Back to Journals » Journal of Inflammation Research » Volume 17
TAK1 in Vascular Signaling: “Friend or Foe”?
Authors Fan G, Lu J, Zha J ![]() , Guo W, Zhang Y, Liu Y, Zhang L
, Guo W, Zhang Y, Liu Y, Zhang L
Received 10 January 2024
Accepted for publication 16 April 2024
Published 16 May 2024 Volume 2024:17 Pages 3031—3041
DOI https://doi.org/10.2147/JIR.S458948
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Tara Strutt
Gang Fan,1,* Jingfen Lu,2,* Jinhui Zha,3,* Weiming Guo,1 Yifei Zhang,4 Yuxin Liu,5 Liyuan Zhang1
1Department of Urology, Huazhong University of Science and Technology Union Shenzhen Hospital, Shenzhen, 518052, People’s Republic of China; 2The First Clinical Medical College of Guangzhou University of Chinese Medicine, Guangzhou, 510006, People’s Republic of China; 3Shenzhen University, Shenzhen, 518000, People’s Republic of China; 4The First Teaching Hospital of Tianjin University of Traditional Chinese Medicine, Tianjin, 300193, People’s Republic of China; 5College of Traditional Chinese Medicine, Hunan University of Chinese Medicine, Changsha, 410208, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Liyuan Zhang; Gang Fan, Department of Urology, Huazhong University of Science and Technology Union Shenzhen Hospital, Shenzhen, 518000, People’s Republic of China, Email [email protected]; [email protected]
Abstract: The maintenance of normal vascular function and homeostasis is largely dependent on the signaling mechanisms that occur within and between cells of the vasculature. TGF-β-activated kinase 1 (TAK1), a multifaceted signaling molecule, has been shown to play critical roles in various tissue types. Although the precise function of TAK1 in the vasculature remains largely unknown, emerging evidence suggests its potential involvement in both physiological and pathological processes. A comprehensive search strategy was employed to identify relevant studies, PubMed, Web of Science, and other relevant databases were systematically searched using keywords related to TAK1, TABs and MAP3K7.In this review, we discussed the role of TAK1 in vascular signaling, with a focus on its function, activation, and related signaling pathways. Specifically, we highlight the TA1-TABs complex is a key factor, regulating vascular smooth muscle cells (VSMCs) and endothelial cells (ECs) involved in the processes of inflammation, vascular proliferation and angiogenesis. This mini review aims to elucidate the evidence supporting TAK1 signaling in the vasculature, in order to better comprehend its beneficial and potential harmful effects upon TAK1 activation in vascular tissue.
Keywords: TGF-β-activated kinase 1, vascular disease, mitogen-activated protein kinase, transforming growth factor-β, smooth muscle cell, endothelial cell
Introduction
TGF-β-activated kinase 1 (TAK1) is a protein kinase that exerts fundamental control over numerous cellular processes including inflammation, cell differentiation, and survival.1,2 Recently, emerging evidence has affirmed the critical role of TAK1 in cardiovascular disease, a leading cause of worldwide mortality, comprising an assemblage of heart and blood vessel-related ailments encompassing coronary artery disease, stroke, and heart failure.3,4 The pathophysiological mechanisms underpinning cardiovascular disease are multifaceted and involve a gamut of genetic and environmental determinants interconnecting with cellular signaling pathways. In the cardiovascular apparatus, TAK1 has demonstrated significant participation in the modulation of vascular function of vascular homeostasis, constriction, angiogenesis and so on, as well as targeted participation in regulating both cell death and inflammation.5
TAK1 is a crucial signaling molecule that activates nuclear factor-kappa B (NF-κB) and mitogen-activated protein kinase (MAPK) pathways in response to various signals, such as cytokines and chemokines, and regulates inflammasomes.6 Through TAK1, these diverse conditions and signals influence cell functions, including cell cycle, cell differentiation, immune responses, cell migration, and redox homeostasis. The dysregulation of these pathways is frequently linked to pathological vascular disease.7,8 This review assembles evidence indicating a comparable dual role of TAK1 in cells of the vasculature and identifies potential avenues of inquiry. The objective is to advance our understanding of TAK1 signaling in cardiovascular health and disease.
TAK1 Function and Activation
TAK1 Related Signaling
Transforming growth factor-β (TGF-β) is a multifaceted cytokine with the ability to regulate diverse intracellular signaling pathways. Conventionally, it is known to promote the Smad-dependent signaling pathway.9 However, apart from the canonical Smad-dependent pathways, TGF-β1 can directly activate non-canonical, Smad-independent pathways to regulate an expansive range of downstream cellular responses.10 TGF-β1 can activate TAK1, a serine/threonine kinase and a member of the mitogen-activated protein kinase kinase kinase (MAP3K) family, also named as MAP3K7. In addition to TGF-β1, TAK1 can be activated by multiple stimuli, such as IL-1α, IL-1β, TNF- α, TLR ligands, Wnt, and is responsible for phosphorylating a series of target proteins.11 Consequently, it elicits various signal transduction and cellular responses that differ across stresses or cell types.
It has been well established that the enzyme TAK1 plays an essential role in the activation of key signaling pathways, including NF-kB and MAPKs, by TNF receptor (TNFR), Interleukin-1 receptor (IL-1R), and Toll-like receptors (TLRs).12 TAK1-TABs complexes are activated and assembled by stimulation of TNFR1, IL-1R, and TLR with their ligands. TAK1-TABs then phosphorylated IKKs, thereby activating the transcription factor NF-kB and causing transcription of downstream genes. Unique among members of the MAP3K family, activated TAK1 also activates MAPKKs by phosphorylates.13 TAK1 has been directly implicated in TGF-β1 signaling, whereby its activation occurs in response to TGF-β signaling. Upon activation, TAK1 in turn activates several critical intracellular kinases, including C-Jun N-terminal kinase (JNK), p38 MAPK, and Inhibitor of kappa B kinase complex (IKKβ).14 TAK1 is considered a key mediator of activation in each of these pathways and plays an important role in transmitting the upstream signal from the receptor complex to downstream signalosomes.
