")
Back to Journals » Journal of Inflammation Research » Volume 13
Targeting Chemokines and Chemokine Receptors in Multiple Sclerosis and Experimental Autoimmune Encephalomyelitis
Authors Dhaiban S, Al-Ani M, Elemam NM , Maghazachi AA
Received 14 July 2020
Accepted for publication 27 August 2020
Published 29 September 2020 Volume 2020:13 Pages 619—633
DOI https://doi.org/10.2147/JIR.S270872
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Ning Quan
Sarah Dhaiban, Mena Al-Ani, Noha Mousaad Elemam, Azzam A Maghazachi
College of Medicine and Immuno-Oncology Group, Sharjah Institute for Medical Research, University of Sharjah, Sharjah, United Arab Emirates
Correspondence: Azzam A Maghazachi
Department of Clinical Sciences, College of Medicine, University of Sharjah, Sharjah 27272, United Arab Emirates
Email [email protected]
Abstract: Multiple sclerosis (MS) is an immune-mediated and neurodegenerative disorder that results in inflammation and demyelination of the central nervous system (CNS). MS symptoms include walking difficulties, visual weakening, as well as learning and memory impairment, thus affecting the quality of the patient’s life. Chemokines and chemokine receptors are expressed on the immune cells as well as the CNS resident cells. Several sets of chemokine receptors and their ligands tend to be pathogenic players in MS, including CCL2, CCL3, CCL4, CCL5, CCL7, CCL8, CCL11, CCL17, CCL19, CCL21, CCL22, CXCL1, CXCL8, CXCL9, CXCL10, CXCL11, and CXCL16. Furthermore, current modulatory drugs that are used in the treatment of MS and its animal model, the experimental autoimmune encephalomyelitis (EAE), affect the expression of several chemokine and chemokine receptors. In this review, we highlight the pathogenic roles of chemokines and their receptors as well as utilizing them as potential therapeutic targets through selective agents, such as specific antibodies and receptor blockers, or indirectly through MS or EAE immunomodulatory drugs.
Keywords: multiple sclerosis, experimental autoimmune encephalomyelitis, chemokines, chemokine receptors
Introduction
Multiple sclerosis (MS) is an immune-mediated and neurodegenerative disease, causing demyelination of the central nervous system (CNS) characterized by formation of separated areas of inflammation called MS lesions.1 There are four key pathological characteristics of MS: (a) inflammation, that is proposed to be the essential cause of most of the events of CNS tissue damage; (b) demyelination, the trademark of MS, in which the myelin sheath or the oligodendrocyte cell body is damaged through the inflammatory process; (c) axonal loss or impairment; and (d) gliosis (astrocytic response to CNS damage).1,2 MS is a long-lasting disease that can influence the brain, spinal cord, and the optic nerves.3 It can cause impairment in vision, balance control, muscle control, and other essential body functions. This could be attributed to the damage of the myelin sheath, the protective layer of the nerve fibers, which adversely affects the communication between the brain and the rest of the body as the message or information conveyed through the nerves may arrive slower or may never arrive. Eventually, the nerves themselves may get damaged. Over time, the brain may shrink in size because axons are destroyed.4 Symptoms of MS are different, depending on the affected nerves and the severity of the symptoms. In difficult cases, patients with MS lose the ability to walk or talk.4 Most individuals with MS have attacks when the condition gets recognizably worse (relapse), followed by recovery times when symptoms improve (remission). In certain individuals, the illness may progress into secondary stage.4
There are four main subtypes for MS. The most common subtype is relapsing-remitting MS (RRMS), accounting for about 85% of MS patients. It is characterized by periods of recurring symptoms that are followed by remission phase, during which symptoms could partially or totally disappear.5 The second type is secondary progressive MS (SPMS) which starts with an initial relapse phase, and that could progress gradually leading to neurological worsening.5 The third type is primary progressive MS (PPMS) characterized by a continuous disease deterioration and no specific relapse phase.5 Finally, the fourth subtype is progressive relapsing MS (PRMS). In PRMS, there is a constant deterioration from the initial stage of the disease followed by a relapse phase.5,6
A mouse model for MS has been well established which is known as the experimental autoimmune encephalomyelitis (EAE). This model was extensively used to discover new therapies for MS. In fact, most, if not all experimental MS drugs must be examined in this mouse model before advancing any of these new drugs into clinical trials.7 In this review, we will briefly discuss the chemokines and their receptors in MS and EAE, as well as the impact of current MS and EAE modulatory drugs on the chemokine system.
Targeting Chemokines and Chemokine Receptors as Immunotherapeutic Approach for MS and EAE
Chemokines and Chemokine Receptors
