")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 11
Telomere length in blood cells is related to the chronicity, severity, and recurrence rate of schizophrenia
Authors Pawełczyk T, Szymanska B, Grancow M, Kotlicka-Antczak M, Pawełczyk A
Received 9 February 2015
Accepted for publication 25 March 2015
Published 22 June 2015 Volume 2015:11 Pages 1493—1503
DOI https://doi.org/10.2147/NDT.S82468
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Tomasz Pawelczyk,1 Bozena Szymanska,2 Marta Grancow-Grabka,3 Magdalena Kotlicka-Antczak,1 Agnieszka Pawelczyk1
1Department of Affective and Psychotic Disorders, Medical University of Lodz, Czechoslowacka, Lodz, Poland; 2Central Scientific Laboratory, Medical University of Lodz, Mazowiecka, Lodz, Poland; 3Central Teaching Hospital, Medical University of Lodz, Czechoslowacka, Lodz, Poland
Introduction: Telomere shortening is strongly associated with higher mortality rates and has been shown in a number of age-related diseases, such as cardiovascular disorders, diabetes mellitus, Alzheimer’s disease, and psychiatric disorders. Oxidative stress is known to induce DNA breaks and genome instability. Telomeric DNA rich in guanosine is particularly sensitive to such oxidative damages. Psychosis is associated with a disequilibrium between free radical production and antioxidative defense. Although telomere attrition has been demonstrated in schizophrenia, no relationship has been reported between telomere length and severity of schizophrenia.
Aim: The aim of the present study was to identify differences in telomere length in peripheral blood cells between patients with chronic schizophrenia (C-SCZ) and early schizophrenia (E-SCZ) and to identify any relationship between telomere length and disease chronicity and severity.
Methods: Relative average telomere lengths were determined using qPCR assay in patients with E-SCZ (n=42) and C-SCZ (n=44) hospitalized due to schizophrenia exacerbation. E-SCZ was diagnosed when less than 2 years had passed since the beginning of psychotic symptoms. The severity of symptoms was assessed using appropriate scales.
Results: The severity of schizophrenia symptoms, as well as the number of psychotic episodes and hospital admissions, correlated significantly with telomere length in univariate analyses. Regression analysis revealed that a model incorporating study group (E-SCZ or C-ECZ), sex, and age, as well as the combined number of documented psychotic episodes and hospital admissions, can significantly predict the length of telomeres in patients with schizophrenia, with over 50% of variance in telomere length explained by the model (adjusted R2=0.512).
Conclusion: The results of the current study indicate that the recurrence of psychotic symptoms as well as their intensity and chronicity may be correlated with telomere attrition, which is well known to contribute to the development of premature senescence and age-related diseases.
Keywords: telomere shortening, schizophrenia, exacerbation, aging
Introduction
Telomeres are nucleoprotein complexes that, in mammals, are composed of tandem repeat DNA sequences (TTAGGG)n that form a protective cap at the ends of eukaryotic chromosomes.1 During somatic cell division, DNA polymerase is not able to fully replicate the 3′ end of linear DNA, which results in progressive loss of telomeric repeats. Although on the one hand, this obligatory telomere loss provides a tumor suppressor mechanism that limits cell proliferation, it eventually results in replicative senescence and contributes to aging and cellular apoptosis.2 In 2009, The Nobel Prize in Physiology or Medicine was awarded to Elizabeth H Blackburn, Carol W Greider, and Jack W Szostak for the discovery of how chromosomes are protected by telomeres and the enzyme telomerase. This award highlighted the important role of telomerase and telomeres in genome stability, immortality, and aging.
It has been proposed that telomeres act as molecular clocks. They shorten with each cell division, and with age, due to the difficulty in end replication. Oxidative stress, chronic inflammation, and genotoxic agents can accelerate telomere attrition. Epel et al report that psychological stress – both perceived stress and chronicity of stress – is significantly associated with known determinants of cell senescence and longevity, ie, higher oxidative stress, lower telomerase activity, and shorter telomere length, in peripheral blood mononuclear cells from healthy premenopausal women.3 The telomeres of participants with the highest level of perceived stress were found to be shorter by the equivalent of at least one decade of additional aging compared to women reporting a low stress level. The authors conclude that increased oxidative stress was responsible for telomere attrition and lower telomerase activity. This relationship could provide an explanation for the common observation that psychological stress can lead to accelerated aging. These observations have led to telomere length emerging as a novel marker of biological age, which integrates the cumulative lifetime burden of genetic factors and environmental stressors independent of chronological age.4
Aging-related telomere shortening has been detected in cell and tissue types such as fibroblasts, leukocytes, vascular tissues, liver, and kidney tissue.5 Telomere attrition was observed in a number of age-related and degenerative diseases, such as atherosclerosis, diabetes mellitus, cancer, aging syndromes, vascular dementia, and Alzheimer’s disease.6–8 Accelerated telomere shortening and decreased telomerase activity has been reported in chronically stressed individuals,3 and accelerated telomere shortening has been reported in anxiety disorders,9 affective disorders,10,11 and schizophrenia.12
Schizophrenia is a neurodevelopmental and neurodegenerative disease whose pathophysiology is poorly understood. Psychotic symptoms commonly present in schizophrenia were found to be related to overproduction of reactive oxygen species (ROS) and impaired oxidative defense.13 Oxidative stress has also been shown to influence telomere length,14 which was found to be shorter in the blood leukocytes of schizophrenia patients in comparison with healthy controls. Kao et al found that lymphocyte telomere erosion in schizophrenia is rapid and that telomeres of patients in their mid- to late-30s were nearly 1,800 base pairs shorter than those of similarly aged controls: the controls would have to be at least 25 years older for the telomere to be of similar length.12 Associations between schizophrenia and telomere attrition have also been found elsewhere.15 However, other studies do not confirm this observation: two studies report no significant differences in telomere length between healthy controls and schizophrenia patients,16,17 and one large study found significantly longer telomeres in patients with schizophrenia in comparison with healthy controls.18
Previous studies have also assessed the relationship between the ability to achieve remission in schizophrenia and telomere length. It was shown that unremitting patients with chronic schizophrenia (C-SCZ) present with significantly shorter telomeres than either patients who achieve remission or healthy individuals matched by age and sex.19 However, another large study did not find any such relationship between telomere length and schizophrenia severity phenotypes.18
A recent study found a relationship between lifetime depression and leukocyte telomere shortening to be independent of the current severity of depressive symptoms.20 Telomere length of individuals, with or without current depressive symptoms, was shown to be significantly shorter than in healthy individuals without a history of unipolar depression. The authors did not find significant differences in telomere length between participants with a history of major depression presenting with or without the active symptoms of depression.