TAK1-TABs Complex
The activation of TAK1 is contingent on the presence of TAK1-binding protein 1 (TAB1), TAB2, and TAB3.15,16 TAB1 serves as an adaptor protein that is continuously bound to the N-terminal kinase domain of TAK1, even in the absence of stimulation. Conversely, TAB2 and TAB3 are only able to bind to the C terminus of TAK1 via the TAK1-binding domain after stimulation.16 Through different signaling pathways, both TAB1 and TAB2 activate the TAK1 protein. TAB1 is essential for osmotic stress-induced TAK1 activation, whereas TAB2 or TAB3 is required for TNF-α or IL-1-induced TAK1 activation.17 TAB1 constitutively binds to TAK1, with no phosphatase or other enzymatic activity. However, O-GlcNAcylation of TAB1 substantially increases the autophosphorylation of TAK1, phosphorylation of IKKβ and translocation of NF-κB, which results in increased production of cytokines.18 Empirical evidence suggests that excessive production of TAB1 augments the kinase activity of TAK1 and functions as an activator of the NF-kB signaling pathway.19 Conversely, the deficiency of TAB1 drastically impedes TAK1 phosphorylation at residue threonine 187 (Thr187) following TNFa and IL-1b stimulations.20
In contrast to TAB1, TAB2 and TAB3 do not activate TAK1 in vitro.21 Notably, the double deficiency of both TAB2 and TAB3 only marginally affects TAK1 activation and the subsequent production of downstream inflammatory cytokines in the early phase after IL-1b stimulation.22 Therefore, TAB2 and TAB3 are deemed dispensable for early TAK1 activation. However, TAB2 deficiency in mouse embryos led to the abnormal growth of capillary blood vessels, proving that TAB2 is crucial for maintaining normal vascular homeostasis.23 Numerous reports have demonstrated that the TAK1-TABs complex has far-reaching implications in regulating a diverse range of physiological and pathological processes.24 Importantly, Interactions of IL-1R, TLR and TNF receptor (TNFR) trigger the strong interaction of TAB2/3 with K63-linked polyubiquitin chains to activate TAK1, then TAK1-TABs complex phosphorylates IKKβ at Ser177 and Ser181, effectively engendering the activation of NF-kB signaling, ultimately regulating inflammation, proliferation and angiogenesis processes.25 The. Furthermore, the TAK1-TABs complex plays a central role in the activation of MAPKs26 (Figure 1).
 |
Figure 1 Effects of TAK1 activation in vascular. TGF-β1 activates TAK1, resulting in the assembly and activation of the TAK1-TABs complex. Subsequently, it activates NF-κB and MAPK signal transduction, leading to cellular responses that vary across organs or cell types, potentially causing both beneficial and harmful effects in vascular tissue. |
TAK1 and Vascular Physiology and Pathologies
Vascular Smooth Muscle Cells (VSMCs)
TAK1, a protein kinase, assumes a significant function in the mediation of cellular activities, including autophagy.27 A recent association has been discovered between autophagy regulation of vascular smooth muscle cell (VSMC) functions and vasculopathy development. Upon the occurrence of aortic transplantation, the activation of TAK1 within VSMCs represents an early and pivotal action for VSMC migration, proliferation, and neointima composition. This event facilitates regulation of autophagy activation and thus constitutes an attractive molecular mechanism and prospective therapeutic target for restraining transplant vasculopathy.28 Table 1 summarized relevant studies of TAK1 in different blood vessel cells.
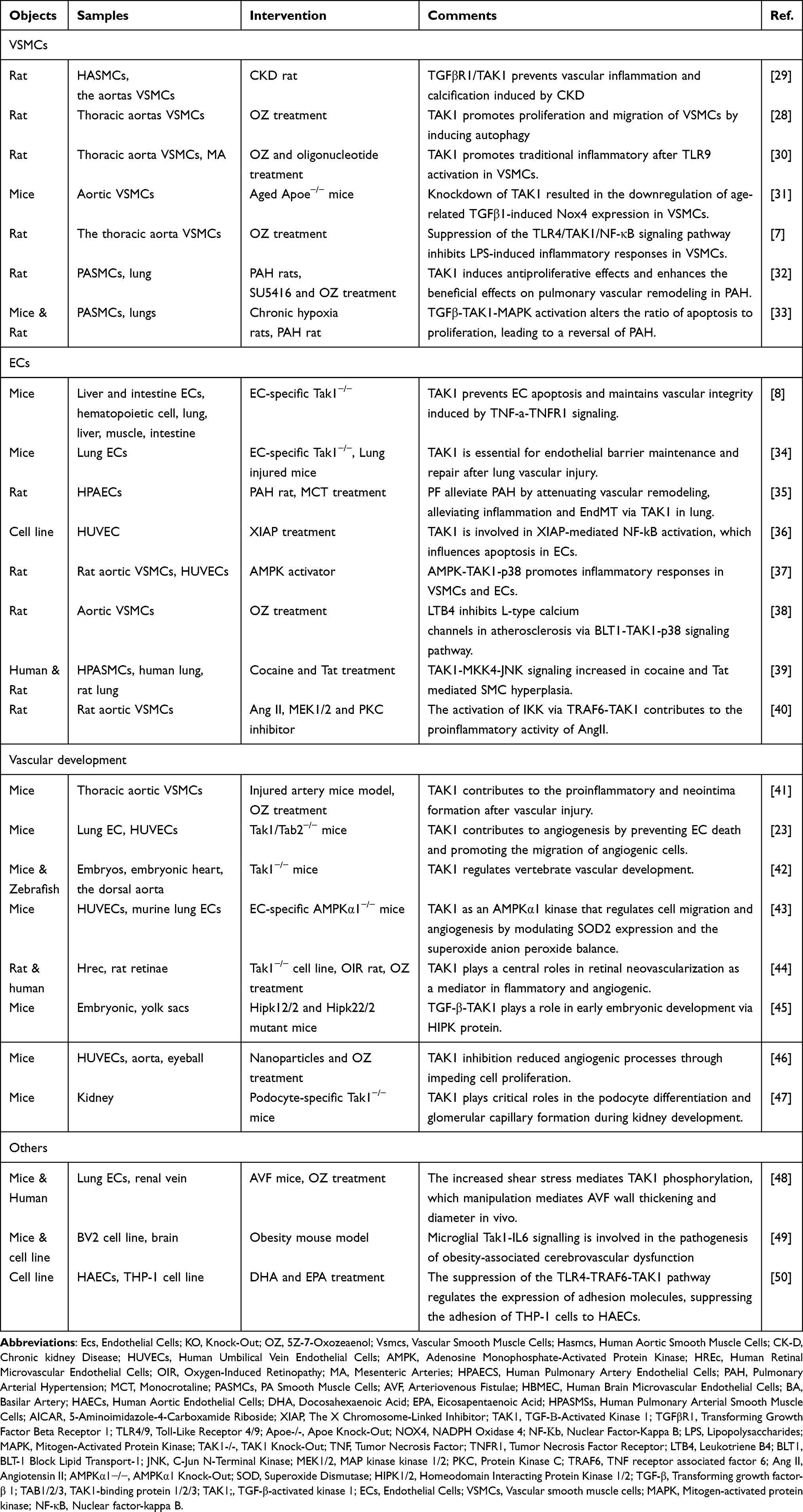 |
Table 1 TAK1 in Vascular |
