Back to Journals » Neuropsychiatric Disease and Treatment » Volume 22
Targeting Programmed Cell Death in Depression: A Review of Traditional Chinese Medicine Interventions Based on Preclinical Evidence
Authors Jia H, Wu M ![]() , Teng L, Song W, Li X, Wang B, Huang Y, Kang X, Zhu L
, Teng L, Song W, Li X, Wang B, Huang Y, Kang X, Zhu L ![]()
Received 23 August 2025
Accepted for publication 22 November 2025
Published 14 January 2026 Volume 2026:22 562047
DOI https://doi.org/10.2147/NDT.S562047
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Taro Kishi
Huanhuan Jia,1 Minmin Wu,1 Lili Teng,1 Wenjing Song,1 Xinyue Li,1 Binhan Wang,1 Ying Huang,1 Xingyu Kang,1 Luwen Zhu2
1Graduate School, Heilongjiang University of Chinese Medicine, Harbin, People’s Republic of China; 2Department of Rehabilitation, The Second Affiliated Hospital of Heilongjiang University of Chinese Medicine, Harbin, People’s Republic of China
Correspondence: Luwen Zhu, Department of Rehabilitation, The Second Affiliated Hospital of Heilongjiang University of Chinese Medicine, Harbin, People’s Republic of China, Email [email protected]
Abstract: Depression is a globally prevalent, highly disabling mental disorder, and its etiology, pathological mechanisms, and treatment approaches have long been a focus of intensive research. Programmed cell death (PCD) refers to a process in which cells actively undergo death to maintain internal homeostasis via specific signals or stimuli. Modern medical studies have demonstrated that the dysregulation of PCD modalities, including apoptosis, pyroptosis, autophagy, ferroptosis, necroptosis, and cuproptosis, is closely associated with the onset and progression of depression. Elucidating the molecular mechanisms of PCD in depression could provide critical insights for an in-depth understanding, prevention, and treatment of this disorder. In recent years, extensive research has been conducted in the field of traditional Chinese medicine (TCM) that targets PCD for depression treatment, yielding significant phased achievements. Therefore, this narrative review, covering literature primarily from the past decade, synthesizes preclinical evidence on six PCD modalities in depression. It further explores the potential mechanisms and current status of TCM interventions that target these PCD pathways, and discusses the crosstalk between them. By emphasizing mechanistic insights, this work aims to provide a theoretical foundation and guidance for future clinical prevention, treatment, and research on depression.
Keywords: depression, programmed cell death, traditional Chinese medicine
Introduction
Depression is a prevalent and disabling mental disorder characterized by persistent low mood, accompanied by a range of heterogeneous symptoms such as anhedonia, feelings of worthlessness, decreased appetite, fatigue, sleep disturbances, impairments in cognitive functions such as attention and memory, and even suicidal tendencies.1 Epidemiological data indicate that depression affects nearly 280 million people worldwide, accounting for 3.8% of the global population, and is projected to become one of the primary burdens of global disability in the future.1 This burden is particularly pronounced in the Asian region. The 2019 Global Burden of Disease Study revealed that depression is the leading cause of mental disorders in Asia, accounting for nearly half of the global cases and demonstrating a continuous upward trend.2 Although the age-standardized incidence rate of depression in China is lower than the global average, the ratio of years lived with disability to incidence is significantly elevated. This indicates that patients face a higher severity of disability and more substantial functional impairment due to the disease.3 The etiology and pathogenesis of depression are extremely complex and involve multiple domains, such as biochemistry, genetics, psychology, and environmental factors. These factors interact to collectively generate a series of pathophysiological mechanism hypotheses, including the monoaminergic neurotransmitter hypothesis, the hypothalamic‒pituitary‒adrenal axis hypothesis, the neuroplasticity hypothesis, the gut‒brain axis hypothesis, the liver‒brain axis hypothesis, and the inflammation and oxidative stress hypothesis.4 Importantly, these pathological processes not only induce the death of neurons and neuroglial cells, but the cell death they trigger further amplifies pathological processes such as neuroinflammation and oxidative stress, forming a vicious cycle that drives disease progression. Cell death plays a crucial role in maintaining physiological homeostasis and regulating the occurrence and progression of diseases. However, abnormal cell death impairs normal cellular communication and activity, and promotes disease progression. Based on its regulatory mechanisms and morphological characteristics, cell death is classified into non-programmed cell death and programmed cell death (PCD). Non-programmed cell death, also referred to as necrosis, is a passive and accidental cellular death process triggered by uncontrollable external factors. It is characterized by cellular swelling, membrane rupture, and the release of intracellular contents, which in turn induce inflammation and tissue damage. Owing to the absence of specific regulatory mechanisms, it is difficult to identify a therapeutic target.5 PCD refers to an active and ordered process of cell death governed by specific genetic programs and signaling pathways. It serves as the primary mechanism by which an organism eliminates damaged, dysfunctional, or potentially harmful cells in various ways.6 Current studies have definitively demonstrated that PCD is involved in the pathological process of depression. For instance, research in animal models of depression has shown significantly elevated levels of pro-apoptotic factors BCL-2–associated X protein (BAX) and cysteine-dependent aspartate-specific protease-3 (Caspase-3),7,8 activation of the NOD-like receptor family pyrin domain–containing 3 (NLRP3) inflammasome,9 and accumulation of iron ions and lipid peroxides.10 Furthermore, relevant studies have also revealed that depression is accompanied by abnormalities in other PCD types, including autophagy,11,12 cuproptosis,13 and necroptosis.14 These PCD types not only function independently but may also jointly regulate their occurrence and development through interactions. Therefore, investigating the regulatory pathways of PCD in depression is conducive to elucidating its pathogenesis and providing a basis for the development of new therapeutic strategies.
In the treatment of depression, current approaches still focus primarily on intervening in the levels of central monoaminergic neurotransmitters. Commonly used medications include monoamine oxidase inhibitors, tricyclic antidepressants, selective serotonin reuptake inhibitors, serotonin-norepinephrine reuptake inhibitors and atypical antidepressants.15 Despite demonstrating symptom alleviation in some patients, these medications present significant limitations: substantial interindividual variability in treatment response, suboptimal efficacy rates, considerable adverse effects, high relapse incidence, poor medication adherence, and severe withdrawal syndrome.16 Furthermore, depression is frequently comorbid with chronic diseases or other mood disorders, such as pain, epilepsy, cardio-cerebrovascular diseases, gastrointestinal disorders, anxiety, and posttraumatic stress disorder, which may contribute to suboptimal responses to antidepressant treatment. Traditional Chinese medicine (TCM) has long been used to treat depression and remains widely applied in clinical practice to this day, particularly in Asian regions. In TCM theory, depression is categorized under the concept of “Yu Zheng”. There are internal and external causes for depression, externally because of emotional factors such as worry, fear, and anger, and internally because the Qi dynamics of the zang-fu organs is easily disturbed. The pathological location is chiefly in the liver, with involvement of the heart, spleen, and kidneys. The disease mechanism is characterized by Qi stagnation and dysfunction of the internal organs. Initially, the most of the condition is solid, where Qi stagnation leads to food accumulation, phlegm coagulation, and blood stasis. As the disease progresses, it transforms from solid to virtual, ultimately resulting in deficiency of the five zang organs and an imbalance of Yin and Yang.17 Compared with Western medicine, TCM has unique value owing to its advantages, such as holistic treatment concept, flexible formula composition, multitarget effects, high safety, and minimal side effects. It has shown remarkable efficacy in alleviating depressive symptoms, improving quality of life, and enhancing synergistic effects. Therefore, in-depth exploration of the mechanisms underlying TCM in the treatment of depression and the optimization of clinical application protocols have become important directions in the field of depression research. In recent years, extensive and fruitful research has been conducted in the field of TCM, focusing on the regulation of PCD. Accordingly, this review focuses on six types of PCD associated with the pathological mechanisms of depression, explores their molecular mechanisms and roles in the onset and progression of depression, systematically summarizes TCM therapies that target PCD to intervene in depression, and discusses the interconnections between different types of PCD. This study aims to provide references for the prevention, diagnosis, and treatment of depression.
Patterns of PCD in Depression
The pathological process of depression involves multiple forms of PCD, primarily including apoptosis, pyroptosis, autophagy, ferroptosis, necroptosis, and cuproptosis (Figure 1).
 |
Figure 1 Molecular mechanisms of PCD in depression. The extrinsic apoptotic pathway (Caspase-8/10), intrinsic apoptotic pathway (BH3-only proteins), and ER pathway can all induce apoptosis. The occurrence of pyroptosis depends on the classical pathway, which is mediated by NLRP3-Caspase-1, and the nonclassical pathway, which is mediated by LPS-Caspase-4/5/11. Ferroptosis is triggered by Fe2+ overload, lipid peroxidation (PUFAs), and dysregulation of the antioxidant defense system (System Xc−/GSH/GPX4). Necroptosis is regulated mainly by the RIPK1-RIPK3-MLKL axis. AMPK can induce autophagy either by inhibiting the phosphorylation of mTORC1 or directly inducing the phosphorylation of the ULK1 complex. Cuproptosis in cells can be caused by increased Cu+ levels, the accumulation of DLAT, and the loss of Fe-S clusters, all of which are induced by FDX1. This diagram was created at https://www.biorender.com. Abbreviations: BH3-only, BCL-2 homology 3 domain-only proteins; BCL-2, B-cell lymphoma-2; BCL-XL, B-cell lymphoma extra-large; BAK, BCL-2 homologous antagonist/killer; MOMP, Mitochondrial outer membrane permeabilization; ER, Endoplasmic reticulum; ERS, Endoplasmic reticulum stress; PERK, Protein kinase RNA–like endoplasmic reticulum kinase; IRE1α, Inositol-requiring enzyme 1α; ATF6, Activating transcription factor 6; LPS, Lipopolysaccharide; IL-1β/18, Interleukin-1β/18; GSDMD, Gasdermin D; Caspase-1/4/5/11, Cysteinyl aspartate specific proteinase; ROS, Reactive oxygen species; PUFAs, Polyunsaturated fatty acids; GSH, Glutathione; GPX4, Glutathione peroxidase 4; RIPK1/3, Receptor-interacting serine/threonine-protein kinase 1/3; MLKL, Mixed lineage kinase domain like pseudokinase; AMPK, Adenosine 5′-monophosphate-activated protein kinase; mTORC1, Mechanistic target of rapamycin complex 1; ULK1, UNC-51-like kinase 1; VPS34, Vesicular protein sorting 34; ATG12/15/16L1, Autophagy-related genes 12/15/16L1; LC3, Microtubule-associated protein 1 light chain 3; FDX1, Ferredoxin 1; DLAT, Dihydrolipoamide S-acetyltransferase; Fe-S, Iron-sulfur proteins. |
Apoptosis
Apoptosis, the earliest discovered form of PCD, plays a crucial role in maintaining tissue homeostasis and eliminating abnormal cells. This mode of cell death is characterized by the activation of Caspases and can be categorized into extrinsic pathway (also known as death receptor pathway), intrinsic pathway (also referred to as mitochondrial pathway), and ER pathway on the basis of differences in activation mechanisms. The extrinsic apoptotic pathway is triggered by the binding of death receptors on the cell surface to their corresponding ligands. Upon binding of cell surface death receptors—such as tumor necrosis factor receptor 1 (TNFR1), FAS receptor (FAS), and TNF-related apoptosis-inducing ligand receptor 1 and 2 (TRAIL-R1/R2)—to their corresponding ligands (tumor necrosis factor-α (TNF-α), Fas ligand (FASL), and TRAIL), conformational changes occur. Through their intracellular death domains, these receptors recruit adaptor proteins such as FAS-associated protein with death domain (FADD), which further recruit and activate pro-Caspase-8/10, leading to the formation of a death-inducing signaling complex. This process promotes the activation of Caspase-8 and Caspase-10, which in turn cleave Caspase-3 and Caspase-7, ultimately triggering apoptosis.18 The intrinsic apoptotic pathway is driven primarily by MOMP, whose core regulation relies on the BCL-2 protein family. When cells are subjected to internal stresses such as DNA damage, oxidative stress, or ERS, BH3-only proteins are activated. These proteins inhibit antiapoptotic proteins and activate the effector proteins BAX/BAK, thereby triggering MOMP.19 MOMP leads to the release of cytochrome c and second mitochondria-derived activator of caspases (SMAC) from mitochondria into the cytoplasm. Cytochrome c then binds to apoptotic peptidase activating factor 1 and pro-Caspase-9 to form an apoptosome, which promotes the activation of Caspase-9. This in turn activates Caspase-3/7, initiating a Caspase cascade that induces apoptosis;20 SMAC promotes the activation of Caspase-9 by binding to inhibitors of apoptosis proteins (IAPs) and abrogating their inhibitory activity, thereby further amplifying apoptotic signals.21 Additionally, accumulating evidence indicates that ERS is another crucial pathway that induces apoptosis. ERS is triggered primarily by the abnormal accumulation of unfolded proteins and can induce apoptosis through the activation of three major unfolded protein response sensors: PERK, IRE1α, and ATF622 (Figure 2).
