of Bax and cleaved caspase 3, and downregulating the expression of Bcl-2. Moreover, combination of TET and PTX obviously induced cell cycle arrest in SKOV3/PTX cells via increasing the level of p21 and decreasing the levels of c-Myc and Cyclin D1. Meanwhile, combination of TET with PTX significantly decreased the expression of β-catenin in SKOV3/PTX cells. In vivo experiments further confirmed that TET enhanced the anti-tumor effect of PTX in SKOV3/PTX xenograft model.
Conclusion: We found that TET could enhance the sensitivity of SKOV3/PTX cells to PTX via inhibiting the β-catenin/c-Myc/Cyclin D1 signaling pathway. Therefore, PTX combined with TET might be considered as a potential approach for the treatment of PTX-resistant ovarian cancer.
Keywords: ovarian cancer, paclitaxel, tetrandrine, apoptosis, cell cycle arrest
Introduction
Ovarian cancer is the fourth most common cancer among women, especially in older women.1 It often occurs in peri- and post-menopausal women.2 Age is the main cause, more than 48% of ovarian cancers occur in women over the age of 65 years.3 According to the International Federation of Gynecology and Obstetrics (FIGO) system, ovarian cancer is divided into four stages: range from stage I to IV.4 Stage I and stage II tumors are called early-stage cancer which spread is limited to the pelvic cavity, and stage III and stage IV tumors that spread outside of the pelvic cavity are called advanced or metastatic cancer.4,5 Currently, chemotherapy and surgery are the main therapeutic methods for the treatment of ovarian cancer.6,7 Previous study found that chemotherapy could decrease the tumor recurrence rate and metastasis rate, whereas drug resistance occurred frequently after long-term chemotherapy.8
Paclitaxel (PTX) is a first-line chemotherapy drug for the treatment of ovarian cancer.9 However, PTX resistance is one of the key factors affecting the effect of chemotherapy, which markedly limit the clinical usage of PTX.10,11 Therefore, searching new agents to overcome cancer chemoresistance is a novel effective therapy for the treatment of ovarian cancer.
Tetrandrine (TET) is one of the most effective component of the dried root of traditional Chinese medicine Stephania tetrandra S.12 Recent studies have revealed that TET exhibited beneficial effects in a number of types of human cancers including lung cancer, breast cancer, colon cancer and cervical cancer.13–16 Li et al indicated that TET administration induced the growth of colon carcinoma cells via inducing apoptosis and cell cycle arrest.17 Meanwhile, Zhang et al indicated that TET could reverse cisplatin resistance in ovarian cancer cells by inducing apoptosis.18 However, the effect of TET in PTX resistance of ovarian cancer remains unclear. Therefore, we aimed to explore the anti-tumor effect of combination of TET and PTX on PTX-resistant human ovarian cancer cells.
Materials and Methods
Cell Culture
The human ovary cancer cell line SKOV-3 was purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). The human ovary cancer cell line A2780 was purchased from Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). PTX-resistant SKOV-3 cell line (SKOV3/PTX) were established by long-term exposure of SKOV-3 cells to a stepwise gradually concentration of PTX over 7 months. Cells were cultured in DMEM medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS, Thermo Fisher Scientific), and 1% penicillin-streptomycin at 37°C in an atmosphere of 5% CO2.
CCK-8 Assay
Cell viability was determined using a Cell Counting Kit 8 (CCK-8, Abcam Cambridge, MA, USA). SKOV-3, A2780 or SKOV3/PTX cells were seeded into 96-well plates at a density of 5 × 103 cells/well and incubated at 37°C overnight. After that, cells were treated with different concentrations of PTX for 72 h. Subsequently, 10 µL of CCK8 reagent was added into each well and then incubated for another 2 h. Finally, the optical density (OD) value of cells was measured at 450 nm using a microplate reader (BioTek, Winooski, VT, USA).
Immunofluorescence Staining Assay
SKOV3/PTX cells (5 × 103 cells/well) were seeded onto 96-well plates and incubated at 37°C overnight. Cells were fixed in 3.7% paraformaldehyde for 15 min at room temperature, and then permeabilized with 0.2% Triton X-100 for 5 min. After that, the cells were washed twice with PBS and then incubated with the primary antibody anti-Ki67 (1:1000, Abcam) at 4°C overnight. Later on, cells were incubated with a fluorescence-conjugated secondary antibody (1:2000, Abcam) at 37°C for 1 h. Nuclei was counterstained with DAPI for 10 min. A fluorescence microscope (Olympus, Tokyo, Japan) was applied to detect the fluorescence signal.
Apoptosis Assay by Flow Cytometry
The Annexin V-FITC apoptosis detection kit was used to measure cell apoptosis (Thermo Fisher Scientific). SKOV3/PTX cells were incubated with PTX or/and TET for 72 h at 37°C. After that, cells were collected and washed twice with PBS, and then stained with 5 μL annexin V-FITC and 5 μL propidium iodide for 30 min in the dark. Finally, apoptosis cells were analyzed using a FACScan flow cytometer (BD Biosciences, Mountain View, CA, USA).