Aging is one of the contributing factors to vascular dysfunction, mediated through mechanisms including oxidative stress, chronic inflammation, apoptosis, and autophagy of VSMCs, resulting in decreased vascular compliance and contractility, vascular sclerosis. These alterations may lead to hypertension, arteriosclerosis, and cardiovascular diseases. The expression and activity of NADPH oxidase 4 (NOX4) during the aging process lead to increased cellular and mitochondrial oxidative stress, vascular inflammation, dysfunction, and atherosclerosis.51 The presence of TGFβ1 substantially upregulates the expression of Nox4 in VSMCs as a consequence of aging. Conversely, the application of melittin-derived peptide p5RHH for the knockdown of TAK1 resulted in the decreased expression of Nox4 in VSMCs, which is induced by age-related TGFβ1.31 Upon sustaining vascular injury, TAK1 phosphorylation (Thr187) is highly induced within the walls of the injured vessels, particularly in the medial VSMCs.41 The upregulation of TAK1 phosphorylation in VSMCs is proposed to play a significant role in the modulation of pro-inflammatory and pro-oxidative effects on these cells, thereby contributing to neointima formation. Notably, siRNA knockdown or 5Z-7-oxozeaenol inhibition of TAK1 resulted in a pronounced reduction of NADPH oxidase activation and subsequent superoxide production triggered by CD40L/CD40 stimulation41 (Table 1).
The involvement of locally produced angiotensin II (Ang II) in chronic inflammatory reactions that contribute to the development of atherosclerosis is postulated to result from the activation of NF-κB transcription factors. This activation is mediated through the phosphorylation of the catalytic subunit IKKβ on serine residues 177 and 181 located in the activation T-loop. The rapid phosphorylation of IKKβ is contingent upon the activation of a second messenger-dependent pathway consisting of PKCα-TRAF6-TAK1, which is crucial for driving the proinflammatory effects of Ang II.40 The stromal-derived factor-1 (SDF-1), derived from VSMCs, plays a critical role in vascular repair and remodeling in diverse vascular ailments. In VSMCs, the IL-1α-induced SDF-1 expression is regulated by TAK1, a pivotal modulator of inflammatory cascades. Furthermore, the suppression of TAK1, an upstream stimulator of IKKβ signaling, significantly amplifies the IL-1α-induced C/EBPβ expression and promotes SDF-1 expression.52
Pulmonary arterial hypertension (PAH) is a cardiovascular disorder characterized by enhanced proliferation and suppressed apoptosis of pulmonary arterial smooth muscle cells (PASMCs).53 The signaling pathway involving TGF-β is widely believed to play a crucial role in pulmonary vascular remodeling. The inhibition of TAK1 results in rescuing abnormal proliferation and apoptosis in PASMCs33 (Table 1). This approach is supported by evidence showing that the use of 5Z-7-oxozeaenol, an inhibitor of TAK1, induces antiproliferative effects. Furthermore, using 5Z-7-oxozeaenol as an adjunct therapy to short-term vasodilator therapy enhances the effects on pulmonary vascular remodeling and right ventricle metabolic reprogramming in experimental PAH.32 In addition, Toll-like receptor 9 (TLR9) signaling has been linked to proinflammatory gene transcription and increased contraction in isolated arteries. TLR9 activation in VSMCs induces AMPK phosphorylation via TAK1, indicating that TLR9-mediated signaling is an extension of the traditional inflammatory pathway regulated by TAK1.30 Paeoniflorin (PF), which possesses several beneficial functions, including vasodilation and anti-inflammation, has been shown to prevent human PASMCs from PDGF-BB-stimulated proliferation and migration. Furthermore, PF inhibits monocrotaline-induced down-regulation of bone morphogenetic protein receptor 2 (BMPR2) and suppresses monocrotaline-induced phosphorylation of TAK1, attenuating monocrotaline-induced PAH in rats.35
Chronic inflammation is an imperative factor in vascular calcification. Calcification in rats with chronic kidney disease (CKD) or in human aortic smooth muscle cells (HASMCs) cultured in an osteogenic medium is induced by inflammation stimulated via the activation of the TGFBR1/TAK1 pathway. Inhibition of the TGFBR1/TAK1 pathway, facilitated by the farnesoid X receptor (FXR), ultimately culminates in the alleviation of both vascular inflammation and calcification in CKD rats29 (Figure 1).
