Back to Journals » International Journal of General Medicine » Volume 15
Systematic Analysis of E2F Expression and Its Relation in Colorectal Cancer Prognosis
Authors Xu Z, Qu H, Ren Y, Gong Z, Ri HJ ![]() , Zhang F, Shao S, Chen X, Chen X
, Zhang F, Shao S, Chen X, Chen X ![]()
Received 7 February 2022
Accepted for publication 22 April 2022
Published 11 May 2022 Volume 2022:15 Pages 4849—4870
DOI https://doi.org/10.2147/IJGM.S352141
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Scott Fraser
ZhaoHui Xu,1,2,* Hui Qu,1,2,* YanYing Ren,1 ZeZhong Gong,1,2 Hyok Ju Ri,1,2 Fan Zhang,1,* Shuai Shao,1 XiaoLiang Chen,1 Xin Chen1
1Department of Hernia and Colorectal Surgery, The Second Hospital of Dalian Medical University, Dalian, People’s Republic of China; 2Graduate School of Dalian Medical University, Dalian, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xin Chen, Tel +86 17709872266, Email [email protected]
Background: The E2 factor (E2F) family of transcription factors is dysregulated in numerous cancer types and may play a vital role in the development of various malignancies. However, to our knowledge, the specific function of each of the E2Fs and their relation to the disease prognosis of colorectal cancer (CRC) patients remain unknown.
Materials and Methods: We used different publicly available databases and tools, such as ONCOMINE, GEPIA2, UALCAN, cBioPortal, Kaplan–Meier plotter, Metascape, and TIMER analysis, to do an in silico exploration of the potential roles of E2Fs in CRC.
Results: In our analyses, we found a downregulation of E2F2 expression and an upregulation of E2F1 and E2F3-8 expression in CRC tissues compared to normal controls. These findings were consistent with our subgroup analysis using the different clinicopathological features of CRC patients. Furthermore, overexpression of E2F3 and E2F4 were significantly correlated with worse overall survival (OS) in colon cancer patients. Meanwhile, low levels of E2F2 resulted in a shorter OS in rectal cancer patients. The E2F family members had varying degrees of genetic alterations with the highest alteration rate observed in E2F1 (23%). Interestingly, a moderate positive express correlation had been found in the following E2F family members: E2F1 with E2F4, E2F2 with E2F7, E2F2 with E2F8, and E2F7 with E2F8. In addition, spearman analysis revealed that E2Fs have a strong positive correlation with the critical oncogenes in CRC patients. Lastly, the expression of E2Fs was significantly associated with the infiltration of six immune cells.
Conclusion: In this study, we found that E2F2, E2F3, and E2F4 have the potential to be novel prognostic biomarkers in CRC. The role of these E2F family members in disease pathology may be related to their functions in cell cycle regulation, therapeutic resistance, immune cell infiltration, and epithelia-to-mesenchymal transition. Further studies are required to validate our results; however, our findings may help provide a foundation for broadening our understanding of CRC pathology.
Keywords: colorectal cancer, E2F, biomarkers, prognosis, bioinformatics
Introduction
Colorectal cancer (CRC) is the third most common aggressive cancer (10.2% of the total cases) and the second leading cause of cancer deaths (9.2% of the total cancer deaths) worldwide.1 In recent years, although CRC screening, diagnosis, and therapies have made encouraging improvements, disease prognosis remains poor, especially in patients with advanced stages.2
Genetic information is increasingly becoming a part of patient medical records due to advances in genomics.2,3 This is further augmented by thousands of gene expression and copy number variation data published online. The availability of these datasets provides the potential for the development of modern diagnostic methodologies. In contrast, the sensitivity and specificity of the traditional laboratory tests, such as the use of the classic serum biomarkers remain limited.4 It is therefore important to identify new potential reliable genetic biomarkers and therapeutic targets to enhance the precision of treatments and ultimately improve patient prognosis.
The E2 factors (E2Fs) correspond to a family of transcription factors in higher eukaryotes, which includes eight subgroups from E2F1 to E2F8.5,6 Some E2Fs have been classified as transcriptional activators, such as E2F1, E2F2, and E2F3a, while others have been shown to act as transcriptional repressors, such as E2F3b and E2F4-8.7 These factors play major regulatory roles in cellular proliferation, differentiation, DNA repair, cell cycle, and apoptosis in mammalian cells.6 Interestingly, the dysregulation of E2Fs has been identified in several human malignancies, including thyroid cancer,8 breast cancer,9–11 lung cancer,12 hepatocellular carcinoma,6 gastrointestinal cancer,7,13 prostate cancer,14,15 and ovarian cancer.16 Recent studies have shown that E2Fs not only regulate the normal cell cycle but also participate in tumor-related metabolic reprogramming and maintenance of the tumor microenvironment, hence playing an important role in the occurrence and development of tumors. Specifically, E2F1 has been suggested to exhibit a key role in the metabolic reprogramming of lung cancer-related fibroblasts and contribute to the development and progression of lung cancer.17 Furthermore, E2F1 has also been suggested to be a major transcriptional activator in renal cell carcinoma.18 However, the specific mechanism of E2Fs in tumor development remains unresolved.7 To the best of our knowledge, no bioinformatic analyses and reports have been made to identify the roles of E2Fs in CRC. In this study, using a variety of online tools and large databases, we evaluated the expression, genetic alterations, and clinical relationships of distinct E2Fs in CRC patients to predict their biological functions and potential prognostic values.
Materials and Methods
No ethics committee approval was required since all the datasets were obtained from online public databases.
ONCOMINE Database
We evaluated the mRNA expression of eight E2F family members in different cancer types and normal samples using data from the publicly online microarray database and data mining platform, Oncomine 4.5.19 The corresponding cut-off threshold values and parameters were set at fold change (FC)=2.0, p<0.01, gene rank= top 10%, and data type is mRNA.
UALCAN
UALCAN20(http://ualcan.path.uab.edu) is an interactive web resource that can be used for various in-depth analyses using -omics and clinical data from 31 cancer types based on the Cancer Genome Atlas (TCGA). In our report, the colon adenocarcinoma (COAD) dataset and rectum adenocarcinoma (READ) datasets were selected to analyze. We further verify the differential expression and methylation patterns of the E2Fs in patients with different CRC subtypes via UALCAN. A p<0.05 was considered statistically significant.