JC-1 Staining
Mitochondrial membrane potential (MMP) was assessed by cationic dye JC-1 using the MMP assay kit (Thermo Fisher Scientific) according to the manufacturer’s protocol. Briefly, SKOV3/PTX cells were collected and washed twice with PBS. Later on, cells were incubated with 10 mg/mL JC-1 reagent at 37°C for 20 min in the dark. After that, cells were resuspended in PBS for analysis by the flow cytometer, as previously described.19,20
Western Blot
BCA protein Assay kit (Beyotime Institute of Biotechnology) was applied to determine the concentration of protein. Proteins from each sample were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and then transferred onto the polyvinylidene difluoride membrane (PVDF, Thermo Fisher Scientific). After that, the membrane was blocked in 5% nonfat milk powder in Tris-buffered saline containing 0.1% Tween 20 (TBST) for 1 h at room temperature. Then, the membrane was incubated in primary antibodies at 4°C overnight, including anti-Cyclin D1 (1:1000, Abcam), anti-p21 (1:1000, Abcam), anti-Bax (1:1000, Abcam), anti-Bcl-2 (1:1000, Abcam), anti-cleaved caspase 3 (1:1000, Abcam), anti-c-Myc (1:1000, Abcam), and anti-β-actin (1:1000, Abcam). After washing with TBST, membranes were incubated with goat anti-rabbit secondary antibodies (1:5000, Abcam) for 1 h at the room temperature. Then, the protein bands were visualized using an ECL Chemiluminescent Substrate Reagent Kit (Thermo Fisher Scientific). β-actin was acted as the internal control.
Wound Healing Assay
SKOV3/PTX cells were seeded into six wells culture plate overnight at 37°C. The cellular monolayer was wounded with a sterile micropipette tip. Then, cells were incubated with PTX or/and TET for 24 h at 37°C. Images of the scratched wounds were observed at 0 h and 24 h using the fluorescence microscope (Olympus CX23 Tokyo, Japan). The migrated areas were determined using ImageJ software (version 1.52).
Cell Cycle
SKOV3/PTX cells were incubated with PTX or/and TET for 24 h at 37°C. After that, cells were fixed in ice-cold 70% ethanol at 4°C overnight. Then, cells were resuspended and washed twice with PBS. Later on, cells were incubated with staining buffer containing 50 μg/mL PI in the dark at 4°C for 30 min. Finally, cell-cycle distribution was analyzed by a flow cytometer (BD Biosciences).
Animal Study
BALB/c nude female mice (n = 12, 6–8 weeks old) were obtained from Animal Center of Chinese Academy of Sciences (Shanghai, China). All mice were maintained following the guidelines of the Institutional Animal Care and Use Committee. These animal experiments were performed approved by the Ethics Committees of Shengjing Hospital Affiliated to China Medical University. SKOV3/PTX cells (5 x 106 cells) were transplanted subcutaneously into the right flank in each animal. When the tumor volume reached about 180 mm3, the mice were randomized into four groups: vehicle, PTX_10 mg/kg, TET_50 mg/kg and PTX (10 mg/kg) + TET (50 mg/kg) groups. TET was administered via intraperitoneal (i.p.) injection daily. PTX was injected intravenously once every three days. Tumor volume was monitored weekly and analyzed using the formula V = (length x width2)/2. After 3 weeks, mice were sacrificed and tumor tissues were weighted.
Statistical Analysis
All data were repeated in triplicate. All statistical analyses were performed using GraphPad Prism software (version 7.0, La Jolla, CA, USA). One-way analysis of variance (ANOVA) and Tukey’s tests were carried out for multiple group comparisons. All data were presented as mean ± standard error (S. D.). A P-value < 0.05 was considered as a statistically significant.
Results
TET Enhanced the Anti-Proliferation Effect of PTX in SKOV3/PTX Cells
First, we established a PTX-resistant SKOV-3 cell line (SKOV3/PTX). Cell proliferation assay indicated that SKOV3/PTX cells were resistant to PTX after gradient treatment compared with SKOV3 parental cells (Figure 1A). To evaluate the effects of TET on the viability of PTX-resistance SKOV3 cells, CCK-8 assay was used. As shown in Figure 1B, TET (2, 4 or 8 μM) significantly inhibited the proliferation of SKOV3/PTX cells. In addition, TET (2, 4 or 8 μM) notably enhanced the anti-proliferation effect of PTX (1 μM) on SKOV3/PTX cells (Figure 1C). Moreover, combination of 1 μM PTX with 2 μM TET induced about 50% growth inhibition in SKOV3/PTX cells (Figure 1C). Therefore, 2 μM TET was utilized in the following experiments.