Endothelial Cells (ECs)
TAK1 activation has been shown to prevent endothelial apoptosis and sustain vascular integrity in the presence of inflammatory stimuli. Upon stimulation with TNF-α, TAK1 promotes the survival of endothelial cells, while endothelial-specific TAK1 knockout mice exhibit a hemorrhagic phenotype in the intestine and liver due to EC apoptosis, resulting in vessel collapse and rapid death.8 The activation of the anti-apoptotic pathway triggers the activation of the transcription factor NF-κB, which governs the expression of certain genes like A20 or members of the IAP gene family.36 These genes function to shield cells from TGFβ-induced apoptosis. Among the IAP gene family, XIAP is an NF-κB-dependent member that enhances the translocation of the p65 subunit of NF-κB to the nucleus. As p65 is one of activators of NF-kB superfamily, and nuclear translocation is a prerequisite for NF-kB activity.54 This process is mediated through its interaction with TAK1 in endothelial cells, influencing both apoptotic and anti-apoptotic signaling pathways36 (Table 1 and Figure 1).
The indispensable role of TAK1 and NEMO in preserving the neurovascular unit is highlighted by the detrimental effects of selectively deleting these proteins in brain endothelial cells.55 Notably, a loss of TAK1 expression in human endothelial cells impairs proliferation, migration, and tube formation without provoking apoptosis.43 TAK1-mediated angiogenesis is also regulated by AMPKα1 and redox balance via SOD2 expression and modulation of the superoxide anion and hydrogen peroxide ratio.43 The critical involvement of endothelial TAK1 and TAB2, but not TAB1, in vascular formation is demonstrated by the observation that TAK1 deficiency in endothelial cells leads to increased cell death and vessel regression at E10.5.23
The maintenance and repair of the lung endothelial barrier rely on the proper functioning of the endothelial TAK1 kinase. Inhibition of TAK1 using 5Z-7-oxozeaenol results in a significant reduction in β-catenin and VE-cadherin expression at the endothelial adherens junctions and an increase in lung vascular permeability triggered by protease-activated receptor-1 (PAR-1) or toll-like receptor-4 (TLR-4). The lack of TAK1 activity leads to a compromised lung endothelial barrier, rendering mice more susceptible to septic shock, as observed in endothelial-specific TAK1 knockout mice.34
Necroptosis is a regulated cell death process that can be triggered in conditions where apoptosis is disrupted.56 When cerebral perfusion is reduced due to arterial occlusion, TAK1, which is a suppressor of RIPK1, is degraded, leading to a shift from necroptosis to apoptosis. Microglial/infiltrated macrophages and neuronal lineages with a conditional knockout of TAK1 become more sensitive to ischemic infarction due to the promotion of apoptosis. This study demonstrates that the sequential activation of necroptosis and apoptosis is involved in the vascular and neural pathology associated with stroke.57
The activation and impairment of vascular endothelial cells is a central causative factor leading to renal dysfunction.58,59 The microRNA-449c-5p has the ability to curb the secretion of cytokines that are triggered by lipopolysaccharides, thereby impeding apoptosis induction and paving the way for cell proliferation via its ability to repress the expression of TAK1.60
Angiogenesis
TAK1 plays an essential role in angiogenesis and is implicated in angiogenesis-related pathological mechanisms. In-depth exploration of TAK1’s modulatory effects on aberrant angiogenesis has led to the identification of therapeutic strategies targeting this protein. The initiation of angiogenesis is driven by endothelial cell proliferation, followed by tube formation, a process enriched by smooth muscle cells, that eventually results in the formation of a specific vascular network.8,23 Activation of the NF-κB and MAPK signaling pathways, under the regulation of TAK1, promotes the expression of various inflammatory response proteins, including cytokines and chemokines, implicated in the regulation of inflammasomes, all of which positively impact the angiogenic process55 (Figure 1).
TAK1 deficiency results in embryonic lethality owing to vascular destruction, underscoring its pivotal function in sustaining vascular integrity throughout embryogenesis.23 The vasculature in Tak1 mutant embryos displays a range of anomalies, including vessel dilation and misbranching, coupled with the absence of vascular smooth muscle in both the yolk sac and the central vasculature of the embryo proper.42 TGF-β has been identified as a potent regulator of multiple aspects of vascular development, including angiogenesis during embryonic development. HIPK1 and HIPK2 (Homeodomain Interacting Protein Kinase 1/2, HIPK1/2) transcriptional corepressors have also been shown to regulate TGF-β-dependent angiogenesis during embryonic development. TGF-β-TAK1 signaling phosphorylates tyrosine residue Y-361 located in HIPK2’s kinase domain to activate it45 (Table 1).
Cancer is commonly characterized by the presence of hypoxia and inflammation. The transcription factor families of Hypoxia Inducible Factor (HIF) and Nuclear Factor of κ-light-chain-enhancer of activated B cells (NF-κB) are of utmost importance in regulating cellular responses under stress stimuli.61,62 Activation of the inflammatory pathway can induce the expression of HIF-1α, resulting in a positive feedback loop.63 TAK1 is a pivotal regulator of the crosstalk between inflammation and hypoxia, further enhancing tumor cell proliferation and angiogenesis. Endothelial cell death induced by TNF-α can be prevented by TAK1, which also plays a significant role in tumor vasculature maintenance.8 Inhibiting or knocking out TAK1 can lead to the apoptosis of endothelial cells and destruction of tumor vasculature, ultimately resulting in tumor regression. Additionally, TGF-β signaling in tumor cells has been shown to play a role in tumor angiogenesis and metastasis by regulating matrix proteolysis. The studies using orthotopic xenograft and SCID mice have demonstrated the efficacy of dn-TAK1 in reducing tumor growth and formation of lung metastases. The mechanism by which TAK1 contributes to TGF-β1-mediated tumor angiogenesis and metastasis involves the TAK1-NF-kappaB-MMP-9 pathway64.