GEPIA2 Database
Gene Expression Profiling Interactive Analysis 2 (http://gepia2.cancer-pku.cn/, GEPIA2) is a web-based tool that can be used for comprehensive analyses of the different TCGA RNA-seq and Genotype-Tissue Expression (GTEx) project data.21 GEPIA2 can provide a series of customized analysis. Subsequently, we used the GEPIA2 to analyze pathological stage analysis, survival analysis, correlative analysis, and similar gene detection of E2Fs family members. The correlation of non-normally distributed variables was determined using the Spearman correlation test. A p<0.05 was considered statistically significant.
Kaplan-Meier Plotter
The Kaplan-Meier plotter (www.kmplot.com, K-M) is a tool that can be used to estimate the effect of various genes on the survival of patients across 21 common cancer types.22 Patients with READ were classified into high and low-expressing groups based on the median mRNA expression and K-M survival plots were obtained for each group. This tool was also used to determine hazard ratios, 95% CI values, and p values. A p<0.05 was considered statistically significant.
cBioPortal
The cBioPortal Cancer Genomics database (www.cbioportal.org) is an open-access web platform for visualization and interactive exploration of multidimensional cancer genomics datasets.23 For our analyses, the TCGA-CRC RNA-seq dataset (TCGA PanCancer Atlas, RNA Seq V2 RSEM with a z-score of threshold ±2.0) was obtained from cBioPortal. Using this dataset, the relationships between E2Fs gene alterations, overall survival (OS), and disease-free survival (DFS) in CRC patients were analyzed. In addition, the Spearman correlation analysis was used to determine the relationship among the different E2F family members.
Metascape
Metascape (http://metascape.org) is a continuously updated web-based portal that is used for gene annotations and enrichment analyses.24 Using the custom analysis function of this tool, the enrichment of E2F genes with other similar genes was demonstrated.
TIMER Analysis
Tumor IMmune Estimation Resource (https://cistrome.shinyapps.io/timer/, TIMER) is a web server used for comprehensive analyses on the clinical impact of immune cell infiltration across diverse cancer types.25 Scatterplots were generated to explore the correlation of E2F expression with the infiltration of various immune cells, such as B cells, CD4+ T cells, CD8+ T cells, neutrophils, macrophages, and dendritic cells, in CRC. A p<0.05 was considered statistically significant.
Results
Differential Gene Expression of E2F Family Members
We evaluated the mRNA expression of the different E2F family members in CRC patients using Oncomine 4.5 (Figure 1). Our analysis found that the expression levels of E2F1, E2F3, and E2F5-8 were significantly increased in CRC samples compared to normal samples, whereas the expression of E2F2 was reduced.
 |
Figure 1 The transcriptional expression of E2Fs in 20 types of cancer (ONCOMINE Database). Notes: The numbers in colored cells show the numbers of datasets with obeviously significant mRNA upregulation (red) or downregulation (blue) of the target gene. Cell color is determined by the best gene rank percentile for the analyses within the cell. |
Detailed information regarding our analyses on the E2Fs in this study is shown in Table 1. One of our findings is a high expression of E2F1 in the TCGA (FC=3.098, p=4.51E-7; FC=2.056, p=1.50E-11) and the Hong statistics datasets26 (FC=2.068, p=2.79E-9). In contrast, E2F2 was found to be downregulated in colon carcinoma epithelia27 (FC=−2.139, p=4.45E-8) and cecum adenocarcinoma (FC=−2.049, p=5.67E-9). Meanwhile, a high E2F3 mRNA expression was also found in the Kaiser colon cancer dataset28 (FC=2.109, p=6.43E-5) as well as in the Hong colorectal cancer dataset26 (FC=2.940, p=5.37E-8). Furthermore, E2F5 mRNA was observed to be overexpressed in CRC with fold changes of 2.061–3.352.27 In the TCGA colorectal datasets, E2F6 was found to be highly expressed in both adenocarcinoma and mucinous adenocarcinoma compared to normal controls (FC=2.294–2.588, p<0.05). Interestingly, E2F7 was found to be highly upregulated in numerous datasets, including the TCGA, Skrzypczak,27 Hong,26 and Gaedcke datasets.29 As for E2F4 and E2F8, no relevant detailed differential expression datasets information in CRC were obtained from Oncomine 4.5.
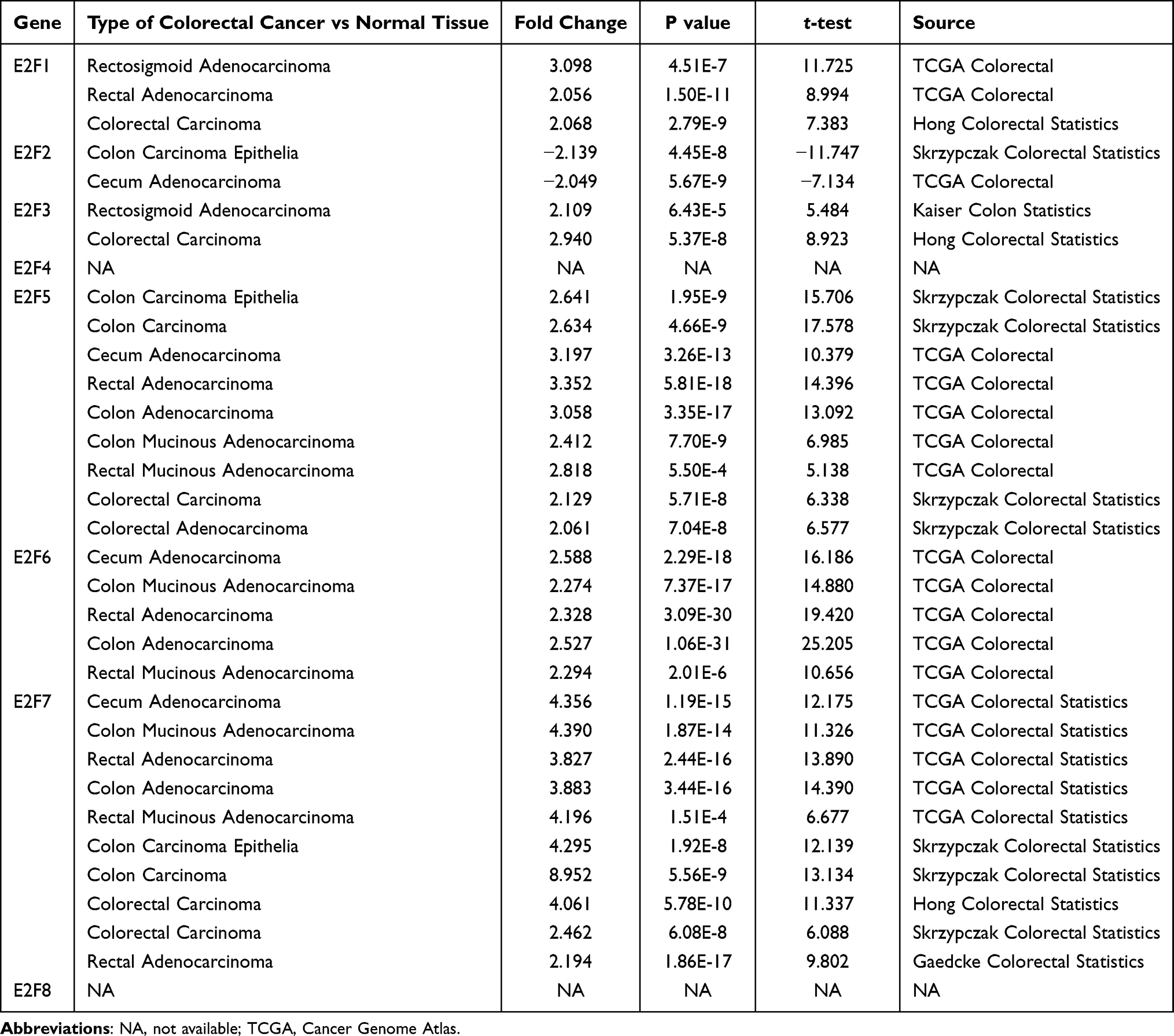 |
Table 1 Datasets of the E2F Family Members in Patients with CRC (ONCOMINE Database) |
A complementary analysis of the TCGA data was done using the UALCAN database to further validate the differential expression levels of E2Fs in tumor and normal tissues. The results were slightly different from that of the Oncomine 4.5 analysis. E2F1 and E2F3-8 were found to be overexpressed in the colon and rectal cancer subgroups compared to the normal controls, whereas the E2F2 mRNA expression was downregulated in these subgroups (Figure 2).