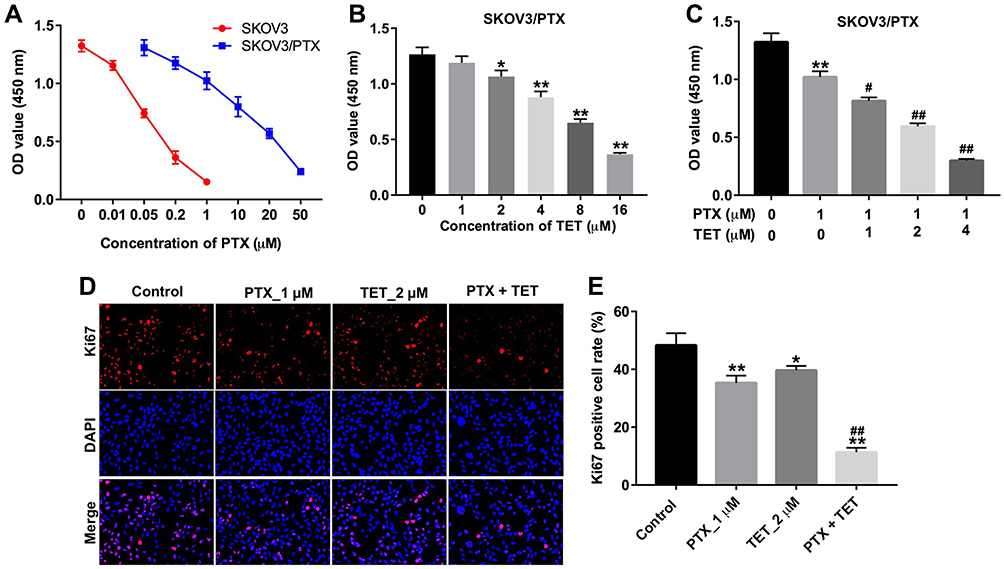 |
Figure 1 TET enhanced the anti-proliferation effect of PTX in SKOV3/PTX cells. (A) CCK-8 assay was used to detect the cell viability. SKOV3 cells were treated with 0, 0.01, 0.05, 0.2, or 1 μM PTX for 72 h. SKOV3/PTX cells were treated with 0, 0.05, 0.2, 1, 10, 20 or 50 μM PTX for 72 h. (B) SKOV3/PTX cells were treated with 0, 1, 2, 4 or 8 μM TET for 72 h. (C) SKOV3/PTX cells were treated with PTX (1 μM) and TET (0, 1, 2 or 4 μM) for 72 h. (D, E) SKOV3/PTX cells were treated with PTX (1 μM) or/and 2 μM TET for 72 h. Quantification of Ki67 expression by immunofluorescence in SKOV3/PTX cells. *P<0.05, **P<0.01 compared with control group; #P<0.05, ##P<0.01 compared with PTX_1 μM group. |
Next, we found PTX significantly decreased the number of Ki67-positive SKOV3/PTX cells compared to control group. As expected, the Ki67-positive cells were further obviously reduced in the presence of TET, compared with PTX treatment group (Figure 1D and E). Furthermore, as shown in Table 1, IC50 value of TET (alone treatment) was 8.48 µM in SKOV3/PTX cells. However, when TET was combined with PTX (1 μM), the IC50 value of TET was decreased to 1.5 μM. In addition, the CI value of TET combined with PTX in SKOV3/PTX cells was less than 0.3, which indicated the strong synergism effect (Table 1). These data indicated that combination of TET with PTX had synergistic effect against SKOV3/PTX cells.
 |
Table 1 Evaluation of Combination of PTX with TET in Ovarian Cancer (72 h Treatment) |
In addition, as shown in Supplementary Figure 1A–D, TET or PTX significantly inhibited the proliferation of parental SKOV3 and A2780 cells. As expected, combination treatment of TET and PTX markedly suppressed the proliferation of SKOV3 and A2780 cells, compared with PTX treatment group (Supplementary Figure 1A–D). These results indicated that TET could enhance the anti-proliferation effect of PTX in ovarian cancer cells.
TET Enhanced the Pro-Apoptotic Effect of PTX in SKOV3/PTX Cells
Flow cytometric assay was used to evaluate the apoptosis in SKOV3/PTX cells. As illustrated in Figure 2A and B, PTX notably induced the apoptosis of SKOV3/PTX cells. As expected, the apoptotic cells were further increased in the presence of TET, compared with PTX treatment group. In addition, combination of PTX with TET markedly decreased the MMP in SKOV3/PTX cells, compared with PTX treatment group (Figure 2C). Moreover, PTX significantly increased the levels of Bax and cleaved caspase 3, but decreased the expression of Bcl-2 in SKOV3/PTX cells. As expected, the levels of Bax and active caspase 3 in cells were further increased in the presence of TET, while the expression of Bcl-2 was further decreased, compared with PTX treatment group (Figure 2D–G). These results indicated that TET could enhance the pro-apoptotic effect of PTX in SKOV3/PTX cells.
