Back to Journals » Journal of Inflammation Research » Volume 18
Targeting PANoptosis in Bacterial-Induced Inflammatory Diseases: Mechanisms and Therapeutic Interventions
Authors Wang T, Lu Y, Zhang X ![]() , Yu F
, Yu F
Received 3 September 2025
Accepted for publication 17 November 2025
Published 27 November 2025 Volume 2025:18 Pages 16713—16728
DOI https://doi.org/10.2147/JIR.S563801
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Anh Ngo
Tianyin Wang,1 Yini Lu,1 Xiong Zhang,2 Feng Yu1
1Department of Basic Medicine and Clinical Pharmacy, China Pharmaceutical University, Nanjing, Jiangsu, 211198, People’s Republic of China; 2Department of Criminal Science and Technology, Jiangsu Police Institute, Nanjing, Jiangsu, 210031, People’s Republic of China
Correspondence: Xiong Zhang, Department of Criminal Science and Technology, Jiangsu Police Institute, Nanjing, Jiangsu, 210031, People’s Republic of China, Email [email protected] Feng Yu, Department of Basic Medicine and Clinical Pharmacy, China Pharmaceutical University, No. 639 Longmian Avenue, Jiangning District, Nanjing, Jiangsu, 211198, People’s Republic of China, Email [email protected]
Abstract: PANoptosis is a novel form of cell death that integrates necroptosis, apoptosis, and pyroptosis, along with their crosstalk, mediated and regulated by the PANoptosome. This review focuses on bacterial PANoptosis, systematically summarizing through distinct pathways and virulence factors, and analyzing its role in inflammatory diseases. Furthermore, we discuss potential therapeutic interventions related to PANoptosis in bacterial inflammation, establishing a framework for novel anti-inflammatory drug development. Understanding the mechanisms of bacteria-induced PANoptosis may provide new strategies for targeted intervention in inflammatory responses to infection.
Keywords: PANoptosis, PANoptosome, gram-positive bacteria, gram-negative bacteria, potential therapeutic targets
Introduction of PANoptosis
PANoptosis
Programmed cell death is a genetically regulated process that maintains cellular homeostasis through distinct death pathways in response to environment-specific stimuli.1 Beyond conventional death modalities, a novel programmed cell death pathway termed PANoptosis has been recognized. The PANoptosis paradigm first emerged in 2019. Researchers analyzing the functions of Z-DNA binding protein 1 (ZBP1) and transforming growth factor-β-activated kinase 1 (TAK1) molecules demonstrated that apoptosis, pyroptosis, and necroptosis could be integrated into a synergistic PANoptosis pathway, establishing that these three forms of cell death are interconnected rather than independent.2 Accumulating data revealed extensive crosstalk between these processes,3 leading to the conceptual maturation of PANoptosis. PANoptosis represents a unique programmed cell death modality that integrates characteristic elements of apoptosis, pyroptosis, and necroptosis while maintaining its own identity distinct from each constituent pathway.
PANoptosome
The PANoptosome serves as the core complex regulating PANoptosis,4 incorporating key regulatory proteins from all three death pathways to initiate downstream necroptosis, apoptosis, and pyroptosis. Currently, four distinct types of PANoptosomes have been identified: ZBP1-, absent in melanoma 2 (AIM2)-, receptor-interacting serine/threonine-protein kinase 1 (RIPK1)-, and NLR family pyrin domain containing 12 (NLRP12)-PANoptosomes, each possessing unique components and activation mechanisms.5 (1) The ZBP1-PANoptosome assembles upon ZBP1 activation through its Zα2 domain recognition of viral Z-RNA. This triggers receptor-interacting protein (RIP) homotypic interaction motif (RHIM) domain mediated homotypic interaction with RIPK3, forming the core complex.6,7 Caspase-6 reinforces this assembly by directly binding RIPK3 and enhancing RHIM-dependent ZBP1-RIPK3 engagement.8 This stabilized core complex subsequently recruits Caspase-8 and activates the NLRP3 inflammasome, thereby amplifying inflammatory signaling and triggering PANoptosis. (2) The AIM2-PANoptosome assembles when AIM2 binds pathogenic or host-derived double-stranded DNA. Through its pyrin domain, AIM2 recruits the apoptosis-associated speck-like protein containing a CARD (ASC) adaptor to form the inflammasome core and activate Caspase-1.9,10 Under specific conditions, AIM2 cooperates with pyrin and ZBP1 to form the AIM2-PANoptosome complex.11,12 This complex integrates ASC, Caspase-1, Caspase-8, RIPK3, RIPK1 and Fas-associated protein with death domain (FADD), collectively activating PANoptosis. (3) The RIPK1-PANoptosome is activated during Yersinia infection or TAK1 inhibition. Its assembly depends on the protein phosphatase 6 (PP6) phosphatase complex, with RIPK1 and RIPK3 serving as the central protein in PANoptosis regulation.13,14 The N-terminal kinase domain of RIPK1 is critical for inducing cell death.15 RIPK3 serves as an essential component of the necrosome, interacts with RIPK1 to trigger programmed necrosis, and plays a pivotal role in PANoptosis regulation.16 (4) The NLRP12-PANoptosome responds to signals including heme, tumor necrosis factor (TNF), and pathogen-associated molecular patterns (PAMPs),17 driving PANoptosis in hemolysis and inflammatory diseases through caspase-8 and RIPK3.18,19
Shared Mechanisms of PANoptosis in Gram-Positive and Gram-Negative Bacteria
Both Gram-positive and Gram-negative bacteria can trigger PANoptosis through distinct molecular mechanisms. Virulence factors from both bacterial types activate host cell death signaling pathways through different molecular mechanisms, ultimately converging on PANoptosome complex formation. This molecular scaffold integrates key components from the inflammasome (ASC, NLRP3, caspase-1), apoptosome (caspase-8), and necrosome (RIPK3), simultaneously activating caspase-3-mediated apoptosis, gasdermin D (GSDMD)-mediated pyroptosis, and mixed lineage kinase domain-like protein (MLKL)-mediated necroptosis (Figure 1).
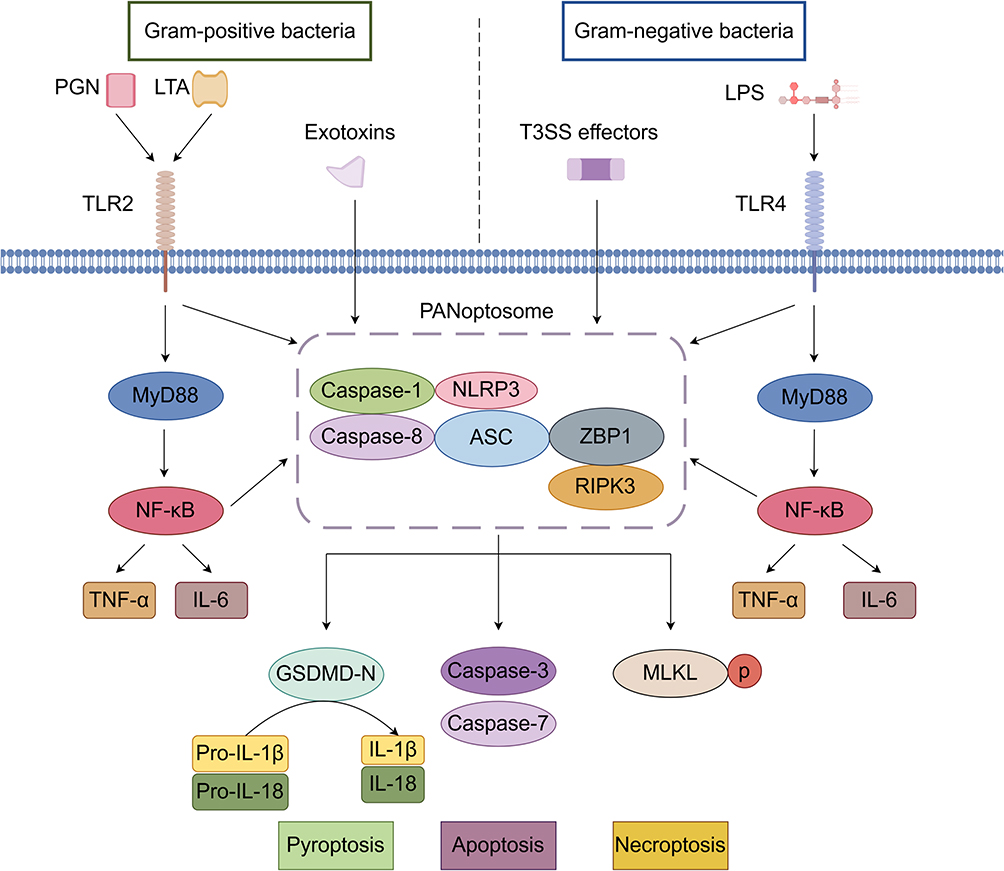 |
Figure 1 Mechanisms of PANoptosis Induction by Gram-negative and Gram-positive Bacteria. |
Bacterial Secretion Systems
The delivery of effectors mediated by bacterial secretion systems constitutes a crucial mechanism for triggering host cell PANoptosis. Predominantly found in Gram-negative bacteria, these secretion systems function to precisely inject effectors into host cells, thereby modulating host signaling pathways to facilitate bacterial invasion, survival, and immune evasion. Gram-negative bacteria employ diverse secretion systems including Type II secretion system (T2SS), T3SS, and T6SS. Notably, Pseudomonas aeruginosa utilizes the T2SS protease LasB to disrupt intercellular junctions20 while simultaneously injecting effectors via T3SS to damage mucosal barriers;21 its T6SS effector VgrG2b targets microtubule structures, with these three systems acting synergistically to enhance bacterial infection.22,23 Similarly, Escherichia coli employs T3SS to inject effectors into the host cytoplasm, inducing impairment of intestinal epithelial tight junctions.24
Toll-Like Receptors (TLRs)
TLRs, a family of pattern recognition receptors (PRRs) in the innate immune system, mediate early pathogen detection.25 While playing pivotal roles in antibacterial immune responses, TLRs exhibit distinct recognition mechanisms and immunoregulatory functions toward Gram-negative versus Gram-positive bacteria. For Gram-negative bacteria, TLR4 serves as the primary receptor that initiates myeloid differentiation primary response 88 (MyD88)-dependent signaling upon lipopolysaccharide (LPS) binding, eliciting robust inflammatory responses, which may progress to cytokine storms when hyperactivated.26 In contrast, Gram-positive bacteria are predominantly recognized by TLR2 through specific components such as lipoproteins and peptidoglycan. For instance, TLR2-mediated detection of Staphylococcus aureus enhances neutrophil phagocytic activity to combat infection,27 whereas excessive TLR2 recognition of Clostridioides difficile toxins may exacerbate tissue damage.28
PAMPs
PRRs detect PAMPs or damage-associated molecular patterns (DAMPs), triggering signaling cascades that initiate PANoptosis.2,4 In Gram-positive bacteria, the predominant PAMPs are peptidoglycan (PGN) and lipoteichoic acid (LTA), which are exposed on the bacterial surface29–31 and activate inflammatory pathways through TLR2 or nucleotide-binding oligomerization domain (NOD)-like receptors. The secreted exotoxins from these bacteria serve as major effector molecules that can activate inflammasomes and trigger PANoptosis. The primary PAMP of Gram-negative bacteria is LPS, which is expressed on the bacterial surface. LPS activates inflammasomes through TLR4,29,30,32 inducing robust inflammatory responses that subsequently promote PANoptosis.