The differences in telomere length between C-SCZ and early schizophrenia (E-SCZ) patients have not yet been reported. Since oxidative stress is implicated in the pathophysiology of schizophrenia, the aim of the present study was to assess differences in telomere length between patients with C-SCZ and E-SCZ, and the relationship between telomere attrition and disease chronicity and severity.
Methods
Population
A total of 86 participants were enrolled in the study (female: n=35; 40.7%, aged 18–59 years). Participants were recruited from the inpatient psychiatric setting of the Department of Affective and Psychotic Disorders, Medical University of Lodz, Poland. All the recruited patients had been admitted to the psychiatric wards for exacerbation of schizophrenic symptoms. Eligible patients were diagnosed with schizophrenia according to ICD-10 criteria, which is an obligatory classification of mental disorders in Poland. The diagnosis was further confirmed using the Mini-International Neuropsychiatric Interview.21
A total of 163 patients were screened for eligibility, of whom, 55 were found ineligible based on the following exclusion criteria: being outside the age range of 18–60 years; diagnosed with mental retardation or severe organic brain syndrome; diagnosed with a serious or unstable medical condition; being pregnant or breastfeeding; undergoing treatment with electroconvulsive therapy; diagnosed with other comorbid psychiatric disorders or a mental disorder due to substance use. Among the 108 eligible patients, 22 (25.6%) refused to participate. These 22 eligible but unenrolled patients did not differ significantly from the 86 participating patients in terms of age, sex, or race/ethnic origin. The sample represents a nonrandom, convenience sample of consecutively hospitalized patients diagnosed with schizophrenia in a defined catchment area. The eligible participants were further categorized as E-SCZ or C-SCZ. The E-SCZ group was composed of 42 patients (aged 18–35 years), who were classified as E-SCZ using criteria introduced by Kahn et al22 with less than 2 years since the beginning of psychotic symptoms. The C-SCZ group comprised 44 schizophrenia patients (aged 18–60 years), who were considered as suffering C-SCZ when more than 2 years had passed from the emergence of psychotic symptoms, and at least two distinct psychotic episodes had been observed and documented in patients’ medical files.
Table 1 presents the demographic and socioeconomic data characterizing the study population as well as mean Positive and Negative Syndrome Scale (PANSS), Calgary Depression Scale for Schizophrenia (CDSS), and Global Assessment of Functioning (GAF) scores of the enrolled patients.
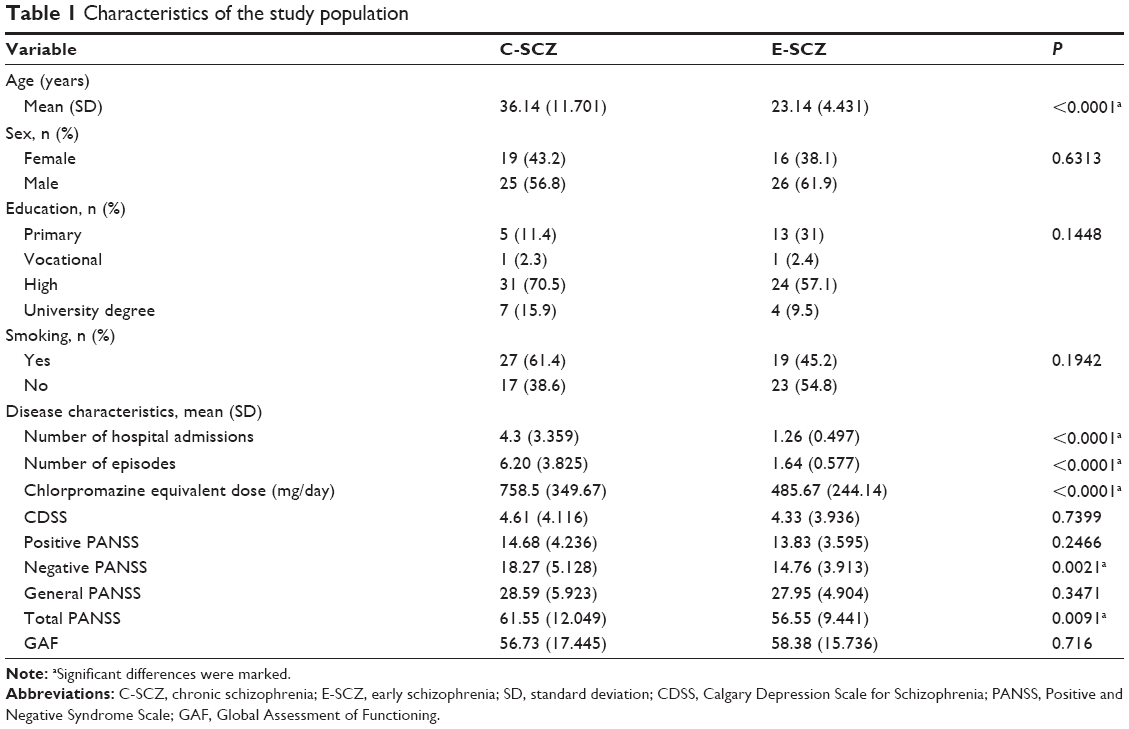 | Table 1 Characteristics of the study population |
Sample size calculation
Previous studies have shown that the clinical significance of the differences in telomere length (measured as T/S ratio) between schizophrenia patients and healthy controls is medium to large.12 Using the results obtained in previous studies, a power calculation reveals that 43 participants per study arm will result in 80% power to detect medium effect size (Cohen’s d=0.5) at a significance level of 0.05 using two-sided tests.