Chemokines and their receptors have many roles in immune regulation, whether in physiological or pathological conditions.8 Chemokines are classified either according to the cysteine residues of the ligands (CC, CXC, C, and CX3C), or their function and expression.9 CC chemokines compose the biggest family containing two adjacent cysteine residues near their N-terminus, while their respective genes are clustered on chromosome 17 in humans. In CXC and CX3C chemokine subfamilies, there are one or three additional amino acids between the first two cysteine residues, respectively. Most of the CXC chemokines are grouped on chromosome 4 in humans.9 The XC chemokine is characterized by the absence of one of the first two N-terminal cysteine residues.10 Chemokines are crucial mediators in the recruitment and migration of immune cells to the inflammatory sites.11 Some chemokines can bind to several chemokine receptors, whereas certain chemokine receptors bind to more than one chemokine ligand. Chemokines are involved in autoimmune diseases, which include rheumatoid arthritis (RA), Graves’ disease (GD), systemic lupus erythematosus (SLE), and MS, among others.12,13 Hence, chemokine receptors represent novel targets for treating autoimmune diseases. Table 1 demonstrates the recent understanding of the 4 chemokine sub-families and their receptors (for reviews, please see).14–30
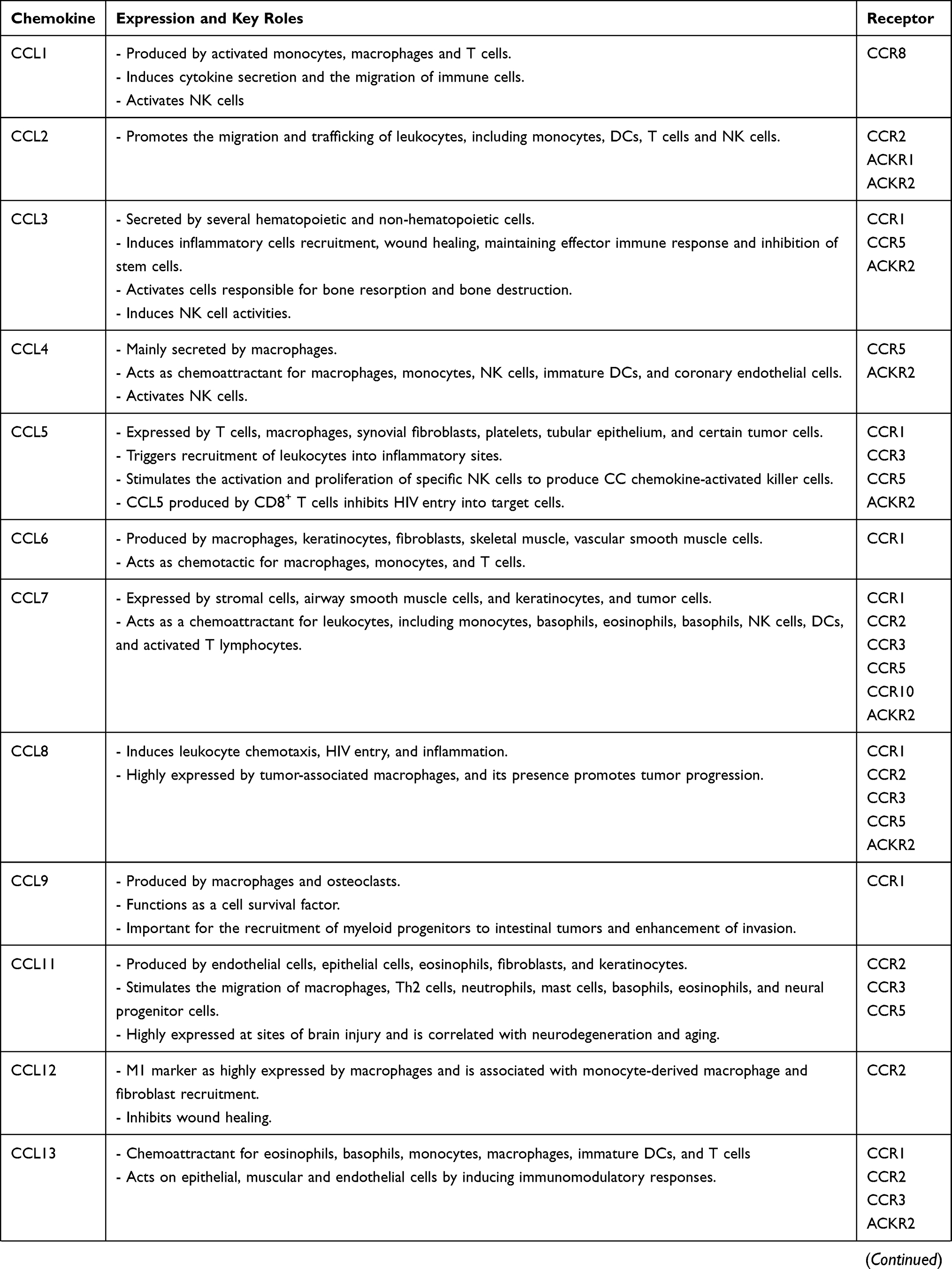 |  | 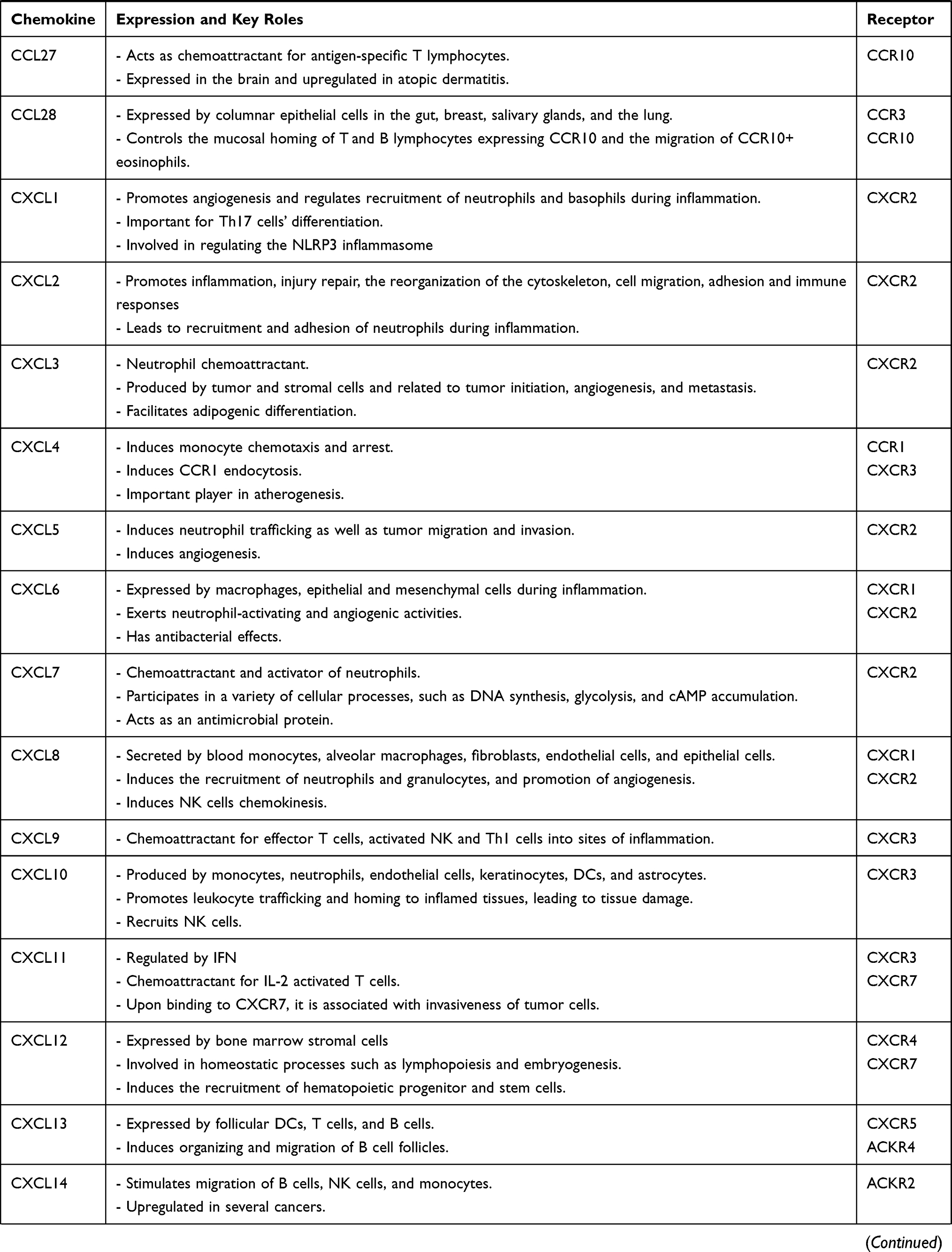 | 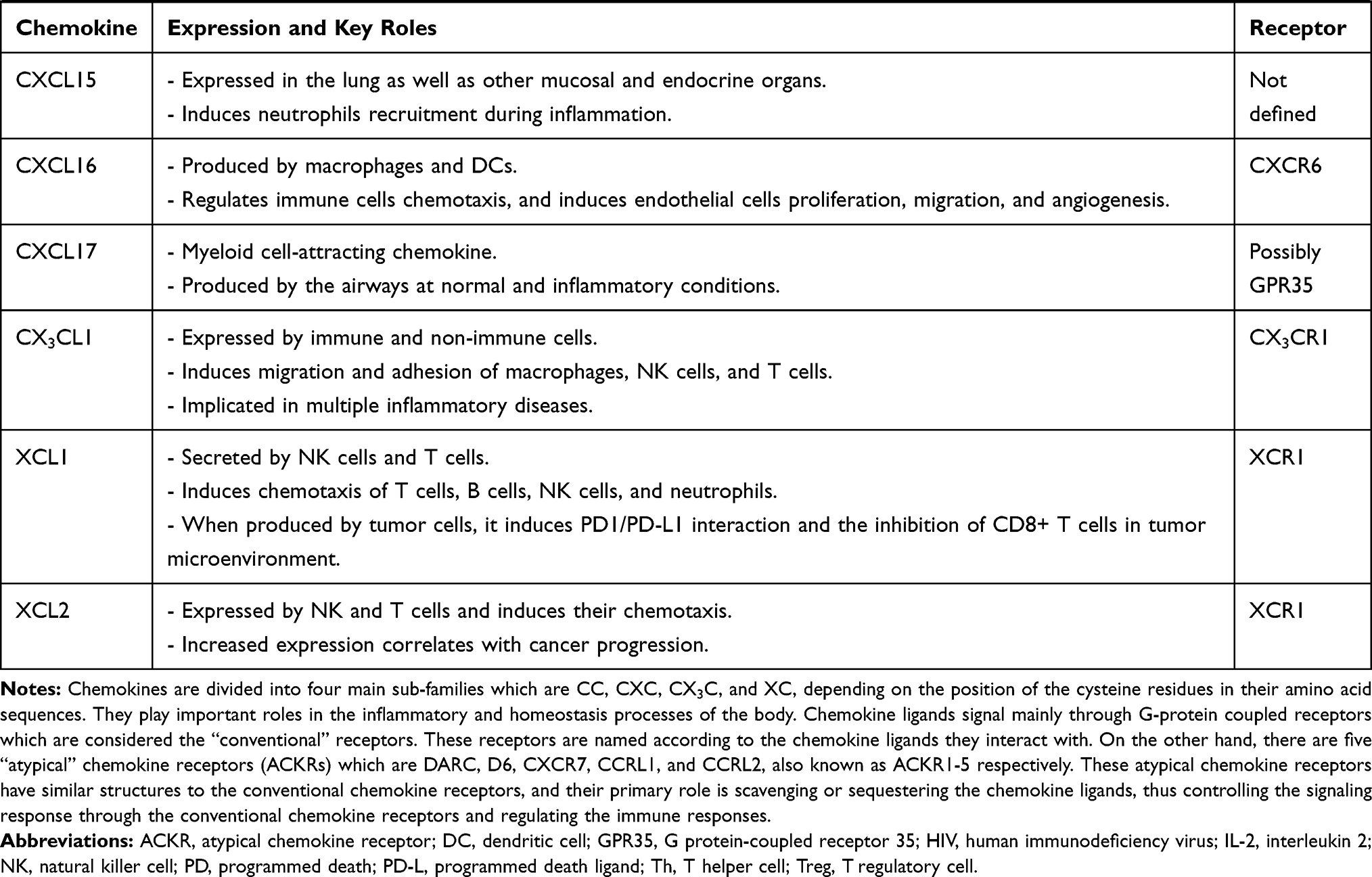 |
Table 1 Key Roles of Chemokines and Chemokine Receptors in Health and Diseases |
Role of Chemokines in the Pathophysiology of MS
Chemokines and chemokine receptors are expressed in different brain areas not only by immune cells but also by neurons and CNS resident cells such as astrocytes and oligodendrocytes. Chemokines and their receptors are increasingly expressed in MS patients.30 The chemokines/chemokine receptors axis expressed in T cells have the capability of recruiting inflammatory cells into CNS, which may lead to its destruction.26,30 Different chemokines play pathogenic roles in MS. These include CCL2, CCL3, CCL4, CCL5, CCL7, CCL8, CCL11, CCL17, CCL19, CCL21, CCL22, CXCL1, CXCL8, CXCL9, CXCL10, CXCL11, and CXCL16.31–43
Targeting of Chemokines and Chemokine Receptors Involved in MS or EAE
Targeting CCR1/CCL3 Axis
CCR1 and its chemokine ligands are implicated in chronic inflammatory disease.44 CCL3 (macrophage inflammatory protein-1α, MIP-1α) and CCL5 (regulated on activation, normal T cell expressed and secreted, RANTES) are the primary CCR1 ligands. Other chemokine ligands for CCR1 include CCL6, CCL9, CCL15, and CCL23.45,46 One study demonstrated that CCR1 is implicated in the pathogenesis of EAE using the CCR1 antagonist BX 471 which showed strong therapeutic effects in EAE animals.47
CCL3 was identified as a macrophage-derived inflammatory mediator48 and has been shown to be implicated in the pathophysiology of MS.30 For instance, treatment of EAE mice with CCL3 antibodies suppressed EAE and prevented the mononuclear cells from accumulating in the CNS.49 However, treatment with Met-RANTES, the CCR1 and CCR5 antagonist, did not affect leukocyte migration and decreased the severity of chronic-relapsing EAE mildly.50
Targeting CCR6/CCL20 Axis
The binding of CCL20 chemokine ligand to CCR6 has been shown to induce homeostatic and inflammatory functions. Elevated CCR6 and CCL20 expression levels were observed in the spinal cord and lymph nodes of EAE mice and were associated with the severity of the disease.51 Additionally, CCL20 binding CCR6 induced the trafficking of T helper 17 (Th17) cells into the CNS through the choroid plexus in the brain of EAE mice.52 CCR6-deficient mice or mice that were treated with a neutralizing anti-CCR6 showed resistance to EAE development.51 Also, elevated CCL20 serum levels were found in MS patients.53
Targeting CCRL2/CCL19 Axis
CC chemokine receptor-like 2 (CCRL2), also known as CRAM, is a member of the atypical chemokine receptor family and thus is a non-classical seven transmembrane spanning domain receptor. The ligand for CCRL2 is the homeostatic chemokine CCL19.54 CCRL2 is found in most immune cells such as dendritic cells (DCs), macrophages, neutrophils, lymphocytes, natural killer (NK) cells, and mast cells.55,56 It has been shown that CCRL2 is overexpressed in mouse microglia and astrocytes as well as being expressed by infiltrating macrophages in EAE.57,58 The knockout of CCRL2 in EAE mice resulted in defective acquisition of M2 phenotype markers by infiltrating macrophages and altered the M1/M2 ratio. This was responsible for the sustained T cell perivascular infiltration and inflammatory reaction resulting in delayed EAE recovery.59
Targeting CXCR3/CXCL10 Axis
CXCR3 is a chemokine receptor that is expressed on lymphocytes and plays a significant role in T cell trafficking and function. The gamma interferon-inducible chemokines which include CXCL9 (monokine induced by gamma interferon, Mig), CXCL10 (interferon gamma-induced protein 10, IP-10), and CXCL11 (interferon-inducible T-cell alpha chemoattractant, I-TAC) bind CXCR3.60,61 The increased expression of CXCR3 on peripheral blood CD4+ lymphocytes correlated with MS relapses.45 Additionally, CXCR3+ T cells levels are increased in RRMS patients during the MS attacks.45
The chemokine CXCL10 is a chemoattractant for activated T cells and NK cells through binding to CXCR3.62 CXCL10 levels in the CNS have been associated with EAE development and the recruitment of T cells expressing CXCR3 receptor into the CNS.63 In SJL mice treated with a neutralizing antibody against CXCL10, less accumulation of mononuclear inflammatory cells was observed along with a decreased incidence of EAE indicating that CXCL10 could contribute to CNS pathology via the recruitment and accumulation of T cells.63 Furthermore, upon ELISA analysis of the supernatant fluid collected from the cerebrospinal fluid (CSF) of 43 MS patients, elevated levels of CXCL10 were correlated with relapse phase of MS. Hence, CXCL10 or its receptor CXCR3 may be therapeutic targets for MS.64 In contrast, a study in C57BL/6J mice deficient in CXCR3 and CXCL10, found that these mice developed a severe myelin oligodendrocyte glycoprotein (MOG)-induced EAE disease.65