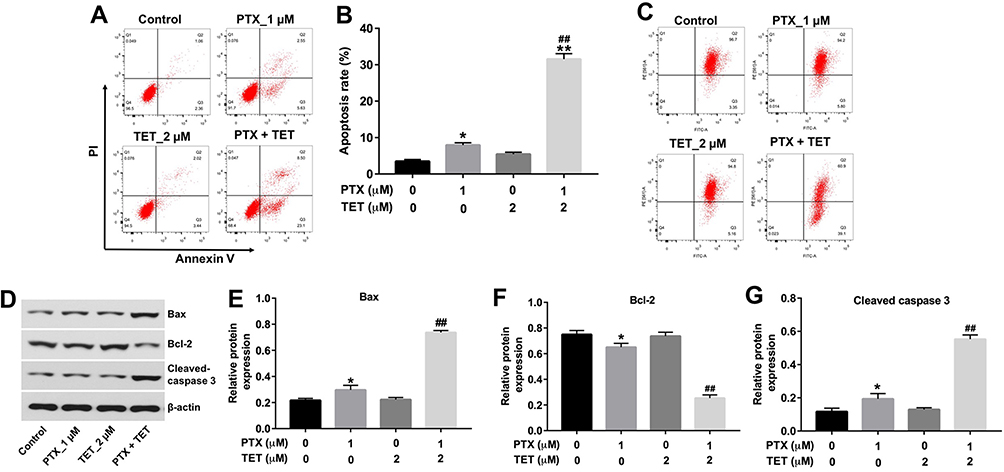 |
Figure 2 TET enhanced the pro-apoptotic effect of PTX in SKOV3/PTX cells. SKOV3/PTX cells were treated with PTX (1 μM) or/and 2 μM TET for 72 h. (A, B) Apoptotic cells were detected by Annexin V/PI double staining and flow cytometry in SKOV3/PTX cells. (C) JC-1 staining was used to determine MMP loss. (D) Expression levels of Bax, Bcl-2 and cleaved caspase 3 in SKOV3/PTX cells were detected with Western blotting. (E–G) The relative expressions of Bax, Bcl-2 and cleaved caspase 3 were quantified via normalization to β-actin. *P<0.05, **P<0.01 compared with control group; ##P<0.01 compared with PTX_1 μM group. |
TET Enhanced the Anti-Migratory Effect of PTX in SKOV3/PTX Cells
Next, the effect of TET and PTX on the migration of SKOV3/PTX cells was analyzed by wound-healing assay. As indicated in Figure 3A and B, PTX or TET obviously suppressed the migration ability of SKOV3/PTX cells. Meanwhile, combination of PTX with TET further inhibited the migration ability of SKOV3/PTX cells, compared with PTX alone treatment groups (Figure 3A and B). These results indicated that TET could enhance the anti-migratory effect of PTX in SKOV3/PTX cells.
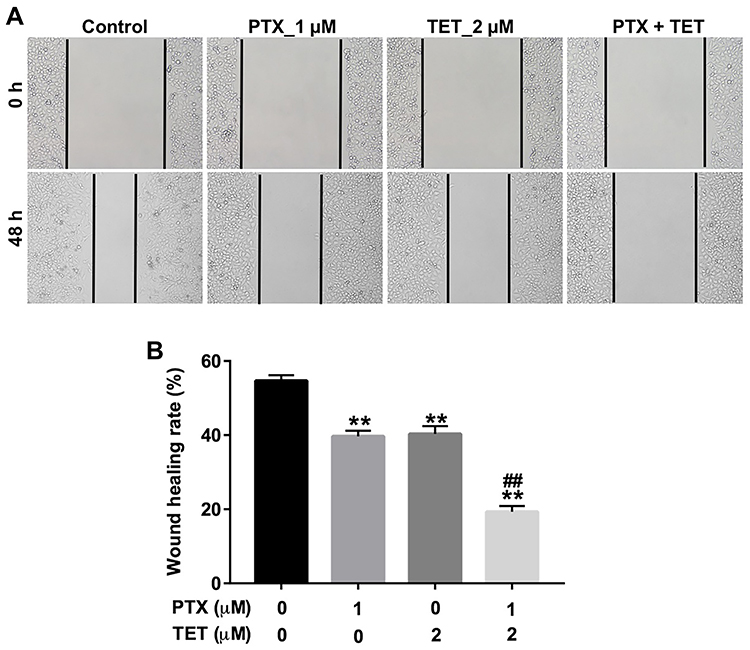 |
Figure 3 TET enhanced the anti-migratory effect of PTX in SKOV3/PTX cells. SKOV3/PTX cells were treated with PTX (1 μM) or/and 2 μM TET for 72 h. (A, B) Cell migration was determined by wound healing assay. **P<0.01 compared with control group; ##P<0.01 compared with PTX_1 μM group. |
TET Enhanced the Sensitivity of SKOV3/PTX Cells to PTX by Inhibiting the β-Catenin/c-Myc/Cyclin D1 Signaling Pathway
The effect of TET and PTX on cell cycle progression was determined by flow cytometry and Western blotting. As shown in Figure 4A and B, the percentage of cells in the G0-G1 phase was obviously increased, with a concomitant decrease of cells in the S phase in the combination treatment group, compared to PTX treatment group. In addition, PTX had very limited effects on the expressions of β-catenin, c-Myc, cyclin D1 and p21 in SKOV3/PTX cells, whereas TET or combination treatment significantly downregulated the expressions of β-catenin, c-Myc and cyclin D1 and upregulated the level of p21 in SKOV3/PTX cells (Figure 4C–G). Furthermore, combination treatment significantly inhibited the proliferation of SKOV3/PTX cells via inducing apoptosis, which were markedly reversed in the presence of R-spondin 1 (β-catenin signal pathway activator) (Supplementary Figure 2A–C). These results indicated that TET could attenuate PTX resistance in SKOV3/PTX cells via inhibiting the β-catenin/c-Myc/Cyclin D1 signaling pathway.