Assessments
Telomere length measurement
Genomic DNA samples were isolated from 200 μL of blood using the GeneMATRIX Quick Blood DNA Purification Kit (EURx Ltd, Gdansk, Poland) according to the manufacturer’s protocol. DNA was quantified using a Picodrop spectrophotometer (Picodrop Limited, Hinxton, UK), and all samples were diluted in Tris-EDTA (TE) buffer to a concentration of 3.5 ng/μL.
Relative average telomere lengths were determined using qPCR assay.23 This method amplifies the telomeric (T) DNA, and a single-copy (S) control gene (36B4) was used to normalize values. Cycle threshold values are converted to nanograms of DNA using a standard curve of serial dilutions. Using these values, a relative measure of telomere length is computed as a T/S ratio.
qPCR reactions were carried out in a 25 μL volume containing 5 μL DNA. The telomere reaction mixture consisted of 1× KAPA SYBR FAST qPCR Kit Master Mix, 2.5 mM of DTT, 270 nM of Tel-1 primer (GGTTTTT GAGGGTGAGGGTGAGGGTGAGGGTGAGGGT), and 900 nM of Tel-2 primer (TCCCGACTATCCCTATC CCTATCCCTATCCCTATCCCTA). The reaction proceeded for one cycle at 95°C for 5 minutes, followed by 18 cycles at 95°C for 15 seconds, and incubation at 54°C for 2 minutes. The 36B4, single-copy gene, reaction consisted of 1× KAPA SYBR FAST qPCR Kit Master Mix, 300 nM of 36B4U primer (CAGCAAGTGGGAAGGTGTAATCC), and 500 nM of 36B4D primer (CCCATTCTATCATCAACGGGTACAA). The 36B4 reaction proceeded for one cycle at 95°C for 5 minutes, followed by 30 cycles at 95°C for 15 seconds, and the mix was incubated at 58°C for 1 minute. All the samples for both the telomere and single-copy gene reactions were performed in triplicate on different 96-well plates in the same well positions. Reference DNA samples were diluted serially to produce five concentrations ranging from 0.625 ng/μL to 10 ng/μL, which were then distributed to the standard curve wells on each plate. All PCRs were performed on the 7900 HT Fast Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA) and Sequence Detection System version 2.0 software was then used to generate the standard curve for each plate and to determine the dilution factors of standards corresponding to the T and S amounts in each sample.
Chlorpromazine dose equivalents
Daily doses of antipsychotics used were converted into chlorpromazine equivalents using an equivalency table provided by Gardner et al.24 According to a recent systematic review prepared by Patel et al25 there is no universally accepted method of calculating antipsychotic dose equivalents. Comparison of methods for calculating antipsychotic dose equivalence have suggested that different methods yield different equivalencies, and the evidence is not sufficiently robust for any to be considered a gold standard method. Hence, the most recent and comprehensive consensus has been chosen for this study, which according to Patel et al25 appears to provide the most complete estimation of equivalencies of the four reviewed methods.
Clinical assessments
Clinical scales were used to assess several domains of symptoms and patient functioning. The severity of schizophrenia symptoms was measured using the PANSS,26 which is composed of 30 items with scores 1–7, with a total score ranging from 30 (indicating absence of psychopathology) to 208. PANSS is a reliable and valid scale commonly used to score symptom severity in schizophrenia.27
Depressive features were measured by the CDSS developed specifically for schizophrenia patients.28 It is an observer-rated scale that consists of nine items, each with four anchor points. The validity and reliability of the scale has been assessed in adults29 as well as in adolescents.30
Patient functioning status was measured by means of the GAF scale. The GAF is used to represent a clinician’s judgment of the overall level of psychosocial functioning of a patient.31 The GAF requires a clinician to make an overall judgment about the current psychological, social, and occupational functioning of a patient. In the Diagnostic and Statistical Manual of Mental Disorders 4th edition (DSM-IV), this rating was made on a scale from 1 to 100, with ratings of 1–10 indicating severe impairment and ratings of 91–100 indicating superior functioning.
Descriptive variables included socioeconomic and demographic characteristics, variables concerning psychiatric therapy, and history. Information concerning psychiatric history was extracted from the medical files of the patients.
The raters (TP and MG) undertook intensive training in the scoring methods used in the present study as part of the Programme of Recognition and Therapy (PORT): the first Polish program for people at high clinical risk of psychosis development, described in detail elsewhere.32 The raters achieved good reliability scores (coefficient of agreement >0.82) and were all within 20% of the standard scores.
All the participants were assessed as inpatients during the early recovery phase of the current episode over the week preceding discharge from hospital. Hence, all clinical assessments and psychopharmacology data represent the optimal improvement achieved during the current schizophrenia episode.
Statistical analysis
The distributions of the recorded clinical values were first assessed for normality by the Shapiro–Wilk test. Mean and standard deviations were used as measures of central tendency and dispersion of data. Due to a skewed distribution, telomere length expressed as T/S ratio was transformed using natural logarithm to permit comparisons of normally distributed values within each study group. Student’s t−test was used to evaluate the significance of differences in the distributions of continuous variables characterizing the groups. Comparisons on categorical variables were made using the chi-square test with or without Yates’ correction or Fisher’s exact test depending on the number of observations. Correlations were assessed using Pearson’s correlation coefficient followed by a suitable significance test. As expected, a negative correlation was found between telomere length and age (r=−0.472, P<0.0001). Since both age and sex33 can affect telomere length, the results were analyzed using a one-way analysis of covariance (ANCOVA) to control the effects of these two variables. Since telomere length significantly associated with age, the variables describing severity of symptoms (negative and total PANSS score), as well as the number of hospitalizations and episodes of disease, linear regression technique was used to control the confounding variables, to decrease redundancy of data, and to find variables with the highest relation to telomere length. Thus, the stepwise backward and forward linear regression method was used, which revealed that the same models best fitted to the data. Because of a high intercorrelation between the two variables: number of psychotic episodes and number of hospital admissions (Pearson’s r=0.96, P<0.0001) and low tolerance values (0.055 and 0.066), the information from the two variables was combined (addition), since it makes conceptual sense and can serve as a proxy measure of disease recurrence rate and severity. This approach is a recommended solution to avoid multicollinearity problems.34 No significant collinearity was present in the final adjusted model, with all pairwise correlation coefficients less than 0.6 and all tolerance values less than 1- R2. The assumption of linearity was checked by visual inspection of component plus residual plots. The normality assumption was checked by review of residual histograms and normal quantile–quantile plots. A significance level of 0.05 was used for all statistical tests, and two-tailed test probabilities were applied.