Corneal neovascularization (CoNV), a debilitating complication associated with various corneal diseases, is a leading cause of irreversible visual impairment.65 Studies have identified TAK1 as playing a crucial role in CoNV pathogenesis, and inhibition of TAK1 presents a potential therapeutic approach for retinal neovascular pathologies. TAK1 is essential for activation of NFκB signaling and downstream gene expression related to angiogenesis and endothelial activation, as demonstrated in TAK1 knockout human endothelial cells.44 Inhibition of TAK1 by 5Z-7-oxozeaenol, a potent small molecule inhibitor, was found to reduce angiogenesis by suppressing DNA replication and cell cycle, thereby inhibiting cell proliferation. Importantly, pharmacological inhibition of TAK1 by 5Z-7-oxozeaenol significantly attenuated aberrant retinal angiogenesis in OIR rats.44 Furthermore, 5Z-7-oxozeaenol effectively blocked TNFα-mediated NFκB signaling and downstream gene expression related to angiogenesis and inflammation by inhibiting TAK1 activity46 (Table 1).
The TGF-β activated kinase 1 (TAK1) signaling pathway is a fundamental player in the processes governing podocyte differentiation, glomerular microvascular development, and the maintenance of the glomerular filtration barrier homeostasis.47,66 Notably, the targeted deletion of Tak1 in podocytes resulted in marked disruptions to the structural integrity of the podocyte architecture, primarily characterized by a compromise in podocyte foot process formation leading to effacement. Furthermore, the Tak1 (∆/∆) mice exhibited atypical enlargement of glomerular capillaries with concomitant upregulation of vascular endothelial growth factor expression within the glomerulus. These findings highlight the central importance of the TAK1 pathway in podocyte function and renal microvascular development.47
Others
Arteriovenous fistula (AVF) serve as the most effective conduit for hemodialysis access.67 The process of AVF maturation is reliant on the augmentation of the wall’s thickness through the deposition of extracellular matrix (ECM) components such as collagen and fibronectin, alongside lumen dilation. The in vivo modification of TAK1 instigated variations in AVF wall thickening and luminal diameter. A decline in TAK1 function was concomitant with reduced thickness and smaller diameter, whereas activation of TAK1 function led to amplified thickness and broader diameter48 (Table 1). Protracted obesity is linked to cerebrovascular impairment, as evidenced by abnormalities in the basilar artery (BA) of a mouse model demonstrating prolonged obesity. Over-activation of microglial Tak1 in the brainstem was observed in this model. This activation of Tak1 resulted in an escalation of interleukin-18 (IL-18) production. As a proinflammatory cytokine, the action of IL-18 on various vascular cells could collectively contribute to the cerebrovascular dysfunction in animals with prolonged obesity. Since the chronic, low-grade inflammation is induced by IL-18, protection from cerebrovascular dysfunction was afforded by blocking the IL-18 receptor in the brain, despite continued obesity in the model49 (Figure 1).
Conclusions
Based on the aforementioned investigations, it has been established that TAK1 is a multifaceted signaling molecule that elicits the activation of NF-κB and MAPK pathways, exerting varying influences on the regulation of vascular smooth muscle cells (VSMCs), endothelial cells (ECs), angiogenesis, and the vascular wall. Activation of TAK1, on the one hand, is implicated in autophagy, pro-inflammatory and pro-oxidative responses, thereby potentially contributing to transplant vasculopathy and vascular senescence, vascular calcification, atherosclerosis, pulmonary hypertension, and tumor progression. Conversely, inhibiting or down-regulating TAK1 can mitigate and alleviate these factors with detrimental consequences for the organism.
Nevertheless, TAK1 activation serves as a protective measure against endothelial cell apoptosis and sustains vascular integrity, thereby playing a crucial role in angiogenesis and embryonic development. Additionally, TAK1 safeguards neurovascular units and participates in the restoration of pulmonary endothelial barrier function, as well as the prevention of renal vascular endothelial cell injury. Consequently, TAK1 acts as an indispensable and prominent intermediary within numerous physiological mechanisms that uphold normal bodily functions, hence earning the moniker of a “Friend”. Although uncovering these mechanisms offers novel molecular insights and targets for potential disease control, the multifaceted involvement of TAK1 necessitates careful regulation, without outright inhibition or excessive activation of its functions. Consequently, the targeted manipulation and maintenance of TAK1 levels within specific cells and tissues represents a crucial area of investigation that will enable effective utilization of its beneficial properties while minimizing potential harm. It’s also one of the directions that future research efforts need to focus on.