Targeting CXCL12/CXCR4/CXCR7 Axis
CXCL12 is continuously expressed in the healthy CNS where it is produced mainly by glial cells and neurons. In addition, its receptors CXCR4 and CXCR7 are abundantly expressed in diverse brain area.66 CXCL12 expression is upregulated in the MS lesions especially in astrocytes, that are likely to attract macrophages and lymphocytes into the CNS inflamed areas.67
CXCR4 is expressed on T and NK cells and has an important role in the homeostasis of the immune system.68 A study by Meiron et al69 demonstrated that the use of CXCR4 antagonist AMD3100, blocked CXCL12-induced production of IL-10 completely, thus interfering with the selection of the IL-10 producing regulatory T (Treg) cells. These results suggest that CXCL12 acts as a regulatory chemokine that redirects the polarization of effector Th1 cells into Treg cells which in turn suppress the autoimmune responses through IL-10 secretion.69
Moreover, treatment of C57BL/6 mice with the AMD3100 during EAE induction led to increased deteriorating of the disease along with parenchymal infiltration and demyelination.70 Along these findings, another study demonstrated that CXCL12 is up-regulated in spinal cord tissue after injection of MOG (which is the glycoprotein responsible for myelination of the nerves), in EAE-resistant Albino Oxford rats. Treatment of these rats with CXCL12 antagonist AMD3100 rendered them susceptible to EAE, indicating that CXCL12 is able to suppress inflammatory responses within the CNS.71
On the other hand, CXCR7 is a non-classical chemokine receptor and was proposed to have more than five natural ligands, although research focused on CXCL11 and CXCL12.72 It was observed that the main function of CXCR7 is to sequester CXCL12, thus regulating the function of CXCR4.72 Treatment of EAE mice with NSC-87877 a SHP-2 inhibitor induced the expression of CXCR7 which resulted in decreased T cell trafficking in response to CXCL12.73 This finding suggests that CXCR7, present on CD8+ T cells, is upregulated as a result of SHP-2 inhibition and is negatively regulated in these cells.73 It is worth mentioning that the initiation of treatment of NSC-87877 at day 0 after immunization blocked the incidence of EAE effectively, while that starting from day 13 did not. These observations suggest that NSC-87877 treatment does not change the encephalitogenic response of peripheral T cells but affects their ability to migrate into the CNS and cause inflammation.73
It has also been demonstrated that CXCL12 inhibits the trafficking of infiltrating cells from the perivascular space into the brain parenchyma, thereby limiting inflammation.74 A study reported loss of CXCL12 from abluminal surfaces of the blood–brain barrier in MS lesions.74 Therefore, treatment with a CXCR7 antagonist CCX771 prevented CXCR7 from sequestering CXCL12, resulting in elevated abluminal levels of CXCL12 and reduced leukocyte infiltration, which ameliorate the severity of EAE.75
Targeting CXCL13/CXCR5 Axis
CXCL13 is a homeostatic chemokine that is continuously expressed in lymphoid tissues, which has a vital role in activating the migration of lymphocytes and antigen-presenting cells.76,77 Upon binding of CXCL13 to CXCR5, CNS inflammation developed.78 In MS patients, the levels of CXCL13 were found to be significantly increased and consequently, CXCL13 is considered a biomarker for the disease severity.79,80
Blocking CXCL13 binding to its receptor results in inhibition of B, T follicular helper (Tfh), and Th17 cell migration and subsequent development of tissue inflammation. The use of anti-human CXCL13 antibody, MAb 5261, inhibited CXCL13-induced migration of B cells into secondary lymphoid organs resulting in halting the progression and severity of Th17-induced EAE in SJL mice.81
Targeting CXCL16/CXCR6 Axis
The CXC chemokine ligand CXCL16 is the ligand for CXC chemokine receptor 6 (CXCR6). CXCL16 in a soluble form acts as a chemoattractant for activated CD8+ T cells, NKT cells and Th1 cells that express CXCR6.82 CXCL16 is highly expressed in the brain during MS.83 Serum levels of CXCL16 reflect disease activity in MS, suggesting that CXCL16 could be a potential marker of MS disease.43 Moreover, animals treated with anti-CXCL16 antibodies were found to be resistant to the induction of EAE.84
Effects of the Current MS and EAE Modulatory Drugs on the Chemokine System
Currently, several drugs are used for the treatment of MS or EAE, these include: 1) Steroids such as methylprednisolone (MP); 2) Disease-modifying agents such as glatiramer acetate (GA), 4-methylumbelliferone (4MU) and interferon-beta (IFN-β); 3) Immunosuppressants such as mitoxantrone and cladribine; and 4) Monoclonal antibodies such as natalizumab and rituximab.85–88 Some of these modulatory agents were found to have regulatory effects on the chemokines and chemokine receptor levels in MS patients or EAE models.26,30 In the following section, we focus on the effects of these agents on modifying the expression of chemokines and chemokine receptors.