 |
Figure 2 Molecular mechanisms of apoptosis. Apoptosis is activated through three pathways: the extrinsic, intrinsic, and ER pathways. The extrinsic apoptotic pathway is triggered by the binding of death receptors on the cell surface to their corresponding ligands, which activates Caspase-8/10 and subsequently Caspase-3/7 to induce apoptosis. The core regulation of the intrinsic apoptotic pathway relies on the activation of the BCL-2 protein family, which then activates the proapoptotic proteins BAX and BAK, inhibits antiapoptotic proteins, promotes the activation of Caspase-9, and further activates Caspase-3/7 to trigger apoptosis. The ER apoptotic pathway induces apoptosis by inducing ERS, thereby activating PERK, IRE1α, and ATF6. This diagram was created at https://www.biorender.com. Abbreviations: BID, BH3 interacting domain death agonist; tBID, Truncated BID. |
Apoptosis constitutes a crucial pathological basis for brain functional impairment. Studies have shown that the number of apoptotic neurons in the hippocampus and prefrontal cortex (PFC) of depressed rats induced by chronic unpredictable mild stress (CUMS) is significantly increased,23 accompanied by an imbalance in the expression of apoptosis-related proteins. This is manifested by elevated levels of the proapoptotic factors BAX and Caspase-3 and decreased levels of the antiapoptotic factor BCL-2,7,8 suggesting that apoptosis is one of the important pathological mechanisms contributing to the development of depression. Microglia are crucial for maintaining brain function. As resident immune cells in brain tissue, their apoptosis exacerbates neuroinflammation and neuronal damage, thereby participating in the onset and progression of depression.24 Astrocytes, on the other hand, play a central role in the pathological mechanisms of depression by regulating neuroinflammation, neural activity, synaptic plasticity, and energy metabolism.25 When astrocytes undergo apoptosis, the survival and function of neurons are severely impaired, which in turn exacerbates depressive states. In animal models of depression induced by chronic social defeat stress, the apoptosis of both astrocytes and microglia is clearly observed,24,26 suggesting that glial cell damage may be one of the core links in the pathology of depression. Further studies revealed that in the hippocampus and PFC of mice subjected to CUMS-induced depression, the expression of TNFR1 was significantly upregulated. Upon binding to TNF-α, TNFR1 can trigger the transformation of astrocytes into the proinflammatory A1 phenotype, promote microglial activation, induce neuroinflammation, and ultimately lead to neuronal apoptosis, thereby exacerbating depressive symptoms.27,28 In addition, the ER pathway has been identified as a crucial driver of the onset and progression of depression. The ER is activated when cells are exposed to various stressors, and prolonged or severe ERS can trigger apoptotic programs. In CUMS-induced rats, the expression levels of ERS marker proteins (C/EBP-homologous protein (CHOP), Caspase-12, and c-Jun N-terminal kinase 1) in the hippocampus are abnormally elevated, which is significantly correlated with an increased number of apoptotic hippocampal neurons. These findings suggest that the activation of the ER pathway is also an important mechanism promoting neuronal apoptosis.29 Therefore, targeted regulation of apoptosis in neurons and glial cells is expected to become a key entry point for improving depressive symptoms and their associated brain functional impairments.
Pyroptosis
Pyroptosis is a lytic and inflammatory form of PCD that is typically triggered by inflammasomes, mediated by Caspases, and executed by Gasdermins. It is characterized by progressive cellular swelling until membrane rupture and the release of intracellular contents, which in turn activates a robust inflammatory response.30 Currently, pyroptosis is generally classified into the classical pathway, which is mediated by Caspase-1, and the nonclassical pathway, which is mediated by Caspase-4/5/11. In the canonical pyroptotic pathway, upon stimulation by pathogenic microorganisms, ROS, or other stimuli, intracellular pattern recognition receptors (PRRs) recognize pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (DAMPs). These PRRs then bind to apoptosis-associated speck-like protein containing a CARD (ASC) and pro-Caspase-1 to assemble into inflammasomes. This complex further activates Caspase-1; on the one hand, activated Caspase-1 cleaves GSDMD to release the active N-terminal fragment GSDMD-N, which forms pores in the plasma membrane, induces inflammatory responses, and triggers pyroptosis. On the other hand, it cleaves pro-IL-1β/18 to generate mature IL-1β/IL-18, which are released extracellularly through pores to amplify the inflammatory response.31,32 The noncanonical pyroptotic pathway is primarily activated by LPS. LPS specifically binds to and activates Caspase-4/5/11; activated Caspase-4/5/11 can cleave GSDMD to generate GSDMD-N, thereby executing pyroptosis. Although they do not directly process pro-IL-1β/IL-18, activated Caspase-4/5/11 triggers K+ efflux via cleavage of GSDMD, which in turn induces the assembly of the NLRP3 inflammasome, a process analogous to the classical pyroptotic pathway.31,32 In contrast to the canonical pathway, in noncanonical pyroptosis, only the cleavage of pro-IL-1β/IL-18 is dependent on Caspase-1, whereas the cleavage of GSDMD is executed by other activated inflammatory Caspases. Pannexin-1 represents another critical mediator of pyroptosis induced by noncanonical Caspase-11. Exposure to LPS activates Caspase-11, which in turn cleaves the Pannexin-1 channel to facilitate ATP efflux, thereby activating purinergic receptor P2X7 (P2X7) and ultimately mediating the occurrence of pyroptosis.33 In recent years, it has been discovered that Caspase-3/8 and granzymes can also mediate pyroptosis by cleaving Gasdermins, but the specific mechanisms require further exploration34,35 (Figure 3).
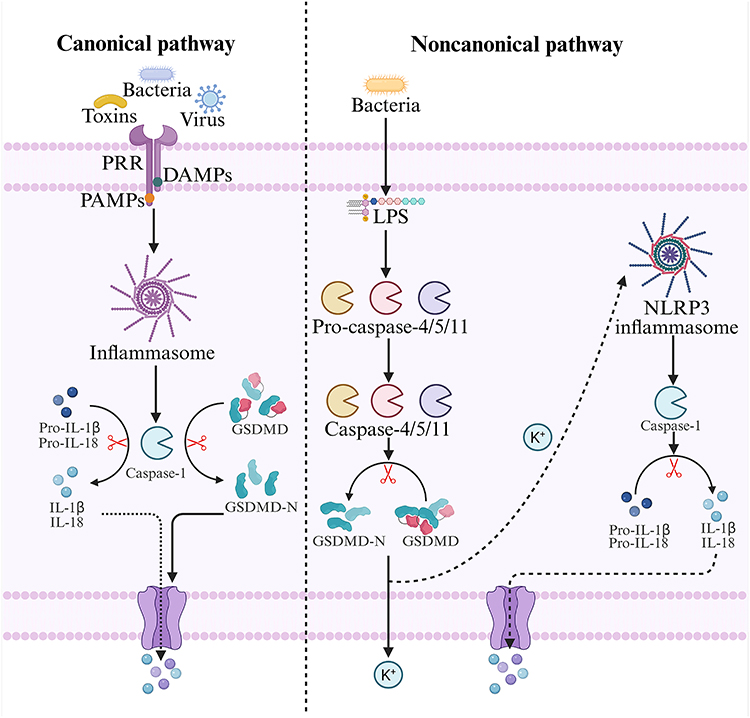 |
Figure 3 Molecular mechanisms of pyroptosis. In the canonical pyroptotic pathway, intracellular PRRs recognize PAMPs or DAMPs to activate inflammasomes, which in turn induce the activation of Caspase-1. Activated Caspase-1 cleaves GSDMD to generate GSDMD-N and promotes the maturation of IL-1β and IL-18, thereby triggering pyroptosis. The noncanonical pyroptotic pathway is activated mainly by LPS. LPS binds to and activates Caspase-4/5/11; activated Caspase-4/5/11 cleaves GSDMD to produce GSDMD-N, which elicits K+ efflux and leads to the formation of the NLRP3 inflammasome. Like the classical pyroptotic pathway, this process activates Caspase-1, promotes the maturation of IL-1β and IL-18, and ultimately induces pyroptosis. This diagram was created at https://www.biorender.com. |
In pyroptosis, inflammasomes are regarded as key therapeutic targets for depression, among which the NLRP3 inflammasome has been the most extensively studied. Research has revealed that in depression models, the NLRP3 inflammasome is highly activated, which in turn induces pyroptosis in the hippocampal neurons of depressed rats in an NLRP3/Caspase-1/GSDMD-dependent manner. This is accompanied by elevated levels of GSDMD-N, NLRP3, ASC, active Caspase-1, cleaved IL-1β, and cleaved IL-18, thereby promoting the development of depression.9 Notably, this NLRP3 inflammasome-driven pyroptotic mechanism not only occurs in neurons but also dominates the death process of glial cells, further amplifying the pathological damage in depression. Pyroptosis in microglia further confirms the role of the NLRP3 inflammasome. Microglia promote the activation of Caspase-1 through the activation of the NLRP3 inflammasome, which in turn cleaves GSDMD to generate GSDMD-N. This leads to microglial rupture and the release of proinflammatory factors such as IL-1β, triggering neuroinflammation, impairing synaptic plasticity in the hippocampus, and inducing depressive behaviors.36,37 Additionally, in depression models, hippocampal astrocytes also exhibit characteristic pyroptotic changes, manifested by the activation of the NLRP3 inflammasome, elevated expression of pyroptotic executive proteins such as Caspase-1 and GSDMD-N, and increased release of proinflammatory factors such as IL-1β. These changes result in a reduction in the number of hippocampal astrocytes, disruption of the neural microenvironment, and damage to synaptic structures.38,39 Genetic knockout of GSDMD, Caspase-1, or astrocyte-specific NLRP3 significantly alleviates depressive-like behaviors in mice.38 Therefore, treatments for depression can target pyroptosis-inhibiting pathways, such as suppressing the activation of the NLRP3 inflammasome, antagonizing the maturation of inflammatory Caspases, and inhibiting GSDMD cleavage.