Exemplars of PANoptosis in Bacteria
Gram-Positive Cocci
Staphylococcus aureus (S. aureus)
S. aureus dynamically modulates host cell PANoptosis through multiple virulence factors. Regarding apoptosis, S. aureus secreted alpha-toxin (α-toxin) and staphylococcal enterotoxins (SEs) directly trigger apoptosis.33 Its extracellular vesicles (EVs) invade cells via lipid raft-mediated endocytosis, inducing mitochondrial damage and activating mitogen-activated protein kinase (MAPK) and nuclear factor-κB (NF-κB) pathways to promote apoptosis.34 Conversely, this pathogen can also suppress apoptosis by inhibiting caspase-8 to facilitate immune evasion,35 or disrupt the crosstalk between neutrophil autophagy and apoptosis to sustain infection.36 In pyroptotic regulation, the pore-forming toxin Panton-Valentine leukocidin (PVL) binds the LukS-PV subunit on monocytes and macrophages to activate the NLRP3 inflammasome, driving caspase-1-dependent interleukin-1 beta (IL-1β) and IL-18 release.37 While LTA activates caspase-11 and caspase-1 via NLRP6 to regulate IL-18 secretion, demonstrating inflammasome synergy.38 Regarding necroptosis, toxin-mediated activation of RIPK1/RIPK3/MLKL signaling is the key mechanism,39 and AIM2 inflammasome-mediated macrophage necroptosis enhances bacterial pathogenicity.40 This multi-pathway cell death modulation reveals S. aureus infection strategies: either promoting PANoptosis to facilitate toxin dissemination or suppressing PANoptosis to evade host clearance.
Streptococcus pneumoniae (S. pneumoniae)
S. pneumoniae modulates host cell PANoptosis through multiple pore-forming toxins. In apoptotic regulation, S. pneumoniae cholesterol-dependent cytolysins induce apoptosis in neurons and epithelial cells.41 In chronic obstructive pulmonary disease (COPD) patients, S. pneumoniae triggers macrophage apoptosis and reactive oxygen species (ROS) release;42 whereas alveolar macrophage caspase-dependent apoptosis associated with ROS and nitric oxide (NO) production is crucial for bacterial clearance.43 This differential regulation demonstrates that S. pneumoniae-induced apoptosis disrupts tissue barriers, while macrophage apoptosis facilitates bacterial clearance and inflammation control. Pyroptosis is primarily activated by pneumolysin (PLY) through NIMA-related kinase 7 (NEK7)–NLRP3 inflammasome assembly, leading to caspase-dependent IL-1β maturation.44 Different inflammasomes exhibit synergistic defense: NLRP6-deficient mice show reduced pyroptosis and higher survival rates than wild-type mice during S. pneumoniae infection,45 while AIM2-deficiency impairs caspase-1 activation and IL-1β secretion, increasing infection susceptibility.46 Regarding necroptosis, hemolysins and other pore-forming toxins activate both the typical RIPK1/RIPK3/MLKL pathway47 and direct ion dysregulation.48 RIPK3 forms complexes with RIPK1, MLKL, and mitochondrial calcium uniporter (MCU) to stimulate Ca2+ uptake into mitochondria and subsequent mitochondrial ROS (mROS) production in S. pneumoniae infection, promoting bacterial clearance.49 However, aberrant necroptosis with low NF-κB activation exacerbates lung injury.50 This dual role makes PANoptosis both a defensive weapon and potential contributor to infection pathology.
Gram-Positive Bacilli
Bacillus anthracis (B. anthracis)
B. anthracis exerts its pathogenic effects primarily through lethal toxin (LeTx)-mediated modulation of PANoptosis during infection. LeTx activates the NLRP3 inflammasome via TNF signaling, with caspase-8/RIPK3-deficient mice demonstrating enhanced resistance to infection, confirming this pathway’s significance.51 Simultaneously, LeTx specifically activates NLRP1b and NLR family CARD domain containing 4 (NLRC4) inflammasomes, exhibiting dual functionality: it recruits FADD through ASC to trigger caspase-8-dependent apoptosis52 while also inducing caspase-1-dependent pyroptosis and IL-1β release through potassium efflux.53,54 Regarding necroptosis, LeTx enhances TNF-α-induced intestinal epithelial cell death by suppressing p38α MAPK, concurrently activating both caspase-dependent apoptosis and RIPK3/MLKL-mediated necroptosis.55 These findings reveal that B. anthracis orchestrates cell death through multifaceted regulation involving toxins and inflammasomes, simultaneously promoting pathogen dissemination and exacerbating tissue damage.
Listeria monocytogenes (L. monocytogenes)
L. monocytogenes primarily induces host cell death through its key virulence factor listeriolysin O (LLO). In lymphocytes, LLO directly triggers dose-dependent apoptosis and enhances host susceptibility.56 In glial cells, L. monocytogenes induces apoptosis, while inhibition of bacterial ROS further promotes apoptotic cell death.57 Regarding pyroptosis, in macrophages, L. monocytogenes promotes the interaction between Nek7 and NLRP3 through the c-Jun N-terminal kinase (JNK) signaling pathway, driving caspase-1-dependent pyroptosis.58 Bacterial DNA released during lysis triggers AIM2 inflammasome-dependent pyroptosis.59 Depletion of Mint3 promotes pyroptosis in host macrophages and prevents L. monocytogenes infection spread, indicating that Mint3 deficiency enhances host defense.60 Furthermore, in necroptosis regulation, the RIPK3-MLKL pathway exhibits dual modulation: it can clear infection through classical necroptosis while directly inhibiting intracellular bacterial replication in a cell death-independent manner.61,62
Other Gram-Positive Bacteria
Gram-positive bacteria with specialized morphologies play crucial roles in host–pathogen interactions by modulating programmed cell death. Actinomyces species activate the NLRP3 inflammasome through TLR2-dependent mechanisms, inducing macrophage pyroptosis characterized by GSDMD cleavage and IL-1β/IL-18 release.63 Nocardia produces heme-binding protein (HBP) that targets mitochondria to trigger caspase-3-dependent apoptosis.64 These bacteria display biphasic apoptotic control during macrophage infection: first activating extrinsic and intrinsic pathways with increased caspase-8/9 expression, then suppressing apoptosis.65 Streptomyces-derived secondary metabolites can simultaneously induce tumor cell apoptosis and necroptosis.66
Gram-Negative Cocci
Neisseria
Neisseria gonorrhoeae exhibits cell type-specific regulation of apoptosis, delivering outer membrane protein PorB to mitochondria via outer membrane vesicles (OMVs) to induce THP-1 macrophage apoptosis67 while suppressing apoptosis in U937 cells and primary human macrophages.68 In polymorphonuclear leukocytes (PMNs), it inhibits caspase-3/7/9 to prolong PMN survival and promote bacterial dissemination.69,70 For pyroptosis, this pathogen activates both canonical (caspase-1) and non-canonical (caspase-4) pathways through TLR2-NLRP3 signaling, triggering GSDMD-dependent pyroptosis with IL-1β/IL-18 release; NLRP3 deficiency enhances macrophage bactericidal activity, suggesting that pyroptosis may compromise host defense.71,72 Neisseria meningitidis shows serotype-dependent apoptotic regulation: serogroup W induces stronger apoptosis in human epithelial cells and murine immune cells than serogroup Y, correlating with higher bacteremia levels.73 PorB selectively inhibits intrinsic apoptosis to maintain intracellular survival.74 Pyroptosis depends on the HrpA/HrpB two-partner secretion system (TPS) activating caspase-1/11 to cleave GSDMD, accelerating cerebral dissemination via IL-1β/IL-18 release.75 HrpA interaction with neuronal dynein light-chain, Tctex-type 1 balances apoptosis and pyroptosis,76 while lipooligosaccharide (LOS) coordinates inflammation through TLR4-mediated NF-κB activation and inflammasome assembly.77
Moraxella catarrhalis (M. catarrhalis)
M. catarrhalis activates caspase-4/11-GSDMD-dependent pyroptosis through its LOS and OMVs, a process requiring type I interferon (IFN-I) signaling and guanylate-binding proteins (GBPs) with involvement of NLRP3 inflammasome activation.78 In lung epithelial cells, M. catarrhalis triggers the mitochondrial apoptotic pathway, characterized by caspase-3/6/9 activation, decreased B-cell lymphoma 2 (Bcl-2) expression, and Bcl-2-associated X protein (Bax) translocation.79 Notably, while this bacterium activates B-cell proliferation and surface marker expression, it does not induce B-cell apoptosis.80 Furthermore, in severe asthma patients, M. catarrhalis infection correlates with neutrophil extracellular trap pathway (NETosis) formation alongside activation of MAPK, NF-κB, and necroptosis signaling pathways.81
Gram-Negative Bacilli
Escherichia coli (E. coli)
E. coli regulates host cell PANoptosis through strain-specific mechanisms. Multidrug-resistant strains simultaneously activate apoptotic caspases-3/7/8, pyroptotic caspase-1/GSDMD, and necroptotic RIPK3/MLKL pathways, with amygdalin and chlorogenic acid mitigating this process by inhibiting PANoptosis core proteins.82,83 In apoptosis, enteroaggregative E. coli induces intestinal epithelial cell apoptosis through both Bax/Bak upregulation and caspase-8 activation,84 causing DNA fragmentation and cell cycle arrest.85 Enterotoxigenic E. coli (ETEC) demonstrates selective regulation in piglet models by suppressing caspase-9 while activating caspase-8.86 For pyroptosis, ETEC K88 and mastitis-associated strains trigger canonical pyroptosis via NLRP3/caspase-1 axis,87–89 exacerbating inflammation. Whereas, enteropathogenic E. coli (EPEC) activates non-canonical pyroptosis through T3SS-dependent caspase-4 activation.90 GSDMB binding to caspase-4 promotes non-canonical pyroptosis, while caspase-7 creates negative feedback through GSDMB cleavage.91 Regarding necroptosis, extraintestinal pathogenic E. coli enhances RIPK1/MLKL signaling via outer membrane protein TolC,92 reversible by RIPK1 inhibitor necrostatin-1 (Nec-1).93 Whereas EPEC degrades RIPK1, RIPK3, TIR-domain-containing adapter-inducing interferon-β, and ZBP1 through T3SS to actively suppress necroptosis for immune evasion.94
Salmonella spp.