The study was carried out in accordance with the latest version of the Declaration of Helsinki. The study design was accepted by the Medical University of Łódź Ethics Committee. All the participants provided written informed consent before entering the study. The study, part of a larger project, was financed by the Polish Science National Center, grant No N N402 243435.
Results
The groups differed significantly according to age, number of hospital admissions, number of psychotic episodes, and negative and total PANSS scores. The groups did not differ according to sex, education, smoking status, or other dimensions of clinical scales. The differences in age and psychiatric history are the consequence of the criteria used to classify patients to the study groups. However, each of these variables provides additional information. Age is correlated with the number of psychotic episodes, but that last variable also provides information about the severity of the disease course. Likewise, the number of hospital admissions is correlated with the number of psychotic episodes, but the first variable also gives an insight into the severity of psychotic episodes. Patients also differed according to psychopathology (total and negative PANSS scores) assessed at the end of the hospitalization period. These scores represent possibly the best achievable improvement in the two groups of patients. It is well established that the improvement observed in patients with C-SCZ is lower than that in patients with a shorter duration of symptoms.
Univariate analyses
Univariate analyses did not reveal any significant associations between telomere length and either sex or smoking status (Student’s t-test, t=1.767, P=0.081 and t=0.99, P=0.321, respectively). Telomere length was significantly associated with age (Pearson’s r=−0.472, P<0.001). Significant negative correlation was found between telomere length (ln T/S ratio) and disease severity in the whole group of schizophrenia patients (n=86). Telomere length was found to significantly correlate with negative PANSS score (Pearson’s r=−0.318, P<0.01) and total PANSS (Pearson’s r=−0.238, P<0.05). No significant correlations were observed between telomere length and either positive (r=−0.143) or general PANSS scores (r=−0.097) (P>0.05). No significant correlations were found between telomere length and either patient functioning (GAF) or severity of depressive symptoms (CDSS) (Pearson’s r=−0.036, P=0.744 and r=−0.029, P=0.793, respectively). However, the association between telomere length and both the number of hospital admissions and the number of psychotic episodes was significant (Pearson’s r=−0.466, P<0.001 and r=−0.594, P<0.001, respectively). No significant correlation was observed between telomere length and the equivalent dose of chlorpromazine (Pearson’s r=−0.155, P=0.155). Subgroup analyses carried out in E-SCZ and C-SCZ groups revealed correlation coefficients that did not reach significance level but a trend toward negative correlation was observed between negative PANSS score and telomere length in the C-SCZ group (Pearson’s r=−0.196, P=0.065).
ANCOVA
An ANCOVA was used to assess whether telomere length (expressed as ln T/S ratio) is related to the chronicity of schizophrenia after controlling for the influence of age and sex. Results indicate that after controlling for the possible confounder variables, a significant difference was observed in telomere length between groups of patients with E-SCZ and C-SCZ (F[1,82] =47.08, P<0.001). Patients with shorter disease durations had significantly longer telomeres than patients with C-SCZ. No main effect of age or sex was observed within this analysis. Table 2 presents the adjusted and unadjusted group means and variability of telomere length (ln T/S ratio) after controlling for the influence of age and sex and the results of ANCOVA for telomere length as a function of group, age, and sex.
 | Table 2 Adjusted and unadjusted group means and variability of telomere length (ln T/S ratio) after controlling for the influence of age and sex and the results of ANCOVA for telomere length as a function of group, age, and sex |
Linear regression
Multiple stepwise backward and forward regression procedures were conducted to determine the best linear combination of the disease severity (negative and total PANSS score), chronicity of disease, number of psychotic episodes and hospitalizations, age, and sex for predicting telomere length in patients diagnosed with schizophrenia. All the above mentioned continuous variables correlated significantly with telomere length (described in “Univariate analyses” section). The cutoff probabilities (F) in stepwise regression procedures for inclusion and exclusion of variables were assumed as 0.05 and 0.1, respectively. Both backward and forward stepwise regression procedures revealed the same combination of variables, which significantly predicted telomere length, F(4,85) =23.263, P<0.0001, with three variables significantly contributing to the prediction. Age, although not significant, is known to be associated with telomere length and so was included in the model to control its effects. The beta weights, presented in Table 3, suggest that longer duration of the disease (belonging to C-SCZ group), higher combined number of psychotic episodes and hospital admissions, and being female are related to shorter telomere length. The adjusted R2 value was 0.512, which indicates that over 50% of the variance in telomere length was explained by the model. According to Cohen,35 this is a large effect.
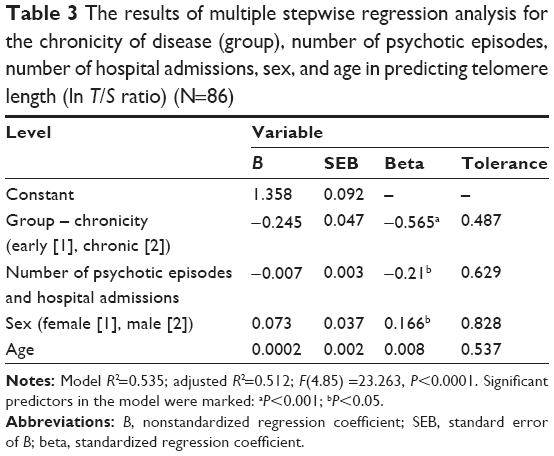 | Table 3 The results of multiple stepwise regression analysis for the chronicity of disease (group), number of psychotic episodes, number of hospital admissions, sex, and age in predicting telomere length (ln T/S ratio) (N=86) |
Discussion
The results of the present study suggest that telomere length of peripheral blood cells in patients with schizophrenia is related to the chronicity and severity of the disease course. Severity of psychopathology measured by the total and negative PANSS scores was correlated negatively with telomere length. It was shown that the number of psychotic episodes and the number of the documented hospital admissions were also significantly associated with telomere length. ANCOVA analysis showed that telomere erosion in schizophrenia is significantly higher in patients with C-SCZ than with E-SCZ even after controlling for the effects of the confounding variables: age and sex. The magnitude of the effect (η2 =0.365) is large according to the rule of thumb provided by Cohen.35 Moreover, this was further confirmed in regression analysis, which showed that the model composed of the study group (E-SCZ or C-SCZ), sex, age, and the number of the documented psychotic episodes and hospital admissions can significantly predict the length of telomeres in patients with schizophrenia with over 50% of variance in telomere length explained by the model (adjusted R2=0.512).