Role of Methylprednisolone on the Modulation of Chemokines and Chemokine Receptors
Methylprednisolone (MP) is a glucocorticosteroid drug that has been used for the treatment of MS patients. It is an anti-inflammatory drug that inhibits the activation of T cells and their trafficking into the CNS while reducing the inflammatory cytokine levels.89,90 Treatment of MS patients in the active phase with MP decreased the migratory ability of CD4+CCR5+ T cells.91 Additionally, the level of CXCL10 was reduced in the sera of MS patients after intravenous administration of MP.92 Moreover, the application of MP in vitro and in vivo increased the chemotaxis of monocytes towards CCL2, CCL5, and CX3CL1 in MS patients compared to healthy controls. This boost was independent of chemokine receptor levels, suggesting that MP induces monocytes polarization towards the anti-inflammatory form and enhances their trafficking into inflamed CNS, where they plausibly suppress the pathogenic immune responses.93 Additionally, a study by Sellebjerg et al94 found that CXCL13 concentrations in CSF of MS patients were reduced after treatment with high-dose MP and natalizumab. On the other hand, CXCR1 and CXCR2, which are up-regulated in MS patients, showed a further increase in their expression upon MP treatment.95
Effect of Glatiramer Acetate (GA) on Chemokines/Chemokine Receptors and NK Cell Cytolytic Activity
Glatiramer acetate (GA), is an immunomodulating drug used for the treatment of MS patients.96 This drug increased the levels of the chemokine receptor CCR7 and decreased the levels of CCR5, CXCR3 and CXCR6, on T cells.31 Further, GA activates in vitro human NK cell lysis of DCs.97 Similarly, this drug stimulates NK cell killing of DCs in GA-treated MS patients.98 In mice, GA reduces the EAE clinical score corroborated with enhancing NK cell cytolysis of DCs, hence impeding antigen presentation to autoreactive T cells.99
Role of IFN-β on the Modulation of Chemokines and Chemokine Receptors
IFN-β was the first agent that was approved for treatment of relapsing-remitting MS.100 IFN-β-1b therapy reduced the rate of clinical exacerbations and the number of lesions in brain magnetic resonance imaging (MRI), and it has been established as an effective drug for treatment of RRMS patients.101
Treatment with IFN-β reduced the expression of CCR5 on T cells of MS patients, whereas treatment of T cells with IFN-β in vitro, inhibited their expression of this chemokine receptor as well its ligands CCL5 and CCL3.102 Furthermore, the expression of CXCR3 on CD4+ and CD8+ T cells was considerably decreased in RRMS patients after treatment with IFN-β.103 However, IFN-β induced a transient strong increase of CXCL10 level in MS patients.104 Moreover, it was found that in vitro treatment of peripheral blood mononuclear cells with IFN-β-1b lowered the production of CXCL8, CXCL9, CXCL10, CCL2, and CCL7 chemokines.105
Effect of 4-Methylumbelliferone on Chemokines and Chemokine Receptors
4-methylumbelliferone (4-MU) is a drug that hinders the synthesis of hyaluronan, which is an extracellular glycosaminoglycan that is present in the pericellular and extracellular matrix of vertebrate tissues and is implicated in autoimmunity. In MS patients, hyaluronan has proinflammatory roles in the CNS, as it accumulates in demyelinated MS lesions, inducing antigen presentation by antigen-presenting cells leading to T cell activation and proliferation.106 4-MU inhibits hyaluronan production in vitro and in vivo.107,108 In EAE, this drug reduces the polarization of Th1 phenotype and increases FoxP3+ Treg cells that are correlated with the suppression of EAE. Moreover, 4-MU inhibits the reactive response of astrocytes, immunocompetent resident cells of the CNS, and prevents activated T cells from migrating into the CNS.85
Additionally, spinal CXCL12 protein levels were raised in non-inflamed CNS tissues and in inflamed CNS tissues of lipopolysaccharide (LPS)-injected mice after the oral 4MU administration. Feeding EAE mice with 4MU induced protective effects against EAE as the disease clinical scores were considerably lower in 4MU-fed mice compared to untreated EAE mice.106
Role of Mitoxantrone in Modulating Chemokines and Chemokine Receptors
Mitoxantrone (Novantrone) is an anticancer drug that inhibits the DNA topoisomerase II activity which results in preventing the DNA synthesis process, and inhibiting cellular proliferation.109 This drug was approved for the treatment of RRMS, SPMS, and PRMS, where it acts as an immunosuppressant inhibiting immune cells proliferation, their antigen presentation and inflammatory cytokines secretion.110 A study by Scott et al111 showed that intravenous treatment of MS patients with mitoxantrone reduced the relapse rate and delayed the progression of the disease.
It has been demonstrated that therapy of MS patients with mitoxantrone resulted in decreased CXCR1 and CXCR2 expression on peripheral blood mononuclear cells, suggesting a pathogenic role for these chemokine receptors in MS that can be inhibited by this drug.95 Another study indicated that in vitro treatment of primary astrocytes with mitoxantrone, inhibited LPS-induced production of CCL2 and other inflammatory molecules.112 On the other hand, it was observed that the chemokine receptor CCR2 expression was significantly upregulated on monocytes of two out of eight mitoxantrone treated patients who had active inflammation when compared to placebo patients who showed no significant changes in the expression of this chemokine receptor.113
Effects of Cladribine on the Modulation of Chemokines and Chemokine Receptors
Cladribine (2-chlorodeoxyadenosine) is a deoxyadenosine analogue pro-drug that selectively depletes lymphocytes such as B and T cells. It was identified as an anticancer drug for the treatment of B and T cell lymphoid malignancies, but was later used for the treatment of RRMS due to its immunosuppressive activity for these immune cells that play a significant role in MS pathogenesis.114 Regarding its effects on the chemokine system of MS patients, it was demonstrated that treatment of RRMS patients with Cladribine resulted in declined levels of CXCL8 in their CSF, and a significant decrease in CCL5 levels in the CSF and serum of those patients.115
Effects of Natalizumab on Chemokines and Chemokine Receptors
Natalizumab (Tysabri) is a humanized monoclonal antibody that selectively binds to α4 integrins expressed on the surface of human leukocytes.116 The drug is approved in North America for relapsing MS and in Europe for the treatment of RRMS.117 Studies in relapsing MS patients indicated that natalizumab markedly reduced the relapse rate, and the accumulation of new brain MRI lesions of these patients.118,119 In another study, natalizumab suppressed the development of new gadolinium-enhanced lesions in patients with relapsing MS and reduced the rate of brain volume loss and enhanced brain tissue integrity in RRMS.120 It was also demonstrated that RRMS patients receiving natalizumab therapy, had significantly reduced levels of plasma pro-inflammatory cytokines and chemokines. After natalizumab treatment, there was a significant decline in the levels of chemokines correlated with Th1 chemokines such as CXCL9, CXCL10, and CXCL11, or Th2 chemokine CCL22 in the CSF, in addition to the expected decline of the proinflammatory cytokines.121
Rituximab Effects on the Chemokines and Chemokine Receptors
Rituximab is a chimeric monoclonal antibody that binds to the protein CD20 and depletes B cells from the circulation.122 This drug was initially used to treat lymphomas but currently is being used for the treatment of certain autoimmune diseases.123 The effect of rituximab results from its regulatory effect on the cell cycle, including apoptosis stimulation, complement-dependent B cell lysis, and antibody-dependent cell-mediated cytotoxicity.124 Recent studies showed that rituximab not only affects B cells but has another possible mechanism of action against CD20+ T cells in MS patients,125 or in EAE mice.88
Several studies indicated the high efficacy of rituximab in relapsing-remitting MS patients. Other studies have showed the efficacy of rituximab in treating progressive MS patients.126,127 Treatment of MS patients with rituximab affects chemokine levels since infusion therapy resulted in a significant decrease of CXCL13 and CCL19 in those patients.128,129 Further, the levels of CXCL8 and CXCL10 chemokines decreased significantly after treatment with rituximab.130
Conclusions
Several chemokines and chemokine receptors have been involved in the pathophysiology of MS, some of which have been targeted directly (Table 2) or modulated indirectly by immunomodulatory agents and drugs (Figure 1). This targeting strategy demonstrates promising future approaches in the treatment of MS and EAE. However, due to the promiscuity and the multiple functions of the chemokine system, targeting chemokines and their receptors with selective agents such as antibodies or receptor blockers could present a challenge and might not be effective.131,132 On the other hand, the general chemokine blockage might affect the host defense dramatically.133 Moreover, chemotaxis results obtained from studies on EAE mouse model may not be fully applicable for MS disease in humans, as sometimes the findings of studies by different research groups can be contradictory. Further research should be done to understand the precise functions of the chemokines and chemokine receptors that are involved in this disease, in order to develop better therapeutic strategies to treat MS patients.