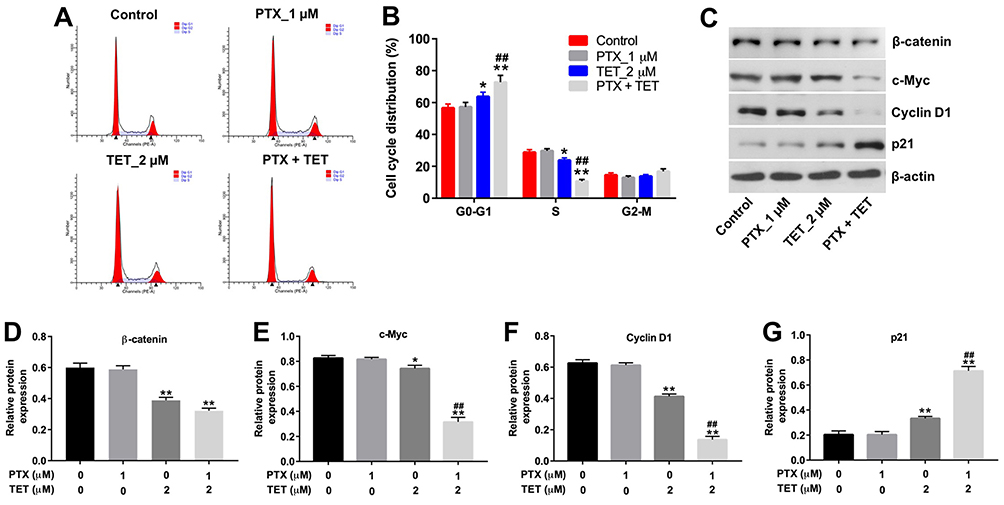 |
Figure 4 TET enhanced the sensitivity of SKOV3/PTX cells to PTX by inhibiting the β-catenin/c-Myc/Cyclin D1 signaling pathway. SKOV3/PTX cells were treated with PTX (1 μM) or/and 2 μM TET for 72 h. (A, B) Flow cytometry was used to measure cell cycle phase. (C) Expression levels of c-Myc, cyclin D1 and p21 in SKOV3/PTX cells were detected with Western blotting. (D–G) The relative expressions of β-catenin, c-Myc, cyclin D1 and p21 were quantified via normalization to β-actin. *P<0.05, **P<0.01 compared with control group; ##P<0.01 compared with PTX_1 μM group. |
TET Enhanced Anti-Tumor Effect of PTX in SKOV3/PTX Xenograft
Cell line-derived xenograft model was established to investigate the effect of TET and PTX on ovarian tumor growth in vivo. As indicated in Figure 5A–C, the tumor volume and tumor weight in PTX group was significantly decreased compared with control group, which were further reduced in the presence of TET. In addition, Western blotting results indicated that combination of PTX with TET treatment significantly downregulated the levels of c-Myc and Cyclin D1, but notably upregulated the expression of p21 in tumor tissues compared with PTX treatment group (Figure 5D–G). These results illustrated that TET could enhance the anti-tumor effect of PTX in SKOV3/PTX xenograft.
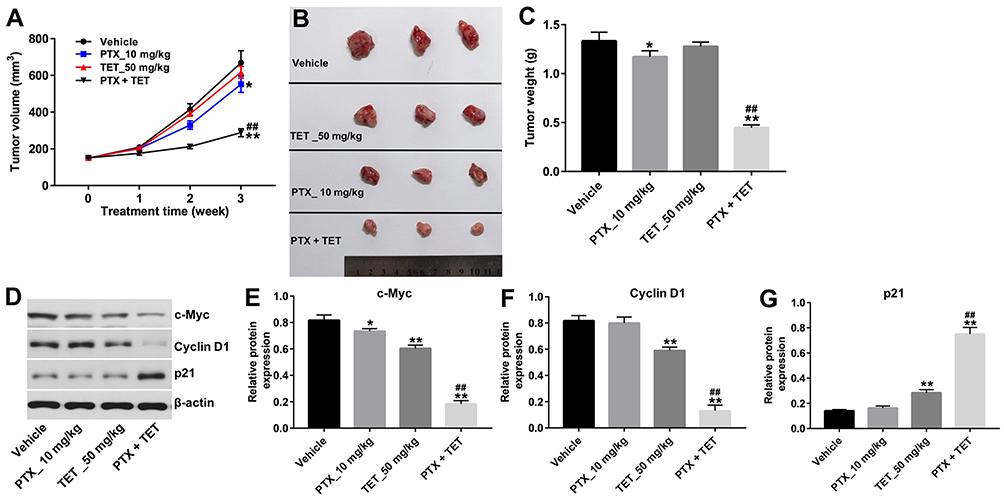 |
Figure 5 TET enhanced anti-tumor effect of PTX in SKOV3/PTX xenograft. SKOV3/PTX cells were transplanted subcutaneously into nude mice to establish a xenograft model of ovarian cancer. (A) Subcutaneous tumor xenograft volumes were monitored and measured weekly. (B) Representative photographs of SKOV3/PTX xenograft tumor were captured on day 21. (C) Tumor weights in each group were calculated. (D) Expression levels of c-Myc, cyclin D1 and p21 in tumor tissues were detected with Western blotting. (E–G) The relative expressions of c-Myc, cyclin D1 and p21 were quantified via normalization to β-actin. *P<0.05, **P<0.01 compared with vehicle group; ##P<0.01 compared with PTX_10 mg/kg group. |
Discussion
Chemotherapy plays an important role in the treatment of human cancers, whereas drug resistance is one of the major challenges in the fight against ovarian cancer.10,21 Recently, traditional Chinese medicine (TCM) combined with conventional chemotherapeutic agents has received growing attention.22 Previous studies indicated that combination of TCM and chemotherapeutic drugs could overcome drug resistance.23,24 Clinically, patients with advanced non-small cell lung cancer received TET combined with gemcitabine and cisplatin treatment. The results indicated that TET could mitigate adverse reactions to chemotherapy for patients with advanced non-small cell lung cancer.25 However, the effect of the combination of PTX and TET on PTX-resistance ovarian cancer cells remains unclear. In this study, we found that combination of TET and PTX significantly enhanced the anti-tumor effect of PTX in vitro and in vivo.