Among the strengths of the current study is that it is the first to compare the telomere length of blood cells in patients with C-SCZ and E-SCZ and to assess the relationship between both the severity and chronicity of the disease with telomere length. However, several limitations should be considered in the interpretation of our results. First, the association reported in this study is observational, and therefore, no definitive conclusions can be made regarding causality. Although adjustments were made for potential confounding variables, the possibility of residual confounding by measured or unmeasured covariates cannot be excluded. For example, the influence of parental age, physical activity, systemic inflammation, obesity, and red meat consumption are thought to cumulatively determine telomere length through adult life. These potential confounders were not adjusted for in the analyses. Second, our measurements were restricted to telomere length in blood cells and do not necessarily reflect telomere trajectory in other cell compartments such as nervous or other tissues. Third, our study sample comprises hospitalized patients with psychotic exacerbation, which may represent a population especially susceptible to the effects of psychosis on telomere length; hence, the findings cannot be generalized to other populations of patients, and further studies are required to verify our findings in other populations of patients. Fourth, the method of measuring telomere length based on quantitative RT-PCR developed by Cawthon has been criticized for being difficult to optimize and extremely sensitive to technical errors, resulting in high variability within and between samples.36 However, despite the comparative inaccuracy of the technique, general trends between population groups can be made as long as errors are assumed to be similar between groups. The strength of this method is its practical simplicity in obtaining, analyzing, and interpreting qPCR data, making it one of the most common methods for studying telomere dynamics. There are four main methods of measuring telomere length, and each approach has its strengths and weaknesses.37 The optimal method of measuring telomere length is still debated.
Finally, the sample size is not large for genetic studies. However, comparable sample sizes were used in many other studies of telomere length in patients with schizophrenia.12,15–17,19,38,39 Small sample size may contribute to conflicting results of telomere length, which was observed in patients with schizophrenia. Thus, the results of the present study should be regarded as preliminary and further studies are needed using larger samples.
Few studies have assessed the relationship between schizophrenia symptom severity and telomere length. Yu et al compared telomere length in C-SCZ patients who responded well and poorly to antipsychotic therapy.38 The groups differed significantly according to symptom severity, with poor responders presenting higher Brief Psychiatric Rating Scale scores than good responders. Telomere length (expressed as telomere restriction fragments) was found to be significantly higher in less severely affected patients (good responders) in comparison to patients with worse outcome (poor responders). Moreover, telomere length was significantly shorter in poor responders than in control subjects. The authors did not find significant differences in telomere length between schizophrenia patients with better outcome (good responders) and control subjects. Interestingly, the authors did not observe any negative correlation between age and telomere length in either patient group, with the slopes of regression lines in fact being positive. A well-established negative correlation between age and telomere length was observed only in the control group. An inverse relationship between telomere length and poor outcome was recently observed by Kota et al in patients with schizophrenia,19 who showed that unremitting schizophrenia patients present with significantly lower relative telomere length than patients who achieve remission of psychotic episode and individuals without neuropsychiatric illness. The authors concluded that the results of their study may indicate that schizophrenia shares biological pathways with other neurodegenerative disorders that are also characterized by increased cellular senescence. Lower telomerase levels were also observed in schizophrenia patients.40
The findings of the present study are generally in line with those of previous studies. However, a few differences between the studies warrant attention. Different methods were used to assess psychopathology in the study by Yu et al38 and the current one. The Brief Psychiatric Rating Scale method used by Yu et al concentrates mainly on positive symptoms, whereas the PANSS used in the present study covers different symptomatic domains, including positive and negative symptoms as well as general psychopathology. In addition, while Yu et al assessed C-SCZ patients, the present study enrolled individuals with both early and chronic disease. The results of the current study are also in line with those of Kota et al.19
However, the relationship between disease severity and telomere length is not consistent. In contrast with the present findings, Nieratschker et al in a large sample of schizophrenia patients (n=539), did not find any significant association between telomere length and the presence of any schizophrenia severity subphenotypes.18 The authors suggest that their findings could be biased by antipsychotic use, excess mortality associated with schizophrenia, or might be explained by an underlying association with endophenotypes, which acts beyond the currently available diagnostic boundaries for schizophrenia.18 The last suggestion may be supported by the results of the study mentioned earlier, which was carried out by Yu et al38 in which only C-SCZ patients who responded poorly to antipsychotic therapy presented with shorter telomere length than healthy subjects. In addition, although the study population enrolled by Nieratschker et al is not characterized precisely enough to allow comparisons with patients enrolled in the present study, generally speaking, younger patients were enrolled in the present study: the mean age of both groups in the present study being 29 years with a range of 18–59 years, compared to a mean age 37 years and range of 17–80 years for Nieratschker et al. This difference together with the possible bias suggested by the authors may be responsible for the observed differences between the results of the current study and the study by Nieratschker et al.