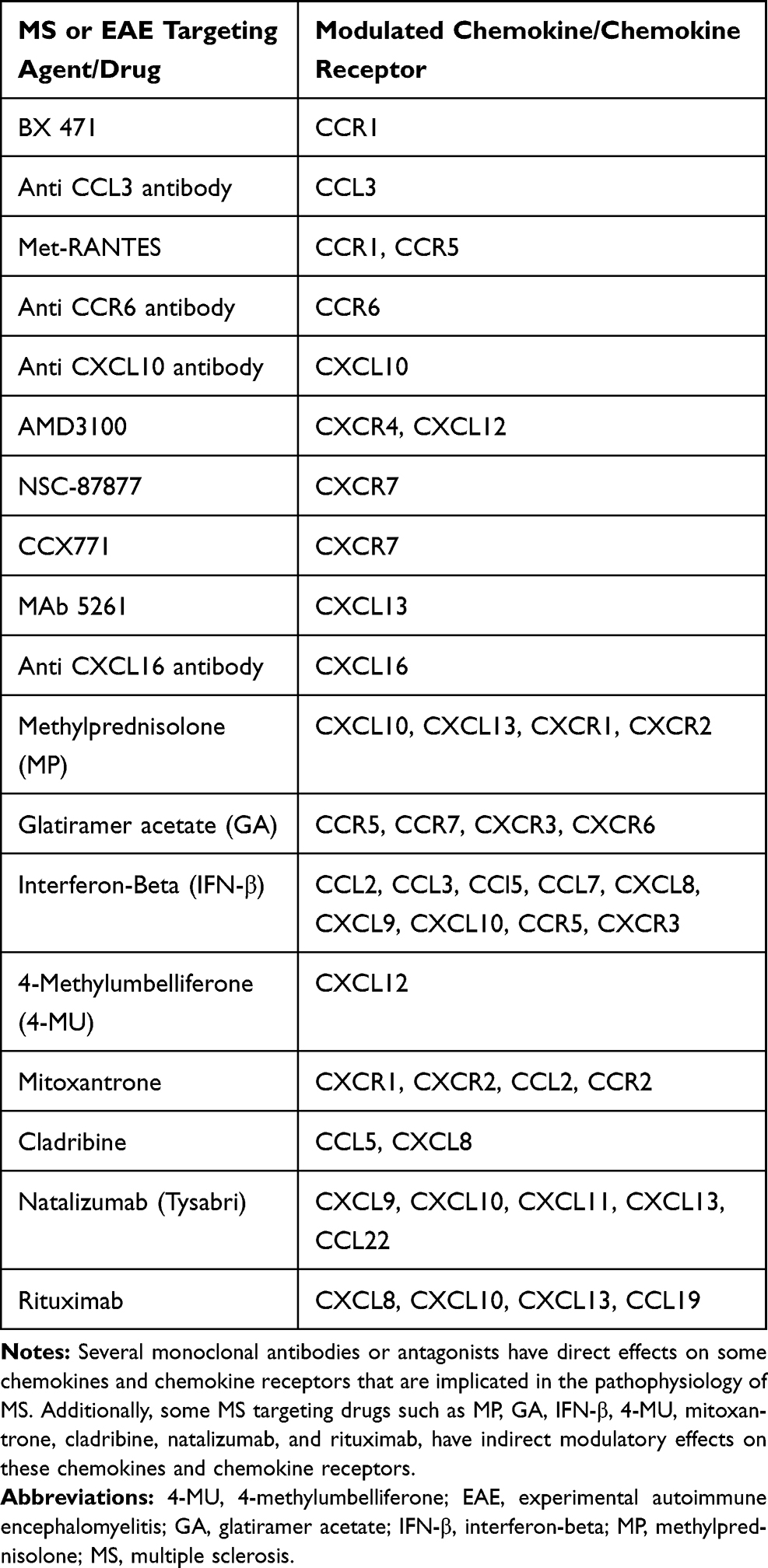 |
Table 2 Agents and Drugs That Have Modulatory Effects on Chemokines and Chemokine Receptors Levels in MS Patients, in vivo EAE Mouse Model, or in vitro Assays |
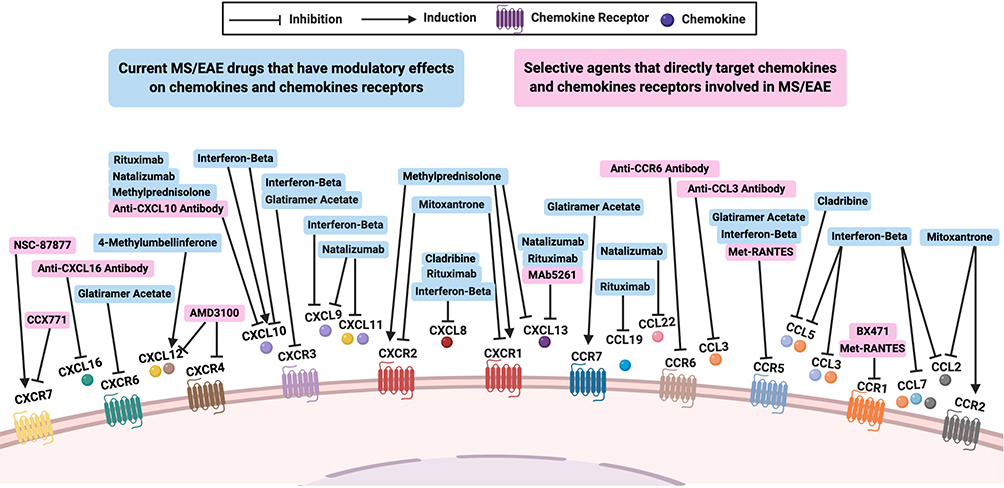 |
Figure 1 The effects of modulating chemokines and chemokine receptors involved in MS or EAE. Several studies investigated the roles of chemokines and chemokine receptors on the pathogenesis of multiple sclerosis (MS) and experimental autoimmune encephalomyelitis (EAE) through targeting them directly (pink boxes), with specific agents acting as antagonists, receptors blockers, or antibodies. Moreover, some current MS or EAE drugs could indirectly modulate the expression of these chemokines and chemokine receptors (blue boxes). |
Abbreviations
4-MU, 4-methylumbelliferone; ACKR, atypical chemokine receptor; APCs, antigen presenting cells; CCL/R, CC chemokine ligand/receptor; CCRL2, CC chemokine receptor-like 2; CNS, central nervous system; CSF, cerebrospinal fluid; CX3CL/R, CX3C chemokine ligand/receptor; CXCL/R, CXC chemokine ligand/receptor; DCs, dendritic cells; EAE, experimental autoimmune encephalitis; GA, glatiramer acetate; GD, Graves’ disease; GPR35, G protein-coupled receptor 35; HIV, human immunodeficiency virus; IL, interleukin; I-TAC, interferon-inducible T-cell alpha chemoattractant; IFN-β, interferon-beta; IP-10, interferon gamma-induced protein 10; LPS, lipopolysaccharide; MIG, monokine induced by gamma interferon; MIP-1α, macrophage inflammatory protein-1α; MOG, myelin oligodendrocyte glycoprotein; MP, methylprednisolone; MRI, magnetic resonance imaging; MS, multiple sclerosis; NK, natural killer; PD, programmed death; PD-L, programmed death ligand; PPMS, primary progressive MS; PRMS, progressive relapsing MS; RA, rheumatoid arthritis; RANTES, regulated on activation, normal T cell expressed and secreted; RRMS, relapsing remitting MS; SDF-1, stromal cell-derived factor 1; SLE, systemic lupus erythematosus; SPMS, secondary progressive MS; Treg, T regulatory; Th, T helper; Tfh, T follicular helper; XCL/R, XC chemokine ligand/receptor.
Acknowledgments
The work in the authors’ laboratory is supported by a grant from Terry Fox Foundation number MISC051.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Hestvik ALK. The double-edged sword of autoimmunity: lessons from multiple sclerosis. Toxins. 2010;2(4):856–877. doi:10.3390/toxins2040856
2. Hachim MY, Elemam NM, Maghazachi AA. The beneficial and debilitating effects of environmental and microbial toxins, drugs, organic solvents and heavy metals on the onset and progression of multiple sclerosis. Toxins. 2019;11(3):147–163. doi:10.3390/toxins11030147
3. Miller DH. Imaging multiple sclerosis outside the brain. Funct Neurol. 2013;28(2):72.
4. Nazem G, Shahnaz R, Elham N. Multiple sclerosis: pathogenesis, symptoms, diagnoses and cell-based therapy. Cell J. 2017;19(1):1–10. doi:10.22074/cellj.2016.4867
5. Loma I, Heyman R. Multiple sclerosis: pathogenesis and treatment. Curr Neuropharmacol. 2011;9(3):409–416. doi:10.2174/157015911796557911
6. Burtchell J, Fetty K, Miller K, Minden K, Kantor D. Two sides to every story: perspectives from four patients and a healthcare professional on multiple sclerosis disease progression. Neurol Ther. 2019;8(2):185–205. doi:10.1007/s40120-019-0141-4
7. Constantinescu CS, Farooqi N, O’Brien K, Gran B. Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). Br J Pharmacol. 2011;164(4):1079–1106. doi:10.1111/j.1476-5381.2011.01302.x
8. Raman D, Sobolik-Delmaire T, Richmond A. Chemokines in health and disease. Exp Cell Res. 2011;317(5):575–589. doi:10.1016/j.yexcr.2011.01.005
9. Nomiyama H, Osada N, Yoshie O. A family tree of vertebrate chemokine receptors for a unified nomenclature. Dev Comp Immunol. 2011;35(7):705–715. doi:10.1016/j.dci.2011.01.019
10. Nomiyama H, Osada N, Yoshie O. The evolution of mammalian chemokine genes. Cytokine Growth Factor Rev. 2010;21(4):253–262. doi:10.1016/j.cytogfr.2010.03.004
11. Maghazachi AA. G protein-coupled receptors in natural killer cells. J Leukoc Biol. 2003;74(1):16–24. doi:10.1189/jlb.0103019
12. Li X, Qi Y, Ma X, et al. Chemokine (C-C motif) ligand 20, a potential biomarker for Graves’ disease, is regulated by osteopontin. PLoS One. 2013;8(5):e64277. doi:10.1371/journal.pone.0064277
13. Sellam J, Rouanet S, Hendel-Chavez H, et al. CCL19, a B-cell chemokine, is related to the decrease of blood memory B-cells and predicts the clinical response to rituximab in rheumatoid arthritis. Arthritis Rheum. 2013;65(9):2253–2261. doi:10.1002/art.38023
14. Schall TJ, Proudfoot AE. Overcoming hurdles in developing successful drugs targeting chemokine receptors. Nat Rev Immunol. 2011;11(5):355–363. doi:10.1038/nri2972
15. Schall TJ, Bacon KB. Chemokines, leukocyte trafficking, and inflammation. Curr Opin Immunol. 1994;6(6):865–873. doi:10.1016/0952-7915(94)90006-x
16. Murphy PM. Viral chemokine receptors. Front Immunol. 2015;6:281. doi:10.3389/fimmu.2015.00281
17. Zlotnik A, Yoshie O. The chemokine superfamily revisited. Immunity. 2012;36(5):705–716. doi:10.1016/j.immuni.2012.05.008
18. Fantuzzi L, Tagliamonte M, Gauzzi MC, Lopalco L. Dual CCR5/CCR2 targeting: opportunities for the cure of complex disorders. Cell Mol Life Sci. 2019;76(24):4869–4886. doi:10.1007/s00018-019-03255-6
19. Aldinucci D, Borghese C, Casagrande N. The CCL5/CCR5 axis in cancer progression. Cancers. 2020;12(7):E1765. doi:10.3390/cancers12071765
20. Zimmerman KA, Hopp K, Mrug M. Role of chemokines, innate and adaptive immunity. Cell Signal. 2020;73:109647. doi:10.1016/j.cellsig.2020.109647
21. Capucetti A, Albano F, Bonecchi R. Multiple roles for chemokines in neutrophil biology. Front Immunol. 2020;11:1259. doi:10.3389/fimmu.2020.01259
22. Coperchini F, Chiovato L, Croce L, Magri F, Rotondi M. The cytokine storm in COVID-19: an overview of the involvement of the chemokine/chemokine-receptor system. Cytokine Growth Factor Rev. 2020;53:25–32. doi:10.1016/j.cytogfr.2020.05.003
23. Maghazachi AA. Role of chemokines in the biology of natural killer cells. Curr Top Microbiol Immunol. 2010;341:37–58. doi:10.1007/82_2010_20
24. Sokol CL, Luster AD. The chemokine system in innate immunity. Cold Spring Harb Perspect Biol. 2015;7(5):a016303. doi:10.1101/cshperspect.a016303
25. Gustavsson M. New insights into the structure and function of chemokine receptor: chemokine complexes from an experimental perspective. J Leukoc Biol. 2020;107(6):1115–1122. doi:10.1002/JLB.2MR1219-288R
26. Karpus WJ. Cytokines and chemokines in the pathogenesis of experimental autoimmune encephalomyelitis. J Immunol. 2020;204(2):316–326. doi:10.4049/jimmunol.1900914
27. Elemam NM, Hannawi S, Maghazachi AA. Role of chemokines and chemokine receptors in rheumatoid arthritis. Immunotargets Ther. 2020;9:43–56. doi:10.2147/ITT.S243636
28. Miyabe Y, Lian J, Miyabe C, Luster AD. Chemokines in rheumatic diseases: pathogenic role and therapeutic implications. Nat Rev Rheumatol. 2019;15(12):731–746. doi:10.1038/s41584-019-0323-6