Salmonella Typhimurium induces apoptosis, pyroptosis, and necroptosis in host cells. This pathogen triggers both apoptosis and necroptosis during Caco-2 cell invasion while promoting the release of proinflammatory cytokines IL-8 and TNF-α.95 Additionally, Salmonella drives pyroptosis through inflammasome pathway activation, which worsens gut inflammation. A novel heteropolysaccharide (EPS 7–4) inhibits Salmonella Typhimurium-induced pyroptosis by blocking ASC oligomerization, helping to reduce intestinal damage.96 These findings reveal how Salmonella differentially regulates cell death pathways to promote infection, offering new perspectives for targeted therapies.
Yersinia spp.
Yersinia modulates host cell death programs through multifaceted mechanisms. In PANoptosis regulation, these pathogens trigger RIPK1-dependent PANoptosome assembly, coordinately activating caspase-1, caspase-8, and MLKL pathways.13 RIPK1 plays a central regulatory role, where its deficiency suppresses pyroptosis and apoptosis while enhancing necroptosis.13 The Yersinia effector protein YopJ induces caspase-8-dependent GSDMD cleavage, driving non-canonical pyroptosis,97 with RIPK1 autophosphorylation at T169 being critical for this process.98 Metabolically, Yersinia pseudotuberculosis activates AMP-activated protein kinase (AMPK) through glucose depletion, promoting RIPK1 phosphorylation at S321 to inhibit pyroptosis.99 Lysosomal supercomplexes precisely regulate pyroptosis intensity by recruiting RIPK1-caspase-8 assemblies.100 This cell death regulation exhibits species specificity. Yersinia-infected murine macrophages undergo extrinsic apoptosis via the RIPK1-caspase-8 axis, while human macrophages trigger RIPK1-independent apoptosis through inhibitor of kappa B kinase (IKK) inhibition.101
Pseudomonas aeruginosa (P. aeruginosa)
In P. aeruginosa-induced PANoptosis, macrophage cell death can only be completely blocked when caspase-1, −11, −8 and RIPK3 are simultaneously absent.102 The RIPK1/MLKL pathway shows compensatory enhancement when neuronal apoptosis inhibitory protein 5 and NLRC4 are deficient.102 Regarding apoptosis, the T3SS effector ExoT activates JNK1/2 signaling through its GTPase-activating protein (GAP) domain, inducing caspase-9/caspase-3-mediated apoptosis,103 while exotoxin A markedly increases caspase-3 activity to promote hemocyte apoptosis.104 In pyroptosis regulation, flagellin activates the NLRC4 inflammasome, triggering caspase-1/GSDMD-dependent pyroptosis in macrophages and neutrophils.105,106 In keratitis models, caspase-4/5/11-mediated non-canonical pyroptosis causes corneal cell death.107 For necroptosis, P. aeruginosa employs quorum sensing-mediated precise control, where rhl subsystem deficiency enhances RIPK3/MLKL-dependent necroptosis.108 During respiratory infection, CYP450 activation triggers club cell necroptosis which disrupts epithelial barriers, with Nec-1 showing reversal effects.109 These mechanisms collectively represent P. aeruginosa’s multifaceted strategies to breach host defenses.
Other Gram-Negative Bacteria
Vibrio species, as a major group of Gram-negative bacteria, modulate host PANoptosis through diverse virulence mechanisms. Vibrio alginolyticus triggers caspase-8/-9/-3-dependent apoptosis and cytolysis in fish cells via T3SS.110 Vibrio proteolyticus hemolysin VPRH activates caspase-1 by NLRP3 inflammasome, inducing IL-1β release and pyroptosis.111 Vibrio fluvialis hemolysin activates the NLRP3-caspase-1-GSDMD axis via K+ efflux and Ca2+ influx.112 For necroptosis, Vibrio vulnificus cytolysin upregulates pMLKL to drive necroptosis, which resveratrol can ameliorate in sepsis models.113
PANoptosis in Bacterial Inflammatory Diseases
Sepsis
Sepsis is defined as life-threatening organ dysfunction caused by a dysregulated host response to infection.114 This condition represents a major global health challenge, with annual estimates of 31.5 million cases and 5 million deaths worldwide,115 prompting the World Health Organization to designate sepsis as a global health priority.116
Analysis of PANoptosis-related genes (PRGs) and associated immune signatures in sepsis datasets reveals dysregulated PRG expression in patients, highlighting the complex interplay between sepsis and PANoptosis.117 Our previous studies also revealed that severe apoptosis occurs in the liver, kidney, and other tissues during the progression of sepsis.118 Proinflammatory cytokines play pivotal roles in sepsis-associated PANoptosis. Combined TNF-α and IFN-γ treatment induces PANoptosis, while neutralizing antibodies against these cytokines suppresses PANoptosis progression and improves survival in septic mice.119
Addressing sepsis-induced organ dysfunction remains a critical challenge, with distinct PANoptosis activation patterns across organs. In cardiac tissue, Xiao Chai Hu Tang exerts protective effects by suppressing ZBP1-mediated PANoptosis, whereas ZBP1 overexpression exacerbates cardiomyocyte death.120 During renal injury, AIM2 expression is significantly upregulated and promotes PANoptosis in renal tubular epithelial cells through eukaryotic translation initiation factor 2-alpha kinase 2 (EIF2AK2) regulation.121 In the lungs, which represent the most frequently affected organ in sepsis, the ZBP1 and RIPK1 play a key role in pulmonary epithelial PANoptosis.122 Lactic acidosis aggravates ZBP1-dependent PANoptosis in pulmonary vascular endothelial cells during acute lung injury.123 Collectively, these findings demonstrate marked organ-specificity in PANoptosis activation during sepsis: ZBP1 predominantly mediates cardiac and pulmonary injury, while AIM2 plays a more prominent role in renal damage. This organ-specific pattern provides a theoretical basis for developing targeted organ-protective strategies.
Lung Diseases
Community-acquired pneumonia (CAP) represents the most common infectious cause of adult hospitalizations and deaths in the United States. A substantial proportion of CAP cases progress to sepsis and acute respiratory distress syndrome (ARDS).124 Pneumonia serves as the primary infectious trigger for ARDS.125 As a frequent cause of respiratory failure in critically ill patients, ARDS meets diagnostic criteria in approximately 10% of global intensive care unit admissions, with mortality rates reaching 30–40%.126
Abnormal PANoptosis activation is closely associated with bacterial pneumonia progression, involving coordinated regulation of multiple signaling pathways. Transcriptomic analysis reveals significantly altered expression of PANoptosis signature genes including zinc finger protein 304 (ZNF304), AKT serine/threonine kinase 3 (AKT3), MAPK8, and Rho GTPase activating protein 10 (ARHGAP10) in CAP patients, indicating their biomarker and therapeutic potential.127 During pathogen infection, IFN-γ is a key PANoptosis regulator. In Pasteurella multocida toxin (PMT)-induced pneumonia models, IFN-γ deficiency significantly reduces expression of executioner proteins including caspase-3, GSDMD and MLKL, while decreasing lung myeloperoxidase activity and IL-1β release, thereby alleviating lung injury.128
In pulmonary diseases, PANoptosis activation involves coordinated regulation through multiple pathways. The stimulator of interferon genes (STING) agonist activates NF-κB to promote IFN-I and pro-inflammatory cytokine production, inducing ZBP1-dependent PANoptosis.129 Overactivation of the cyclic GMP-AMP synthase (cGAS)-STING pathway promotes PANoptosome assembly, whereas the kaempferol-3-O-α-L-(4″-E-p-coumaroyl)-rhamnoside (KAE) significantly attenuates lung injury by inhibiting this pathway.130 MiR-29a-3p targets ZBP1 expression, substantially reducing levels of key PANoptosis execution proteins including Caspase-3, Caspase-8, GSDMD, and MLKL in alveolar epithelial cells.131 Baicalin effectively suppresses PANoptosis in macrophages by blocking mitochondrial Z-DNA formation and ZBP1-PANoptosome assembly.132 Collectively, these studies demonstrate that PANoptosis in lung tissue is regulated by multiple signaling nodes including STING, cGAS, and ZBP1, suggesting that targeting these key nodes may represent a promising therapeutic strategy for bacterial pneumonia.
Gastrointestinal Diseases
The global epidemiological evolution of inflammatory bowel disease (IBD) shows developing countries in the emerging stage, while newly industrialized nations are experiencing accelerating incidence.133 Concurrently, Clostridium difficile infection (CDI), an important bacterial gastrointestinal disease, remains primarily hospital-associated but shows significantly increasing community-associated cases affecting younger, healthier populations.134
Bacterial infections contribute to gastrointestinal disease pathogenesis by modulating PANoptosis in intestinal epithelial cells (IECs). Studies reveal significant PANoptosis activation in ulcerative colitis (UC) patients’ IECs, characterized by upregulated key genes including ZBP1, AIM2, caspase-1/8, and interferon regulatory factor 1 (IRF1), where IRF1 potentially acts as a transcription factor promoting PANoptosome complex formation and subsequent intestinal barrier disruption.135 In IBD, PANoptosis signature genes strongly correlate with TNF, NF-κB signaling, and immune cell infiltration.136 IBD-associated mitochondrial dysfunction and mtROS accumulation exacerbate this process, while selenium-modified exosomes can suppress PANoptosis by scavenging mtROS.137 Mechanistically, the Salmonella effector protein SopF inhibits caspase-8 via phosphoinositide-dependent protein kinase-1 (PDK1)-p90 ribosomal S6 kinase signaling, blocking apoptosis and pyroptosis while promoting necroptosis to facilitate bacterial immune evasion.138 Colitis mice exhibit elevated expression of PANoptosis-related genes including ZBP1 and caspase-1, which the natural compound diosmin can reverse while maintaining epithelial barrier function and modulating gut microbiota, demonstrating therapeutic potential for colitis.139 These findings illustrate how bacteria activate PANoptosis either directly by hijacking cell death programs or indirectly through dysbiosis, ultimately compromising intestinal barrier integrity, with targeting PANoptosis key components representing a promising strategy against bacterial gastrointestinal infections.
Potential Therapeutic Targets and Drugs for PANoptosis in Bacterial Inflammatory Diseases
Bacterial infection-induced inflammatory diseases can activate PANoptosis through multiple mechanisms, with their molecular targets and intervention strategies emerging as research priorities. At the target level, PRR pathways, cell death regulatory nodes, and immune modulators constitute core components of PANoptosis regulation. Potential therapeutic agents can be categorized into three classes: chemical drugs, natural extracts, and traditional Chinese medicine formulations, which differentially modulate inflammatory factors and cell death execution proteins to provide novel therapeutic possibilities for conditions like sepsis and acute lung injury. The following tables systematically summarize the mechanisms of relevant targets and potential drugs (Tables 1 and 2).