 |
Figure 2 The mRNA expression of E2F family members in patients with COAD (A–H) or READ (I–P) and normal control (UALCAN Database). Notes: p-value significant code:*p<0.05; ***p < 0.001. |
Correlation of E2Fs with Clinicopathological Features in CRC
We evaluated the expression levels of E2Fs and their correlation with the tumor staging in CRC using data from the GEPIA2 database. Significant differential expression levels of E2F3 and E2F5 were found between the different tumor stages of CRC (p<0.05), whereas other E2F family members had no such differential expression between stage groups (Figure 3).
 |
Figure 3 The expression of E2Fs in patients with CRC across different cancer stages (GEPIA2). Notes: E2F1 (A), E2F2 (B), E2F3 (C), E2F4 (D), E2F5 (E), E2F6 (F), E2F7 (G), E2F8 (H). |
Subgroup analyses based on different histological subtypes (adenocarcinoma and mucinous adenocarcinoma), gender, TP53 mutation status, and nodal metastasis status (N0, N1, N2, and N3) also showed high expression levels of E2F1 and E2F3-8, and low expression levels of E2F2 in CRC patients (Figures 4 and 5). These data further suggest that E2Fs may exert a significant contribution to the tumorigenesis and progression of CRC.
 |
Figure 4 Relationships between mRNA expression levels of eight E2F family members and the clinicopathological features of colonic cancers (UALCAN Database). Notes: p-value significant code:*p<0.05; **p<0.01; ***p<0.001. |
 |
Figure 5 Relationships between mRNA expression levels of eight E2F family members and the clinicopathological features of rectal cancers (UALCAN Database). Notes: p-value significant code:*p<0.05; **p<0.01; ***p<0.001. |
Prognostic Value of the Differential Expression of E2Fs
We evaluated the prognostic value of the differentially expressed E2F genes in CRC patients using GEPIA2 and K-M plotter (Figure 6). Overexpression of E2F3 and E2F4 mRNAs were significantly associated with a shorter OS in the colon cancer subgroup. Meanwhile, a low E2F2 mRNA expression was significantly correlated with a worse OS in the rectal cancer subgroup.
 |
Figure 6 The prognostic value of E2Fs famliy in patients with COAD (A–H) and READ (I–P) by using GEPIA2 (A–H) and K-M plotter (I–P). Notes: E2F1 (A and I), E2F2 (B and J), E2F3 (C and K), E2F4 (D and L), E2F5 (E and M), E2F6 (F and N), E2F7 (G and O), E2F8 (H and P). |
E2Fs Promoter Methylation Status in CRC
As previously described, differential expression of the various E2Fs can be observed in CRC patients. To further elucidate the mechanism which leads to the aberrant expression of these genes, we examined the methylation status of the E2F family members using UALCAN. In the COAD subgroup, significantly lower promoter methylation levels for E2F5 and E2F6 were observed in tumor samples compared with normal controls (Figure 7). Similarly, the methylation levels of E2F1, E2F3, E2F5, and E2F6 promoters in READ were significantly lower than those in the normal samples (Figure 7). Furthermore, a similar methylation status for these E2F promoters was also observed during subgroup analyses according to age, weight, nodal metastasis status, and tumor stage (Figures S1 and S2). It is well-known that DNA methylation is a type of epigenetic modification that can silence gene expression.30 In our study, the hypomethylated status of E2F1, E2F3, E2F5, and E2F6 promoters in CRC may partially explain the observed elevated expression levels of these genes.
 |
Figure 7 The promoter methylation of E2Fs in COAD (A–H) and READ (I–P) (UALCAN Database). E2F1 (A and I), E2F2 (B and J), E2F3 (C and K), E2F4 (D and L), E2F5 (E and M), E2F6 (F and N), E2F7 (G and O), and E2F8 (H and P). Abbreviation: ns, no significance. Notes: p-value significant code:*p < 0.05; **p<0.01;***p < 0.001. |
Correlations Among E2Fs Gene Alterations, OS, and DFS for CRC Patients
We investigated the relationship between the genetic alterations in CRC and the disease prognosis of patients using the cBioPortal online tool. At least five genetic alterations were identified in different subtypes of CRC, with the primary alteration type being mRNA amplification (Figure 8A). In the 592 total sequenced CRC patients’ samples, genetic alterations were detected in 324 CRC patients with the alteration rate observed at 55% (Figure 8B). In addition, the alteration rates of E2F1, E2F2, E2F3, E2F4, E2F5, E2F6, E2F7, and E2F8 were 23%, 4%, 8%, 9%, 19%, 8%, 9%, and 7%, respectively. However, no significant relationships were observed between the alteration frequency of E2Fs and OS or DFS of CRC patients (Figure 8C and D).
 |
Figure 8 E2F family members gene expression and alteration analysis in patients with CRC (cBioportal). Notes: (A and B) Summary of the visualized results of different alteration types in the E2F family. (C) Survival plots of OS in cases with or without E2F alterations. (D) Survival plots of DFS in cases with or without E2F alterations. (E) Spearman correlation of E2F family members. (F–I) showed the correlation between E2F1 and E2F4, E2F2 and E2F7, E2F2 and E2F8, and E2F7 and E2F8, respectively. |
We also determined the Spearman correlations among the different E2F family members in CRC patients using the cBioPortal online tool (Figure 8E). Moderate positive correlations in the following E2F family members were observed: E2F1 with E2F4, E2F2 with E2F7, E2F2 with E2F8, and E2F7 with E2F8 (Figure 8F–I).