29. Miao M, De Clercq E, Li G. Clinical significance of chemokine receptor antagonists. Expert Opin Drug Metab Toxicol. 2020;16(1):11–30. doi:10.1080/17425255.2020.1711884
30. Cui LY, Chu SF, Chen NH. The role of chemokines and chemokine receptors in multiple sclerosis. Int Immunopharmacol. 2020;83:106314. doi:10.1016/j.intimp.2020.106314
31. Cheng W, Chen G. Chemokines and chemokine receptors in multiple sclerosis. Mediators Inflamm. 2014;2014:659206. doi:10.1155/2014/659206
32. Sørensen TL, Ransohoff RM, Strieter RM, Sellebjerg F. Chemokine CCL2 and chemokine receptor CCR2 in early active multiple sclerosis. Eur J Neurol. 2004;11(7):445–449. doi:10.1111/j.1468-1331.2004.00796.x
33. Szczuciński A, Losy J. Chemokines and chemokine receptors in multiple sclerosis. Potential targets for new therapies. Acta Neurol Scand. 2007;115(3):137–146. doi:10.1111/j.1600-0404.2006.00749.x
34. Pittaluga A. CCL5-glutamate cross-talk in astrocyte-neuron communication in multiple sclerosis. Front Immunol. 2017;8:1079. doi:10.3389/fimmu.2017.01079
35. Banisor I, Leist TP, Kalman B. Involvement of beta-chemokines in the development of inflammatory demyelination. J Neuroinflammation. 2005;2(1):7. doi:10.1186/1742-2094-2-7
36. Bielecki B, Jatczak-Pawlik I, Wolinski P, Bednarek A, Glabinski A. Central nervous system and peripheral expression of CCL19, CCL21 and their receptor CCR7 in experimental model of multiple sclerosis. Arch Immunol Ther Exp. 2015;63(5):367–376. doi:10.1007/s00005-015-0339-9
37. Tejera-Alhambra M, Casrouge A, de Andrés C, et al. Plasma biomarkers discriminate clinical forms of multiple sclerosis. PLoS One. 2015;10(6):e0128952. doi:10.1371/journal.pone.0128952
38. Scheu S, Ali S, Ruland C, Arolt V, Alferink J. The C-C chemokines CCL17 and CCL22 and their receptor CCR4 in CNS autoimmunity. Int J Mol Sci. 2017;18(11):2306. doi:10.3390/ijms18112306
39. Grist JJ, Marro BS, Skinner DD, et al. Induced CNS expression of CXCL1 augments neurologic disease in a murine model of multiple sclerosis via enhanced neutrophil recruitment. Eur J Immunol. 2018;48(7):1199–1210. doi:10.1002/eji.201747442
40. Lund BT, Ashikian N, Ta HQ, et al. Increased CXCL8 (IL-8) expression in multiple sclerosis. J Neuroimmunol. 2004;155(1–2):161–171. doi:10.1016/j.jneuroim.2004.06.008
41. Koper OM, Kamińska J, Sawicki K, Kemona H. CXCL9, CXCL10, CXCL11, and their receptor (CXCR3) in neuroinflammation and neurodegeneration. Adv Clin Exp Med. 2018;27(6):849–856. doi:10.17219/acem/68846
42. Sørensen TL, Trebst C, Kivisäkk P, et al. Multiple sclerosis: a study of CXCL10 and CXCR3 co-localization in the inflamed central nervous system. J Neuroimmunol. 2002;127(1–2):59–68. doi:10.1016/s0165-5728(02)00097-8
43. Holmøy T, Løken-Amsrud KI, Bakke SJ, et al. Inflammation markers in multiple sclerosis: CXCL16 reflects and may also predict disease activity. PLoS One. 2013;8(9):e75021. doi:10.1371/journal.pone.0075021
44. Berahovich RD, Miao Z, Wang Y, Premack B, Howard MC, Schall TJ. Proteolytic activation of alternative CCR1 ligands in inflammation. J Immunol. 2005;174(11):7341–7351. doi:10.4049/jimmunol.174.11.7341
45. Balashov KE, Rottman JB, Weiner HL, Hancock WW. CCR5+ and CXCR3+ T cells are increased in multiple sclerosis and their ligands MIP-1alpha and IP-10 are expressed in demyelinating brain lesions. Proc Natl Acad Sci U S A. 1999;96(12):6873–6878. doi:10.1073/pnas.96.12.6873
46. Furuichi K, Gao JL, Horuk R, Wada T, Kaneko S, Murphy PM. Chemokine receptor CCR1 regulates inflammatory cell infiltration after renal ischemia-reperfusion injury. J Immunol. 2008;181(12):8670–8676. doi:10.4049/jimmunol.181.12.8670
47. Liang M, Mallari C, Rosser M, et al. Identification and characterization of a potent, selective, and orally active antagonist of the CC chemokine receptor-1. J Biol Chem. 2000;275(25):19000–19008. doi:10.1074/jbc.M001222200
48. Baba T, Mukaida N. Role of macrophage inflammatory protein (MIP)-1α/CCL3 in leukemogenesis. Mol Cell Oncol. 2014;1(1):e29899. doi:10.4161/mco.29899
49. Karpus WJ, Lukacs NW, McRae BL, Strieter RM, Kunkel SL, Miller SD. An important role for the chemokine macrophage inflammatory protein-1 alpha in the pathogenesis of the T cell-mediated autoimmune disease, experimental autoimmune encephalomyelitis. J Immunol. 1995;155(10):5003–5010.
50. Matsui M, Weaver J, Proudfoot AE, et al. Treatment of experimental autoimmune encephalomyelitis with the chemokine receptor antagonist Met-RANTES. J Neuroimmunol. 2002;128(1–2):16–22. doi:10.1016/s0165-5728(02)00121-2
51. Liston A, Kohler RE, Townley S, et al. Inhibition of CCR6 function reduces the severity of experimental autoimmune encephalomyelitis via effects on the priming phase of the immune response. J Immunol. 2009;182(5):3121–3130. doi:10.4049/jimmunol.0713169
52. Reboldi A, Coisne C, Baumjohann D, et al. C-C chemokine receptor 6-regulated entry of TH-17 cells into the CNS through the choroid plexus is required for the initiation of EAE. Nat Immunol. 2009;10(5):514–523. doi:10.1038/ni.1716
53. Jafarzadeh A, Bagherzadeh S, Ebrahimi HA, et al. Higher circulating levels of chemokine CCL20 in patients with multiple sclerosis: evaluation of the influences of chemokine gene polymorphism, gender, treatment and disease pattern. J Mol Neurosci. 2014;53(3):500–505. doi:10.1007/s12031-013-0214-2
54. Leick M, Catusse J, Follo M, et al. CCL19 is a specific ligand of the constitutively recycling atypical human chemokine receptor CRAM-B. Immunology. 2010;129(4):536–546. doi:10.1111/j.1365-2567.2009.03209.x
55. Salvi V, Sozio F, Sozzani S, Del Prete A. Role of atypical chemokine receptors in microglial activation and polarization. Front Aging Neurosci. 2017;9:148. doi:10.3389/fnagi.2017.00148
56. Otero K, Vecchi A, Hirsch E, et al. Nonredundant role of CCRL2 in lung dendritic cell trafficking. Blood. 2010;116(16):2942–2949. doi:10.1182/blood-2009-12-259903
57. Zuurman MW, Heeroma J, Brouwer N, Boddeke HW, Biber K. LPS-induced expression of a novel chemokine receptor (L-CCR) in mouse glial cells in vitro and in vivo. Glia. 2003;41(4):327–336. doi:10.1002/glia.10156
58. Brouwer N, Zuurman MW, Wei T, Ransohoff RM, Boddeke HW, Biber K. Induction of glial L-CCR mRNA expression in spinal cord and brain in experimental autoimmune encephalomyelitis. Glia. 2004;46(1):84–94. doi:10.1002/glia.10352
59. Mazzon C, Zanotti L, Wang L, et al. CCRL2 regulates M1/M2 polarization during EAE recovery phase. J Leukoc Biol. 2016;99(6):1027–1033. doi:10.1189/jlb.3MA0915-444RR
60. Kuo PT, Zeng Z, Salim N, Mattarollo S, Wells JW, Leggatt GR. The role of CXCR3 and its chemokine ligands in skin disease and cancer. Front Med. 2018;5:271. doi:10.3389/fmed.2018.00271
61. Tokunaga R, Zhang W, Naseem M, et al. CXCL9, CXCL10, CXCL11/CXCR3 axis for immune activation - A target for novel cancer therapy. Cancer Treat Rev. 2018;63:40–47. doi:10.1016/j.ctrv.2017.11.007