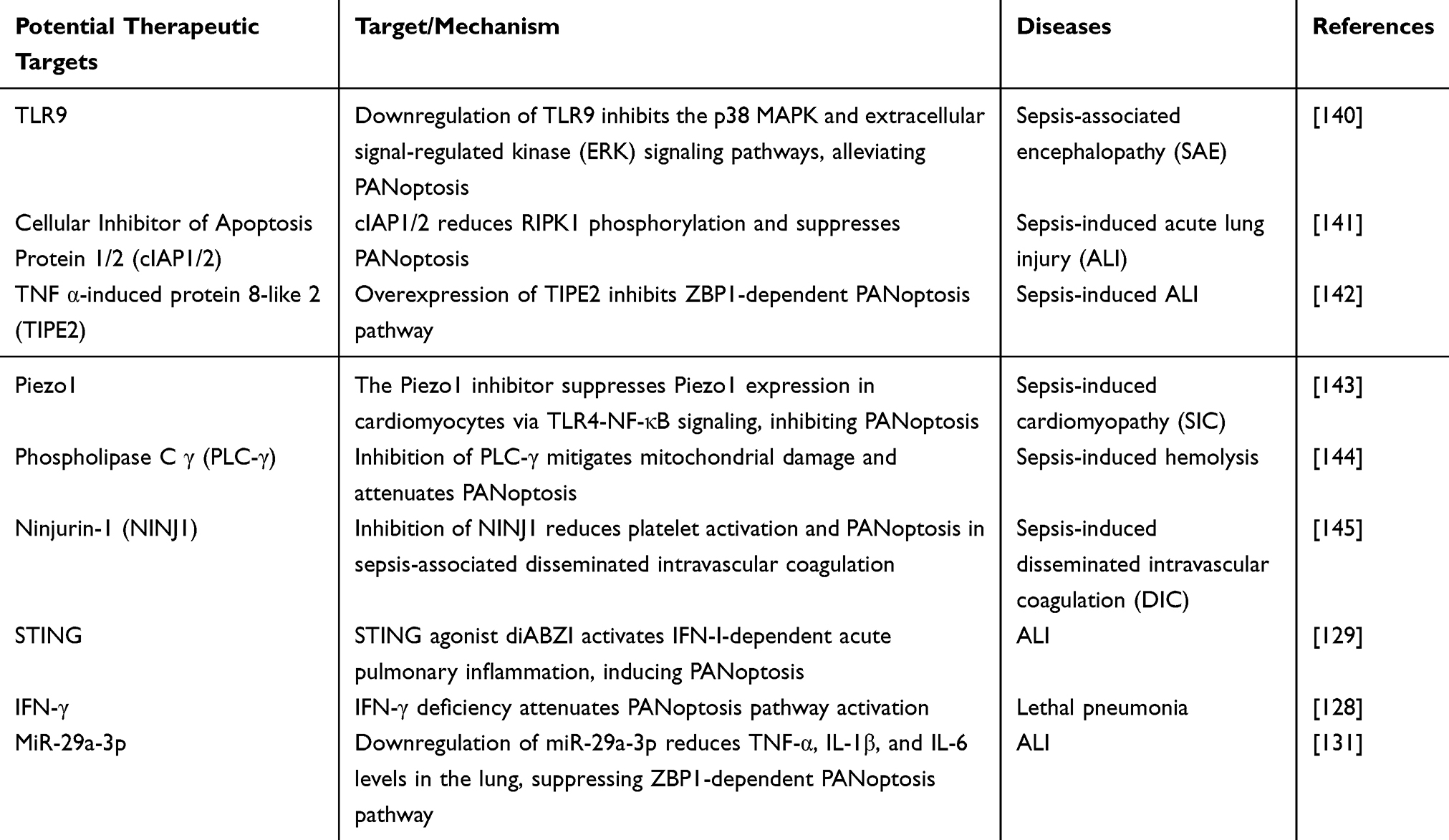 |
Table 1 Potential Therapeutic Targets for PANoptosis in Bacterial Inflammatory Diseases |
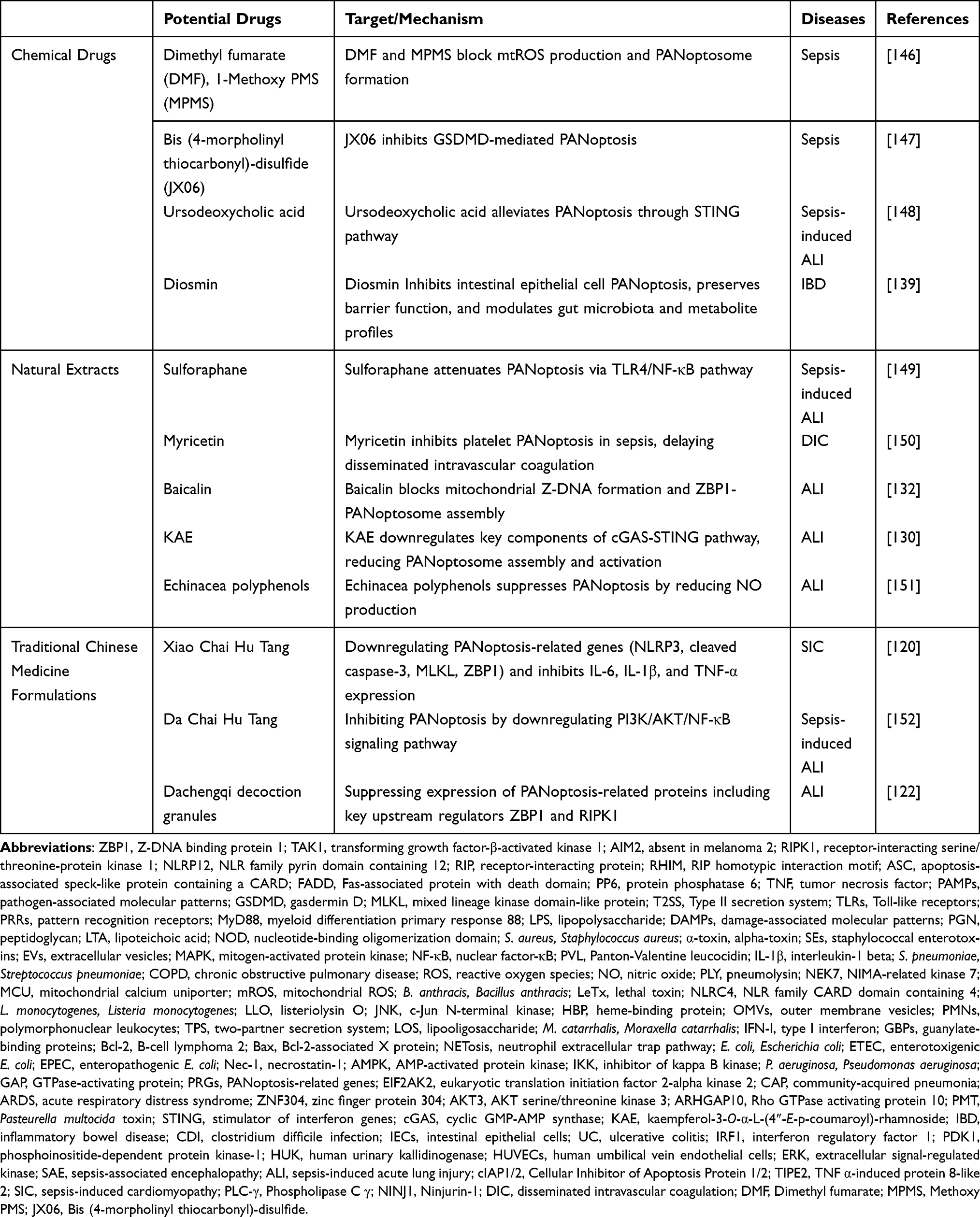 |
Table 2 Potential Therapeutic Drugs for PANoptosis in Bacterial Inflammatory Diseases |
Discussion
PANoptosis, as an inflammatory programmed cell death pathway that integrates pyroptosis, apoptosis, and necroptosis, forms a sophisticated PANoptosome complex. ZBP1-PANoptosome activates by binding RIPK3 through its Zα2 domain. AIM2-PANoptosome detects double-stranded DNA and integrates pyrin with ZBP1. RIPK1-PANoptosome activates during Yersinia infection or TAK1 inhibition. NLRP12-PANoptosome responds to heme and TNF signals. These complexes possess distinct characteristic components while sharing common elements including caspase-8, RIPK3, and ASC, creating molecular crosstalk that synchronizes multiple cell death programs. Bacterial pathogens engage this network through their characteristic signatures. Gram-positive bacteria primarily activate TLR2/NOD pathways, while Gram-negative bacteria activate LPS-TLR4 signaling. Despite their different starting points, all these pathways eventually lead to activation of the core PANoptosis process.
Currently, the primary challenge in PANoptosis-targeted therapy lies in the lack of specific inhibitors. Existing drugs mainly exhibit two limitations: (1) Uncontrollability of single-target agents. Certain inhibitors like necroptosis inhibitor Nec-1 only act on a single node of one cell death pathway. The crosstalk among cell death pathways enables mutual regulation through negative feedback mechanisms. Disruption of this dynamic balance may trigger abnormal activation or suppression of compensatory pathways, thereby increasing therapeutic unpredictability and potential risks. (2) Nonspecificity of multi-target drugs. Some natural extracts or traditional Chinese medicine components exhibit broad-spectrum regulatory effects. For example, our previous studies demonstrated that human urinary kallidinogenase (HUK), a natural substance extracted from human urine, can inhibit PANoptosis-related apoptosis and necroptosis, thereby protecting damaged human umbilical vein endothelial cells (HUVECs). It also exerts protective effects by modulating cell migration and oxidative stress.153 Their polypharmacology may cause systemic effects, making it difficult to precisely distinguish direct actions on PANoptosis, while failing to achieve personalized intervention for different bacterial infectious diseases. This highlights the urgent need to develop highly selective PANoptosis modulators through computational modeling and biological approaches to identify specific binding domains that balance efficacy and safety.
As an integral part of host defense mechanisms, complete inhibition of PANoptosis may impair immune clearance functions, potentially increasing risks of persistent bacterial infection or dissemination. Therefore, avoiding complete PANoptosis suppression while preventing infection risks remains a critical challenge. Future studies could explore targeted modulation strategies rather than complete blockade, such as locally selective PANoptosis inhibition at inflammatory foci to improve overall infection outcomes. Additionally, combined antibiotic therapy may achieve sequential bactericidal and anti-inflammatory effects. Antibiotics first eliminate pathogens to reduce infection burden, followed by precise PANoptosis modulator intervention on overactivated cell death pathways, thereby controlling inflammatory damage while preserving host defense functions.