Author Contributions
Gang Fan, Jingfen Lu and Jinhui Zha share first authorship. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
GF is supported by the National Natural Science Foundation of China (82001489), Shenzhen Natural Science Foundation (JCYJ 20220530141613031), Shenzhen Nanshan District Science and Technology Plan Project (NS044), Huazhong University of Science and Technology Union Shenzhen Hospital Foundation. Weiming Guo is supported by the Health Bureau of Nanshan District, Shenzhen (NSZD2023010) and Shenzhen Natural Science Foundation (JCYJ20230807115817037).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Tzavlaki K, Moustakas A. TGF-β Signaling. Biomolecules. 2020;10(3):487. doi:10.3390/biom10030487
2. Wang J, Zhao X, Wan YY. Intricacies of TGF-β signaling in Treg and Th17 cell biology. Cell Mol Immunol. 2023;20(9):1002–1022. doi:10.1038/s41423-023-01036-7
3. Bai B, Ji Z, Wang F, et al. CTRP12 ameliorates post-myocardial infarction heart failure through down-regulation of cardiac apoptosis, oxidative stress and inflammation by influencing the TAK1-p38 MAPK/JNK pathway. Inflamm Res. 2023;72(7):1375–1390. doi:10.1007/s00011-023-01758-4
4. Zhe-Wei S, Li-Sha G, Yue-Chun L, Coşkun H, Koyuncu O, Walker S. The role of necroptosis in cardiovascular disease. Front Pharm. 2018;9:9. doi:10.3389/fphar.2018.00009
5. Roh YS, Song J, Seki E. TAK1 regulates hepatic cell survival and carcinogenesis. J Gastroenterol. 2014;49(2):185–194. doi:10.1007/s00535-013-0931-x
6. Zou W, Yin P, Shi Y, et al. A novel biological role of α-mangostin via TAK1–NF-κB pathway against inflammatory. Inflammation. 2019;42(1):103–112. doi:10.1007/s10753-018-0876-6
7. Meng Z, Si CY, Teng S, Yu XH, Li HY. Tanshinone IIA inhibits lipopolysaccharide‑induced inflammatory responses through the TLR4/TAK1/NF‑κB signaling pathway in vascular smooth muscle cells. IntJ Mol Med. 2019;43(4):1847–1858. doi:10.3892/ijmm.2019.4100
8. Naito H, Iba T, Wakabayashi T, et al. TAK1 prevents endothelial apoptosis and maintains vascular integrity. Dev Cell. 2019;48(2):151–166.e7. doi:10.1016/j.devcel.2018.12.002
9. Jiang T, Wang Z, Sun J. Human bone marrow mesenchymal stem cell-derived exosomes stimulate cutaneous wound healing mediates through TGF-β/Smad signaling pathway. Stem Cell Res Ther. 2020;11(1):1–10. doi:10.1186/s13287-020-01723-6
10. Azadian Z, Hosseini S, Dizjikan ZP, et al. Computational and in vitro validation of cardiogenic induction of quercetin on adipose-derived mesenchymal stromal cells through the inhibition of Wnt and non-Smad-dependent TGF-β pathways. J Cell Biochem. 2022;123(2):450–468. doi:10.1002/jcb.30189
11. Yang J, Li Y, Wang L, Zhang Z, Li Z, Jia HQ. LncRNA H19 aggravates TNF-α-induced inflammatory injury via TAK1 pathway in MH7A cells. BioFactors. 2020;46(5):813–820. doi:10.1002/biof.1659
12. Rodrigues M, Petrova T, Tibbs B, Arthur JSC, Cohen P. TAK1 protein kinase activity is required for TLR signalling and cytokine production in myeloid cells. Biochem J. 2022;479(17):1891–1907. doi:10.1042/BCJ20220314
13. Ninomiya-Tsuji J, Kishimoto K, Hiyama A, Inoue J, Cao Z, Matsumoto K. The kinase TAK1 can activate the NIK-I kappaB as well as the MAP kinase cascade in the IL-1 signalling pathway. Nature. 1999;398(6724):252–256. doi:10.1038/18465
14. Farhan M, McIntosh KA, Cunningham MR, Plevin R, Tinto K. TAK1 is an upstream MAP 3 K regulator of a novel non-canonical NFkB pathway stimulated by IL-1b: 19th world congress of basic & clinical pharmacology 2023. Br J Pharmacol. 2023;180(S1):708–709. doi:10.1111/bph.16110
15. Shibuya H, Yamaguchi K, Shirakabe K, et al. TAB1: an activator of the TAK1 MAPKKK in TGF-beta signal transduction. Science. 1996;272(5265):1179–1182. doi:10.1126/science.272.5265.1179
16. Besse A, Lamothe B, Campos AD, et al. TAK1-dependent signaling requires functional interaction with TAB2/TAB3. J Biol Chem. 2007;282(6):3918–3928. doi:10.1074/jbc.M608867200
17. Zhu L, Lama S, Tu L, Dusting GJ, Wang JH, Liu GS. TAK1 signaling is a potential therapeutic target for pathological angiogenesis. Angiogenesis. 2021;24(3):453–470. doi:10.1007/s10456-021-09787-5
18. Pathak S, Borodkin VS, Albarbarawi O, Campbell DG, Ibrahim A, van Aalten DM. O-GlcNAcylation of TAB1 modulates TAK1-mediated cytokine release. EMBO J. 2012;31(6):1394–1404. doi:10.1038/emboj.2012.8
19. Zhao J, Cai B, Shao Z, et al. TRIM26 positively regulates the inflammatory immune response through K11-linked ubiquitination of TAB1. Cell Death Differ. 2021;28(11):3077–3091. doi:10.1038/s41418-021-00803-1
20. Scholz R, Sidler CL, Thali RF, Winssinger N, Cheung PCF, Neumann D. Autoactivation of transforming growth factor β-activated Kinase 1 Is a sequential bimolecular process. J Biol Chem. 2010;285(33):25753–25766. doi:10.1074/jbc.M109.093468
21. Shim JH, Xiao C, Paschal AE, et al. TAK1, but not TAB1 or TAB2, plays an essential role in multiple signaling pathways in vivo. Genes Dev. 2005;19(22):2668–2681. doi:10.1101/gad.1360605
22. Zhang J, Macartney T, Peggie M, Cohen P. Interleukin-1 and TRAF6-dependent activation of TAK1 in the absence of TAB2 and TAB3. Biochem J. 2017;474(13):2235–2248. doi:10.1042/BCJ20170288