Correlation of E2Fs with Specific Key Oncogenes in CRC
Emerging evidence suggests that E2Fs are involved in cell cycle progression. To support this, we explored the correlations of E2Fs with well-known cell cycle and proliferation oncogenes, such as FOXM1,31 MKI67,32 TRIP13,33 XPO5,34 MELK,35 and AURKA36 in CRC patients. Furthermore, we also determined the correlation of E2Fs with key oncogenes in CRC, including the cancer stem cell (CSC) related oncogenes CD26,37 CD44,38 Lgr5,39,40 EPHB2,40 NANOG,41 and OLFM4,42 the potential therapeutic resistance oncogenes BMI1,43 SNHG15,42 EPHA2,44 LRP1,45 IDO1,46 and MRP5,43 and the epithelial-to-mesenchymal transition (EMT) oncogenes SNAI1, SLUG, VIM, CDH1, and CDH2.47 Strong correlations of E2F1, E2F2, E2F7, and E2F8 with six cell cycle oncogenes in both COAD and READ groups were observed (Figures 9 and 10). Moreover, we also found a certain correlation between BMI1, SNHG15, EPHA2, and CDH1 with E2Fs (Figures 9 and 10). Meanwhile, none of the E2Fs were found to be strongly correlated with CSC-associated oncogenes. Our results are summarized in Tables S1 and S2. As observed, the overexpression of E2Fs in CRC patients may correlate with cell proliferation, therapeutic resistance, and EMT of tumor cell lines.
 |
Figure 9 Relationship between E2F family members and well-known oncogenes ((A) FOXM1, (B) MKI67, (C) BMI1, (D) SNHG15, (E) CDH1) in patients with COAD (GEPIA2). |
 |
Figure 10 Relationship between E2F family members and well-known oncogenes ((A) FOXM1, (B) MKI67, (C) BMI1, (D) SNHG15, (E) CDH1) in patients with READ (GEPIA2). |
Functional Enrichment of E2Fs and Other Similar Genes in Patients with CRC
We used GEPIA2 to explore E2F-like genes in CRC samples. The top 100 similar genes for each E2F member were identified for further analysis. Functional pathways involving these genes were predicted using the Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genome (KEGG), and protein-protein interaction (PPI) enrichment analyses in the Metascape (Figures 11 and 12).
 |
Figure 11 The functions and pathway of E2Fs and their similar genes in patients with CRC. Notes: (A) GO anslysis-Biological Process. (B) GO analysis-Cellular Component. (C) GO analysis-Molecular Functions. (D) KEGG analysis. |
 |
Figure 12 The protein-protein interactions (PPI) of E2Fs family and their similar genes (Metascape). Notes: (A) Network of enriched terms of E2Fs and similar genes. (B) PPI enrichment analysis of these genes and MCODE (Molecular Complex Detection) components identified in the gene lists. |
GO enrichment analyses evaluate the functional roles of target genes based on three parts: biological processes (BP), cellular components (CC), and molecular functions (MF). Our analyses revealed that the E2Fs and similar genes in CRC are significantly involved in the regulation of the cell cycle process (GO: 0010564), DNA repair (GO: 0006281), nuclear division (GO: 0000280), DNA replication (GO: 0006260), microtubule cytoskeleton organization (GO: 0000226), signal transduction by p53 class mediator (GO: 0072331), and positive regulation of cell cycle (GO: 0045787). Furthermore, chromosomal region (GO: 0098687), helicase activity (GO: 0004386), and transcription factor binding (GO: 0008134) were also significantly controlled by E2Fs.
The KEGG pathway analysis showed that cell cycle (hsa04110), DNA replication (hsa03030), homologous recombination (hsa03440), and cellular senescence (hsa04218) were correlated with the functions of E2Fs and similar genes, and may contribute in the tumorigenesis and pathogenesis of CRC.
The constructed PI network associated with the identified genes was mainly related to DNA replication, DNA repair, nuclear division, signal transduction by p53 class mediator, and regulation of cell cycle processes (Figure 12).
The Contribution of E2Fs in Immune Infiltration in CRC Patients
The TIMER database was used to determine the correlation between the E2F mRNA expression and immune cell infiltration in CRC tumors (Figure 13). Analysis of patients’ samples in the COAD subgroup showed that the mRNA expression levels of E2F1, E2F2, E2F3, E2F7, and E2F8 were significantly related to B cell infiltration. Meanwhile, the mRNA expression levels of E2F1, E2F3, E2F4, E2F6, E2F7, and E2F8 were correlated with CD8+ T cells infiltration. A significant relationship between the mRNA expression of E2F2, E2F3, E2F4, E2F6, and E2F8, and CD4+ T cells was also observed. Gene expression of E2F1, E2F3, E2F4, E2F5, E2F6, E2F7, and E2F8 were consistent with macrophage infiltration. On the other hand, gene expression of E2F2, E2F3, E2F6, E2F7, and E2F8 were related to neutrophils infiltration. Lastly, the expression of E2F1, E2F2, E2F3, E2F6, E2F7, and E2F8 had significant correlations with dendritic cell infiltration.
 |
Figure 13 Correlations between differentially expressed E2Fs and immune cell infiltration (TIMER). |
In patients belonging to the READ subgroup, a significant correlations between E2F5 and E2F8 expression with B cell infiltration were observed. The mRNA expression of E2F1, E2F3, E2F5, E2F6, E2F7, and E2F8 were found to correlate with CD8+ T cell infiltration. Gene expression of E2F4 was related to the infiltration of CD4+ T cells. The expression of E2F2 and E2F3 was found to be associated with macrophage infiltration. Meanwhile, the mRNA expression of E2F3, E2F6, E2F7, and E2F8 were related to neutrophil infiltration. No significant association was found between E2Fs mRNA expression and dendritic cell infiltration in this subgroup.