Previous study indicated that TET exerted multiple biological activities on cancer cells, such as induction of apoptosis, inhibition of proliferation and reversal of multidrug resistance.26 Wang et al found that TET enhanced the sensitivity of drug-resistant esophageal squamous carcinoma cells to cisplatin via suppressing of multidrug resistance-associated protein 1.27 Zou et al indicated that tetramethylpyrazine enhances the anti-tumor effect of PTX in ovarian cancer via inducing apoptosis.10 In this study, TET enhanced the inhibitory effects of PTX on the proliferation and migration of SKOV3/PTX cells. In addition, TET enhanced the pro-apoptotic effect of PTX in SKOV3/PTX cells via downregulating the expression of Bcl-2, and upregulating the levels of Bax and active caspase 3, which were consistent with previous studies. Meanwhile, combination of PTX with TET notably inhibited the tumor growth in SKOV3/PTX xenograft. These data suggested that TET could enhance the anti-tumor effect of PTX in SKOV3/PTX cells in vitro and in vivo.
Previous study found that overexpression of c-Myc is intently associated with cell proliferation, while downregulation of c-Myc could inhibit the cell cycle progression.28 In addition, evidence has been shown that p21 functioned as a negative regulator of cell cycle progression at G1 and S phase.29 Meanwhile, cyclin D1 is an important regulator in the G1 phase of the cell cycle, increasing the level of Cyclin D1 might contribute to tumorigenesis.30 Zhang et al found that combination of TET with PTX markedly induced apoptosis in ovarian cancer cells via inhibition cell cycle progression.18 Meanwhile, Hou et al indicated that TET enhanced the anti-tumor effect of epirubicin on breast carcinoma multidrug resistance cells.31 Our results found that 1 μM PTX had very limited effect on cell cycle distribution in SKOV3/PTX cells. In addition, 2 μM TET markedly upregulated the level of p21 and downregulated the levels of c-Myc and Cyclin D1 in SKOV3/PTX cells. Singh et al indicated that TET inhibited the growth of pancreatic cancer cells via increasing the level of p21 and decreasing the level of Cyclin D1, which was consistent with our results.32 Meanwhile, Sánchez-Carranza et al found that gallic acid reduced PTX resistance in ovarian carcinoma cells via inducing G2/M cell arrest.33 In this study, by reducing the expressions of cyclin D1 and c-Myc and increasing the level of p21, combination of TET and PTX significantly increased the distribution of G1 phase in cell cycle in SKOV3/PTX cells, and thus induced G1 phase cell cycle arrest. These data indicated that TET enhanced the sensitivity of SKOV3/PTX cells to PTX via inhibiting cell cycle transition at G1/S boundary.
Evidence has been shown that β-catenin, an important transduction factor in the Wnt signaling pathway, plays a vital role in regulating chemoresistance in epithelial ovarian cancer.34 In addition, β-catenin involved in the regulation of cell growth, cell cycle, and apoptosis.35 Moreover, inhibition of β-catenin in cancer cells could suppress the cell cycle distribution via downregulating the expressions of Cyclin D1 and c-Myc.35 Our data indicated that PTX had no effects on the expressions of β-catenin, c-Myc, and Cyclin D1 in SKOV3/PTX cells, whereas TET or combination treatment significantly downregulated the expressions of β-catenin, c-Myc and cyclin D1 in SKOV3/PTX cells, indicating that TET enhanced anti-tumor effect of PTX in SKOV3/PTX cells via inhibition the β-catenin/c-Myc/Cyclin D1 signaling pathway. Meanwhile, Liao et al found that TET could reverse multidrug resistance in cancer cells via downregulating the expression of ABCB1.36 In the future, the association between β-catenin and ABCB1 in SKOV3/PTX cells should be investigated, which may further confirm the roles and mechanisms of TET in regulating ovarian cancer cells resistance.
Conclusion
In this study, we found that TET could enhance the sensitivity of SKOV3/PTX cells to PTX via inducing apoptosis and cell cycle arrest. In addition, TET enhanced anti-tumor effect of PTX in SKOV3/PTX cells via inhibition of the β-catenin/C-myc/Cyclin D1 signaling pathway. Therefore, PTX combined with TET might be considered as a potential approach for the treatment of PTX-resistant ovarian cancer.