Autophagy
Autophagy is a lysosome-dependent cellular degradation process tightly regulated by ATGs. It maintains intracellular homeostasis by clearing and recycling misfolded or aggregated proteins, damaged organelles, and pathogens.40 Autophagy can be classified into nonselective autophagy and selective autophagy. Nonselective autophagy can be further divided into macroautophagy, microautophagy, and chaperone-mediated autophagy on the basis of the distinct ways in which degraded substances are transported to lysosomes. Macroautophagy, the most prominent type, is referred to simply as “autophagy” in this article.41 Selective autophagy can be further categorized into mitophagy, peroxisome autophagy, and endoplasmic reticulum autophagy, among others, on the basis of differences in the subcellular locations where autophagy occurs.42 The process of autophagy, from initiation to functional execution, primarily involves three stages: phagophore formation, autophagosome formation, and degradation. During the phagophore formation stage, AMPK is activated under nutrient deprivation conditions, which inhibits the downstream negative regulator of autophagy, mTORC1. This inhibition leads to the dissociation of mTORC1 from the ULK1 complex (comprising ULK1, ATG13, focal adhesion kinase family interacting protein of 200kDa (FIP200), and ATG101), thereby resulting in mTORC1-mediated inhibitory phosphorylation of ULK1 and ATG13, allowing their activation. On the other hand, AMPK directly phosphorylates ULK1, promoting the activation of the ULK1 complex and the formation of autophagosomal membranes. This complex subsequently activates the downstream VPS34 complex (Beclin-1-ATG14-VPS15-VPS34), which catalyzes the production of phosphatidylinositol-3-phosphate (PI3P) on autophagosomal membranes, initiating the nucleation and initial expansion of the phagophore. During the autophagosome formation stage, the phagophore elongates and closes to form an autophagosome, a process that requires two ubiquitin-like systems. The first system involves the formation of the ATG12-ATG5-ATG16L1 protein complex, which acts on the outer membrane of the phagophore and participates in the elongation of the outer membrane of the phagophore. The second system begins with ATG4 cleaving LC3 to generate LC3-I. Upon autophagy activation, LC3-I is conjugated with phosphatidylethanolamine (PE) to form lipid-soluble LC3-II under the action of ATG3, ATG7, and the ATG12-ATG5-ATG16L1 complex. LC3-II interacts with the ATG12-ATG5-ATG16L1 protein complex to promote phagophore elongation, enabling it to enwrap autophagic substrates and form a spherical structure. Finally, with the closure of the pore, a double-membraned vesicle-like autophagosome is formed. Eventually, the autophagosome is transported to the lysosome, where its outer membrane fuses with the lysosomal membrane to form an autolysosome, and the contents are degraded43 (Figure 4).
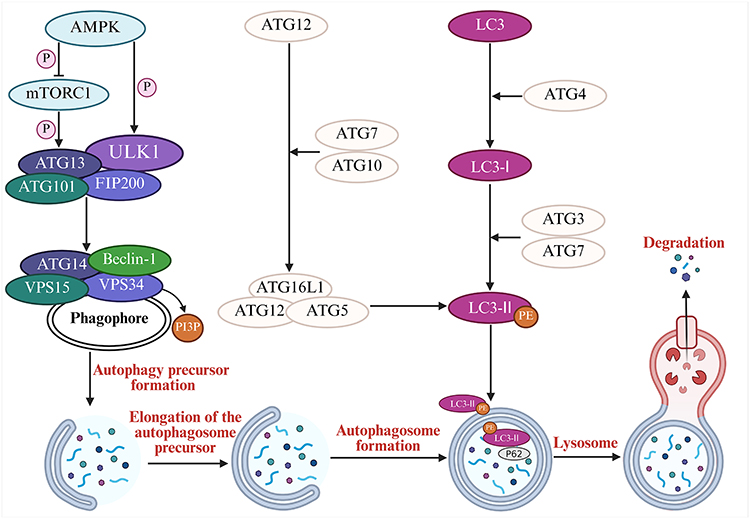 |
Figure 4 Molecular mechanisms of autophagy. AMPK activates autophagy by inhibiting mTORC1 or directly phosphorylating the ULK1 complex. The activated ULK1 complex further activates the VPS34 complex (Beclin-1-ATG14-VPS15-VPS34), which catalyzes the generation of PI3P to initiate the nucleation and initial expansion of the phagophore. Two ubiquitin-like systems subsequently drive autophagosome formation: 1. The ATG12-ATG5-ATG16L1 complex localizes to the outer membrane of the phagophore to promote its elongation; 2. LC3-I is conjugated with PE to form membrane-bound LC3-II under the action of ATG7, ATG3, and the ATG12-ATG5-ATG16L1 complex. LC3-II acts synergistically with the ATG12-ATG5-ATG16L1 complex to facilitate phagophore elongation, the enclosure of substrates, and closure to form double-membraned autophagosomes. Finally, autophagosomes fuse with lysosomes to form autolysosomes, and their contents are degraded. This diagram was created at https://www.biorender.com. Abbreviation: P62, Sequestosome 1. |
In recent years, a growing body of research has confirmed that dysregulated autophagy is closely associated with depression. As an intracellular “waste clearance system,” autophagy plays a central role in maintaining neuronal homeostasis. However, in the complex context of depression, it acts as a “double-edged sword”: both abnormally elevated and reduced activity can act as key drivers in the development of depression. On the one hand, excessive autophagy has destructive effects. In neurons, hyperactivated autophagy may significantly reduce the synthesis of brain-derived neurotrophic factor, directly impairing neuroplasticity and neurogenesis. In glial cells, particularly microglia, excessive autophagy drives their transformation into a proinflammatory phenotype, leading to the release of large amounts of proinflammatory cytokines and triggering neuroinflammatory responses. Thus, inhibiting excessive autophagy can exert antidepressant therapeutic effects.11,44 On the other hand, insufficient autophagic function leads to cumulative toxicity. In the state of depression, autophagic activity often is insufficient, preventing timely clearance of damaged proteins and organelles. These substances thus abnormally accumulate within neurons, directly causing neuronal damage. Therefore, moderate enhancement of autophagy can exert a key neuroprotective effect by eliminating harmful substances, thereby helping to alleviate depression-related neuronal damage.12,45 Notably, dysregulation of mitophagy has been proven to be deeply involved in the progression of depression. When mitophagic function is impaired, dysfunctional mitochondria cannot be effectively recognized and cleared, resulting in their massive accumulation in cells, excessive production of reactive oxygen species, and severe oxidative stress. Moreover, as an inflammatory stimulus, ROS can activate the NLRP3 inflammasome, triggering neuroinflammatory responses and pyroptosis, which ultimately lead to neuronal damage. Hence, enhancing mitophagic function has been identified as an effective strategy to ameliorate depressive symptoms.46,47
Ferroptosis
Ferroptosis is an iron-dependent form of PCD induced by lipid peroxidation. Currently, three major pathways have been identified as critical in ferroptosis: the iron metabolism dysregulation pathway, the lipid peroxidation pathway, and the antioxidant defense system failure pathway.48 Under normal circumstances, extracellular iron ions exist in the form of Fe3+, which can enter cells via the transferrin receptor protein 1 (TFR1) transport system and be reduced to Fe2+ by six-transmembrane epithelial antigen of prostate 3 (STEAP3) before being released into the cellular labile iron pool. When TFR1 is abnormally activated, cells take up excessive iron ions, a phenomenon known as “iron overload,” which triggers the “Fenton reaction” to generate large amounts of ROS and induce lipid peroxidation damage. Intracellular excess Fe2+ is mainly stored in ferritin, while nuclear receptor coactivator 4 (NCOA4)-mediated ferritinophagy can specifically degrade ferritin to release free Fe2+. When ferritinophagy is activated, increased ferritin degradation leads to elevated intracellular free Fe2+ concentrations, ultimately promoting the occurrence of ferroptosis.49 PUFAs are important substrates for lipid metabolism in ferroptosis. They are converted to polyunsaturated-fatty-acid–containing phospholipids (PUFA-PLs) under the action of Acyl-CoA synthetase long chain family member 4 (ACSL4) and lysophosphatidylcholine acyltransferase 3 (LPCAT3). Fe2+ generates hydroxyl radicals through the Fenton reaction, activating lipoxygenases (LOXs), which catalyze the oxidation of PUFA-PLs to form lipid peroxides polyunsaturated fatty acid–containing phospholipid hydroperoxide (PUFA-PL-OOH), leading to ferroptosis.49 Lipid peroxidation can be inhibited by the intracellular antioxidant defense system, among which the system Xc−/GSH/GPX4 axis has been the most extensively studied. System Xc−, composed of solute carrier family 7 member 11 (SLC7A11) and SLC3A2, is involved in the extracellular uptake of cystine and the release of glutamate (Glu), promoting the synthesis of GSH.50 GPX4, a major intracellular antioxidant enzyme, uses GSH to reduce toxic lipid peroxides to nontoxic lipid alcohols while oxidizing GSH, thereby negatively regulating ferroptosis. The expression and activity of GPX4 during ferroptosis depend on the presence of GSH.51 Therefore, depletion of intracellular GSH indirectly inhibits GPX4 activity, and decreased GPX4 expression is a key marker of ferroptosis (Figure 5).
 |
Figure 5 Molecular mechanisms of ferroptosis. Ferroptosis involves three key pathways: dysregulated iron metabolism, lipid peroxidation, and failure of antioxidant defense. In iron metabolism, abnormal activation of TFR1 leads to iron overload, triggering the Fenton reaction to generate ROS; NCOA4-mediated ferritinophagy results in the degradation of ferritin and the release of free Fe2+, which together promote ferroptosis. In the lipid peroxidation pathway, PUFAs are converted to PUFA-PLs via ACSL4 and LPCAT3; the Fenton reaction activates LOXs, which catalyze the formation of lipid peroxides (PUFA-PL-OOH), directly inducing ferroptosis. The failure of the antioxidant defense system, however, cannot prevent this process. Among them, System Xc− mediates cystine uptake for GSH synthesis; GSH, as an essential substrate, maintains the activity of GPX4, which is responsible for reducing lipid peroxides. Depletion of GSH or decreased GPX4 expression causes the collapse of this defense system, collectively leading to cell death. This diagram was created at https://www.biorender.com. Abbreviations: PUFA-CoA, polyunsaturated fatty acid–coenzyme A. |
Ferroptosis can participate in and influence the pathological process of depression through multiple pathways, including neuroinflammation, mitochondrial dysfunction, and gut microbiota dysbiosis.52 Ferroptosis-related genes can also serve as biomarkers for the auxiliary diagnosis of depression.53 Studies have shown that in the hippocampus of LPS-induced depressed mice, the expression of ferritin heavy chain (FTH1) is significantly lower than that in the normal control group. FTH1 is an important intracellular iron storage protein; a decrease in its content often leads to abnormal intracellular iron storage, resulting in iron deposition and overload, generating lipid peroxides. The accumulation of these peroxides damages DNA and proteins, ultimately inducing ferroptosis.54 Proteomic analysis of CUMS-induced mice revealed that the ferroptosis marker ferritin light chain 1 and the lipid peroxidation product malondialdehyde (MDA) are significantly increased, whereas the levels of the antioxidants GSH and GPX4 are significantly decreased,10 suggesting that CUMS-induced depressive-like behaviors may impair neuronal function by inhibiting the GPX4-GSH axis, exacerbating ferroptosis and lipid peroxidation. In addition, a meta-analysis indicates that dietary iron intake is negatively associated with depression risk.55 These studies provide important clues and evidence for the involvement of ferroptosis in the pathological process of depression.