Discussion
The E2Fs family of transcription factors, which is composed of eight different members, has been implicated in numerous human cancer types. Although the roles of some E2F family members in human malignancies have been partially confirmed, no bioinformatics analysis has been done to demonstrate the function of these factors in CRC. To bridge this science gap, we performed an in-depth analyses on the expression patterns, co-expression, genetic alterations, prognostic values, relation to immune infiltration, and potential functions of all accessible E2F family members in CRC patients based on high-quality datasets from public databases. We hope that our work will contribute substantially in the diagnosis and treatment of CRC.
The E2F1, the most classic E2F member, is an intriguing protein. It is a known downstream effector of the retinoblastoma protein (RB) and it drives the progression of the cell cycle from G1 to the S phase.48 Aside from these, it has been demonstrated to regulate cellular proliferation, differentiation, and apoptosis in CRC by activating different downstream effectors.49 For instance, E2F1 overexpression has been shown to enhance DNAJA1 expression in CRC cells, which in turn promotes serosa invasion and lymph node metastasis of malignant cells.50 Notably, E2F1 is a direct functional target of multiple microRNAs (miRNAs), such as miR-526b-3p51 and miR-328-5p,52 which is positively correlated with the malignant phenotypes of CRC. In addition to miRNAs, the oncogenic H19 noncoding RNA can also promote cell proliferation in CRC via the RB1/E2F1 signaling pathway.53 Furthermore, it has been found that E2F1 may contribute to the EMT and aggressiveness of CRC cells by enhancing IL4/STAT6 signaling.54 Another study demonstrated that Mdm2 inhibition could trigger apoptosis by activating the p73- and E2F1-mediated expression of pro-apoptotic proteins in p53 deficient human colon cancer cells.55 Growing evidence suggests that E2F1 overexpression is closely related to the expression of thymidylate synthase in human colorectal primary cancer,56 lung metastasis of colon cancer,57 and promotion of 5-FU resistance. In line with this, it has been found that 5-FU induced antitumor activities in CRC work at least partially by lowering E2F1 expression.49 In this study, we found that the E2F1 expression was higher in CRC samples than in normal samples; however, its expression has no significant association with tumor stage and has very little prognostic value. We hypothesize that it functions as an oncogenic switch in CRC.
E2F2 has been shown to have a dual role by acting as an oncogene when it is upregulated and acting as a suppressor when it is downregulated.58 However, its specific role in CRC is not yet completely elaborated. Studies have shown that E2F2 expression was extremely low in colorectal carcinoma, and it may act as a tumor suppressor.7,59 For example, it has been shown to suppress the proliferation of T lymphocytes.60 Furthermore, E2F2 was proven to be the direct target of miR-155 and miR-31, inhibiting the cell proliferation and cell cycle process in colorectal carcinoma.58,59 Consistent with previous studies, we found that E2F2 mRNA expression was lower in CRC samples compared to normal controls. Moreover, the survival analysis results revealed that decreased E2F2 expression was significantly linked to a worse OS in rectal cancers.
The E2F3 locus expresses two alternative proteins, E2F3a and E2F3b. Its overexpression is a frequent carcinogenic event in human tumorigenesis.8,14,61 E2F3 was identified as a direct target of miR-503. This associated miRNA has been shown to inhibit cellular proliferation, induce apoptosis, and arrest the G0/G1 phase in CRC tissues and cell lines.61 Recently, emerging evidence suggests that circPRTM5 promotes CRC cell proliferation by sponging miR-377 to induce E2F3 expression.62 In our study, E2F3 mRNA was shown to be upregulated in CRC samples compared to normal controls, and its expression was significantly associated with tumor stage. Furthermore, overexpression of E2F3 was significantly correlated with a worse OS in CRC patients. Moreover, we found that E2F3 mRNA expression was significantly correlated with the infiltration of six immune cells. Taken together, we suggest that E2F3 may act as an oncogene in CRC and a new disease prognostic marker.
E2F4 is generally recognized as a potential oncogene in CRC. Previous studies have found that the expression of E2F4 and its target cyclin A were both upregulated in human CRC cells.63 Furthermore, E2F4 knockdown inhibits tumor growth by slowing down the G1/S phase transition and decreasing the proliferation rate of colon cancer cells.63 Notably, it has been shown that E2F4 exhibits frequent tumor-specific mutations in CRC with microsatellite instability, which in turn may accelerate tumorigenesis for this type of tumor.64 Similarly, this genetic or epigenetic phenomenon has also been observed in gastric cancer65 and hepatocellular carcinoma.66 In this report, E2F4 expression was found to be upregulated in colon cancer cells compared to the normal counterparts. Moreover, upregulation of E2F4 mRNA was significantly related to a shorter OS, supporting its role as an oncogene in colon cancer.
The high expression and pro-oncogenic activity of E2F5 have been shown in several tumors, including CRC.9,15,67 The miRNAs miR-32, miR-34a, and miR-181a-5p can regulate the proliferation, migration, and invasion of CRC cell lines by targeting E2F5.67–69 Consistent with this, silencing E2F5 inhibited the growth of the human colon cancer cell line SW-948 by inducing apoptosis-related processes and arresting the G2/M step in the cell cycle.68 In our analyses, we found that E2F5 was overexpressed in CRC tissues, but no significant relationships were found between E2F5, tumor stage, OS, and DFS.
An upregulation of E2F6 has been reported in several tumors, such as gastric carcinoma,13 and breast cancer.10 However, no studies have tackled the role of E2F6 in CRC. In this study, the expression levels of E2F6 were increased in CRC; however, no significant correlations were seen between E2F6 and OS or DFS. Future work will be necessary to demonstrate whether E2F6 plays an oncogenic role in CRC.
E2F7 can function both as a transcriptional repressor70 and as an activator.11 In one particular study, a low-frequency missense variant in E2F7 has been associated with CRC susceptibility in a Chinese Han population, suggesting that E2F7 may play a vital role in tumorigenesis and disease development.71 In line with this, it has been demonstrated that ATRA suppresses the genesis and development of human CRC by regulating the miR-3666/E2F7 axis.70 Furthermore, miR-3666/E2F7 may also inhibit the activation of the MAPK/ERK signaling pathway. In our study, we observed that E2F7 was highly expressed in CRC samples, and its expression was not related to tumor stage and has very little prognostic value.
The last member of the family, E2F8, controls the DNA damage-related cell-cycle processes by regulating E2F1 activity. In addition, it is essential for angiogenesis, lymphangiogenesis, and embryonic development.6,72 It has been previously found that E2F8 is upregulated in CRC patients and is a specific target of miR-1258.72 One study has reported that knocking down E2F8 reduces the expression of cell cycle regulators.73 The same group further discovered that E2F8 may promote the growth of human colon cancer cells by modulating the NF-κB pathway. Another recent study also discovered that the small nuclear ribonucleoprotein polypeptide N, an imprinted gene transcriptionally upregulated by E2F8, was found to promote malignant progression of CRC by cell cycle regulation.74 In our analyses, results showed that E2F8 was markedly upregulated in patients with CRC. However, its prognostic role in CRC remains unknown.