By systematically elucidating the mechanisms through which Gram-negative and Gram-positive bacteria activate PANoptosis via distinct pathways, and summarizing PANoptosis occurrence in bacterial inflammatory diseases along with therapeutic drugs and targets, this review aims to create more possibilities for precision medicine targeting PANoptosis.
Data Sharing Statement
No new data were generated for this review. All discussed information is based on previously published studies cited in the references.
Acknowledgments
Figure 1 was created with Figdraw (www.figdraw.com).
Author Contributions
Wang T: Conceptualization, Writing – original draft, Writing – review & editing. Lu Y: Conceptualization, Formal analysis, Writing – review & editing. Zhang X and Yu F: Conceptualization, Supervision, Writing – review & editing. All authors took part in drafting, revising or critically reviewing the article, gave final approval of the version to be published and agreed on the journal for submission, accepted full accountability for the content.
Funding
This work was supported by The Hospital Pharmaceutical Research Fundation of Nanjing Pharmaceutical Association – Changzhou Siyao (grant number: 2024YX011).
Disclosure
The authors declare no known competing financial or personal interests that could influence this work.
References
1. Yuan J, Ofengeim D. A guide to cell death pathways. Nat Rev Mol Cell Biol. 2023;25(5):379–395. doi:10.1038/s41580-023-00689-6
2. Malireddi RKS, Kesavardhana S, Kanneganti T-D. ZBP1 and TAK1: master regulators of NLRP3 inflammasome/pyroptosis, apoptosis, and necroptosis (PAN-optosis). Front Cell Infect Microbiol. 2019;9. doi:10.3389/fcimb.2019.00406
3. Hughes SA, Lin M, Weir A, et al. Caspase‐8‐driven apoptotic and pyroptotic crosstalk causes cell death and IL‐1β release in X‐linked inhibitor of apoptosis (XIAP) deficiency. EMBO J. 2023;42(5). doi:10.15252/embj.2021110468
4. Christgen S, Zheng M, Kesavardhana S, et al. Identification of the PANoptosome: a molecular platform triggering pyroptosis, apoptosis, and necroptosis (PANoptosis). Front Cell Infect Microbiol. 2020. doi:10.3389/fcimb.2020.00237
5. Pandeya A, Kanneganti T-D. Therapeutic potential of PANoptosis: innate sensors, inflammasomes, and RIPKs in PANoptosomes. Trends Mol Med. 2024;30(1):74–88. doi:10.1016/j.molmed.2023.10.001
6. Kesavardhana S, Malireddi RKS, Burton AR, et al. The Zα2 domain of ZBP1 is a molecular switch regulating influenza-induced PANoptosis and perinatal lethality during development. J Biol Chem. 2020;295(24):8325–8330. doi:10.1074/jbc.RA120.013752
7. Karki R, Sundaram B, Sharma BR, et al. ADAR1 restricts ZBP1-mediated immune response and PANoptosis to promote tumorigenesis. Cell Rep. 2021;37(3). doi:10.1016/j.celrep.2021.109858
8. Zheng M, Karki R, Vogel P, Kanneganti T-D. Caspase-6 is a key regulator of innate immunity, inflammasome activation, and host defense. Cell. 2020;181(3):674–687.e13. doi:10.1016/j.cell.2020.03.040
9. Man SM, Karki R, Malireddi RK, et al. The transcription factor IRF1 and guanylate-binding proteins target activation of the AIM2 inflammasome by Francisella infection. Nat Immunol. 2015;16(5):467–475. doi:10.1038/ni.3118
10. Gao H, Wang L, Lyu Y, et al. The P2X7R/NLRP3 inflammasome axis suppresses enthesis regeneration through inflammatory and metabolic macrophage-stem cell cross-talk. Sci Adv. 2025;11(17):eadr4894. doi:10.1126/sciadv.adr4894
11. Lee S, Karki R, Wang Y, Nguyen LN, Kalathur RC, Kanneganti T-D. AIM2 forms a complex with pyrin and ZBP1 to drive PANoptosis and host defence. Nature. 2021;597(7876):415–419. doi:10.1038/s41586-021-03875-8
12. Ma C, Zhou X, Pan S, Liu L. AIM2 mediated neuron PANoptosis plays an important role in diabetes cognitive dysfunction. Behav Brain Res. 2025;491:115651. doi:10.1016/j.bbr.2025.115651
13. Malireddi RKS, Kesavardhana S, Karki R, Kancharana B, Burton AR, Kanneganti T-D. RIPK1 distinctly regulates yersinia-induced inflammatory cell death, PANoptosis. Immunohorizons. 2020;4(12):789–796. doi:10.4049/immunohorizons.2000097
14. Bynigeri RR, Malireddi RKS, Mall R, Connelly JP, Pruett-Miller SM, Kanneganti T-D. The protein phosphatase PP6 promotes RIPK1-dependent PANoptosis. BMC Biol. 2024;22(1). doi:10.1186/s12915-024-01901-5
15. Yao K, Shi Z, Zhao F, et al. RIPK1 in necroptosis and recent progress in related pharmaceutics. Front Immunol. 2025;16:1480027. doi:10.3389/fimmu.2025.1480027
16. Zhou Y, Xiang Y, Liu S, et al. RIPK3 signaling and its role in regulated cell death and diseases. Cell Death Discov. 2024;10(1):200. doi:10.1038/s41420-024-01957-w
17. Henkel FDR, O’Neill LAJ. NLRP12 drives PANoptosis in response to heme. Trends Immunol. 2023;44(8):574–576. doi:10.1016/j.it.2023.06.008
18. Sundaram B, Pandian N, Mall R, et al. NLRP12-PANoptosome activates PANoptosis and pathology in response to heme and PAMPs. Cell. 2023;186(13):2783–2801.e20. doi:10.1016/j.cell.2023.05.005
19. Nadendla EK, Alluri P, Sundaram B, et al. HCK regulates NLRP12-mediated PANoptosis. Proc Natl Acad Sci U S A. 2025;122(21). doi:10.1073/pnas.2422079122
20. Kazmierczak BI, Golovkine G, Faudry E, et al. VE-cadherin cleavage by LasB protease from Pseudomonas aeruginosa facilitates type III secretion system toxicity in endothelial cells. PLoS Pathog. 2014;10(3). doi:10.1371/journal.ppat.1003939
21. Engel J, Balachandran P. Role of Pseudomonas aeruginosa type III effectors in disease. Curr Opin Microbiol. 2009;12(1):61–66. doi:10.1016/j.mib.2008.12.007
22. Sana TG, Baumann C, Merdes A, et al. Internalization of Pseudomonas aeruginosa strain PAO1 into epithelial cells is promoted by interaction of a T6SS effector with the microtubule network. mBio. 2015;6(3). doi:10.1128/mBio.00712-15
23. Wettstadt S, Wood TE, Fecht S, Filloux A. Delivery of the Pseudomonas aeruginosa phospholipase effectors PldA and PldB in a VgrG- and H2-T6SS-dependent manner. Front Microbiol. 2019. doi:10.3389/fmicb.2019.01718
24. Ugalde-Silva P, Gonzalez-Lugo O, Navarro-Garcia F. Tight junction disruption induced by type 3 secretion system effectors injected by enteropathogenic and enterohemorrhagic Escherichia coli. Front Cell Infect Microbiol. 2016;6. doi:10.3389/fcimb.2016.00087
25. Wang Y, Zhang S, Li H, et al. Small-molecule modulators of toll-like receptors. Acc Chem Res. 2020;53(5):1046–1055. doi:10.1021/acs.accounts.9b00631
26. Ciesielska A, Matyjek M, Kwiatkowska K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell Mol Life Sci. 2020;78(4):1233–1261. doi:10.1007/s00018-020-03656-y
27. Rooijakkers SHM, Chen Y-G, Zhang Y, et al. Control of methicillin-resistant staphylococcus aureus Pneumonia utilizing TLR2 agonist Pam3CSK4. PLoS One. 2016;11(3). doi:10.1371/journal.pone.0149233
28. Cowardin CA, Buonomo EL, Saleh MM, et al. The binary toxin CDT enhances Clostridium difficile virulence by suppressing protective colonic eosinophilia. Nat Microbiol. 2016;1(8). doi:10.1038/nmicrobiol.2016.108
29. Wu Y, Sun Y, Dong X, et al. The synergism of PGN, LTA and LPS in inducing transcriptome changes, inflammatory responses and a decrease in lactation as well as the associated epigenetic mechanisms in bovine mammary epithelial cells. Toxins (Basel). 2020;12(6). doi:10.3390/toxins12060387
30. Parolini C. Sepsis and high-density lipoproteins: pathophysiology and potential new therapeutic targets. Biochim Biophys Acta Mol Basis Dis. 2025;1871(5). doi:10.1016/j.bbadis.2025.167761
31. Im J, Jeon JH, Lee D, et al. Muramyl dipeptide potentiates Staphylococcus aureus lipoteichoic acid-induced nitric oxide production via TLR2/NOD2/PAFR signaling pathways. Front Immunol. 2024;15. doi:10.3389/fimmu.2024.1451315
32. Talreja J, Kabir MH, Filla MB, Stechschulte DJ, Dileepan KN. Histamine induces toll‐like receptor 2 and 4 expression in endothelial cells and enhances sensitivity to gram‐positive and gram‐negative bacterial cell wall components. Immunology. 2004;113(2):224–233. doi:10.1111/j.1365-2567.2004.01946.x
33. Zhang X, Hu X, Rao X. Apoptosis induced by Staphylococcus aureus toxins. Microbiol Res. 2017;205:19–24. doi:10.1016/j.micres.2017.08.006
34. Wang X, Li H, Wang J, et al. Staphylococcus aureus extracellular vesicles induce apoptosis and restrain mitophagy-mediated degradation of damaged mitochondria. Microbiol Res. 2023;273. doi:10.1016/j.micres.2023.127421
35. Van Krüchten A, Wilden JJ, Niemann S, et al. Staphylococcus aureus triggers a shift from influenza virus–induced apoptosis to necrotic cell death. FASEB J. 2018;32(5):2779–2793. doi:10.1096/fj.201701006R
36. Mulcahy ME, O’Brien EC, O’Keeffe KM, Vozza EG, Leddy N, McLoughlin RM. Manipulation of autophagy and apoptosis facilitates intracellular survival of Staphylococcus aureus in human neutrophils. Front Immunol. 2020;11. doi:10.3389/fimmu.2020.565545