Disclosure
The authors declare no competing financial interests.
References
1. Amini E, Baharara J, Afzali M, Nikdel N. The p53 modulated cytotoxicity of ophiocoma scolopendrina polysaccharide against resistance ovarian cancer cells. Avicenna J Med Biotechnol. 2019;11(3):208–214.
2. Pokhriyal R, Hariprasad R, Kumar L, Hariprasad G. Chemotherapy resistance in advanced ovarian cancer patients. Biomark Cancer. 2019;11:1179299x19860815. doi:10.1177/1179299X19860815
3. Yancik R. Ovarian cancer. age contrasts in incidence, histology, disease stage at diagnosis, and mortality. Cancer. 1993;71(2 Suppl):517–523. doi:10.1002/cncr.2820710205
4. Prat J. FIGO’s staging classification for cancer of the ovary, fallopian tube, and peritoneum: abridged republication. J Gynecol Oncol. 2015;26(2):87–89. doi:10.3802/jgo.2015.26.2.87
5. Meinhold-Heerlein I, Fotopoulou C, Harter P, et al. Statement by the kommission ovar of the AGO: the new FIGO and WHO classifications of ovarian, fallopian tube and primary peritoneal cancer. Geburtshilfe Frauenheilkd. 2015;75(10):1021–1027. Doi:10.1055/s-0035-1558079
6. Cowan RA, O’Cearbhaill RE, Zivanovic O, Chi DS. Current status and future prospects of hyperthermic intraoperative intraperitoneal chemotherapy (HIPEC) clinical trials in ovarian cancer. Int J Hyperthermia. 2017;33(5):548–553. doi:10.1080/02656736.2017.1283066
7. Makar AP, Trope CG, Tummers P, Denys H, Vandecasteele K. Advanced ovarian cancer: primary or interval debulking? Five categories of patients in view of the results of randomized trials and tumor biology: primary debulking surgery and interval debulking surgery for advanced ovarian cancer. Oncologist. 2016;21(6):745–754. doi:10.1634/theoncologist.2015-0239
8. Tong X, Zhao J, Zhang Y, Mu P, Wang X. Expression levels of MRP1, GST-pi, and GSK3beta in ovarian cancer and the relationship with drug resistance and prognosis of patients. Oncol Lett. 2019;18(1):22–28. doi:10.3892/ol.2019.10315
9. Guo W, Dong W, Li M, Shen Y. Mitochondria P-glycoprotein confers paclitaxel resistance on ovarian cancer cells. Onco Targets Ther. 2019;12:3881–3891. doi:10.2147/OTT.S193433
10. Zou L, Liu X, Li J, et al. Tetramethylpyrazine enhances the antitumor effect of paclitaxel by inhibiting angiogenesis and inducing apoptosis. Front Pharmacol. 2019;10:707. doi:10.3389/fphar.2019.00707
11. Yusuf RZ, Duan Z, Lamendola DE, Penson RT, Seiden MV. Paclitaxel resistance: molecular mechanisms and pharmacologic manipulation. Curr Cancer Drug Targets. 2003;3(1):1–19. doi:10.2174/1568009033333754
12. Cao Z, Li D, Liu L, Yang P. Effect of five novel 5-substituted tetrandrine derivatives on P-glycoprotein-mediated inhibition and transport in Caco-2 cells. Oncol Lett. 2018;16(5):6808–6814. doi:10.3892/ol.2018.9492
13. Chen Z, Zhao L, Zhao F, Yang G, Wang JJ. Tetrandrine suppresses lung cancer growth and induces apoptosis, potentially via the VEGF/HIF-1alpha/ICAM-1 signaling pathway. Oncol Lett. 2018;15(5):7433–7437. doi:10.3892/ol.2018.8190
14. Guo Y, Pei X. Tetrandrine-induced autophagy in MDA-MB-231 triple-negative breast cancer cell through the inhibition of PI3K/AKT/mTOR signaling. Evid Based Complement Alternat Med. 2019;2019:7517431. doi:10.1155/2019/7517431
15. Juan TK, Liu KC, Kuo CL, et al. Tetrandrine suppresses adhesion, migration and invasion of human colon cancer SW620 cells via inhibition of nuclear factor-kappaB, matrix metalloproteinase-2 and matrix metalloproteinase-9 signaling pathways. Oncol Lett. 2018;15(5):7716–7724. doi:10.3892/ol.2018.8286
16. Zhang H, Xie B, Zhang Z, Sheng X, Zhang S. Tetrandrine suppresses cervical cancer growth by inducing apoptosis in vitro and in vivo. Drug Des Devel Ther. 2019;13:119–127. doi:10.2147/DDDT.S187776
17. Li J, Wang Q, Wang Z, et al. Tetrandrine inhibits colon carcinoma HT-29 cells growth via the Bcl-2/Caspase 3/PARP pathway and G1/S phase. Biosci Rep. 2019;39(5).