Roles of Other Types of PCD in Depression
In addition to the four types of PCD mentioned above, the roles of necroptosis and cuproptosis in depression have gradually been discovered. Necroptosis is a process of self-destruction that occurs when apoptosis is blocked. Morphologically similar to necrosis, it is characterized by membrane rupture, organelle swelling, and the release of intracellular contents. Necroptosis can be activated by various stimuli, among which the RIPK1/RIPK3/MLKL pathway is the most classic and extensively studied.56 Necroptotic signaling is triggered by death receptors, such as TNFR1, TRAIL-R, FAS, and Toll-like receptors 3/4 (TLR3/4).57 For example, upon stimulation by necrotic signals, TNF-α binds to the TNFR1 receptor and recruits RIPK1, tumor necrosis factor receptor-associated death domain protein (TRADD), cellular inhibitor of apoptosis proteins 1/2 (cIAP1/2), and TNF receptor associated factor 2/5 (TRAF2/5) to form complex I on the cell membrane. After complex I formation, RIPK1, TRADD, Caspase-8, and FADD form complex II under the action of deubiquitinases. When Caspase-8 activity is inhibited, the necroptotic pathway is initiated, inducing the phosphorylation of RIPK1. Phosphorylated RIPK1 binds to RIPK3 to form a necrosome; RIPK3 then undergoes autophosphorylation, followed by phosphorylating MLKL and inducing its oligomerization. MLKL subsequently translocates to the cell membrane, increasing Na+ and Ca2+ influx to alter membrane permeability, form membrane pores, and disrupt membrane integrity, ultimately leading to necroptosis.58 (Figure 6) Proteomic analysis of the hippocampus of CUMS rats revealed that the necroptosis pathway is significantly activated, with increased levels of RIPK3 and p-MLKL.10 In a corticosterone-induced depressed rat model, hippocampal RIPK3 expression is elevated; knockdown of RIPK3 or inhibition of its activity can ameliorate depressive-like behaviors by reducing oxidative stress, restoring synaptic function, and decreasing neuronal death.14 Taken together, these findings indicate that necroptosis is involved in the pathogenesis of depression and that targeted inhibition of the RIPK1/RIPK3/MLKL axis may serve as a novel strategy for the treatment of depression.
 |
Figure 6 Molecular mechanisms of necroptosis and cuproptosis. Necroptosis is triggered by death receptors. For example, the binding of TNF-α to TNFR1 first promotes the assembly of Complex I at the plasma membrane, which contains RIPK1. Subsequently, upon deubiquitination of RIPK1, this membrane-associated complex dissociates and reforms in the cytosol as Complex II, which contains RIPK1, caspase-8, FADD, and other components. When the activity of Caspase-8 is inhibited, the necroptosis pathway is initiated: RIPK1 is phosphorylated and binds to RIPK3 to form necrosomes; phosphorylated RIPK3 then phosphorylates MLKL, leading to its oligomerization and translocation to the cell membrane; and oligomerized MLKL alters membrane permeability by increasing Na+ and Ca2+ influx, forming membrane pores, and ultimately disrupting membrane integrity, triggering necroptosis. Cuproptosis is triggered by excessive copper, and its occurrence depends on intracellular copper accumulation and the lipoylation state of mitochondrial tricarboxylic acid cycle enzymes. When the copper ionophore Elesclomol disrupts copper homeostasis regulated by the copper transporters SLC31A1/ATP7α/β, it leads to intracellular copper accumulation; the mitochondrial matrix reductase FDX1 reduces Cu2+ to Cu+ and releases it into mitochondria, whereas FDX1, as an acylation effector, promotes the accumulation of lipoylated DLAT proteins. Cu+ binds to lipoylated DLAT, triggering disulfide bond-dependent aggregation to form toxic aggregates, accompanied by the loss of iron‒sulfur cluster proteins, ultimately inducing proteotoxic stress and leading to cell death. This diagram was created at https://www.biorender.com. |
Cuproptosis is a novel form of programmed cell death triggered by copper and was first proposed by Science in 2022.59 Copper, an essential trace element in the human body, plays a crucial role in biological processes such as mitochondrial respiration, antioxidant defense, and neurotransmitter production. However, excessive copper can be toxic to cells or tissues, leading to copper overload or copper poisoning.60 Cuproptosis depends on intracellular copper levels and the lipoylation status of tricarboxylic acid cycle enzymes and is mediated by two mitochondrial protein toxicity stress pathways. When cells are exposed to the copper ionophore elesclomol, copper homeostasis regulated by the copper transporters SLC31A1 and ATPase copper transporting alpha/beta (ATP7α/β) is disrupted, resulting in intracellular copper accumulation.49 Mitochondrial matrix reductase FDX1 catalyzes the reduction of Cu2+ to Cu+ and increases mitochondrial Cu+ availability.49,61 FDX1 also acts as a novel acylation effector, contributing to the accumulation of lipoylated DLAT.49,61 Cu+ binds to lipoylated DLAT and promotes disulfide bond-dependent aggregation of lipoylated DLAT, thereby leading to the accumulation of toxic acylated DLAT.49 Concurrently, the loss of Fe‒S protein induces proteotoxic stress, ultimately resulting in cuproptosis (Figure 6). Research indicates elevated levels of copper ions in the serum of depressed animals.13 This disruption of copper homeostasis may be involved in the pathogenesis of depression by inducing oxidative stress and inflammatory responses.62 In summary, cuproptosis is a newly discovered pattern of PCD; however, research on this mechanism in the field of depression is still in its infancy and urgently requires in-depth verification.
Connections Between Different Types of PCD
Apoptosis, pyroptosis, autophagy, ferroptosis, necroptosis, and cuproptosis are involved in the pathophysiological progression of depression, with each mode exhibiting unique cell biological, morphological, and biochemical characteristics. In depression, multiple types of PCD may coexist and interact through complex mechanisms.
As a key regulatory molecule, Caspase-8 connects apoptosis, pyroptosis, and necroptosis. It serves as a core initiator of the extrinsic apoptotic pathway and inhibits the RIPK3/MLKL-mediated necroptotic pathway. However, when the enzymatic activity of Caspase-8 is lost and necroptosis is blocked, its expression triggers ASC formation, Caspase-1 activation, and IL-1β release, thereby initiating pyroptosis.63 This “molecular switch” property suggests that Caspase-8 could serve as a therapeutic target for integrating multiple forms of PCD. Ferroptosis and cuproptosis, as metal ion-dependent forms of PCD, are interconnected through the action of GSH. As a core antioxidant, GSH plays dual roles in both ferroptosis and cuproptosis. It can inhibit ferroptosis by reducing lipid peroxides via the GSH/GPX4 pathway,51 and can also chelate excessive intracellular Cu+ to reduce its aggregation of lipoylated proteins and damage Fe-S protein, thereby blocking cuproptosis.59 GSH depletion not only enhances ferroptosis sensitivity but also leads to Cu+ accumulation. The latter can degrade SLC7A11 by increasing ROS levels, preventing cystine from entering cells, further inhibiting GSH synthesis, and triggering ferroptosis.64 Furthermore, ferroptosis and necroptosis are linked through ROS and lipid metabolism. ROS are key factors that induce necroptosis; iron overload activates necroptosis by increasing ROS levels and stimulating the opening of the mitochondrial permeability transition pore (MPTP).65 MLKL may inhibit ferroptosis by altering PUFA-containing phospholipids, while the overexpression of ACSL4, a key enzyme in ferroptosis, can downregulate GPX4 and weaken the cellular membrane response to MLKL, thereby inhibiting necroptosis.66 Conversely, knockdown of ACSL4 suppresses ferroptosis and activates necroptosis.67
Autophagy serves as a core regulatory node in the PCD network, interacting extensively with other death modalities. Autophagy and apoptosis can influence each other through mitochondrial function, antiapoptotic proteins, and the mammalian target of rapamycin (mTOR) signaling pathway. The antiapoptotic proteins BCL-2/BCL-XL prevent the formation of the autophagy-initiating complex between Beclin-1 and VPS34 by binding to Beclin-1, a key autophagic molecule, while also inhibiting apoptosis by suppressing proapoptotic proteins such as BAX/BAK.67 In contrast, BH3-only proteins competitively bind to BCL-2/BCL-XL, relieving the inhibition of both Beclin-1 and BAX/BAK, thereby coordinately activating autophagy and apoptosis.68 mTOR is a core inhibitor of autophagy. Ablation of Raptor, a positive regulator of mTORC1, can activate Caspase-3, induce mitochondrial dysfunction, and promote both apoptosis and autophagy.69 Mitophagy can inhibit apoptosis by degrading mitochondria with increased permeability, preventing the release of apoptosis-inducing factors such as cytochrome c into the cytoplasm.70 The relationship between autophagy and necroptosis remains controversial. Studies have shown that necroptosis is often accompanied by autophagy activation, and its inhibitor necrostatin-1 can reduce the level of the autophagy marker LC3-II and block autophagy.71 However, the activated necroptotic effector protein MLKL is partially retained in autophagosomes, interfering with the fusion and degradation of autophagosomes and lysosomes and thereby inhibiting the completion of autophagy.72 In addition, mitophagy can regulate intracellular ROS levels, trigger the opening of the MPTP, and induce necroptosis through the RIP1/RIP3/MLKL pathway.73 There is also a complex bidirectional regulatory relationship between autophagy and pyroptosis. On the one hand, autophagy can activate Caspase-3 through activating the intrinsic Caspase-9 and extrinsic Caspase-8 apoptotic pathways and then cleave GSDME to promote pyroptosis.74 On the other hand, it can degrade the NLRP3 inflammasome and inhibit Caspase-1 activity to suppress pyroptosis.75 Conversely, IL-1β released by pyroptosis can induce autophagy, whereas activated NLRP4 inhibits the initiation of autophagy by binding to Beclin-1.70 Ferroptosis and cuproptosis also interact with autophagy. Ferroptosis is regulated mainly by iron homeostasis and oxidative stress, with iron homeostasis partially controlled by ferritin. During ferroptosis, autophagy induces the release of intracellular free iron through NCOA4-mediated ferritinophagy, triggering the Fenton reaction.76,77 Inhibiting NCOA4 or disrupting autophagy can eliminate the accumulation of Fe2+ and ROS caused by ferroptosis.77 Additionally, Beclin-1 can bind to SLC7A11, reducing GSH synthesis and thereby triggering ferroptosis.77 Cu+ induces autophagy by generating ROS through the Fenton reaction. Excessive ROS can overactivate autophagy, accelerating cell death by selectively degrading damage-resistant or antioxidant proteins.78 Excess copper can also promote autophagy by increasing ATG expression, regulating the AMPK/mTOR pathway, activating transcription factor EB, and binding to ULK1/2.79
The aforementioned research indicates that different modes of PCD do not exist in isolation during the development and progression of depression. Instead, they may form an intricate interactive network that collectively regulates its pathological process, underscoring the complexity and significance of cell death in depression. Shifting the focus from single pathways to multi-pathway interactive networks will not only contribute to a comprehensive understanding of the pathogenesis of depression but may also provide a theoretical basis for developing multi-target therapeutic strategies.