37. Holzinger D, Gieldon L, Mysore V, et al. Staphylococcus aureus panton-valentine leukocidin induces an inflammatory response in human phagocytes via the NLRP3 inflammasome. J Leukoc Biol. 2012;92(5):1069–1081. doi:10.1189/jlb.0112014
38. Hara H, Seregin SS, Yang D, et al. The NLRP6 inflammasome recognizes lipoteichoic acid and regulates gram-positive pathogen infection. Cell. 2018;175(6):1651–1664.e14. doi:10.1016/j.cell.2018.09.047
39. Miller LS, Kitur K, Parker D, et al. Toxin-induced necroptosis is a major mechanism of Staphylococcus aureus lung damage. PLoS Pathog. 2015;11(4). doi:10.1371/journal.ppat.1004820
40. Feng S, Yang Y, Liu Z, et al. Intracellular bacteriolysis contributes to pathogenicity of Staphylococcus aureus by exacerbating AIM2-mediated inflammation and necroptosis. Virulence. 2022;13(1):1684–1696. doi:10.1080/21505594.2022.2127209
41. Marriott HM, Dockrell DH. Streptococcus pneumoniae: the role of apoptosis in host defense and pathogenesis. Int J Biochem Cell Biol. 2006;38(11):1848–1854. doi:10.1016/j.biocel.2006.06.001
42. Lea S, Beech A, Baker J, et al. Differential responses of COPD macrophages to respiratory bacterial pathogens. ERJ Open Res. 2022;8(3). doi:10.1183/23120541.00044-2022
43. Preston JA, Bewley MA, Marriott HM, et al. Alveolar macrophage apoptosis–associated bacterial killing helps prevent murine Pneumonia. Am J Respir Crit Care Med. 2019;200(1):84–97. doi:10.1164/rccm.201804-0646OC
44. Wang X, Zhao Y, Wang D, et al. ALK-JNK signaling promotes NLRP3 inflammasome activation and pyroptosis via NEK7 during Streptococcus pneumoniae infection. Mol Immunol. 2023;157:78–90. doi:10.1016/j.molimm.2023.03.016
45. Xu D, Wu X, Peng L, et al. The critical role of NLRP6 inflammasome in Streptococcus pneumoniae infection in vitro and in vivo. Int J Mol Sci. 2021;22(8). doi:10.3390/ijms22083876
46. Feng S, Chen T, Lei G, et al. Absent in melanoma 2 inflammasome is required for host defence against Streptococcus pneumoniae infection. Innate Immun. 2019;25(7):412–419. doi:10.1177/1753425919860252
47. Riegler AN, Brissac T, Gonzalez-Juarbe N, Orihuela CJ. Necroptotic cell death promotes adaptive immunity against colonizing Pneumococci. Front Immunol. 2019. doi:10.3389/fimmu.2019.00615
48. González-Juarbe N, Bradley KM, Shenoy AT, et al. Pore-forming toxin-mediated ion dysregulation leads to death receptor-independent necroptosis of lung epithelial cells during bacterial pneumonia. Cell Death Differ. 2017;24(5):917–928. doi:10.1038/cdd.2017.49
49. Huang H-R, Cho SJ, Harris RM, et al. RIPK3 activates MLKL-mediated necroptosis and inflammasome signaling during streptococcus infection. Am J Respir Cell Mol Biol. 2021;64(5):579–591. doi:10.1165/rcmb.2020-0312OC
50. Hakansson AP, Bergenfelz C. Low NF-κB activation and necroptosis in alveolar macrophages: a new virulence property of Streptococcus pneumoniae. J Infect Dis. 2017;216(4):402–404. doi:10.1093/infdis/jix161
51. Van hauwermeiren F, Van Opdenbosch N, Van Gorp H, et al. Bacillus anthracis induces NLRP3 inflammasome activation and caspase-8–mediated apoptosis of macrophages to promote lethal anthrax. Proc Natl Acad Sci U S A. 2022;119(2). doi:10.1073/pnas.2116415119
52. Van Opdenbosch N, Van Gorp H, Verdonckt M, et al. Caspase-1 engagement and TLR-induced c-FLIP expression suppress ASC/Caspase-8-dependent apoptosis by inflammasome sensors NLRP1b and NLRC4. Cell Rep. 2017;21(12):3427–3444. doi:10.1016/j.celrep.2017.11.088
53. Greaney AJ, Portley MK, O’Mard D, et al. Frontline science: anthrax lethal toxin-induced, NLRP1-mediated IL-1β release is a neutrophil and PAD4-dependent event. J Leukoc Biol. 2020;108(3):773–786. doi:10.1002/jlb.4hi0320-028r
54. Guey B, Bodnar M, Manié SN, Tardivel A, Petrilli V. Caspase-1 autoproteolysis is differentially required for NLRP1b and NLRP3 inflammasome function. Proc Natl Acad Sci U S A. 2014;111(48):17254–17259. doi:10.1073/pnas.1415756111
55. Gao X, Teng T, Liu Y, et al. Anthrax lethal toxin and tumor necrosis factor-α synergize on intestinal epithelia to induce mouse death. Protein Cell. 2024;15(2):135–148. doi:10.1093/procel/pwad050
56. Carrero JA, Unanue ER. Mechanisms and immunological effects of apoptosis caused by Listeria monocytogenes. Adv Immunol. 2012;113:157–174. doi:10.1016/B978-0-12-394590-7.00001-4
57. Li S, Li Y, Chen G, Zhang J, Xu F, Wu M. Restraining reactive oxygen species in Listeria monocytogenes promotes the apoptosis of glial cells. Redox Rep. 2017;22(4):190–196. doi:10.1080/13510002.2016.1173327
58. Gao A, Tang H, Zhang Q, et al. Mst1/2-ALK promotes NLRP3 inflammasome activation and cell apoptosis during Listeria monocytogenes infection. J Microbiol. 2021;59(7):681–692. doi:10.1007/s12275-021-0638-2
59. Sauer JD, Witte CE, Zemansky J, Hanson B, Lauer P, Portnoy DA. Listeria monocytogenes triggers AIM2-mediated pyroptosis upon infrequent bacteriolysis in the macrophage cytosol. Cell Host Microbe. 2010;7(5):412–419. doi:10.1016/j.chom.2010.04.004
60. Uematsu T, Tsuchiya K, Kobayashi N, et al. Mint3 depletion-mediated glycolytic and oxidative alterations promote pyroptosis and prevent the spread of Listeria monocytogenes infection in macrophages. Cell Death Dis. 2021;12(4):404. doi:10.1038/s41419-021-03691-y
61. Sai K, Parsons C, House JS, Kathariou S, Ninomiya-Tsuji J. Necroptosis mediators RIPK3 and MLKL suppress intracellular Listeria replication independently of host cell killing. J Cell Biol. 2019;218(6):1994–2005. doi:10.1083/jcb.201810014
62. Zhang T, Balachandran S. Bayonets over bombs: RIPK3 and MLKL restrict Listeria without triggering necroptosis. J Cell Biol. 2019;218(6):1773–1775. doi:10.1083/jcb.201905047
63. Wu Z, Takigawa H, Maruyama H, Nambu T, Mashimo C, Okinaga T. TLR2-dependent and independent pyroptosis in dTHP-1 cells induced by Actinomyces oris MG-1. Biochem Biophys Rep. 2024;38:101680. doi:10.1016/j.bbrep.2024.101680
64. Wen Y, Kang X, Li Z, Xia L, Lu Y. Identification of a secretory heme-binding protein from Nocardia seriolae involved in cell apoptosis. J Fish Dis. 2022;45(8):1189–1199. doi:10.1111/jfd.13654
65. Liu W, Deng Y, Tan A, et al. Intracellular behavior of Nocardia seriolae and its apoptotic effect on RAW264.7 macrophages. Front Cell Infect Microbiol. 2023;13:1138422. doi:10.3389/fcimb.2023.1138422
66. Potapenko K, Lisiutin G, Vasylieva N, et al. Antimicrobial and anticancer activity of Streptomyces ambofaciens (Myt 8) and S. globisporus ONU 1019 (Myt 11) secondary metabolites isolated from the Odesa Bay, the Black Sea: an in vitro study. Biomed Pharmacother. 2025;186:117981. doi:10.1016/j.biopha.2025.117981
67. Deo P, Chow SH, Hay ID, et al. Outer membrane vesicles from Neisseria gonorrhoeae target PorB to mitochondria and induce apoptosis. PLoS Pathog. 2018;14(3):e1006945. doi:10.1371/journal.ppat.1006945
68. Chateau A, Seifert HS. Neisseria gonorrhoeae survives within and modulates apoptosis and inflammatory cytokine production of human macrophages. Cell Microbiol. 2016;18(4):546–560. doi:10.1111/cmi.12529
69. Chen A, Seifert HS. Neisseria gonorrhoeae-mediated inhibition of apoptotic signalling in polymorphonuclear leukocytes. Infect Immun. 2011;79(11):4447–4458. doi:10.1128/IAI.01267-10
70. Simons MP, Nauseef WM, Griffith TS, Apicella MA. Neisseria gonorrhoeae delays the onset of apoptosis in polymorphonuclear leukocytes. Cell Microbiol. 2006;8(11):1780–1790. doi:10.1111/j.1462-5822.2006.00748.x
71. Li LH, Lin JS, Chiu HW, et al. Mechanistic insight into the activation of the NLRP3 inflammasome by neisseria gonorrhoeae in macrophages. Front Immunol. 2019;10:1815. doi:10.3389/fimmu.2019.01815
72. Ritter JL, Genco CA. Neisseria gonorrhoeae-induced inflammatory pyroptosis in human macrophages is dependent on intracellular Gonococci and Lipooligosaccharide. J Cell Death. 2018;11:1179066017750902. doi:10.1177/1179066017750902
73. Eriksson L, Stenmark B, Deghmane AE, et al. Difference in virulence between Neisseria meningitidis serogroups W and Y in transgenic mice. BMC Microbiol. 2020;20(1):92. doi:10.1186/s12866-020-01760-4
74. Massari P, Gunawardana J, Liu X, Wetzler LM. Meningococcal porin PorB prevents cellular apoptosis in a toll-like receptor 2- and NF-kappaB-independent manner. Infect Immun. 2010;78(3):994–1003. doi:10.1128/IAI.00156-09
75. Pagliuca C, Colicchio R, Resta SC, et al. Neisseria meningitidis activates pyroptotic pathways in a mouse model of meningitis: role of a two-partner secretion system. Front Cell Infect Microbiol. 2024;14:1384072. doi:10.3389/fcimb.2024.1384072
76. Talà A, Guerra F, Calcagnile M, et al. HrpA anchors meningococci to the dynein motor and affects the balance between apoptosis and pyroptosis. J Biomed Sci. 2022;29(1). doi:10.1186/s12929-022-00829-8