The possible explanation of the observed relationship between the severity and chronicity of psychotic episodes and reduced telomere length may be found in the paradigm of oxidative stress. It is understood as a disequilibrium between the increased production of ROS and decreased oxidative defense represented mainly by the activity of free radical scavenging enzymes, ie, superoxide dismutase, catalase, and glutathione.41 Increased levels of oxidative damage and decreased oxidative defense were repeatedly observed in schizophrenia.13 It was shown that psychosis is connected with neurotoxic effects induced by ROS.42 Recent studies show that oxidative stress might serve as a potential biomarker in the etiopathophysiology and clinical course of schizophrenia.13 Telomeric G triplets are especially sensitive to cleavage by oxidative damage, which leads to single-strand breaks and the loss of distal fragments of telomeric DNA following replication.14,33,43
Accelerated telomere shortening may have important consequences and may shed light on the excess mortality observed in patients with schizophrenia. Telomere length is regarded as a marker of biological aging. Shortening of telomeres triggers multiple aging-related processes, such as cell growth arrest, apoptosis, and decreased capacity in response to stress in highly replicative cells.33 Excess mortality, expressed as standardized mortality rate, has repeatedly and consistently been found in persons with schizophrenia.44–47 In most countries, standardized mortality rate of schizophrenia patients is about 2.5, which leads to shortened life expectancy of about 15 years to nearly 20 years compared with general population.48,49 The main contributor of the shortened life expectancy in schizophrenia is cardiovascular disease (CVD), diabetes, and their complications, such as cardiovascular accidents.50–53
Patients with schizophrenia show higher incidence of metabolic abnormalities characteristic of CVD, diabetes, and obesity even before antipsychotic therapy initiation.54,55 Recent studies suggest that schizophrenia and CVD risk factors may share pathophysiology and genetics.54,55 Obesity, insulin resistance, dyslipidemia, and hypertension are observed more often in antipsychotic-naïve first-episode schizophrenia (FES) patients compared with age-matched healthy controls.53 Recent molecular studies have shown the abnormalities in the glucose metabolism and insulin signaling pathways in nonmedicated subgroups of people with schizophrenia indicating a shared genetic vulnerability between type 2 diabetes and schizophrenia.56 FES patients also show increased cortisol levels due to increased environmental stress, which further increases the risk of glucose and lipid abnormalities.57 Not only CVD, diabetes mellitus, and schizophrenia share plenty of environmental and genetic risk factors as well as pathophysiology but also telomere pathology. Telomere erosion is observed in schizophrenia,12,19,38 coronary artery disease,58 insulin resistance,59 diabetes mellitus,60 and many other degenerative disorders, which are related to accelerated senescence.61,62
Many factors are related to telomere erosion, such as sex, age, obesity, systemic inflammation, lack of physical activity, plasma leptin and irisin levels, total and abdominal fat percentage, and oxidative stress.63–65 Many of the above risk factors are the same as for CVD and diabetes mellitus. Not only schizophrenia and CVD share pathophysiology but also recurrent psychotic episodes may lead to reduced telomere length, which further increases the risk of developing CVD, diabetes, their complications, and other age-related conditions.
It was shown that short telomeres trigger cell senescence, and senescent cells accumulate in aged tissues. These cells produce factors that affect not only the local tissue but also the whole organism. Senescence-associated secretory phenotype includes secretion of many proinflammatory cytokines, chemokines, growth factors, and proteases that have the potential to cause or exacerbate age-related pathology, both degenerative and hyperplastic.66 Senescence in peripheral tissues has recently been linked to a number of age-related pathologies, including atherosclerosis and diabetes mellitus.67 Recent data generated by several laboratories suggest that both aging and age-related neurodegenerative diseases are accompanied by an increase in senescence-associated secretory phenotype-expressing senescent cells of nonneuronal origin in the brain.68 These complex alterations triggered by telomere attrition may explain association between psychosis severity and chronicity, oxidative stress, age-related diseases, neurodegenerative diseases, and reduced life expectancy in schizophrenia.
The relationship between telomere attrition and schizophrenia severity and chronicity observed in the current study may suffer from different kinds of bias typical for observational studies, since it is not possible to foresee and control all relevant confounding factors. Thus, longitudinal cohort studies of FES individuals are needed to further explain the relationship between psychopathology and telomere length in schizophrenia. The results of the present study, together with those of a previous report69 of reduced telomere attrition in patients diagnosed with mild cognitive impairment who were supplemented with either n-3 polyunsaturated fatty acids (PUFA), emphasize the need to assess the influence of PUFA supplementation on telomere shortening and telomerase activity in schizophrenia patients. Moreover, an understanding of the relationship between dietary habits in patients with schizophrenia, in particular n-3 and n-6 PUFA consumption and telomere length, would be a valuable influence on dietary recommendations in schizophrenia and reduce the burden of disease, premature aging, and excess mortality in schizophrenia patients.
The connection between chronicity and severity of schizophrenia and telomere length, as well as the possible consequences, premature cellular senescence, and increased cardiovascular mortality risk, reinforce the need for effective therapy of schizophrenia. It is well documented that achieving partial remission with the presence of residual psychotic symptoms may increase the risk of schizophrenia relapse. The results of the current preliminary study indicate that the recurrence of psychotic symptoms, as well as their intensity and chronicity, may be related to telomere attrition of cells, which activates degenerative processes, cellular senescence, or apoptosis. These may, in turn, lead to premature aging, the development of age-related diseases and their complications, leading to shortened life expectancy in schizophrenia.
Acknowledgments
This paper was supported by grant no N N402 243435 obtained from the Polish Science National Center. The authors would like to express their deep gratitude for Professor Jolata Rabe-Jabłońska, the former head of the department, who was deeply involved in the preparation of the study and unfortunately died in May 2014.
Disclosure
The authors report no conflicts of interest in this work.