Research on TCM Targeting PCD for the Treatment of Depression
The dysregulation of PCD has been identified as a core pathological mechanism of depression, and TCM, with its therapeutic characteristics of multiple components and multiple targets, has shown significant advantages in the precise regulation of the PCD network.
Apoptosis
Targeting apoptosis is a key strategy for alleviating neural damage in depression, with mechanisms involving the intrinsic, extrinsic, and ER pathways. TCM can effectively ameliorate depressive symptoms by regulating the aforementioned networks to reduce apoptosis. Berberine is a natural isoquinoline alkaloid extracted from Chinese medicinal materials such as Curcuma longa L., Coptis chinensis Franch. and Phellodendron chinense. It has multiple pharmacological effects, including anti-inflammatory, antioxidant, and antibacterial properties. Studies have confirmed that it can ameliorate depressive symptoms by activating the phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT)/cyclic adenosine monophosphate-response element binding protein (CREB)/brain-derived neurotrophic factor (BDNF) signaling pathway, inhibiting inflammatory responses, downregulating the expression levels of the proapoptotic proteins BAX and Caspase-3, and increasing the expression of the antiapoptotic protein BCL-2.80 In addition to its anti-inflammatory, bacteriostatic, antiviral, and antioxidant effects, syringa oblata Lindl. can exert antidepressant effects by regulating the cyclic adenosine monophosphate-protein kinase A/CREB/BDNF pathway, reducing the levels of BAX and Caspase-3, increasing the level of BCL-2, alleviating neuronal apoptosis, and promoting the recovery of neuroplasticity.81 Gastrodin, an extract from the dried rhizomes of Gastrodia elata, has been shown in network pharmacology-based animal experiments to increase the levels of serotonin, dopamine, and norepinephrine and ameliorate depressive-like behaviors in CUMS rats via a mechanism involving the inhibition of the Caspase-3-mediated apoptotic pathway.82 Rehmannioside, the main active component of Rehmannia glutinosa, exerts antidepressant effects by inhibiting the expression of BAX, Caspase-3, and ERS-related proteins such as glucose regulated protein 78kDa, X-box binding protein 1, ATF6, ATF4, and CHOP, increasing the level of BCL-2, and simultaneously inhibiting the excessive activation of the mitogen-activated protein kinase signaling pathway.83 Arctigenin, the main pharmacologically active component of Arctium lappa L., possesses various biological properties, such as anti-inflammatory and antiviral effects. It can regulate extrinsic apoptosis by modulating the TNF-α/TNFR1/nuclear factor kappa-B (NF-kB) signaling pathway and inhibiting microglial activation and neuroinflammation, thereby exerting antidepressant effects.28 According to studies on Chinese herbal compounds, Baihe Dihuang decoction can alleviate depressive symptoms by regulating the silent information regulator sirtuin 1 (SIRT1)/high mobility group box 1 protein signaling pathway, inducing M2 polarization of microglia, and reducing neuroinflammation and neuronal apoptosis.8 Additionally, in the field of acupuncture therapy, acupuncture at the Shangxing (GV23) and Fengfu (GV16) acupoints can activate the extracellular regulated protein kinases (ERK) signaling pathway, regulate the expression of apoptosis-related proteins (upregulating BCL-2 and downregulating BAX and Caspase-3), reduce hippocampal neuronal apoptosis, and produce antidepressant effects.7 Therefore, targeting apoptotic pathways can serve as an effective strategy for depression intervention and provide novel combinatorial targets for subsequent multimodal combination therapies.
Pyroptosis
Currently, the core mechanism by which TCM intervenes in the pyroptosis process to improve depression lies in the inhibition of the NLRP3-mediated pyroptosis pathway. Saikosaponin, derived from the roots of Bupleurum scorzonerifolium Willd., possesses pharmacological properties such as anti-inflammatory and antibacterial effects. Studies have shown that it can target and bind to the P2X7 receptor, downregulate its expression, and suppress activation of downstream pyroptosis-related proteins such as NLRP3, Caspase-1, and GSDMD, thereby reducing microglial pyroptosis, repairing the morphology of hippocampal neurons, and ultimately ameliorating depressive-like behaviors.37 Salidroside, an active extract from Rhodiola rosea L., has antioxidant and immunomodulatory effects. This component exerts antidepressant effects by downregulating the expression of proteins related to the P2X7/NF-kB/NLRP3 signaling pathway and simultaneously increasing hippocampal BDNF levels.84 Isoliquiritin, a major flavonoid glycoside extracted from licorice, possesses pharmacological activities such as antiangiogenic, antineurotoxic, and antitumor effects. It exerts antidepressant effects by upregulating MicroRNA-27a and targeting and inhibiting spleen tyrosine kinase gene transcription, thereby blocking the NF-kB-mediated NLRP3 pyroptotic cascade, reducing the release of inflammatory factors, inhibiting neuronal pyroptosis.85 Paeoniflorin, the main active component of Radix Paeoniae Rubra and Radix Paeoniae Alba and it exhibits anti-inflammatory and sedative effects. It can reduce the levels of proinflammatory cytokines in the hippocampus, inhibit excessive activation of microglia and neuronal damage, and simultaneously ameliorate depressive symptoms by upregulating SIRT1 protein expression and downregulating the levels of pyroptosis-related proteins.36 Additionally, acupuncture at Shangyang (LI1) and Fengfu (GV16) can inhibit the NLRP3-mediated pyroptotic pathway in the hippocampus, alleviate neuroinflammation and neuronal damage, and exert antidepressant effects.86 In summary, TCM can effectively inhibit the activation and expression of pyroptosis-related proteins by targeting the NLRP3 inflammasome and its regulatory network, ultimately improving depressive-like behaviors. This provides an important target for the clinical treatment of depression on the basis of TCM theory.
Autophagy
In terms of exerting antidepressant effects, Chinese medicine monomers and compounds promote mainly autophagy. Saffron is a TCM with anti-inflammatory and antibacterial properties. Studies have shown that kaempferol-3-O-sophoroside, an extract from saffron petals, can specifically bind to AMPK, activate its downstream pathways, promote BDNF-mediated neuroplasticity, and induce enhanced autophagy—characterized by an increased LC3-II/LC3-I ratio, upregulated Beclin-1 expression, and downregulated p62 expression—thereby ameliorating depressive symptoms.12 Ginsenoside Rb1, an active component of the Chinese herb ginseng, has multiple effects, such as anti-inflammatory, antitumor, and antibacterial properties. It clears damaged mitochondria by activating mitophagy, reduces ROS production, inhibits NF-kB pathway-mediated NLRP3 activation, alleviates astrocyte pyroptosis, protects hippocampal neurons, and improves synaptic plasticity.39 Gypenosides, triterpenoid saponins extracted from Gynostemma pentaphyllum (Thunb.) Makino, have antitumor, anti-aging, and antioxidant effects. Studies have confirmed that gypenosides can activate SIRT1, enhance the mitochondrial PTEN-induced putative kinase 1/Parkin pathway, and promote mitophagy, thereby inhibiting oxidative stress and neuroinflammation and ameliorating depressive-like behaviors.87 Resveratrol, a natural polyphenolic phytoalexin, reshapes mitochondrial dynamics by activating the SIRT1/Peroxisome prolilerator-activated receptor γ coactivator lalpha/SIRT3 pathway, promotes mitophagy, alleviates inflammatory responses and neuronal apoptosis, and relieves depressive symptoms.88 The Chinese herbal compound Xiaoyaosan can enhance microglial autophagy by inhibiting the activation of the PI3K/AKT/mTOR pathway, thereby suppressing M1 polarization and neuroinflammation and ultimately ameliorating LPS-induced depressive-like behaviors.89 In contrast, acupuncture therapy mainly inhibits excessive autophagy. Electroacupuncture at Baihui (GV20) and Governor Vessel can improve CUMS-induced depressive-like behaviors by inhibiting the autophagy in the hippocampus. However, it primarily downregulates LC3-II/LC3-I ratio to inhibit autolysosome formation, with little effect on Beclin-1, suggesting that its action may be independent of the Beclin-1 pathway.90 Therefore, regulating the bidirectional balance of autophagic function is an important approach to improve depressive-like behaviors, which provides a more comprehensive perspective for TCM in the treatment of depression.
Ferroptosis
Ferroptosis is an important link in the pathogenesis of depression, and TCM has shown significant effects in regulating ferroptosis to improve depression. Resveratrol, a natural polyphenolic substance, has antioxidant, anti-inflammatory, anticancer, and cardiovascular protective effects. Studies have shown that resveratrol can effectively inhibit lipid peroxidation and iron overload by activating the AKT/nuclear factor erythroid 2-related factor 2 (NRF2) pathway and increasing the expression of antioxidant and iron metabolism-related molecules such as heme oxygenase-1 (HO-1), GPX4, SLC7A11, and FTH1, thereby blocking the ferroptosis process and ameliorating depressive-like behaviors.91 Asiaticoside, a triterpenoid compound extracted from Centella asiatica, has multiple pharmacological effects, such as antitumor, anti-inflammatory, and antifibrotic properties. This component can exert antidepressant effects by activating the BDNF/NRF2/GPX4 signaling pathway to inhibit ferroptosis in hippocampal neurons while repairing mitochondrial damage and alleviating neuroinflammation.92 Hydroxysafflor yellow A, a natural flavonoid compound isolated from safflower, can activate the SIRT1/NRF2/NF-κB signaling pathway to promote the recovery of GPX4 and SLC7A11 protein expression, reduce the hippocampal Fe2+ concentration, block ferroptosis, simultaneously inhibit excessive activation of microglia, alleviate neuroinflammation, and ameliorate depressive symptoms.93 Saikosaponin B2 has been shown to upregulate the levels of GPX4, SLC7A11, and FTH1; downregulate the levels of ACSL4 and TFR1; reduce the intracellular Fe2+ concentration and ROS levels; and inhibit iron-dependent lipid peroxidation. Moreover, saikosaponin B2 can alleviate depression through dual regulation of inflammatory responses: it inhibits the TLR4/NF-κB pathway; reduces excessive activation of microglia and the release of IL-1β, IL-6, and TNF-α; and increases IL-10 levels.94 The Chinese herbal compound Xiaoyaosan can improve depressive-like behaviors in CUMS model mice by inhibiting phosphatidylethanolamine binding protein 1-GPX4-mediated ferroptosis, regulating the ERK signaling pathway, and modulating the functions of neuroglial cells.95 In acupuncture therapy, acupuncture at the Shangxing (GV23) and Fengfu (GV16) acupoints inhibits ferroptosis by activating the hippocampal SIRT1/NRF2/HO-1/GPX4 pathway, suppressing the TLR4/NF-κB pathway, reducing glial cell activation, decreasing the hippocampal Fe2+ content, and thus relieving ferroptosis-induced depressive-like behaviors.96 TCM regulates ferroptosis-related processes, highlighting its multidimensional antidepressant effects and indicating that targeting ferroptosis has important research value and application prospects in the treatment of depression.