77. John CM, Phillips NJ, Jarvis GA. Predominant phosphorylation patterns in Neisseria meningitidis lipid A determined by top-down MS/MS. J Lipid Res. 2020;61(11):1437–1449. doi:10.1194/jlr.RA120001014
78. Enosi Tuipulotu D, Feng S, Pandey A, et al. Immunity against Moraxella catarrhalis requires guanylate-binding proteins and caspase-11-NLRP3 inflammasomes. EMBO J. 2023;42(6):e112558. doi:10.15252/embj.2022112558
79. N’Guessan PD, Vigelahn M, Bachmann S, et al. The UspA1 protein of Moraxella catarrhalis induces CEACAM-1-dependent apoptosis in alveolar epithelial cells. J Infect Dis. 2007;195(11):1651–1660. doi:10.1086/514820
80. Jendholm J, Samuelsson M, Cardell LO, Forsgren A, Riesbeck K. Moraxella catarrhalis-dependent tonsillar B cell activation does not lead to apoptosis but to vigorous proliferation resulting in nonspecific IgM production. J Leukoc Biol. 2008;83(6):1370–1378. doi:10.1189/jlb.1107788
81. Versi A, Azim A, Ivan FX, et al. A severe asthma phenotype of excessive airway Haemophilus influenzae relative abundance associated with sputum neutrophilia. Clin Transl Med. 2024;14(9). doi:10.1002/ctm2.70007
82. Yan X, Jin L, Zhou H, Wan H, Wan H, Yang J. Amygdalin reverses macrophage panoptosis induced by drug-resistant Escherichia coli. J Microbiol Biotechnol. 2023;33(10):1281–1291. doi:10.4014/jmb.2306.06030
83. Lu C, Jin L, Zhou H, Yang J, Wan H. Chlorogenic acid inhibits macrophage PANoptosis induced by cefotaxime-resistant Escherichia coli. Arch Microbiol. 2024;206(2):67. doi:10.1007/s00203-023-03777-5
84. Priya A, Chandel S, Joon A, Ghosh S. Molecular mechanism of Enteroaggregative Escherichia coli induced apoptosis in cultured human intestinal epithelial cells. J Med Microbiol. 2023;72(10). doi:10.1099/jmm.0.001760
85. Priya A, Kaur K, Bhattacharyya S, Chakraborti A, Ghosh S. Cell cycle arrest and apoptosis induced by enteroaggregative Escherichia coli in cultured human intestinal epithelial cells. J Med Microbiol. 2017;66(2):217–225. doi:10.1099/jmm.0.000405
86. Xia Y, Bin P, Liu S, et al. Enterotoxigenic Escherichia coli infection promotes apoptosis in piglets. Microb Pathog. 2018;125:290–294. doi:10.1016/j.micpath.2018.09.032
87. Li Y, Zhu Y, Chu B, Liu N, Chen S, Wang J. Lactobacillus rhamnosus GR-1 prevents Escherichia coli-induced apoptosis through PINK1/Parkin-mediated mitophagy in bovine mastitis. Front Immunol. 2021;12:715098. doi:10.3389/fimmu.2021.715098
88. Zhuang C, Zhao J, Zhang S, Shahid M. Escherichia coli infection mediates pyroptosis via activating p53-p21 pathway-regulated apoptosis and cell cycle arrest in bovine mammary epithelial cells. Microb Pathog. 2023;184:106338. doi:10.1016/j.micpath.2023.106338
89. Cheng Y, Xiao X, Fu J, Zong X, Lu Z, Wang Y. Escherichia coli K88 activates NLRP3 inflammasome-mediated pyroptosis in vitro and in vivo. Biochem Biophys Rep. 2024;38:101665. doi:10.1016/j.bbrep.2024.101665
90. Goddard PJ, Sanchez-Garrido J, Slater SL, et al. Enteropathogenic Escherichia coli stimulates effector-driven rapid caspase-4 activation in human macrophages. Cell Rep. 2019;27(4):1008–1017e6. doi:10.1016/j.celrep.2019.03.100
91. Li X, Zhang T, Kang L, et al. Apoptotic caspase-7 activation inhibits non-canonical pyroptosis by GSDMB cleavage. Cell Death Differ. 2023;30(9):2120–2134. doi:10.1038/s41418-023-01211-3
92. Hu J, Wang D, Huang X, et al. Effects of TolC on the pathogenicity of porcine extraintestinal pathogenic Escherichia coli. Front Immunol. 2022;13:929740. doi:10.3389/fimmu.2022.929740
93. Li Z, Shang W, Mei T, et al. Outer membrane vesicles of avian pathogenic Escherichia coli induce necroptosis and NF-kappaB activation in chicken macrophages via RIPK1 mediation. Res Vet Sci. 2024;170:105185. doi:10.1016/j.rvsc.2024.105185
94. Pearson JS, Giogha C, Muhlen S, et al. EspL is a bacterial cysteine protease effector that cleaves RHIM proteins to block necroptosis and inflammation. Nat Microbiol. 2017;2:16258. doi:10.1038/nmicrobiol.2016.258
95. Rahiminejad M, Montaseri M, Yousefi MH, Nazifi S, Wagemans J, Hosseinzadeh S. Inflammation, toxicity, and apoptosis reducing potential of bacteriophage Ariobarzanes on intestinal cells infected with Salmonella Typhimurium. Sci Rep. 2025;15(1):14958. doi:10.1038/s41598-025-99116-3
96. Ding C, Wu H, Cao X, et al. Lactobacillus crispatus-derived exopolysaccharides with antibacterial activity limit Salmonella typhimurium invasion by inhibiting inflammasome-mediated pyroptosis. Food Funct. 2022;13(20):10501–10515. doi:10.1039/d2fo02125k
97. Orning P, Weng D, Starheim K, et al. Pathogen blockade of TAK1 triggers caspase-8-dependent cleavage of gasdermin D and cell death. Science. 2018;362(6418):1064–1069. doi:10.1126/science.aau2818
98. Jetton D, Muendlein HI, Connolly WM, et al. Non-canonical autophosphorylation of RIPK1 drives timely pyroptosis to control Yersinia infection. Cell Rep. 2024;43(8):114641. doi:10.1016/j.celrep.2024.114641
99. Yang Y, Fang H, Xie Z, et al. Yersinia infection induces glucose depletion and AMPK-dependent inhibition of pyroptosis in mice. Nat Microbiol. 2024;9(8):2144–2159. doi:10.1038/s41564-024-01734-6
100. Zheng Z, Deng W, Bai Y, et al. The lysosomal rag-ragulator complex licenses RIPK1 and Caspase-8-mediated pyroptosis by Yersinia. Science. 2021;372(6549). doi:10.1126/science.abg0269
101. Nataraj NM, Sillas RG, Herrmann BI, Shin S, Brodsky IE. Blockade of IKK signaling induces RIPK1-independent apoptosis in human macrophages. PLoS Pathog. 2024;20(8):e1012469. doi:10.1371/journal.ppat.1012469
102. Sundaram B, Karki R, Kanneganti T-D. NLRC4 deficiency leads to enhanced phosphorylation of MLKL and necroptosis. Immunohorizons. 2022;6(3):243–252. doi:10.4049/immunohorizons.2100118
103. Wood SJ, Goldufsky JW, Bello D, Masood S, Shafikhani SH. Pseudomonas aeruginosa ExoT induces mitochondrial apoptosis in target host cells in a manner that depends on its GTPase-activating Protein (GAP) domain activity. J Biol Chem. 2015;290(48):29063–29073. doi:10.1074/jbc.M115.689950
104. Iwanski B, Mizerska-Kowalska M, Andrejko M. Pseudomonas aeruginosa exotoxin A induces apoptosis in Galleria mellonella hemocytes. J Invertebr Pathol. 2023;197:107884. doi:10.1016/j.jip.2023.107884
105. Santoni K, Pericat D, Gorse L, et al. Caspase-1-driven neutrophil pyroptosis and its role in host susceptibility to Pseudomonas aeruginosa. PLoS Pathog. 2022;18(7):e1010305. doi:10.1371/journal.ppat.1010305
106. Minns MS, Liboro K, Lima TS, et al. NLRP3 selectively drives IL-1beta secretion by Pseudomonas aeruginosa infected neutrophils and regulates corneal disease severity. Nat Commun. 2023;14(1):5832. doi:10.1038/s41467-023-41391-7
107. Xu S, Liu X, Liu X, et al. Wedelolactone ameliorates Pseudomonas aeruginosa-induced inflammation and corneal injury by suppressing caspase-4/5/11/GSDMD-mediated non-canonical pyroptosis. Exp Eye Res. 2021;211:108750. doi:10.1016/j.exer.2021.108750
108. Liu Z, Sun L, Li L, et al. Pseudomonas aeruginosa mediates host necroptosis through Rhl-Pqs quorum sensing interaction. Immunohorizons. 2024;8(9):721–728. doi:10.4049/immunohorizons.2400012
109. Guan J, Yao W, Zhang L, et al. Contribution of Pseudomonas aeruginosa - mediated club cell necroptosis to the bias of type 17 inflammation and steroid insensitivity in asthma. J Adv Res. 2024. doi:10.1016/j.jare.2024.10.020
110. Zhao Z, Liu J, Deng Y, et al. The Vibrio alginolyticus T3SS effectors, Val1686 and Val1680, induce cell rounding, apoptosis and lysis of fish epithelial cells. Virulence. 2018;9(1):318–330. doi:10.1080/21505594.2017.1414134
111. Cohen H, Baram N, Edry-Botzer L, Munitz A, Salomon D, Gerlic M. Vibrio pore-forming leukocidin activates pyroptotic cell death via the NLRP3 inflammasome. Emerg Microbes Infect. 2020;9(1):278–290. doi:10.1080/22221751.2020.1720526
112. Wang Y, Luo J, Zhao Y, Zhang J, Guan X, Sun L. Haemolysins are essential to the pathogenicity of deep-sea Vibrio fluvialis. iScience. 2024;27(5):109558. doi:10.1016/j.isci.2024.109558
113. Qin KW, Liu JF, Wu CL, Zhang C, Zhou LJ. Resveratrol prevents vibrio vulnificus-induced sepsis by attenuating necroptosis. Biomed Environ Sci. 2023;36(2):135–145. doi:10.3967/bes2023.017
114. Singer M, Deutschman CS, Seymour CW, et al. The third international consensus definitions for sepsis and septic shock (Sepsis-3). JAMA. 2016;315(8):801–810. doi:10.1001/jama.2016.0287
115. Liu B, Du H, Zhang J, et al. Developing a new sepsis screening tool based on lymphocyte count, international normalized ratio and procalcitonin (LIP score). Sci Rep. 2022;12(1):20002. doi:10.1038/s41598-022-16744-9
116. Reinhart K, Daniels R, Kissoon N, Machado FR, Schachter RD, Finfer S. Recognizing sepsis as a global health priority - A WHO resolution. N Engl J Med. 2017;377(5):414–417. doi:10.1056/NEJMp1707170