62. Maghazachi AA, Skalhegg BS, Rolstad B, Al-Aoukaty A. Interferon-inducible protein-10 and lymphotactin induce the chemotaxis and mobilization of intracellular calcium in natural killer cells through pertussis toxin-sensitive and -insensitive heterotrimeric G-proteins. FASEB J. 1997;11(10):765–774. doi:10.1096/fasebj.11.10.9271361
63. Fife BT, Kennedy KJ, Paniagua MC, et al. CXCL10 (IFN-γ-inducible protein-10) control of encephalitogenic CD4+ T cell accumulation in the central nervous system during experimental autoimmune encephalomyelitis. J Immunol. 2001;166(12):7617–7624. doi:10.4049/jimmunol.166.12.7617
64. Mahad DJ, Howell SJ, Woodroofe MN. Expression of chemokines in the CSF and correlation with clinical disease activity in patients with multiple sclerosis. J Neurol Neurosurg Psychiatry. 2002;72(4):498–502. doi:10.1136/jnnp.72.4.498
65. Müller M, Carter SL, Hofer MJ, et al. CXCR3 signaling reduces the severity of experimental autoimmune encephalomyelitis by controlling the parenchymal distribution of effector and regulatory T cells in the central nervous system. J Immunol. 2007;179(5):2774–2786. doi:10.4049/jimmunol.179.5.2774
66. Guyon A. CXCL12 chemokine and its receptors as major players in the interactions between immune and nervous systems. Front Cell Neurosci. 2014;8:65. doi:10.3389/fncel.2014.00065
67. Calderon TM, Eugenin EA, Lopez L, et al. A role for CXCL12 (SDF-1alpha) in the pathogenesis of multiple sclerosis: regulation of CXCL12 expression in astrocytes by soluble myelin basic protein. J Neuroimmunol. 2006;177(1–2):27–39. doi:10.1016/j.jneuroim.2006.05.003
68. Maghazachi AA. Role of the heterotrimeric G proteins in stromal-derived factor-1alpha-induced natural killer cell chemotaxis and calcium mobilization. Biochem Biophys Res Commun. 1997;236(2):270–274. doi:10.1006/bbrc.1997.6937
69. Meiron M, Zohar Y, Anunu R, Wildbaum G, Karin N. CXCL12 (SDF-1α) suppresses ongoing experimental autoimmune encephalomyelitis by selecting antigen-specific regulatory T cells. J Exp Med. 2008;205(11):2643–2655. doi:10.1084/jem.20080730
70. McCandless EE, Wang Q, Woerner BM, Harper JM, Klein RS. CXCL12 limits inflammation by localizing mononuclear infiltrates to the perivascular space during experimental autoimmune encephalomyelitis. J Immunol. 2006;177(11):8053–8064. doi:10.4049/jimmunol.177.11.8053
71. Miljković D, Stanojević Z, Momcilović M, Odoardi F, Flügel A, Mostarica-Stojković M. CXCL12 expression within the CNS contributes to the resistance against experimental autoimmune encephalomyelitis in Albino Oxford rats. Immunobiology. 2011;216(9):979–987. doi:10.1016/j.imbio.2011.03.013
72. Wang C, Chen W, Shen J. CXCR7 targeting and its major disease relevance. Front Pharmacol. 2018;9:641. doi:10.3389/fphar.2018.00641
73. Luo Q, Sun Y, Gong FY, et al. Blocking initial infiltration of pioneer CD8+ T-cells into the CNS via inhibition of SHP-2 ameliorates experimental autoimmune encephalomyelitis in mice. Br J Pharmacol. 2014;171(7):1706–1721. doi:10.1111/bph.12565
74. McCandless EE, Piccio L, Woerner BM, et al. Pathological expression of CXCL12 at the blood-brain barrier correlates with severity of multiple sclerosis. Am J Pathol. 2008;172(3):799–808. doi:10.2353/ajpath.2008.070918
75. Cruz-Orengo L, Holman DW, Dorsey D, et al. CXCR7 influences leukocyte entry into the CNS parenchyma by controlling abluminal CXCL12 abundance during autoimmunity. J Exp Med. 2011;208(2):327–339. doi:10.1084/jem.20102010
76. Irani DN. Regulated production of CXCL13 within the central nervous system. J Clin Cell Immunol. 2016;7(5):460. doi:10.4172/2155-9899.1000460
77. Kazanietz MG, Durando M, Cooke M. CXCL13 and its receptor CXCR5 in cancer: inflammation, immune response, and beyond. Front Endocrinol. 2019;10:471. doi:10.3389/fendo.2019.00471
78. Griffin DE. Cytokines and Chemokines. In: Encyclopedia of Virology.
79. Khademi M, Kockum I, Andersson ML, et al. Cerebrospinal fluid CXCL13 in multiple sclerosis: a suggestive prognostic marker for the disease course. Mult Scler. 2011;17(3):335–343. doi:10.1177/1352458510389102
80. Alvarez E, Piccio L, Mikesell RJ, et al. CXCL13 is a biomarker of inflammation in multiple sclerosis, neuromyelitis optica, and other neurological conditions. Mult Scler. 2013;19(9):1204–1208. doi:10.1177/1352458512473362
81. Klimatcheva E, Pandina T, Reilly C, et al. CXCL13 antibody for the treatment of autoimmune disorders. BMC Immunol. 2015;16(1):6. doi:10.1186/s12865-015-0068-1
82. Tohyama M, Sayama K, Komatsuzawa H, et al. CXCL16 is a novel mediator of the innate immunity of epidermal keratinocytes. Int Immunol. 2007;19(9):1095–1102. doi:10.1093/intimm/dxm083
83. Di Castro MA, Trettel F, Milior G, Maggi L, Ragozzino D, Limatola C. The chemokine CXCL16 modulates neurotransmitter release in hippocampal CA1 area. Sci Rep. 2016;6:34633. doi:10.1038/srep34633
84. Fukumoto N, Shimaoka T, Fujimura H, et al. Critical roles of CXC chemokine ligand 16/scavenger receptor that binds phosphatidylserine and oxidized lipoprotein in the pathogenesis of both acute and adoptive transfer experimental autoimmune encephalomyelitis. J Immunol. 2004;173(3):1620–1627. doi:10.4049/jimmunol.173.3.1620
85. Kuipers HF, Rieck M, Gurevich I, et al. Hyaluronan synthesis is necessary for autoreactive T-cell trafficking, activation, and Th1 polarization. Proc Natl Acad Sci U S A. 2016;113(5):1339–1344. doi:10.1073/pnas.1525086113
86. Tavazzi E, Rovaris M, La Mantia L. Drug therapy for multiple sclerosis. CMAJ. 2014;186(11):833–840. doi:10.1503/cmaj.130727
87. Hermann R, Karlsson MO, Novakovic AM, Terranova N, Fluck M, Munafo A. The clinical pharmacology of cladribine tablets for the treatment of relapsing multiple sclerosis. Clin Pharmacokinet. 2019;58(3):283–297. doi:10.1007/s40262-018-0695-9
88. Al-Ani MR, Raju TK, Hachim MY, et al. Rituximab prevents the development of experimental autoimmune encephalomyelitis (EAE): comparison with prophylactic, therapeutic or combinational regimens. J Inflamm Res. 2020;13:151–164. doi:10.2147/JIR.S243514
89. Martínez-Cáceres EM, Barrau MA, Brieva L, Espejo C, Barberà N, Montalban X. Treatment with methylprednisolone in relapses of multiple sclerosis patients: immunological evidence of immediate and short-term but not long-lasting effects. Clin Exp Immunol. 2002;127(1):165–171. doi:10.1046/j.1365-2249.2002.01725.x
90. Wandinger KP, Wessel K, Trillenberg P, Heindl N, Kirchner H. Effect of high-dose methylprednisolone administration on immune functions in multiple sclerosis patients. Acta Neurol Scand. 1998;97(6):359–365. doi:10.1111/j.1600-0404.1998.tb05966.x
91. Jalosinski M, Karolczak K, Mazurek A, Glabinski A. The effects of methylprednisolone and mitoxantrone on CCL5-induced migration of lymphocytes in multiple sclerosis. Acta Neurol Scand. 2008;118(2):120–125. doi:10.1111/j.1600-0404.2008.00998.x
92. Michałowska-Wender G, Losy J, Szczuciński A, Biernacka-Łukanty J, Wender M. Effect of methylprednisolone treatment on expression of sPECAM-1 and CXCL10 chemokine in serum of MS patients. Pharmacol Rep. 2006;58(6):920–923.