Necroptosis and Cuproptosis
Currently, research on improving depression through targeting necroptosis and cuproptosis is in its infancy. For the regulation of necroptosis, studies have focused mainly on inhibiting the activation of the RIPK1/RIPK3/MLKL pathway. Xiaoyaosan can downregulate the expression levels of hippocampal RIPK1, RIPK3, MLKL, and p-MLKL, inhibit excessive activation of microglia, reduce hippocampal neuroinflammation, and thus relieve depressive-like behaviors.97 Notably, copper homeostasis imbalance and copper-mediated cuproptosis are also considered potential mechanisms involved in the pathogenesis and progression of depression. Studies have confirmed that electroacupuncture at Baihui (GV20), Cuanzhu (BL2), Fenglong (ST40), and Taichong (LR3) can effectively reduce serum copper levels, accompanied by the relief of depressive symptoms.13 Given that both pathways are still in the early stages of research, future studies should further clarify their activation characteristics and crosstalk with apoptosis, ferroptosis, and autophagy to construct more targeted combined intervention strategies.
Conclusion
Depression is a multifactorial psychiatric disorder, whose pathogenesis involves the dysregulation of multiple PCD pathways. These aberrant PCD processes drive core pathological changes such as neuroinflammation, oxidative stress, and impaired synaptic plasticity, thereby forming a crucial pathophysiological basis for depression. Notably, different PCD pathways exhibit extensive crosstalk at the molecular level and can even be co-activated under specific conditions, forming a complex regulatory network. Therefore, in-depth investigation into the molecular mechanisms and interactions of PCD in depression is of great significance for unraveling the complex etiology of the disease and expanding effective therapeutic strategies. The complexity and interconnectivity of PCD in depression highlight the limitations of single-target therapies, while underscoring the unique value of the holistic perspective and multi-target therapeutic concept of TCM. Current studies have achieved substantial progress in describing how TCM monomers, formulae, and acupuncture regulate specific PCD pathways. By targeting core signaling pathways, TCM enables the precise regulation of PCD, providing abundant candidate regimens and therapeutic targets for clinical practice. However, several obvious limitations remain. First, existing studies are predominantly focused on classical pathways such as apoptosis and autophagy, whereas the regulatory mechanisms of newer forms of PCD (e.g., cuproptosis and necroptosis) in depression remain poorly understood and lack experimental validation. Research on how TCM influences these novel PCD types is also scarce. Furthermore, there is a lack of studies exploring the interactions among different PCD pathways in depression from the perspective of shared molecular mechanisms and how TCM may exert multi-target regulatory effects across these pathways. In the future, on the basis of clarifying the mechanisms of various PCD pathways and conducting balanced TCM-related research, more attention should be paid to the crosstalk between different PCD modes. Additionally, large-scale clinical studies are required to validate the diagnostic value of PCD biomarkers in patients with depression, and to further clarify the efficacy of TCM in improving patients’ clinical symptoms and quality of life through regulating these biomarkers. Second, variations in dosage, purity, and formulation exist in preclinical experiments, which affect the reproducibility and reliability of study results. Future efforts should strengthen the standardization of TCM intervention protocols to ensure the scientific rigor of research findings and their clinical applicability. Finally, current studies mainly focus on verifying the regulatory effects of TCM on PCD, with almost no “head-to-head” comparisons between TCM and conventional antidepressants in terms of their impact on PCD modulation, nor relevant analyses of their combined application. Strengthening research on the synergistic and complementary mechanisms between TCM and modern antidepressants will not only help optimize clinical treatment strategies but also provide new scientific evidence for personalized therapy and nursing interventions, thereby better improving patients’ prognosis and quality of life.
Abbreviations
ACSL4, Acyl-CoA synthetase long chain family member 4; AKT, Protein kinase B; AMPK, Adenosine 5′-monophosphate (AMP)-activated protein kinase; ASC, Apoptosis-associated speck-like protein containing a CARD; ATF4/6, Activating transcription factor 4/6; ATG12/15/7/3/10/14/16L1, Autophagy-related genes 12/15/7/3/10/14/16L1; ATP7α/β, ATPase copper transporting alpha/beta; BAK, BCL-2 homologous antagonist/killer; BAX, BCL-2-associated X protein; BCL-2, B-cell lymphoma-2; BCL-XL, B-cell lymphoma extra-large; BDNF, Brain-derived neurotrophic factor; BH3-only, BCL-2 homolog3 domain only protein; BID, BH3 interacting domain death agonist; CHOP, C/EBP-homologous protein; CREB, Cyclic adenosine monophosphate-response element binding protein; CUMS, Chronic unpredictable mild stress; Caspase, Cysteinyl aspartate specific proteinase; DAMPs, Damage-associated molecular patterns; DLAT, Dihydrolipoamide S-acetyltransferase; ERK, Extracellular regulated protein kinases; ER, Endoplasmic reticulum; ERS, Endoplasmic reticulum stress; FADD, Fas-associating protein with a novel death domain; FAS, FAS receptor; FDX1, Ferredoxin 1; Fe-S, Iron-sulfur proteins; FIP200, Focal adhesion kinase family interacting protein of 200 kD; FTH1, Ferritin heavy chain; Glu, Glutamate; GPX4, Glutathione peroxidase 4; GSH, Glutathione; GSDMD, Gasdermin D; HO-1, Heme oxygenase-1; IAPs, Inhibitors of apoptosis proteins; IL-1β/18/6/10, Interleukin-1β/18/6/10; IRE1α, Inositol-requiring enzyme 1α; LC3, Microtubule-associated protein 1 light chain 3; LOXs, Lipoxygenases; LPCAT3, Lysophosphatidylcholine acyltransferase 3; LPS, Lipopolysaccharide; MDA, Malondialdehyde; MLKL, Mixed lineage kinase domain like pseudokinase; MOMP, Mitochondrial outer membrane permeabilization; MPTP, Mitochondrial permeability transition pore; mTORC1, Mechanistic target of rapamycin complex 1; mTOR, mammalian target of rapamycin; NCOA4, Nuclear receptor coactivator 4; NF-κB, Nuclear factor kappa-B; NLRP3, NOD-like receptor family pyrin domain–containing 3; NRF2, Nuclear factor erythroid 2-related factor 2; P62, Sequestosome 1; P2X7, Purinergic receptor P2X7; PAMPs, Pathogen-associated molecular patterns; PCD, Programmed cell death; PE, Phosphatidylethanolamine; PERK, Protein kinase RNA–like endoplasmic reticulum kinase; PFC, Prefrontal cortex; PI3K, Phosphoinositide 3-kinase; PI3P, Phosphatidylinositol-3-phosphate; PRRs, Pattern recognition receptors; PUFA-PLs, Polyunsaturated fatty acid-phospholipids; PUFAs, Polyunsaturated fatty acids; PUFA-PL-OOH, Polyunsaturated fatty acid–containing phospholipid hydroperoxide; PUFA-CoA, polyunsaturated fatty acid–coenzyme A; RIPK1/3, Receptor-interacting serine/threonine-protein kinase 1/3; ROS, Reactive oxygen species; SIRT1/3, Silent information regulator sirtuin 1/3; SLC7A11, Solute carrier family 7 member 11; SLC3A2, Solute carrier family 3 member 2; SLC31A1, Solute carrier family 31 member 1; SMAC, Second mitochondria-derived activator of caspases; STEAP3, Six-transmembrane epithelial antigen of prostate 3; tBID, Truncated BID; TCM, Traditional Chinese medicine; TFR1, Transferrin receptor protein 1; TLR3/4, Toll like receptor 3/4; TNF-α, Tumor necrosis factor-α; TNFR1, Tumor necrosis factor receptor 1; TRADD, Tumor necrosis factor receptor-associated death domain protein; TRAF2/5, TNF receptor associated factor 2/5; TRAIL, TNF-related apoptosis-inducing ligand; TRAIL-R1/R2, TNF-related apoptosis-inducing ligand receptor 1/2; ULK1, UNC-51-like kinase 1; VPS34/15, Vesicular protein sorting 34/15.
Funding
This study was supported by the National Natural Science Foundation of China (82174477) and Heilongjiang Provincial Key Research and Development Program (Innovation Base) (No. JD2023SJ41).
Disclosure
The authors declare no competing interests in this work.
References
1. World Health Organization. Depressive disorder (depression). 2023. Available from: https://www.who.int/news-room/fact-sheets/detail/depression.