117. Xu J, Zhu M, Luo P, Gong Y. Machine learning screening and validation of PANoptosis-related gene signatures in sepsis. J Inflamm Res. 2024;17:4765–4780. doi:10.2147/jir.S461809
118. Zhang X, Su C, Zhao S, Li J, Yu F. Combination therapy of Ulinastatin with Thrombomodulin alleviates endotoxin (LPS) - induced liver and kidney injury via inhibiting apoptosis, oxidative stress and HMGB1/TLR4/NF-kappaB pathway. Bioengineered. 2022;13(2):2951–2970. doi:10.1080/21655979.2021.2024686
119. Karki R, Sharma BR, Tuladhar S, et al. Synergism of TNF-alpha and IFN-gamma triggers inflammatory cell death, tissue damage, and mortality in SARS-CoV-2 infection and cytokine shock syndromes. Cell. 2021;184(1):149–168e17. doi:10.1016/j.cell.2020.11.025
120. Wang Y, Fu X, Shang Z, et al. In vivo and in vitro study on the regulatory mechanism of XiaoChaiHu decoction on PANoptosis in sepsis-induced cardiomyopathy. J Ethnopharmacol. 2025;336:118740. doi:10.1016/j.jep.2024.118740
121. Wei S, Wu L, Xiang Z, et al. EIF2AK2 protein targeted activation of AIM2-mediated PANoptosis promotes sepsis-induced acute kidney injury. Ren Fail. 2024;46(2):2403649. doi:10.1080/0886022X.2024.2403649
122. Zhang M, Shang L, Zhou F, et al. Dachengqi decoction dispensing granule ameliorates LPS-induced acute lung injury by inhibiting PANoptosis in vivo and in vitro. J Ethnopharmacol. 2025;336:118699. doi:10.1016/j.jep.2024.118699
123. Gong T, Wang QD, Loughran PA, et al. Mechanism of lactic acidemia-promoted pulmonary endothelial cells death in sepsis: role for CIRP-ZBP1-PANoptosis pathway. Mil Med Res. 2024;11(1):71. doi:10.1186/s40779-024-00574-z
124. Vaughn VM, Dickson RP, Horowitz JK, Flanders SA. Community-acquired pneumonia: a review. JAMA. 2024;332(15):1282–1295. doi:10.1001/jama.2024.14796
125. Bos LDJ, Ware LB. Acute respiratory distress syndrome: causes, pathophysiology, and phenotypes. Lancet. 2022;400(10358):1145–1156. doi:10.1016/S0140-6736(22)01485-4
126. Matthay MA, Zemans RL, Zimmerman GA, et al. Acute respiratory distress syndrome. Nat Rev Dis Primers. 2019;5(1):18. doi:10.1038/s41572-019-0069-0
127. Hao Q, Gao W, Zhang P, Yan P. Identification of PANoptosis-related genes in community-acquired Pneumonia diagnosis. J Inflamm Res. 2024;17:10289–10304. doi:10.2147/JIR.S491315
128. Xiao H, Zhao Q, Yuan J, et al. IFN-gamma promotes PANoptosis in Pasteurella multocida toxin-induced pneumonia in mice. Vet Microbiol. 2023;285:109848. doi:10.1016/j.vetmic.2023.109848
129. Messaoud-Nacer Y, Culerier E, Rose S, et al. STING agonist diABZI induces PANoptosis and DNA mediated acute respiratory distress syndrome (ARDS). Cell Death Dis. 2022;13(3):269. doi:10.1038/s41419-022-04664-5
130. Chen Y, Wu X, Jiang Z, Li X. KAE ameliorates LPS-mediated acute lung injury by inhibiting PANoptosis through the intracellular DNA-cGAS-STING axis. Front Pharmacol. 2024;15:1461931. doi:10.3389/fphar.2024.1461931
131. Cui Y, Wang X, Lin F, et al. MiR-29a-3p improves acute lung injury by reducing alveolar epithelial cell PANoptosis. Aging Dis. 2022;13(3):899–909. doi:10.14336/AD.2021.1023
132. You YP, Yan L, Ke HY, et al. Baicalin inhibits PANoptosis by blocking mitochondrial Z-DNA formation and ZBP1-PANoptosome assembly in macrophages. Acta Pharmacol Sin. 2025;46(2):430–447. doi:10.1038/s41401-024-01376-8
133. Kaplan GG, Windsor JW. The four epidemiological stages in the global evolution of inflammatory bowel disease. Nat Rev Gastroenterol Hepatol. 2021;18(1):56–66. doi:10.1038/s41575-020-00360-x
134. Martin JS, Monaghan TM, Wilcox MH. Clostridium difficile infection: epidemiology, diagnosis and understanding transmission. Nat Rev Gastroenterol Hepatol. 2016;13(4):206–216. doi:10.1038/nrgastro.2016.25
135. Wang JM, Yang J, Xia WY, et al. Comprehensive analysis of PANoptosis-related gene signature of ulcerative colitis. Int J Mol Sci. 2023;25(1). doi:10.3390/ijms25010348
136. Zhang M, Liu T, Luo L, Xie Y, Wang F. Biological characteristics, immune infiltration and drug prediction of PANoptosis related genes and possible regulatory mechanisms in inflammatory bowel disease. Sci Rep. 2025;15(1):2033. doi:10.1038/s41598-024-84911-1
137. Gong W, Liu Z, Wang Y, et al. Reprogramming of Treg cell-derived small extracellular vesicles effectively prevents intestinal inflammation from PANoptosis by blocking mitochondrial oxidative stress. Trends Biotechnol. 2025;43(4):893–917. doi:10.1016/j.tibtech.2024.11.017
138. Yuan H, Zhou L, Chen Y, et al. Salmonella effector SopF regulates PANoptosis of intestinal epithelial cells to aggravate systemic infection. Gut Microbes. 2023;15(1):2180315. doi:10.1080/19490976.2023.2180315
139. Tan C, Xiang Z, Wang S, et al. Diosmin alleviates colitis by inhibiting PANoptosis of intestinal epithelial cells and regulating gut microbiota and metabolites. Phytomedicine. 2025;141:156671. doi:10.1016/j.phymed.2025.156671
140. Zhou R, Ying J, Qiu X, et al. A new cell death program regulated by toll-like receptor 9 through p38 mitogen-activated protein kinase signaling pathway in a neonatal rat model with sepsis associated encephalopathy. Chin Med J. 2022;135(12):1474–1485. doi:10.1097/CM9.0000000000002010
141. Liu X, Li Y, Zhang W, et al. Inhibition of cIAP1/2 reduces RIPK1 phosphorylation in pulmonary endothelial cells and alleviate sepsis-induced lung injury and inflammatory response. Immunol Res. 2024;72(4):841–850. doi:10.1007/s12026-024-09491-8
142. Wang Y, Yuan M, Qin J, et al. TIPE2 alleviates sepsis-induced lung injury by inhibiting PANoptosis in murine alveolar macrophages. Inflammation. 2025. doi:10.1007/s10753-025-02288-3
143. Zhang Y-T, Li -H-H, Teng F. Inhibition of Piezo1 ameliorates septic cardiomyopathy by blocking calcium-dependent PANoptosis. Eur J Pharmacol. 2025;996. doi:10.1016/j.ejphar.2025.177438
144. Li T, Adams J, Zhu P, et al. The role of heme in sepsis induced Kupffer cell PANoptosis and senescence. Cell Death Dis. 2025;16(1):284. doi:10.1038/s41419-025-07637-6
145. Zhou X, Yu X, Wan C, et al. NINJ1 regulates platelet activation and PANoptosis in septic disseminated intravascular coagulation. Int J Mol Sci. 2023;24(4). doi:10.3390/ijms24044168
146. Shi FL, Li Q, Xu R, et al. Blocking reverse electron transfer-mediated mitochondrial DNA oxidation rescues cells from PANoptosis. Acta Pharmacol Sin. 2024;45(3):594–608. doi:10.1038/s41401-023-01182-8
147. Chen S, Wu G-D, Li T, et al. Targeting GSDMD JX06 inhibits PANoptosis and multiple organ injury. Biochem Pharmacol. 2025;233. doi:10.1016/j.bcp.2025.116765
148. He YQ, Deng JL, Zhou CC, et al. Ursodeoxycholic acid alleviates sepsis-induced lung injury by blocking PANoptosis via STING pathway. Int Immunopharmacol. 2023;125(Pt B):111161. doi:10.1016/j.intimp.2023.111161
149. Wang Y, Liu H, Feng Y, Wu S, He J, Cao L. Sulforaphane inhibits LPS-induced macrophage PANoptosis via TLR4/NFkappaB pathway: a potential therapeutic strategy for acute lung injury. Tohoku J Exp Med. 2025;265(4):239–248. doi:10.1620/tjem.2024.J105
150. Zhou X, Xin G, Wan C, et al. Myricetin reduces platelet PANoptosis in sepsis to delay disseminated intravascular coagulation. Biochem Biophys Res Commun. 2024;724:150140. doi:10.1016/j.bbrc.2024.150140
151. Guo J, Luo Y, Zuo J, Teng J, Shen B, Liu X. Echinacea polyphenols inhibit NLRP3-dependent pyroptosis, apoptosis, and necroptosis via suppressing no production during lipopolysaccharide-induced acute lung injury. J Agric Food Chem. 2023;71(19):7289–7298. doi:10.1021/acs.jafc.2c08382
152. Yang Z, Kao X, Zhang L, Huang N, Chen J, He M. Exploring the anti-PANoptosis mechanism of dachaihu decoction against sepsis-induced acute lung injury: network pharmacology, bioinformatics, and experimental validation. Drug Des Devel Ther. 2025;19:349–368. doi:10.2147/DDDT.S495225
153. Zhang X, Yang J, Lu Y, Liu Y, Wang T, Yu F. Human Urinary Kallidinogenase improves vascular endothelial injury by activating the Nrf2/HO-1 signaling pathway. Chem Biol Interact. 2024;403:111230. doi:10.1016/j.cbi.2024.111230
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Comparison of Autof Ms1000 and EXS3000 MALDI-TOF MS Platforms for Routine Identification of Microorganisms
Xiong L, Long X, Ni L, Wang L, Zhang Y, Cui L, Guo J, Yang C
Infection and Drug Resistance 2023, 16:913-921
Published Date: 15 February 2023
PANoptosis: Cross-Talk Among Apoptosis, Necroptosis, and Pyroptosis in Neurological Disorders
Li P, Gao Y, Tao Z, Mu Z, Du S, Zhao X
Journal of Inflammation Research 2025, 18:8131-8140
Published Date: 19 June 2025
Promising Future of Novel Beta-Lactam Antibiotics Against Bacterial Resistance
Tayeb SM, Alharbi JA, Alattas BB, Alotaibi DM, Althibaiti NM, Alharbi JF, SafirAldeen AE, Alqurashi IR, Wali SM
Drug Design, Development and Therapy 2025, 19:9185-9197
Published Date: 9 October 2025