23. Morioka S, Inagaki M, Komatsu Y, Mishina Y, Matsumoto K, Ninomiya-Tsuji J. TAK1 kinase signaling regulates embryonic angiogenesis by modulating endothelial cell survival and migration. Blood. 2012;120(18):3846–3857. doi:10.1182/blood-2012-03-416198
24. Xu YR, Lei CQ. TAK1-TABs complex: A central Signalosome in inflammatory responses. Front Immunol. 2021;11:608976. doi:10.3389/fimmu.2020.608976
25. Braun H, Staal J. Stabilization of the TAK1 adaptor proteins TAB2 and TAB3 is critical for optimal NF-κB activation. FEBS J. 2020;287(15):3161–3164. doi:10.1111/febs.15210
26. Feng ZH, bo ZH, Wu X, ling GY, dong CW, Qian F. Fisetin alleviates sepsis-induced multiple organ dysfunction in mice via inhibiting p38 MAPK/MK2 signaling. Acta Pharmacol Sin. 2020;41(10):1348–1356. doi:10.1038/s41401-020-0462-y
27. Ran D, Ma Y, Liu W, et al. TGF-β-activated kinase 1 (TAK1) mediates cadmium-induced autophagy in osteoblasts via the AMPK / mTORC1 / ULK1 pathway. Toxicology. 2020;442:152538. doi:10.1016/j.tox.2020.152538
28. Zheng X, Yu Q, Shang D, et al. TAK1 accelerates transplant arteriosclerosis in rat aortic allografts by inducing autophagy in vascular smooth muscle cells. Atherosclerosis. 2022;343:10–19. doi:10.1016/j.atherosclerosis.2022.01.009
29. Li C, Zhang S, Chen X, et al. Farnesoid X receptor activation inhibits TGFBR1/TAK1-mediated vascular inflammation and calcification via miR-135a-5p. Commun Biol. 2020;3(1):327. doi:10.1038/s42003-020-1058-2
30. McCarthy CG, Wenceslau CF, Ogbi S, Szasz T, Webb RC. Toll-like receptor 9-Dependent AMPKα Activation Occurs via TAK1 and Contributes to RhoA/ROCK Signaling and Actin polymerization in vascular smooth muscle cells. J Pharmacol Exp Ther. 2018;365(1):60–71. doi:10.1124/jpet.117.245746
31. Lozhkin A, Vendrov AE, Pan H, Wickline SA, Madamanchi NR, Runge MS. NADPH oxidase 4 regulates vascular inflammation in aging and atherosclerosis. J Mol Cell Cardiol. 2017;102:10–21. doi:10.1016/j.yjmcc.2016.12.004
32. Morales-Cano D, Izquierdo-García JL, Barreira B, et al. Impact of a TAK-1 inhibitor as a single or as an add-on therapy to riociguat on the metabolic reprogramming and pulmonary hypertension in the SUGEN5416/hypoxia rat model. Front Pharm. 2023;14:1021535. doi:10.3389/fphar.2023.1021535
33. Nasim MT, Ogo T, Chowdhury HM, et al. BMPR-II deficiency elicits pro-proliferative and anti-apoptotic responses through the activation of TGFβ-TAK1-MAPK pathways in PAH. Hum Mol Genet. 2012;21(11):2548–2558. doi:10.1093/hmg/dds073
34. Wang DM, Soni D, Regmi SC, Vogel SM, Tiruppathi C, Hardin J. TAK1 is essential for endothelial barrier maintenance and repair after lung vascular injury. Mol Biol Cell. 2022;33(7):ar65. doi:10.1091/mbc.E21-11-0563
35. Yu M, Wu X, Wang J, et al. Paeoniflorin attenuates monocrotaline-induced pulmonary arterial hypertension in rats by suppressing TAK1-MAPK/NF-κB pathways. Int J Med Sci. 2022;19(4):681–694. doi:10.7150/ijms.69289
36. Hofer-Warbinek R, Schmid JA, Stehlik C, Binder BR, Lipp J, de Martin R. Activation of NF-kappa B by XIAP, the X chromosome-linked inhibitor of apoptosis, in endothelial cells involves TAK1. J Biol Chem. 2000;275(29):22064–22068. doi:10.1074/jbc.M910346199
37. Chang MY, Ho FM, Wang JS, et al. AICAR induces cyclooxygenase-2 expression through AMP-activated protein kinase-transforming growth factor-beta-activated kinase 1-p38 mitogen-activated protein kinase signaling pathway. Biochem Pharmacol. 2010;80(8):1210–1220. doi:10.1016/j.bcp.2010.06.049
38. Liu X, Yang T, Miao L, Mei YA, Hu C. Leukotriene B4 Inhibits L-type calcium channels via p38 Signaling pathway in vascular smooth muscle cells. Cell Physiol Biochem. 2015;37(5):1903–1913. doi:10.1159/000438551
39. Dalvi P, Sharma H, Konstantinova T, Sanderson M, Brien-Ladner AO, Dhillon NK. Hyperactive TGF-β signaling in smooth muscle cells exposed to HIV-protein(s) and Cocaine: Role in pulmonary vasculopathy. Sci Rep. 2017;7(1):10433. doi:10.1038/s41598-017-10438-3
40. Doyon P, Servant MJ. Tumor necrosis factor receptor-associated factor-6 and ribosomal S6 kinase intracellular pathways link the angiotensin II AT1 receptor to the phosphorylation and activation of the IkappaB kinase complex in vascular smooth muscle cells. J Biol Chem. 2010;285(40):30708–30718. doi:10.1074/jbc.M110.126433
41. Song Z, Zhu X, Jin R, et al. Roles of the Kinase TAK1 in CD40-mediated effects on vascular oxidative stress and neointima formation after vascular injury. PLoS One. 2014;9(7):e101671. doi:10.1371/journal.pone.0101671
42. Jadrich JL, O’Connor MB, Coucouvanis E. The TGF beta activated kinase TAK1 regulates vascular development in vivo. Development. 2006;133(8):1529–1541. doi:10.1242/dev.02333
43. Zippel N, Malik RA, Frömel T, et al. Transforming growth factor-β-activated kinase 1 regulates angiogenesis via AMP-activated protein kinase-α1 and redox balance in endothelial cells. Arterios Thromb Vasc Biol. 2013;33(12):2792–2799. doi:10.1161/ATVBAHA.113.301848