2. Chen Q, Huang S, Xu H, et al. The burden of mental disorders in Asian countries, 1990–2019: an analysis for the global burden of disease study 2019. Transl Psychiatry. 2024;14(1):167. doi:10.1038/s41398-024-02864-5
3. Chen S, Sun H, Song Y, et al. Transition and trend analysis of the burden of depression in China and different income countries: based on GBD database and joinpoint regression model. J Affect Disord. 2024;362:437–19. doi:10.1016/j.jad.2024.06.067
4. Cui L, Li S, Wang S, et al. Major depressive disorder: hypothesis, mechanism, prevention and treatment. Signal Transduct Target Ther. 2024;9(1):30. doi:10.1038/s41392-024-01738-y
5. Vanden Berghe T, Linkermann A, Jouan-Lanhouet S, Walczak H, Vandenabeele P. Regulated necrosis: the expanding network of non-apoptotic cell death pathways. Nat Rev Mol Cell Biol. 2014;15(2):135–147. doi:10.1038/nrm3737
6. Yuan J, Ofengeim D. A guide to cell death pathways. Nat Rev Mol Cell Biol. 2024;25(5):379–395. doi:10.1038/s41580-023-00689-6
7. Li J, Wu X, Yan S, et al. Understanding the antidepressant mechanisms of acupuncture: targeting hippocampal neuroinflammation, oxidative stress, neuroplasticity, and apoptosis in CUMS Rats. Mol Neurobiol. 2025;62(4):4221–4236. doi:10.1007/s12035-024-04550-5
8. Cao LH, Wang ZZ, Zhao H, et al. The microglial state transition as a novel mechanism by which fresh Baihe Dihuang decoction prevents depression by regulating SIRT1/HMGB1 signaling. Phytomedicine. 2025;141:156718. doi:10.1016/j.phymed.2025.156718
9. Hu Y, Zhao M, Zhao T, Qi M, Yao G, Dong Y. The protective effect of pilose antler peptide on CUMS-induced depression through AMPK/Sirt1/NF-κB/NLRP3-mediated pyroptosis. Front Pharmacol. 2022;13:815413. doi:10.3389/fphar.2022.815413
10. Cao H, Zuo C, Huang Y, et al. Hippocampal proteomic analysis reveals activation of necroptosis and ferroptosis in a mouse model of chronic unpredictable mild stress-induced depression. Behav Brain Res. 2021;407:113261. doi:10.1016/j.bbr.2021.113261
11. Zhang K, Wang F, Zhai M, et al. Hyperactive neuronal autophagy depletes BDNF and impairs adult hippocampal neurogenesis in a corticosterone-induced mouse model of depression. Theranostics. 2023;13(3):1059–1075. doi:10.7150/thno.81067
12. Wang R, Hu X, Liu S, et al. Kaempferol-3-O-sophoroside (PCS-1) contributes to modulation of depressive-like behaviour in C57BL/6J mice by activating AMPK. Br J Pharmacol. 2024;181(8):1182–1202. doi:10.1111/bph.16283
13. Zhou HH, Lu F, Chen SD, Zhou ZH, Han YZ, Hu JY. Effect of electroacupuncture on serum copper, zinc, calcium and magnesium levels in the depression rats. J Tradit Chin Med. 2011;31(2):112–114. doi:10.1016/s0254-6272(11)60023-x
14. Xu X, Yan Y, Yang Z, Zhang T. Down-regulation of RIPK3 prevents depression-like behaviors by restoring the synaptic plasticity and suppressing neuronal loss. J Affect Disord. 2024;365:213–221. doi:10.1016/j.jad.2024.08.088
15. Rosas-Sánchez GU, Germán-Ponciano LJ, Guillen-Ruiz G, et al. Neuroplasticity and mechanisms of action of acute and chronic treatment with antidepressants in preclinical studies. Biomedicines. 2024;12(12):2744. doi:10.3390/biomedicines12122744
16. Kendrick T. Strategies to reduce use of antidepressants. Br J Clin Pharmacol. 2021;87(1):23–33. doi:10.1111/bcp.14475
17. Ding W, Wang L, Li L, et al. Pathogenesis of depression and the potential for traditional Chinese medicine treatment. Front Pharmacol. 2024;15:1407869. doi:10.3389/fphar.2024.1407869
18. Wang M, Yu F, Zhang Y, Li P. Programmed cell death in tumor immunity: mechanistic insights and clinical implications. Front Immunol. 2023;14:1309635. doi:10.3389/fimmu.2023.1309635
19. Liu D, Liu Z, Hu Y, Xiong W, Wang D, Zeng Z. MOMP: a critical event in cell death regulation and anticancer treatment. Biochim Biophys Acta Rev Cancer. 2025;1880(2):189280. doi:10.1016/j.bbcan.2025.189280
20. Sever AIM, Alderson TR, Rennella E, et al. Activation of caspase-9 on the apoptosome as studied by methyl-TROSY NMR. Proc Natl Acad Sci U S A. 2023;120(51):e2310944120. doi:10.1073/pnas.2310944120
21. Du C, Fang M, Li Y, Li L, Wang X. Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition. Cell. 2000;102(1):33–42. doi:10.1016/s0092-8674(00)00008-8
22. Zhang J, Guo J, Yang N, Huang Y, Hu T, Rao C. Endoplasmic reticulum stress-mediated cell death in liver injury. Cell Death Dis. 2022;13(12):1051. doi:10.1038/s41419-022-05444-x
23. Ge C, Wang S, Wu X, Lei L. Quercetin attenuates brain apoptosis in mice with chronic unpredictable mild stress-induced depression. Behav Brain Res. 2024;465:114934. doi:10.1016/j.bbr.2024.114934
24. Zhou Z, Ye Q, Ren H, et al. CircDYM attenuates microglial apoptosis via CEBPB/ZC3H4 axis in LPS-induced mouse model of depression. Int J Biol Macromol. 2024;254(Pt 3):127922. doi:10.1016/j.ijbiomac.2023.127922
25. Yao J, Chen C, Guo Y, et al. A review of research on the association between neuron-astrocyte signaling processes and depressive symptoms. Int J Mol Sci. 2023;24(8):6985. doi:10.3390/ijms24086985
26. Shen SY, Liang LF, Shi TL, et al. Microglia-derived Interleukin-6 triggers astrocyte apoptosis in the hippocampus and mediates depression-like behavior. Adv Sci. 2025;12(11):e2412556. doi:10.1002/advs.202412556
27. Gao M, Song Y, Liu Y, Miao Y, Guo Y, Chai H. TNF-α/TNFR1 activated astrocytes exacerbate depression-like behavior in CUMS mice. Cell Death Discov. 2024;10(1):220. doi:10.1038/s41420-024-01987-4
28. Xu X, Piao HN, Aosai F, et al. Arctigenin protects against depression by inhibiting microglial activation and neuroinflammation via HMGB1/TLR4/NF-κB and TNF-α/TNFR1/NF-κB pathways. Br J Pharmacol. 2020;177(22):5224–5245. doi:10.1111/bph.15261
29. Sun KH, Jin Y, Mei ZG, et al. Antidepressant-like effects of chaihu shugan powder (柴胡疏肝散) on rats exposed to chronic unpredictable mild stress through inhibition of endoplasmic reticulum stress-induced apoptosis. Chin J Integr Med. 2021;27(5):353–360. doi:10.1007/s11655-020-3228-y
30. Rao Z, Zhu Y, Yang P, et al. Pyroptosis in inflammatory diseases and cancer. Theranostics. 2022;12(9):4310–4329. doi:10.7150/thno.71086
31. Liu Y, Pan R, Ouyang Y, et al. Pyroptosis in health and disease: mechanisms, regulation and clinical perspective. Signal Transduct Target Ther. 2024;9(1):245. doi:10.1038/s41392-024-01958-2
32. Long J, Sun Y, Liu S, et al. Targeting pyroptosis as a preventive and therapeutic approach for stroke. Cell Death Discov. 2023;9(1):155. doi:10.1038/s41420-023-01440-y
33. Yang D, He Y, Muñoz-Planillo R, Liu Q, Núñez G. Caspase-11 requires the Pannexin-1 channel and the purinergic P2X7 pore to mediate pyroptosis and endotoxic shock. Immunity. 2015;43(5):923–932. doi:10.1016/j.immuni.2015.10.009
34. Kang L, Dai J, Wang Y, et al. Blocking Caspase-1/Gsdmd and Caspase-3/-8/Gsdme pyroptotic pathways rescues silicosis in mice. PLoS Genet. 2022;18(12):e1010515. doi:10.1371/journal.pgen.1010515
35. Zhou Z, He H, Wang K, et al. Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science. 2020;368(6494):eaaz7548. doi:10.1126/science.aaz7548
36. Wang X, Su L, Liu S, et al. Paeoniflorin inhibits the activation of microglia and alleviates depressive behavior by regulating SIRT1-NF-kB-NLRP3/pyroptosis pathway. Int J Mol Sci. 2024;25(23):12543. doi:10.3390/ijms252312543
37. Bi Y, Li M, Wang Y, et al. Saikosaponins from Bupleurum scorzonerifolium Willd. alleviates microglial pyroptosis in depression by binding and inhibiting P2X7 expression. Phytomedicine. 2025;136:156240. doi:10.1016/j.phymed.2024.156240
38. Li S, Sun Y, Song M, et al. NLRP3/caspase-1/GSDMD-mediated pyroptosis exerts a crucial role in astrocyte pathological injury in mouse model of depression. JCI Insight. 2021;6(23):e146852. doi:10.1172/jci.insight.146852
39. Li Y, Li J, Yang L, et al. Ginsenoside Rb1 protects hippocampal neurons in depressed rats based on mitophagy-regulated astrocytic pyroptosis. Phytomedicine. 2023;121:155083. doi:10.1016/j.phymed.2023.155083
40. Tang M, Liu T, Jiang P, Dang R. The interaction between autophagy and neuroinflammation in major depressive disorder: from pathophysiology to therapeutic implications. Pharmacol Res. 2021;168:105586. doi:10.1016/j.phrs.2021.105586
41. Fleming A, Bourdenx M, Fujimaki M, et al. The different autophagy degradation pathways and neurodegeneration. Neuron. 2022;110(6):935–966. doi:10.1016/j.neuron.2022.01.017
42. Liu J, Wu Y, Meng S, et al. Selective autophagy in cancer: mechanisms, therapeutic implications, and future perspectives. Mol Cancer. 2024;23(1):22. doi:10.1186/s12943-024-01934-y
43. Galluzzi L, Green DR. Autophagy-Independent Functions of the Autophagy Machinery. Cell. 2019;177(7):1682–1699. doi:10.1016/j.cell.2019.05.026
44. Xu K, Wang M, Wang H, et al. HMGB1/STAT3/p65 axis drives microglial activation and autophagy exert a crucial role in chronic Stress-Induced major depressive disorder. J Adv Res. 2024;59:79–96. doi:10.1016/j.jare.2023.06.003
45. Peng S, Su P, Liu L, et al. Formononetin ameliorates depression-like behaviors through rebalancing microglia M1/M2 polarization and inhibiting NLRP3 inflammasome: involvement of activating PPARα-mediated autophagy. Mol Med. 2025;31(1):153. doi:10.1186/s10020-025-01217-2
46. Su L, Guo P, Guo X, et al. Paeoniflorin alleviates depression by inhibiting the activation of NLRP3 inflammasome via promoting mitochondrial autophagy. Chin J Nat Med. 2024;22(6):515–529. doi:10.1016/s1875-5364(24)60654-0
47. Su L, Lu H, Zhang D, et al. Total paeony glycoside relieves neuroinflammation to exert antidepressant effect via the interplay between NLRP3 inflammasome, pyroptosis and autophagy. Phytomedicine. 2024;128:155519. doi:10.1016/j.phymed.2024.155519
48. Jacquemyn J, Ralhan I, Ioannou MS. Driving factors of neuronal ferroptosis. Trends Cell Biol. 2024;34(7):535–546. doi:10.1016/j.tcb.2024.01.010
49. Liu N, Chen M. Crosstalk between ferroptosis and cuproptosis: from mechanism to potential clinical application. Biomed Pharmacother. 2024;171:116115. doi:10.1016/j.biopha.2023.116115
50. Li FJ, Long HZ, Zhou ZW, Luo HY, Xu SG, Gao LC. System X(c) (-)/GSH/GPX4 axis: an important antioxidant system for the ferroptosis in drug-resistant solid tumor therapy. Front Pharmacol. 2022;13:910292. doi:10.3389/fphar.2022.910292
51. Yang WS, SriRamaratnam R, Welsch ME, et al. Regulation of ferroptotic cancer cell death by GPX4. Cell. 2014;156(1–2):317–331. doi:10.1016/j.cell.2013.12.010