18. Zhang Y, Wang C, Wang H, et al. Combination of tetrandrine with cisplatin enhances cytotoxicity through growth suppression and apoptosis in ovarian cancer in vitro and in vivo. Cancer Lett. 2011;304(1):21–32. doi:10.1016/j.canlet.2011.01.022
19. Liang WL, Xiao L, Gu HW, et al. Solid lipid nanoparticle induced apoptosis of macrophages via a mitochondrial-dependent pathway in vitro and in vivo. Int J Nanomedicine. 2019;14:3283–3295. doi:10.2147/IJN.S200395
20. Wu D, Zhou WY, Lin XT, Fang L, Xie CM. Bufalin induces apoptosis via mitochondrial ROS-mediated caspase-3 activation in HCT-116 and SW620 human colon cancer cells. Drug Chem Toxicol. 2019;42(4):444–450. doi:10.1080/01480545.2018.1512611
21. Wang W, Gao Y, Hai J, Yang J, Duan S. HER2 decreases drug sensitivity of ovarian cancer cells via inducing stem cell-like property in an NFkappaB-dependent way. Biosci Rep. 2019;39(3).
22. Chen S, Zhang Z, Zhang J. Emodin enhances antitumor effect of paclitaxel on human non-small-cell lung cancer cells in vitro and in vivo. Drug Des Devel Ther. 2019;13:1145–1153. doi:10.2147/DDDT.S196319
23. Huang MY, Zhang LL, Ding J, Lu JJ. Anticancer drug discovery from Chinese medicinal herbs. Chin Med. 2018;13:35. doi:10.1186/s13020-018-0192-y
24. Sui H, Duan P, Guo P, et al. Zhi Zhen Fang formula reverses hedgehog pathway mediated multidrug resistance in colorectal cancer. Oncol Rep. 2017;38(4):2087–2095. doi:10.3892/or.2017.5917
25. Liu W, Zhang J, Ying C, et al. Tetrandrine combined with gemcitabine and cisplatin for patients with advanced non-small cell lung cancer improve efficacy. Int J Biomed Sci. 2012;8(1):28–35.
26. Liu T, Liu X, Li W. Tetrandrine, a Chinese plant-derived alkaloid, is a potential candidate for cancer chemotherapy. Oncotarget. 2016;7(26):40800–40815. doi:10.18632/oncotarget.8315
27. Wang TH, Wan JY, Gong X, Li HZ, Cheng Y. Tetrandrine enhances cytotoxicity of cisplatin in human drug-resistant esophageal squamous carcinoma cells by inhibition of multidrug resistance-associated protein 1. Oncol Rep. 2012;28(5):1681–1686. doi:10.3892/or.2012.1999
28. Bretones G, Delgado MD, Leon J. Myc and cell cycle control. Biochim Biophys Acta. 2015;1849(5):506–516. doi:10.1016/j.bbagrm.2014.03.013
29. Gartel AL, Radhakrishnan SK. Lost in transcription: p21 repression, mechanisms, and consequences. Cancer Res. 2005;65(10):3980–3985. doi:10.1158/0008-5472.CAN-04-3995
30. Zhang Y, Chen S, Wei C, et al. Dietary compound proanthocyanidins from Chinese bayberry (Myrica rubra Sieb. et Zucc.) leaves attenuate chemotherapy-resistant ovarian cancer stem cell traits via targeting the Wnt/beta-catenin signaling pathway and inducing G1 cell cycle arrest. Food Funct. 2018;9(1):525–533. doi:10.1039/C7FO01453H
31. Hou Y, Guo T, Wu C, He X. Effect of tetrandrine combined with epirubicin on the growth of human breast carcinoma multidrug resistance cell line. Yakugaku Zasshi. 2008;128(4):663–666. doi:10.1248/yakushi.128.663
32. Singh K, Dong Q, TimiriShanmugam PS, Koul S, Koul HK. Tetrandrine inhibits deregulated cell cycle in pancreatic cancer cells: differential regulation of p21(Cip1/Waf1), p27(Kip1) and cyclin D1. Cancer Lett. 2018;425:164–173. doi:10.1016/j.canlet.2018.03.042
33. Sanchez-Carranza JN, Diaz JF, Redondo-Horcajo M, et al. Gallic acid sensitizes paclitaxel-resistant human ovarian carcinoma cells through an increase in reactive oxygen species and subsequent downregulation of ERK activation. Oncol Rep. 2018;39(6):3007–3014. doi:10.3892/or.2018.6382
34. Nguyen VHL, Hough R, Bernaudo S, Peng C. Wnt/beta-catenin signalling in ovarian cancer: insights into its hyperactivation and function in tumorigenesis. J Ovarian Res. 2019;12(1):122. doi:10.1186/s13048-019-0596-z
35. Liang T, Ye X, Liu Y, et al. FAM46B inhibits cell proliferation and cell cycle progression in prostate cancer through ubiquitination of beta-catenin. Exp Mol Med. 2018;50(12):163. doi:10.1038/s12276-018-0184-0
36. Liao D, Zhang W, Gupta P, et al. Tetrandrine Interaction with ABCB1 reverses multidrug resistance in cancer cells through competition with anti-cancer drugs followed by downregulation of ABCB1 expression. Molecules. 2019;24(23):23. doi:10.3390/molecules24234383
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.