References
Blackburn EH. Switching and signaling at the telomere. Cell. 2001; 106(6):661–673. | ||
Blackburn EH. Telomeres. Trends Biochem Sci. 1991;16(10): 378–381. | ||
Epel ES, Blackburn EH, Lin J, et al. Accelerated telomere shortening in response to life stress. Proc Natl Acad Sci U S A. 2004;101(49): 17312–17315. | ||
Olovnikov AM. Telomeres, telomerase, and aging: origin of the theory. Exp Gerontol. 1996;31(4):443–448. | ||
Zhao Z, Pan X, Liu L, Liu N. Telomere length maintenance, shortening, and lengthening. J Cell Physiol. 2014;229(10):1323–1329. | ||
Fyhrquist F, Saijonmaa O, Strandberg T. The roles of senescence and telomere shortening in cardiovascular disease. Nat Rev Cardiol. 2013; 10(5):274–283. | ||
Thomas P, O’Callaghan NJ, Fenech M. Telomere length in white blood cells, buccal cells and brain tissue and its variation with ageing and Alzheimer’s disease. Mech Ageing Dev. 2008;129(4):183–190. | ||
Zekry D, Herrmann FR, Irminger-Finger I, et al. Telomere length and ApoE polymorphism in mild cognitive impairment, degenerative and vascular dementia. J Neurol Sci. 2010;299(1–2):108–111. | ||
Kananen L, Surakka I, Pirkola S, et al. Childhood adversities are associated with shorter telomere length at adult age both in individuals with an anxiety disorder and controls. PLoS One. 2010;5(5):e10826. | ||
Simon NM, Smoller JW, McNamara KL, et al. Telomere shortening and mood disorders: preliminary support for a chronic stress model of accelerated aging. Biol Psychiatry. 2006;60(5):432–435. | ||
Garcia-Rizo C, Fernandez-Egea E, Miller BJ, et al. Abnormal glucose tolerance, white blood cell count, and telomere length in newly diagnosed, antidepressant-naive patients with depression. Brain Behav Immun. 2013;28:49–53. | ||
Kao H, Cawthon RM, Delisi LE, et al. Rapid telomere erosion in schizophrenia. Mol Psychiatry. 2008;13(2):118–119. | ||
Flatow J, Buckley P, Miller BJ. Meta-analysis of oxidative stress in schizophrenia. Biol Psychiatry. 2013;74(6):400–409. | ||
von Zglinicki T. Role of oxidative stress in telomere length regulation and replicative senescence. Ann N Y Acad Sci. 2000;908:99–110. | ||
Fernandez-Egea E, Bernardo M, Heaphy CM, et al. Telomere length and pulse pressure in newly diagnosed, antipsychotic-naive patients with nonaffective psychosis. Schizophr Bull. 2009;35(2):437–442. | ||
Malaspina D, Dracxler R, Walsh-Messinger J, et al. Telomere length, family history, and paternal age in schizophrenia. Mol Genet Genomic Med. 2014;2(4):326–331. | ||
Mansour H, Chowdari K, Fathi W, et al. Does telomere length mediate associations between inbreeding and increased risk for bipolar I disorder and schizophrenia? Psychiatry Res. 2011;188(1):129–132. | ||
Nieratschker V, Lahtinen J, Meier S, et al. Longer telomere length in patients with schizophrenia. Schizophr Res. 2013;149(1–3):116–120. | ||
Kota LN, Purushottam M, Moily NS, Jain S. Shortened telomere in unremitted schizophrenia. Psychiatry Clin Neurosci. 2015;69(5):292–297. | ||
Karabatsiakis A, Kolassa I, Kolassa S, Rudolph KL, Dietrich DE. Telomere shortening in leukocyte subpopulations in depression. BMC Psychiatry. 2014;14:192. | ||
Sheehan DV, Lecrubier Y, Sheehan KH, et al. The Mini-International Neuropsychiatric Interview (M.I.N.I.): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J Clin Psychiatry. 1998;59(suppl 20):22–33. [quiz 34–57]. | ||
Kahn RS, Fleischhacker WW, Boter H, et al; EUFEST Study Group. Effectiveness of antipsychotic drugs in first-episode schizophrenia and schizophreniform disorder: an open randomised clinical trial. Lancet. 2008;371(9618):1085–1097. | ||
Cawthon RM. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002;30(10):e47. | ||
Gardner DM, Murphy AL, O’Donnell H, Centorrino F, Baldessarini RJ. International consensus study of antipsychotic dosing. Am J Psychiatry. 2010;167(6):686–693. | ||
Patel MX, Arista IA, Taylor M, Barnes TR. How to compare doses of different antipsychotics: a systematic review of methods. Schizophr Res. 2013;149(1–3):141–148. | ||
Kay SR, Fiszbein A, Opler LA. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr Bull. 1987;13(2):261–276. | ||
Kay SR, Opler LA, Lindenmayer JP. Reliability and validity of the positive and negative syndrome scale for schizophrenics. Psychiatry Res. 1988;23(1):99–110. | ||
Addington D, Addington J, Maticka-Tyndale E. Assessing depression in schizophrenia: the Calgary Depression Scale. Br J Psychiatry Suppl. 1993;22:39–44. | ||
Addington D, Addington J, Maticka-Tyndale E, Joyce J. Reliability and validity of a depression rating scale for schizophrenics. Schizophr Res. 1992;6(3):201–208. | ||
Addington J, Shah H, Liu L, Addington D. Reliability and validity of the Calgary Depression Scale for Schizophrenia (CDSS) in youth at clinical high risk for psychosis. Schizophr Res. 2014;153(1–3):64–67. | ||
Jones SH, Thornicroft G, Coffey M, Dunn G. A brief mental health outcome scale-reliability and validity of the Global Assessment of Functioning (GAF). Br J Psychiatry. 1995;166(5):654–659. | ||
Kotlicka-Antczak M, Pawelczyk T, Rabe-Jablonska J, Pawelczyk A. PORT (Programme of Recognition and Therapy): the first Polish recognition and treatment programme for patients with an at-risk mental state. Early Interv Psychiatry. Epub 2014 Apr 11. | ||
von Zglinicki T, Martin-Ruiz CM. Telomeres as biomarkers for ageing and age-related diseases. Curr Mol Med. 2005;5(2):197–203. | ||
Leech NL, Barrett KC, Morgan GA. SPSS for Intermediate Statistics: Use and Interpretation. 2 ed. Mahwah, NJ: Lawrence Erlbaum; 2005. | ||
Cohen J. Statistical Power Analysis for the Behavioral Sciences. 2 ed. Hillsdale, NJ: L. Erlbaum Associates; 1988. | ||
Aviv A. Commentary: raising the bar on telomere epidemiology. Int J Epidemiol. 2009;38(6):1735–1736. | ||