52. Zhang G, Lv S, Zhong X, et al. Ferroptosis: a new antidepressant pharmacological mechanism. Front Pharmacol. 2023;14:1339057. doi:10.3389/fphar.2023.1339057
53. Chen J, Jiang X, Gao X, et al. Ferroptosis-related genes as diagnostic markers for major depressive disorder and their correlations with immune infiltration. Front Med. 2023;10:1215180. doi:10.3389/fmed.2023.1215180
54. Wang Y, Li Y, Wu Z, et al. Ferritin disorder in the plasma and hippocampus associated with major depressive disorder. Biochem Biophys Res Commun. 2021;553:114–118. doi:10.1016/j.bbrc.2021.03.059
55. Li Z, Li B, Song X, et al. Dietary zinc and iron intake and risk of depression: a meta-analysis. Psychiatry Res. 2017;251:41–47. doi:10.1016/j.psychres.2017.02.006
56. Beretta GL, Zaffaroni N. Necroptosis and prostate cancer: molecular mechanisms and therapeutic potential. Cells. 2022;11(7):1221. doi:10.3390/cells11071221
57. Frank D, Vince JE. Pyroptosis versus necroptosis: similarities, differences, and crosstalk. Cell Death Differ. 2019;26(1):99–114. doi:10.1038/s41418-018-0212-6
58. He R, Wang Z, Dong S, et al. Understanding necroptosis in pancreatic diseases. Biomolecules. 2022;12(6):828. doi:10.3390/biom12060828
59. Tsvetkov P, Coy S, Petrova B, et al. Copper induces cell death by targeting lipoylated TCA cycle proteins. Science. 2022;375(6586):1254–1261. doi:10.1126/science.abf0529
60. Gaetke LM, Chow CK. Copper toxicity, oxidative stress, and antioxidant nutrients. Toxicology. 2003;189(1–2):147–163. doi:10.1016/s0300-483x(03)00159-8
61. Zulkifli M, Spelbring AN, Zhang Y, et al. FDX1-dependent and independent mechanisms of elesclomol-mediated intracellular copper delivery. Proc Natl Acad Sci U S A. 2023;120(10):e2216722120. doi:10.1073/pnas.2216722120
62. Chen J, Song W, Zhang W. The emerging role of copper in depression. Front Neurosci. 2023;17:1230404. doi:10.3389/fnins.2023.1230404
63. Fritsch M, Günther SD, Schwarzer R, et al. Caspase-8 is the molecular switch for apoptosis, necroptosis and pyroptosis. Nature. 2019;575(7784):683–687. doi:10.1038/s41586-019-1770-6
64. Gao W, Huang Z, Duan J, Nice EC, Lin J, Huang C. Elesclomol induces copper-dependent ferroptosis in colorectal cancer cells via degradation of ATP7A. Mol Oncol. 2021;15(12):3527–3544. doi:10.1002/1878-0261.13079
65. Tian Q, Qin B, Gu Y, et al. ROS-mediated necroptosis is involved in iron overload-induced osteoblastic cell death. Oxid Med Cell Longev. 2020;2020:1295382. doi:10.1155/2020/1295382
66. Cheng J, Fan YQ, Liu BH, Zhou H, Wang JM, Chen QX. ACSL4 suppresses glioma cells proliferation via activating ferroptosis. Oncol Rep. 2020;43(1):147–158. doi:10.3892/or.2019.7419
67. Müller T, Dewitz C, Schmitz J, et al. Necroptosis and ferroptosis are alternative cell death pathways that operate in acute kidney failure. Cell Mol Life Sci. 2017;74(19):3631–3645. doi:10.1007/s00018-017-2547-4
68. Zhou F, Yang Y, Xing D. Bcl-2 and Bcl-xL play important roles in the crosstalk between autophagy and apoptosis. FEBS J. 2011;278(3):403–413. doi:10.1111/j.1742-4658.2010.07965.x
69. Shende P, Plaisance I, Morandi C, et al. Cardiac raptor ablation impairs adaptive hypertrophy, alters metabolic gene expression, and causes heart failure in mice. Circulation. 2011;123(10):1073–1082. doi:10.1161/circulationaha.110.977066
70. Wang H, Feng X, He H, et al. Crosstalk between autophagy and other forms of programmed cell death. Eur J Pharmacol. 2025;995:177414. doi:10.1016/j.ejphar.2025.177414
71. Degterev A, Huang Z, Boyce M, et al. Chemical inhibitor of nonapoptotic cell death with therapeutic potential for ischemic brain injury. Nat Chem Biol. 2005;1(2):112–119. doi:10.1038/nchembio711
72. Wu X, Poulsen KL, Sanz-Garcia C, et al. MLKL-dependent signaling regulates autophagic flux in a murine model of non-alcohol-associated fatty liver and steatohepatitis. J Hepatol. 2020;73(3):616–627. doi:10.1016/j.jhep.2020.03.023
73. Marshall KD, Baines CP. Necroptosis: is there a role for mitochondria? Front Physiol. 2014;5:323. doi:10.3389/fphys.2014.00323
74. Liu W, Gan Y, Ding Y, et al. Autophagy promotes GSDME-mediated pyroptosis via intrinsic and extrinsic apoptotic pathways in cobalt chloride-induced hypoxia reoxygenation-acute kidney injury. Ecotoxicol Environ Saf. 2022;242:113881. doi:10.1016/j.ecoenv.2022.113881
75. Shi CS, Shenderov K, Huang NN, et al. Activation of autophagy by inflammatory signals limits IL-1β production by targeting ubiquitinated inflammasomes for destruction. Nat Immunol. 2012;13(3):255–263. doi:10.1038/ni.2215
76. Mancias JD, Wang X, Gygi SP, Harper JW, Kimmelman AC. Quantitative proteomics identifies NCOA4 as the cargo receptor mediating ferritinophagy. Nature. 2014;509(7498):105–109. doi:10.1038/nature13148
77. Zhou B, Liu J, Kang R, Klionsky DJ, Kroemer G, Tang D. Ferroptosis is a type of autophagy-dependent cell death. Semin Cancer Biol. 2020;66:89–100. doi:10.1016/j.semcancer.2019.03.002
78. Denton D, Kumar S. Autophagy-dependent cell death. Cell Death Differ. 2019;26(4):605–616. doi:10.1038/s41418-018-0252-y
79. Xue Q, Kang R, Klionsky DJ, Tang D, Liu J, Chen X. Copper metabolism in cell death and autophagy. Autophagy. 2023;19(8):2175–2195. doi:10.1080/15548627.2023.2200554
80. Tang Y, Su H, Nie K, et al. Berberine exerts antidepressant effects in vivo and in vitro through the PI3K/AKT/CREB/BDNF signaling pathway. Biomed Pharmacother. 2024;170:116012. doi:10.1016/j.biopha.2023.116012
81. Ru-Han A, Gong Q, Tuo YJ, et al. Syringa oblata Lindl extract alleviated corticosterone-induced depression via the cAMP/PKA-CREB-BDNF pathway. J Ethnopharmacol. 2025;341:119274. doi:10.1016/j.jep.2024.119274
82. Pei H, Shen H, Bi J, He Z, Zhai L. Gastrodin improves nerve cell injury and behaviors of depressed mice through Caspase-3-mediated apoptosis. CNS Neurosci Ther. 2024;30(3):e14444. doi:10.1111/cns.14444
83. Yuan M, Yuan B. Antidepressant-like effects of Rehmannioside A on rats induced by chronic unpredictable mild stress through inhibition of endoplasmic reticulum stress and apoptosis of hippocampus. J Chem Neuroanat. 2022;125:102157. doi:10.1016/j.jchemneu.2022.102157
84. Chai Y, Cai Y, Fu Y, et al. Salidroside ameliorates depression by suppressing NLRP3-mediated pyroptosis via P2X7/NF-κB/NLRP3 signaling pathway. Front Pharmacol. 2022;13:812362. doi:10.3389/fphar.2022.812362
85. Li Y, Song W, Tong Y, et al. Isoliquiritin ameliorates depression by suppressing NLRP3-mediated pyroptosis via miRNA-27a/SYK/NF-κB axis. J Neuroinflammation. 2021;18(1):1. doi:10.1186/s12974-020-02040-8
86. Chen Y, Hao C, Chen W, et al. Anti-depressant effects of acupuncture: the insights from NLRP3 mediated pyroptosis and inflammation. Neurosci Lett. 2022;785:136787. doi:10.1016/j.neulet.2022.136787
87. Liu Y, Li Y, Li J, et al. Gypenosides alleviate oxidative stress in the hippocampus, promote mitophagy, and mitigate depressive-like behaviors induced by CUMS via SIRT1. J Ethnopharmacol. 2025;337(Pt 2):118823. doi:10.1016/j.jep.2024.118823
88. Tabassum S, Misrani A, Huang HX, Zhang ZY, Li QW, Long C. Resveratrol attenuates chronic unpredictable mild stress-induced alterations in the SIRT1/PGC1α/SIRT3 pathway and associated mitochondrial dysfunction in mice. Mol Neurobiol. 2023;60(9):5102–5116. doi:10.1007/s12035-023-03395-8
89. Su P, Wu M, Yin X, et al. Modified Xiaoyao San reverses lipopolysaccharide-induced depression-like behavior through suppressing microglia M1 polarization via enhancing autophagy involved in PI3K/Akt/mTOR pathway in mice. J Ethnopharmacol. 2023;315:116659. doi:10.1016/j.jep.2023.116659
90. Zhang Z, Cai X, Yao Z, et al. EA ameliorated depressive behaviors in CUMS rats and was related to its suppressing autophagy in the hippocampus. Neural Plast. 2020;2020:8860968. doi:10.1155/2020/8860968
91. Li C, Ge H, Huang J, et al. Resveratrol alleviates depression-like behaviors by inhibiting ferroptosis via AKT/NRF2 pathway. Brain Res Bull. 2025;220:111136. doi:10.1016/j.brainresbull.2024.111136
92. Zhou A, Feng HY, Fan CN, et al. Asiaticoside attenuates chronic restraint stress-induced hippocampal CA1 neuronal ferroptosis via activating BDNF/Nrf2/GPX4 signaling pathway. Drug Des Devel Ther. 2025;19:793–810. doi:10.2147/dddt.S509208
93. He J, He M, Yang P, Shangguan J, Jiang L, Liu Z. Activation of SIRT1 by Hydroxysafflor Yellow A attenuates chronic unpredictable mild stress-induced microglia activation and iron death in depressed rats. Brain Behav. 2025;15(3):e70385. doi:10.1002/brb3.70385
94. Wang X, Li S, Yu J, et al. Saikosaponin B2 ameliorates depression-induced microglia activation by inhibiting ferroptosis-mediated neuroinflammation and ER stress. J Ethnopharmacol. 2023;316:116729. doi:10.1016/j.jep.2023.116729
95. Jiao H, Yang H, Yan Z, et al. Traditional Chinese formula xiaoyaosan alleviates depressive-like behavior in CUMS mice by regulating PEBP1-GPX4-mediated ferroptosis in the hippocampus. Neuropsychiatr Dis Treat. 2021;17:1001–1019. doi:10.2147/ndt.S302443
96. Shen J, Hao C, Yuan S, et al. Acupuncture alleviates CUMS-induced depression-like behaviors of rats by regulating oxidative stress, neuroinflammation and ferroptosis. Brain Res. 2024;1826:148715. doi:10.1016/j.brainres.2023.148715
97. Yan ZY, Jiao HY, Chen JB, et al. Antidepressant mechanism of traditional Chinese medicine formula Xiaoyaosan in CUMS-induced depressed mouse model via RIPK1-RIPK3-MLKL mediated necroptosis based on network pharmacology analysis. Front Pharmacol. 2021;12:773562. doi:10.3389/fphar.2021.773562
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Wenyang-Tianjing-Jieyu Decoction Improves Depression Rats of Kidney Yang Deficiency Pattern by Regulating T Cell Homeostasis and Inflammation Level
Zhang T, Wang J, Wang Y, He L, Lv S, Wang Y, Li W
Neuropsychiatric Disease and Treatment 2024, 20:631-647
Published Date: 22 March 2024
Epigenetics and Herbs: Potential Therapeutic Strategies for Osteoarthritis of the Knee

Zheng Y, Wang J, Wang L, Zheng Y, Qi W
Journal of Pain Research 2025, 18:3217-3261
Published Date: 26 June 2025
Traditional Chinese Medicine Ameliorates Depression via the Gut-Brain Axis: A Review Focus on NLRP3/TLR4-Mediated Inflammatory Pathways and Gut Microbiota Modulation
Che L, Xie J, Xia C, Yu Q
Neuropsychiatric Disease and Treatment 2026, 22:579710
Published Date: 8 January 2026