Conclusions
In summary, our results revealed that E2Fs were aberrantly expressed in CRC tissues – E2F1 and E2F3-8 were upregulated in CRC tissues compared to their normal counterparts, whereas E2F2 was downregulated in the same samples. Moreover, the similar variation trends of E2Fs were also validated in subgroup analysis by the different clinicopathological features. Furthermore, our results suggest that the promoter hypomethylation of E2Fs may be the cause of the aberrant expression from different perspectives. Additionally, the overexpression of E2F3 and E2F4 were significantly correlated with a worse OS in COAD. Meanwhile, low levels of E2F2 resulted in a shorter OS in READ. These suggest that E2F may play an important role in tumorigenesis and progression in CRC. The specific mechanism may be correlated with E2F-mediated cell cycle regulation, therapeutic resistance, immune cell infiltration, genetic alterations, and EMT. We hope these findings may help broaden our insight into CRC. However, further studies are acquired to validate our findings.
Ethics Approval and Informed Consent
All data for this study were obtained from the ONCOMINE, GEPIA2, UALCAN, cBioPortal, Kaplan-Meier plotter, Metascape, and TIMER database, which are publicly available. We did not obtain these data directly from patients or animals. Therefore, following ethical review, the Second Hospital of Dalian Medical University Ethics Committee granted a status of exemption.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the “1+X” program for Clinical Competency enhancement - Interdisciplinary Innovation Project, The Second Hospital of Dalian Medical University (2022JCXKYB01, 2022JCXKYB20), Dalian Medical Science Research Project (2012023).
Disclosure
The authors reported that they have no conflict of interest.
References
1. Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Jeon J, Du M, Schoen RE, et al. Determining risk of colorectal cancer and starting age of screening based on lifestyle, environmental, and genetic factors. Gastroenterology. 2018;154(8):2152–2164.e2119. doi:10.1053/j.gastro.2018.02.021
3. Chute CG, Kohane IS. Genomic medicine, health information technology, and patient care. JAMA. 2013;309(14):1467–1468. doi:10.1001/jama.2013.1414
4. Das V, Kalita J, Pal M. Predictive and prognostic biomarkers in colorectal cancer: a systematic review of recent advances and challenges. Biomed Pharmacother. 2017;87:8–19. doi:10.1016/j.biopha.2016.12.064
5. Trimarchi JM, Lees JA. Sibling rivalry in the E2F family. Nat Rev Mol Cell Biol. 2002;3(1):11–20. doi:10.1038/nrm714
6. Lv Y, Xiao J, Liu J, et al. E2F8 is a potential therapeutic target for hepatocellular carcinoma. J Cancer. 2017;8(7):1205–1213. doi:10.7150/jca.18255
7. Xanthoulis A, Tiniakos DG. E2F transcription factors and digestive system malignancies: how much do we know? World j Gastroenterol. 2013;19(21):3189–3198. doi:10.3748/wjg.v19.i21.3189
8. Xue C, Cheng Y, Wu J, et al. Circular RNA CircPRMT5 accelerates proliferation and invasion of papillary thyroid cancer through regulation of miR-30c/E2F3 axis. Cancer Manag Res. 2020;12:3285–3291. doi:10.2147/CMAR.S249237
9. Cai C, Huo Q, Wang X, et al. SNHG16 contributes to breast cancer cell migration by competitively binding miR-98 with E2F5. Biochem Biophys Res Commun. 2017;485(2):272–278. doi:10.1016/j.bbrc.2017.02.094
10. Lafta IJ. E2F6 is essential for cell viability in breast cancer cells during replication stress. Turkish J Biol. 2019;43(5):293–304. doi:10.3906/biy-1905-6
11. Sun CC, Li SJ, Hu W, et al. Comprehensive analysis of the expression and prognosis for E2Fs in human breast cancer. Mol ther. 2019;27(6):1153–1165. doi:10.1016/j.ymthe.2019.03.019
12. Cooper CS, Nicholson AG, Foster C, et al. Nuclear overexpression of the E2F3 transcription factor in human lung cancer. Lung Cancer. 2006;54(2):155–162. doi:10.1016/j.lungcan.2006.07.005
13. Li Y, Jiang L, Lv S, et al. E2F6-mediated lncRNA CASC2 down-regulation predicts poor prognosis and promotes progression in gastric carcinoma. Life Sci. 2019;232:116649. doi:10.1016/j.lfs.2019.116649
14. Foster CS, Falconer A, Dodson AR, et al. Transcription factor E2F3 overexpressed in prostate cancer independently predicts clinical outcome. Oncogene. 2004;23(35):5871–5879. doi:10.1038/sj.onc.1207800
15. Li SL, Sui Y, Sun J, et al. Identification of tumor suppressive role of microRNA-132 and its target gene in tumorigenesis of prostate cancer. Int J Mol Med. 2018;41(4):2429–2433. doi:10.3892/ijmm.2018.3421
16. Zhou Q, Zhang F, He Z, et al. E2F2/5/8 serve as potential prognostic biomarkers and targets for human ovarian cancer. Front Oncol. 2019;9:161. doi:10.3389/fonc.2019.00161
17. Shen C, Chen X, Xiao K, et al. New relationship of E2F1 and BNIP3 with caveolin-1 in lung cancer-associated fibroblasts. Thoracic Cancer. 2020;11(6):1369–1371. doi:10.1111/1759-7714.13408
18. Pandey N, Lanke V, Vinod PK. Network-based metabolic characterization of renal cell carcinoma. Sci Rep. 2020;10(1):5955. doi:10.1038/s41598-020-62853-8
19. Rhodes DR, Yu J, Shanker K, et al. ONCOMINE: a cancer microarray database and integrated data-mining platform. Neoplasia. 2004;6(1):1–6. doi:10.1016/S1476-5586(04)80047-2
20. Chandrashekar DS, Bashel B, Balasubramanya SAH, et al. UALCAN: a portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia. 2017;19(8):649–658. doi:10.1016/j.neo.2017.05.002
21. Tang Z, Kang B, Li C, et al. GEPIA2: an enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019;47(W1):W556–w560. doi:10.1093/nar/gkz430