44. Wang JH, Lin FL, Chen J, et al. TAK1 blockade as a therapy for retinal neovascularization. Pharmacol Res. 2023;187:106617. doi:10.1016/j.phrs.2022.106617
45. Shang Y, Doan CN, Arnold TD, et al. Transcriptional corepressors HIPK1 and HIPK2 control angiogenesis via TGF-β-TAK1-dependent mechanism. PLoS Biol. 2013;11(4):e1001527. doi:10.1371/journal.pbio.1001527
46. Wang JH, Tseng CL, Lin FL, et al. Topical application of TAK1 inhibitor encapsulated by gelatin particle alleviates corneal neovascularization. Theranostics. 2022;12(2):657–674. doi:10.7150/thno.65098
47. Kim SI, Lee SY, Wang Z, et al. TGF-β-activated kinase 1 is crucial in podocyte differentiation and glomerular capillary formation. J Am Soc Nephrol. 2014;25(9):1966–1978. doi:10.1681/ASN.2013030252
48. Hu H, Lee SR, Bai H, et al. TGFβ (Transforming Growth Factor-Beta)-activated Kinase 1 Regulates Arteriovenous Fistula Maturation. Arterios Thromb Vasc Biol. 2020;40(7):e203–e213. doi:10.1161/ATVBAHA.119.313848
49. Shen Q, Chen Z, Zhao F, et al. Reversal of prolonged obesity-associated cerebrovascular dysfunction by inhibiting microglial Tak1. Nat Neurosci. 2020;23(7):832–+. doi:10.1038/s41593-020-0642-6
50. Huang CY, Sheu WHH, Chiang AN. Docosahexaenoic acid and eicosapentaenoic acid suppress adhesion molecule expression in human aortic endothelial cells via differential mechanisms. Mol Nutr Food Res. 2015;59(4):751–762. doi:10.1002/mnfr.201400687
51. Canugovi C, Stevenson MD, Vendrov AE, et al. Increased mitochondrial NADPH oxidase 4 (NOX4) expression in aging is a causative factor in aortic stiffening. Redox Biol. 2019;26:101288. doi:10.1016/j.redox.2019.101288
52. Yang B, Li W, Zheng Q, et al. Transforming growth factor β-activated kinase 1 negatively regulates interleukin-1α-induced stromal-derived factor-1 expression in vascular smooth muscle cells. Biochem Biophys Res Commun. 2015;463(1–2):130–136. doi:10.1016/j.bbrc.2015.05.047
53. Courboulin A, Tremblay VL, Barrier M, et al. Krüppel-like factor 5 contributes to pulmonary artery smooth muscle proliferation and resistance to apoptosis in human pulmonary arterial hypertension. Respir Res. 2011;12(1):128. doi:10.1186/1465-9921-12-128
54. Yaron A, Hatzubai A, Davis M, et al. Identification of the receptor component of the IkappaBalpha-ubiquitin ligase. Nature. 1998;396:(6711):590–594. doi:10.1038/25159
55. Ridder DA, Wenzel J, Müller K, et al. Brain endothelial TAK1 and NEMO safeguard the neurovascular unit. J Exp Med. 2015;212(10):1529–1549. doi:10.1084/jem.20150165
56. Fritsch M, Gunther SD, Schwarzer R, et al. Caspase-8 is the molecular switch for apoptosis, necroptosis and pyroptosis. Nature. 2019;575:(7784):683–+. doi:10.1038/s41586-019-1770-6
57. Naito MG, Xu D, Amin P, et al. Sequential activation of necroptosis and apoptosis cooperates to mediate vascular and neural pathology in stroke. Proc Natl Acad Sci. 2020;117(9):4959–4970. doi:10.1073/pnas.1916427117
58. Molema G, Zijlstra JG, van Meurs M, JAAM K. Renal microvascular endothelial cell responses in sepsis-induced acute kidney injury. Nat Rev Nephrol. 2022;18(2):95–112. doi:10.1038/s41581-021-00489-1
59. Shu Z, Chen S, Xiang H, et al. AKT/PACS2 Participates in Renal Vascular Hyperpermeability by Regulating Endothelial Fatty Acid Oxidation in Diabetic Mice. Front Pharm. 2022;13:876937. doi:10.3389/fphar.2022.876937
60. Fu Q, Yu W, Fu S, Xu Z, Zhang S. MicroRNA-449c-5p alleviates lipopolysaccharide-induced HUVECs injury via inhibiting the activation NF-κb signaling pathway by TAK1. Mol Immunol. 2022;146:18–26. doi:10.1016/j.molimm.2022.03.123
61. Hoefflin R, Harlander S, Schäfer S, et al. HIF-1α and HIF-2α differently regulate tumour development and inflammation of clear cell renal cell carcinoma in mice. Nat Commun. 2020;11(1):1–21. doi:10.1038/s41467-020-17873-3
62. Rastogi S, Aldosary S, Saeedan AS, Ansari MN, Singh M. NF-κB mediated regulation of tumor cell proliferation in hypoxic microenvironment. Front Pharm. 2023;14. doi:10.3389/fphar.2023.1108915
63. Yeh YH, Hsiao HF, Yeh YC, Chen TW, Li TK. Inflammatory interferon activates HIF-1α-mediated epithelial-to-mesenchymal transition via PI3K/AKT/mTOR pathway. J Exp Clin Cancer Res. 2018;37(1):70. doi:10.1186/s13046-018-0730-6
64. Safina A, Ren M-Q, Vandette E, Bakin AV. TAK1 is required for TGF-β1-mediated regulation of matrix metalloproteinase-9 and metastasis. Oncogene. 2008;27(9):1198–1207. doi:10.1038/sj.onc.1210768
65. Mukwaya A, Mirabelli P, Lennikov A, et al. Repeat corneal neovascularization is characterized by more aggressive inflammation and vessel invasion than in the initial phase. Invest Ophthalmol Vis Sci. 2019;60(8):2990–3001. doi:10.1167/iovs.19-27591
66. Wang Y, Mao Y, Zhang X, et al. TAK1 may promote the development of diabetic nephropathy by reducing the stability of SnoN protein. Life Sci. 2019;228:1–10. doi:10.1016/j.lfs.2019.04.058
67. Lok CE, Huber TS, Lee T, et al. KDOQI Clinical practice guideline for vascular access: 2019 update. Am J Kidney Dis. 2020;75(4 Suppl 2):S1–S164. doi:10.1053/j.ajkd.2019.12.001
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.