93. Fischer HJ, Finck TLK, Pellkofer HL, Reichardt HM, Lühder F. Glucocorticoid therapy of multiple sclerosis patients induces anti-inflammatory polarization and increased chemotaxis of monocytes. Front Immunol. 2019;10:1200. doi:10.3389/fimmu.2019.01200
94. Sellebjerg F, Börnsen L, Khademi M, et al. Increased cerebrospinal fluid concentrations of the chemokine CXCL13 in active MS. Neurology. 2009;73(23):2003–2010. doi:10.1212/WNL.0b013e3181c5b457
95. Bielecki B, Mazurek A, Wolinski P, Glabinski A. Treatment of multiple sclerosis with methylprednisolone and mitoxantrone modulates the expression of CXC chemokine receptors in PBMC. J Clin Immunol. 2008;28(2):122–130. doi:10.1007/s10875-007-9142-7
96. Kala M, Miravalle A, Vollmer T. Recent insights into the mechanism of action of glatiramer acetate. J Neuroimmunol. 2011;235(1–2):9–17. doi:10.1016/j.jneuroim.2011.01.009
97. Sand KL, Knudsen E, Rolin J, Al-Falahi Y, Maghazachi AA. Modulation of natural killer cell cytotoxicity and cytokine release by the drug glatiramer acetate. Cell Mol Life Sci. 2009;66(8):1446–1456. doi:10.1007/s00018-009-8726-1
98. Høglund RA, Holmøy T, Harbo HF, Maghazachi AA. A one year follow-up study of natural killer and dendritic cells activities in multiple sclerosis patients receiving glatiramer acetate (GA). PLoS One. 2013;8(4):e62237. doi:10.1371/journal.pone.0062237
99. Al-Falahi Y, Sand KL, Knudsen E, Damaj BB, Rolin J, Maghazachi AA. Splenic natural killer cell activity in two models of experimental neurodegenerative diseases. J Cell Mol Med. 2009;13(8B):2693–2703. doi:10.1111/j.1582-4934.2008.00640.x
100. Madsen C. The innovative development in interferon beta treatments of relapsing-remitting multiple sclerosis. Brain Behav. 2017;7(6):e00696. doi:10.1002/brb3.696
101. McCormack PL, Scott LJ. Interferon-beta-1b: a review of its use in relapsing-remitting and secondary progressive multiple sclerosis. CNS Drugs. 2004;18(8):521–546. doi:10.2165/00023210-200418080-00004
102. Zang YC, Halder JB, Samanta AK, Hong J, Rivera VM, Zhang JZ. Regulation of chemokine receptor CCR5 and production of RANTES and MIP-1alpha by interferon-beta. J Neuroimmunol. 2001;112(1–2):174–180. doi:10.1016/s0165-5728(00)00397-0
103. Sørensen TL, Sellebjerg F. Selective suppression of chemokine receptor CXCR3 expression by interferon-beta1a in multiple sclerosis. Mult Scler. 2002;8(2):104–107. doi:10.1191/1352458502ms781oa
104. Buttmann M, Merzyn C, Rieckmann P. Interferon-beta induces transient systemic IP-10/CXCL10 chemokine release in patients with multiple sclerosis. J Neuroimmunol. 2004;156(1–2):195–203. doi:10.1016/j.jneuroim.2004.07.016
105. Comabella M, Imitola J, Weiner HL, Khoury SJ. Interferon-beta treatment alters peripheral blood monocytes chemokine production in MS patients. J Neuroimmunol. 2002;126(1–2):205–212. doi:10.1016/s0165-5728(02)00064-4
106. Mueller AM, Yoon BH, Sadiq SA. Inhibition of hyaluronan synthesis protects against central nervous system (CNS) autoimmunity and increases CXCL12 expression in the inflamed CNS. J Biol Chem. 2014;289(33):22888–22899. doi:10.1074/jbc.M114.559583
107. Rilla K, Pasonen-Seppänen S, Rieppo J, Tammi M, Tammi R. The hyaluronan synthesis inhibitor 4-methylumbelliferone prevents keratinocyte activation and epidermal hyperproliferation induced by epidermal growth factor. J Invest Dermatol. 2004;123(4):708–714. doi:10.1111/j.0022-202X.2004.23409.x
108. García-Vilas JA, Quesada AR, Medina MÁ. 4-methylumbelliferone inhibits angiogenesis in vitro and in vivo. J Agric Food Chem. 2013;61(17):4063–4071. doi:10.1021/jf303062h
109. Abu Saleh M, Solayman M, Hoque MM, Khan MA, Sarwar MG, Halim MA. Inhibition of DNA topoisomerase type IIα (TOP2A) by mitoxantrone and its halogenated derivatives: a combined density functional and molecular docking study. Biomed Res Int. 2016;2016:6817502. doi:10.1155/2016/6817502
110. Fox EJ. Mechanism of action of mitoxantrone. Neurology. 2004;63(12 Suppl 6):S15–S18. doi:10.1212/wnl.63.12_suppl_6.s15
111. Scott LJ, Figgitt DP. Mitoxantrone: a review of its use in multiple sclerosis. CNS Drugs. 2004;18(6):379–396. doi:10.2165/00023210-200418060-00010
112. Burns SA, Lee Archer R, Chavis JA, Tull CA, Hensley LL, Drew PD. Mitoxantrone repression of astrocyte activation: relevance to multiple sclerosis. Brain Res. 2012;1473:236–241. doi:10.1016/j.brainres.2012.07.054
113. Pelfrey CM, Cotleur AC, Zamor N, Lee JC, Fox RJ. Immunological studies of mitoxantrone in primary progressive MS. J Neuroimmunol. 2006;175(1–2):192–199. doi:10.1016/j.jneuroim.2006.03.003
114. Deeks ED. Cladribine tablets: a review in relapsing MS. CNS Drugs. 2018;32(8):785–796. doi:10.1007/s40263-018-0562-0
115. Bartosik-Psujek H, Belniak E, Mitosek-Szewczyk K, Dobosz B, Stelmasiak Z. Interleukin-8 and RANTES levels in patients with relapsing-remitting multiple sclerosis (RR-MS) treated with cladribine. Acta Neurol Scand. 2004;109(6):390–392. doi:10.1111/j.1600-0404.2004.00259.x
116. Kawamoto E, Nakahashi S, Okamoto T, Imai H, Shimaoka M. Anti-integrin therapy for multiple sclerosis. Autoimmune Dis. 2012;2012:357101. doi:10.1155/2012/357101
117. Hutchinson M. Natalizumab: A new treatment for relapsing remitting multiple sclerosis. Ther Clin Risk Manag. 2007;3(2):259–268. doi:10.2147/tcrm.2007.3.2.259
118. Polman CH, O’Connor PW, Havrdova E, et al. A randomized, placebo-controlled trial of natalizumab for relapsing multiple sclerosis. N Engl J Med. 2006;354(9):899–910. doi:10.1056/NEJMoa044397
119. Tubridy N, Behan PO, Capildeo R, et al. The effect of anti-alpha4 integrin antibody on brain lesion activity in MS. The UK Antegren Study Group. Neurology. 1999;53(3):466–472. doi:10.1212/wnl.53.3.466
120. Dalton CM, Miszkiel KA, Barker GJ, et al. Effect of natalizumab on conversion of gadolinium enhancing lesions to T1 hypointense lesions in relapsing multiple sclerosis. J Neurol. 2004;251(4):407–413. doi:10.1007/s00415-004-0332-4
121. Mellergård J, Edström M, Vrethem M, Ernerudh J, Dahle C. Natalizumab treatment in multiple sclerosis: marked decline of chemokines and cytokines in cerebrospinal fluid. Mult Scler. 2010;16(2):208–217. doi:10.1177/1352458509355068
122. Cooper N, Arnold DM. The effect of rituximab on humoral and cell mediated immunity and infection in the treatment of autoimmune diseases. Br J Haematol. 2010;149(1):3–13. doi:10.1111/j.1365-2141.2010.08076.x
123. Casan JML, Wong J, Northcott MJ, Opat S. Anti-CD20 monoclonal antibodies: reviewing a revolution. Hum Vaccin Immunother. 2018;14(12):2820–2841. doi:10.1080/21645515.2018.1508624
124. Pescovitz MD. Rituximab, an anti-cd20 monoclonal antibody: history and mechanism of action. Am J Transplant. 2006;6(5 Pt 1):859–866. doi:10.1111/j.1600-6143.2006.01288.x
125. Palanichamy A, Jahn S, Nickles D, et al. Rituximab efficiently depletes increased CD20-expressing T cells in multiple sclerosis patients. J Immunol. 2014;193(2):580–586. doi:10.4049/jimmunol.1400118
126. Ineichen BV, Moridi T, Granberg T, Piehl F. Rituximab treatment for multiple sclerosis. Mult Scler. 2020;26(2):137–152. doi:10.1177/1352458519858604
127. Salzer J, Svenningsson R, Alping P, et al. Rituximab in multiple sclerosis: A retrospective observational study on safety and efficacy. Neurology. 2016;87(20):2074–2081. doi:10.1212/WNL.0000000000003331
128. Piccio L, Naismith RT, Trinkaus K, et al. Changes in B- and T-lymphocyte and chemokine levels with rituximab treatment in multiple sclerosis. Arch Neurol. 2010;67(6):707–714. doi:10.1001/archneurol.2010.99
129. Cross AH, Klein RS, Piccio L. Rituximab combination therapy in relapsing multiple sclerosis. Ther Adv Neurol Disord. 2012;5(6):311–319. doi:10.1177/1756285612461165
130. de Flon P, Söderström L, Laurell K, et al. Immunological profile in cerebrospinal fluid of patients with multiple sclerosis after treatment switch to rituximab and compared with healthy controls. PLoS One. 2018;13(2):e0192516. doi:10.1371/journal.pone.0192516
131. Stone MJ, Hayward JA, Huang C, Huma Z, Sanchez J. Mechanisms of regulation of the chemokine-receptor network. Int J Mol Sci. 2017;18(2):342. doi:10.3390/ijms18020342
132. O’Hayre M, Salanga CL, Handel TM, Hamel DJ. Emerging concepts and approaches for chemokine-receptor drug discovery. Expert Opin Drug Discov. 2010;5(11):1109–1122. doi:10.1517/17460441.2010.525633
133. Opdenakker G, Van Damme J. Probing cytokines, chemokines and matrix metalloproteinases towards better immunotherapies of multiple sclerosis. Cytokine Growth Factor Rev. 2011;22(5–6):359–365. doi:10.1016/j.cytogfr.2011.11.005
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.