22. Nagy Á, Lánczky A, Menyhárt O, et al. Validation of miRNA prognostic power in hepatocellular carcinoma using expression data of independent datasets. Sci Rep. 2018;8(1):9227. doi:10.1038/s41598-018-27521-y
23. Cerami E, Gao J, Dogrusoz U, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012;2(5):401–404. doi:10.1158/2159-8290.CD-12-0095
24. Zhou Y, Zhou B, Pache L, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun. 2019;10(1):1523. doi:10.1038/s41467-019-09234-6
25. Li T, Fan J, Wang B, et al. TIMER: a web server for comprehensive analysis of tumor-infiltrating immune cells. Cancer Res. 2017;77(21):e108–e110. doi:10.1158/0008-5472.CAN-17-0307
26. Hong Y, Downey T, Eu KW, et al. A “metastasis-prone” signature for early-stage mismatch-repair proficient sporadic colorectal cancer patients and its implications for possible therapeutics. Clin Exp Metastasis. 2010;27(2):83–90. doi:10.1007/s10585-010-9305-4
27. Skrzypczak M, Goryca K, Rubel T, et al. Modeling oncogenic signaling in colon tumors by multidirectional analyses of microarray data directed for maximization of analytical reliability. PLoS One. 2010;5:10. doi:10.1371/annotation/8c585739-a354-4fc9-a7d0-d5ae26fa06ca
28. Kaiser S, Park YK, Franklin JL, et al. Transcriptional recapitulation and subversion of embryonic colon development by mouse colon tumor models and human colon cancer. Genome Biol. 2007;8(7):R131. doi:10.1186/gb-2007-8-7-r131
29. Gaedcke J, Grade M, Jung K, et al. Mutated KRAS results in overexpression of DUSP4, a MAP-kinase phosphatase, and SMYD3, a histone methyltransferase, in rectal carcinomas. Genes Chromosomes Cancer. 2010;49(11):1024–1034. doi:10.1002/gcc.20811
30. Moore LD, Le T, Fan G. DNA methylation and its basic function. Neuropsychopharmacology. 2013;38(1):23–38. doi:10.1038/npp.2012.112
31. Wang D, Hu G, Du Y, et al. Aberrant activation of hedgehog signaling promotes cell proliferation via the transcriptional activation of forkhead Box M1 in colorectal cancer cells. J Exp Clin Cancer Res. 2017;36(1):23. doi:10.1186/s13046-017-0491-7
32. Sobecki M, Mrouj K, Colinge J, et al. Cell-cycle regulation accounts for variability in Ki-67 expression levels. Cancer Res. 2017;77(10):2722–2734. doi:10.1158/0008-5472.CAN-16-0707
33. Sheng N, Yan L, Wu K, et al. TRIP13 promotes tumor growth and is associated with poor prognosis in colorectal cancer. Cell Death Dis. 2018;9(3):402. doi:10.1038/s41419-018-0434-z
34. Shigeyasu K, Okugawa Y, Toden S, et al. Exportin-5 functions as an oncogene and a potential therapeutic target in colorectal cancer. Clin Cancer Res. 2017;23(5):1312–1322. doi:10.1158/1078-0432.CCR-16-1023
35. Gong X, Chen Z, Han Q, et al. Sanguinarine triggers intrinsic apoptosis to suppress colorectal cancer growth through disassociation between STRAP and MELK. BMC Cancer. 2018;18(1):578. doi:10.1186/s12885-018-4463-x
36. Wang-Bishop L, Chen Z, Gomaa A, et al. Inhibition of AURKA reduces proliferation and survival of gastrointestinal cancer cells with activated KRAS by preventing activation of RPS6KB1. Gastroenterology. 2019;156(3):662–675.e667. doi:10.1053/j.gastro.2018.10.030
37. Pang R, Law WL, Chu AC, et al. A subpopulation of CD26+ cancer stem cells with metastatic capacity in human colorectal cancer. Cell Stem Cell. 2010;6(6):603–615. doi:10.1016/j.stem.2010.04.001
38. Boman BM, Huang E. Human colon cancer stem cells: a new paradigm in gastrointestinal oncology. J clin oncol. 2008;26(17):2828–2838. doi:10.1200/JCO.2008.17.6941
39. Barker N, van Es JH, Kuipers J, et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449(7165):1003–1007. doi:10.1038/nature06196
40. Merlos-Suárez A, Barriga FM, Jung P, et al. The intestinal stem cell signature identifies colorectal cancer stem cells and predicts disease relapse. Cell Stem Cell. 2011;8(5):511–524. doi:10.1016/j.stem.2011.02.020
41. Zhang J, Espinoza LA, Kinders RJ, et al. NANOG modulates stemness in human colorectal cancer. Oncogene. 2013;32(37):4397–4405. doi:10.1038/onc.2012.461
42. van der Flier LG, Haegebarth A, Stange DE, et al. OLFM4 is a robust marker for stem cells in human intestine and marks a subset of colorectal cancer cells. Gastroenterology. 2009;137(1):15–17. doi:10.1053/j.gastro.2009.05.035
43. Liu T, Zhang X, Du L, et al. Exosome-transmitted miR-128-3p increase chemosensitivity of oxaliplatin-resistant colorectal cancer. Mol Cancer. 2019;18(1):43. doi:10.1186/s12943-019-0981-7
44. Martini G, Cardone C, Vitiello PP, et al. EPHA2 is a predictive biomarker of resistance and a potential therapeutic target for improving antiepidermal growth factor receptor therapy in colorectal cancer. Mol Cancer Ther. 2019;18(4):845–855. doi:10.1158/1535-7163.MCT-18-0539
45. Lee KJ, Ko EJ, Park YY, et al. A novel nanoparticle-based theranostic agent targeting LRP-1 enhances the efficacy of neoadjuvant radiotherapy in colorectal cancer. Biomaterials. 2020;255:120151. doi:10.1016/j.biomaterials.2020.120151
46. Chen B, Alvarado DM, Iticovici M, et al. Interferon-induced IDO1 mediates radiation resistance and is a therapeutic target in colorectal cancer. Cancer Immunol Res. 2020;8(4):451–464. doi:10.1158/2326-6066.CIR-19-0282
47. Hu Y, Dai M, Zheng Y, et al. Epigenetic suppression of E-cadherin expression by Snail2 during the metastasis of colorectal cancer. Clin Epigenetics. 2018;10(1):154. doi:10.1186/s13148-018-0592-y
48. Jiang H, Martin V, Alonso M, et al. RB-E2F1: molecular rheostat for autophagy and apoptosis. Autophagy. 2010;6(8):1216–1217. doi:10.4161/auto.6.8.13695
49. Fang Z, Lin M, Li C, et al. A comprehensive review of the roles of E2F1 in colon cancer. Am J Cancer Res. 2020;10(3):757–768.