Aubert G, Hills M, Lansdorp PM. Telomere length measurement-caveats and a critical assessment of the available technologies and tools. Mutat Res. 2012;730(1–2):59–67. | ||
Yu W, Chang H, Lin C, Cho C. Short telomeres in patients with chronic schizophrenia who show a poor response to treatment. J Psychiatry Neurosci. 2008;33(3):244–247. | ||
Zhang D, Cheng L, Craig DW, Redman M, Liu C. Cerebellar telomere length and psychiatric disorders. Behav Genet. 2010;40(2):250–254. | ||
Porton B, Delisi LE, Bertisch HC, et al. Telomerase levels in schizophrenia: a preliminary study. Schizophr Res. 2008;106(2–3):242–247. | ||
Kawanishi S, Oikawa S. Mechanism of telomere shortening by oxidative stress. Ann N Y Acad Sci. 2004;1019:278–284. | ||
Schiavone S, Colaianna M, Curtis L. Impact of early life stress on the pathogenesis of mental disorders: relation to brain oxidative stress. Curr Pharm Des. 2015;21(11):1404–1412. | ||
Coluzzi E, Colamartino M, Cozzi R, et al. Oxidative stress induces persistent telomeric DNA damage responsible for nuclear morphology change in Mammalian cells. PLoS One. 2014;9(10):e110963. | ||
Tsuang MT, Woolson RF. Excess mortality in schizophrenia and affective disorders. Do suicides and accidental deaths solely account for this excess? Arch Gen Psychiatry. 1978;35(10):1181–1185. | ||
Harris EC, Barraclough B. Excess mortality of mental disorder. Br J Psychiatry. 1998;173:11–53. | ||
Osby U, Correia N, Brandt L, Ekbom A, Sparen P. Mortality and causes of death in schizophrenia in Stockholm county, Sweden. Schizophr Res. 2000;45(1–2):21–28. | ||
Saha S, Chant D, McGrath J. A systematic review of mortality in schizophrenia: is the differential mortality gap worsening over time? Arch Gen Psychiatry. 2007;64(10):1123–1131. | ||
Laursen TM. Life expectancy among persons with schizophrenia or bipolar affective disorder. Schizophr Res. 2011;131(1–3):101–104. | ||
Almeida OP, Hankey GJ, Yeap BB, Golledge J, Norman PE, Flicker L. Mortality among people with severe mental disorders who reach old age: a longitudinal study of a community-representative sample of 37892 men. PLoS One. 2014;9(10):e111882. | ||
Crump C, Winkleby MA, Sundquist K, Sundquist J. Comorbidities and mortality in persons with schizophrenia: a Swedish national cohort study. Am J Psychiatry. 2013;170(3):324–333. | ||
Hennekens CH, Hennekens AR, Hollar D, Casey DE. Schizophrenia and increased risks of cardiovascular disease. Am Heart J. 2005;150(6):1115–1121. | ||
Laursen TM, Nordentoft M, Mortensen PB. Excess early mortality in schizophrenia. Annu Rev Clin Psychol. 2014;10:425–448. | ||
Ringen PA, Engh JA, Birkenaes AB, Dieset I, Andreassen OA. Increased mortality in schizophrenia due to cardiovascular disease – a non-systematic review of epidemiology, possible causes, and interventions. Front Psychiatry. 2014;5:137. | ||
Chen S, Broqueres-You D, Yang G, et al. Relationship between insulin resistance, dyslipidaemia and positive symptom in Chinese antipsychotic-naive first-episode patients with schizophrenia. Psychiatry Res. 2013;210(3):825–829. | ||
Mitchell AJ, Vancampfort D, de Herdt A, Yu W, de Hert M. Is the prevalence of metabolic syndrome and metabolic abnormalities increased in early schizophrenia? A comparative meta-analysis of first episode, untreated and treated patients. Schizophr Bull. 2013;39(2):295–305. | ||
Harris LW, Guest PC, Wayland MT, et al. Schizophrenia: metabolic aspects of aetiology, diagnosis and future treatment strategies. Psychoneuroendocrinology. 2013;38(6):752–766. | ||
Bradley AJ, Dinan TG. A systematic review of hypothalamic-pituitary-adrenal axis function in schizophrenia: implications for mortality. J Psychopharmacol. 2010;24(4 suppl):91–118. | ||
Nilsson PM, Tufvesson H, Leosdottir M, Melander O. Telomeres and cardiovascular disease risk: an update 2013. Transl Res. 2013;162(6):371–380. | ||
Demissie S, Levy D, Benjamin EJ, et al. Insulin resistance, oxidative stress, hypertension, and leukocyte telomere length in men from the Framingham Heart Study. Aging Cell. 2006;5(4):325–330. | ||
Jeanclos E, Krolewski A, Skurnick J, et al. Shortened telomere length in white blood cells of patients with IDDM. Diabetes. 1998;47(3): 482–486. | ||
Hochstrasser T, Marksteiner J, Humpel C. Telomere length is age-dependent and reduced in monocytes of Alzheimer patients. Exp Gerontol. 2012;47(2):160–163. | ||
Schurks M, Buring J, Dushkes R, Gaziano JM, Zee RYL, Kurth T. Telomere length and Parkinson’s disease in men: a nested case-control study. Eur J Neurol. 2014;21(1):93–99. | ||
Serra V, Grune T, Sitte N, Saretzki G, von Zglinicki T. Telomere length as a marker of oxidative stress in primary human fibroblast cultures. Ann N Y Acad Sci. 2000;908:327–330. | ||
Rana KS, Arif M, Hill EJ, et al. Plasma irisin levels predict telomere length in healthy adults. Age (Dordr). 2014;36(2):995–1001. | ||
Gladych M, Wojtyla A, Rubis B. Human telomerase expression regulation. Biochem Cell Biol. 2011;89(4):359–376. | ||
Zhu Y, Armstrong JL, Tchkonia T, Kirkland JL. Cellular senescence and the senescent secretory phenotype in age-related chronic diseases. Curr Opin Clin Nutr Metab Care. 2014;17(4):324–328. | ||
Sikora E, Bielak-Zmijewska A, Mosieniak G. Cellular senescence in ageing, age-related disease and longevity. Curr Vasc Pharmacol. 2014; 12(5):698–706. | ||
Chinta SJ, Woods G, Rane A, Demaria M, Campisi J, Andersen JK. Cellular senescence and the aging brain. Exp Gerontol. Epub 2014 Oct 1. | ||
O’Callaghan N, Parletta N, Milte CM, Benassi-Evans B, Fenech M, Howe PR. Telomere shortening in elderly individuals with mild cognitive impairment may be attenuated with omega-3 fatty acid supplementation: a randomized controlled pilot study. Nutrition. 2014;30(4):489–491. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.