50. Yang S, Ren X, Liang Y, et al. KNK437 restricts the growth and metastasis of colorectal cancer via targeting DNAJA1/CDC45 axis. Oncogene. 2020;39(2):249–261. doi:10.1038/s41388-019-0978-0
51. Fang Z, Yang H, Chen D, et al. YY1 promotes colorectal cancer proliferation through the miR-526b-3p/E2F1 axis. Am J Cancer Res. 2019;9(12):2679–2692.
52. Zhou C, Liu HS, Wang FW, et al. circCAMSAP1 promotes tumor growth in colorectal cancer via the miR-328-5p/E2F1 axis. Mol ther. 2020;28(3):914–928. doi:10.1016/j.ymthe.2019.12.008
53. Ohtsuka M, Ling H, Ivan C, et al. H19 noncoding RNA, an independent prognostic factor, regulates essential Rb-E2F and CDK8-β-catenin signaling in colorectal cancer. EBioMedicine. 2016;13:113–124. doi:10.1016/j.ebiom.2016.10.026
54. Chen J, Gong C, Mao H, et al. E2F1/SP3/STAT6 axis is required for IL-4-induced epithelial-mesenchymal transition of colorectal cancer cells. Int J Oncol. 2018;53(2):567–578. doi:10.3892/ijo.2018.4429
55. Ray RM, Bhattacharya S, Johnson LR. Mdm2 inhibition induces apoptosis in p53 deficient human colon cancer cells by activating p73- and E2F1-mediated expression of PUMA and Siva-1. Apoptosis. 2011;16(1):35–44. doi:10.1007/s10495-010-0538-0
56. Varghese V, Magnani L, Harada-Shoji N, et al. FOXM1 modulates 5-FU resistance in colorectal cancer through regulating TYMS expression. Sci Rep. 2019;9(1):1505. doi:10.1038/s41598-018-38017-0
57. Banerjee D, Gorlick R, Liefshitz A, et al. Levels of E2F-1 expression are higher in lung metastasis of colon cancer as compared with hepatic metastasis and correlate with levels of thymidylate synthase. Cancer Res. 2000;60(9):2365–2367.
58. Li T, Luo W, Liu K, et al. miR-31 promotes proliferation of colon cancer cells by targeting E2F2. Biotechnol Lett. 2015;37(3):523–532. doi:10.1007/s10529-014-1715-y
59. Li T, Yang J, Lv X, et al. miR-155 regulates the proliferation and cell cycle of colorectal carcinoma cells by targeting E2F2. Biotechnol Lett. 2014;36(9):1743–1752. doi:10.1007/s10529-014-1540-3
60. Opavsky R, Tsai SY, Guimond M, et al. Specific tumor suppressor function for E2F2 in Myc-induced T cell lymphomagenesis. Proc Natl Acad Sci U S A. 2007;104(39):15400–15405. doi:10.1073/pnas.0706307104
61. Chang SW, Yue J, Wang BC, et al. miR-503 inhibits cell proliferation and induces apoptosis in colorectal cancer cells by targeting E2F3. Int J Clin Exp Pathol. 2015;8(10):12853–12860.
62. Yang B, Du K, Yang C, et al. CircPRMT5 circular RNA promotes proliferation of colorectal cancer through sponging miR-377 to induce E2F3 expression. J Cell Mol Med. 2020;24(6):3431–3437. doi:10.1111/jcmm.15019
63. Garneau H, Paquin MC, Carrier JC, et al. E2F4 expression is required for cell cycle progression of normal intestinal crypt cells and colorectal cancer cells. J Cell Physiol. 2009;221(2):350–358. doi:10.1002/jcp.21859
64. Paquin MC, Leblanc C, Lemieux E, et al. Functional impact of colorectal cancer-associated mutations in the transcription factor E2F4. Int J Oncol. 2013;43(6):2015–2022. doi:10.3892/ijo.2013.2131
65. Souza RF, Yin J, Smolinski KN, et al. Frequent mutation of the E2F-4 cell cycle gene in primary human gastrointestinal tumors. Cancer Res. 1997;57(12):2350–2353.
66. Park YM, Choi JY, Bae SH, et al. Microsatellite instability and mutations of E2F-4 in hepatocellular carcinoma from Korea. Hepatol Res. 2000;17(2):102–111. doi:10.1016/S1386-6346(99)00066-2
67. Yu C, Sun J, Leng X, et al. Long noncoding RNA SNHG6 functions as a competing endogenous RNA by sponging miR-181a-5p to regulate E2F5 expression in colorectal cancer. Cancer Manag Res. 2019;11:611–624. doi:10.2147/CMAR.S182719
68. Yang F, Chen L, Wang ZJ. MicroRNA-32 inhibits the proliferation, migration and invasion of human colon cancer cell lines by targeting E2F transcription factor 5. Eur Rev Med Pharmacol Sci. 2019;23(10):4156–4163. doi:10.26355/eurrev_201905_17918
69. Lu G, Sun Y, An S, et al. MicroRNA-34a targets FMNL2 and E2F5 and suppresses the progression of colorectal cancer. Exp Mol Pathol. 2015;99(1):173–179. doi:10.1016/j.yexmp.2015.06.014
70. Liu W, Song Y, Zhang C, et al. The protective role of all-transretinoic acid (ATRA) against colorectal cancer development is achieved via increasing miR-3666 expression and decreasing E2F7 expression. Biomed Pharmacother. 2018;104:94–101. doi:10.1016/j.biopha.2018.05.015
71. Guo AY, Zhai K, Xu JL, et al. Identification of a low-frequency missense variant in E2F transcription factor 7 associated with colorectal cancer risk in A Chinese population. APJCP. 2017;18(1):271–275. doi:10.22034/APJCP.2017.18.1.271
72. Zhang Z, Li J, Huang Y, et al. Upregulated miR-1258 regulates cell cycle and inhibits cell proliferation by directly targeting E2F8 in CRC. Cell Prolif. 2018;51(6):e12505. doi:10.1111/cpr.12505
73. Yan PY, Zhang XA. Knockdown of E2F8 suppresses cell proliferation in colon cancer cells by modulating the NF-κB pathway. Ann Clin Lab Sci. 2019;49(4):474–480.
74. Ji M, Ren L, Lv Y, et al. Small nuclear ribonucleoprotein Polypeptide N accelerates malignant progression and poor prognosis in colorectal cancer transcriptionally regulated by E2F8. Front Oncol. 2020;10:561287. doi:10.3389/fonc.2020.561287
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.