")
Back to Journals » Infection and Drug Resistance » Volume 16
Study on the Role of ampG in the Regulation of Plasmid-Mediated ampC -Induced Expression in Klebsiella pneumoniae
Authors Li G, Wang L, Zhang H, Luan Y, Sun Q, Duo L
Received 16 May 2023
Accepted for publication 26 July 2023
Published 24 August 2023 Volume 2023:16 Pages 5587—5598
DOI https://doi.org/10.2147/IDR.S421598
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Guiling Li,1 Li Wang,2 Heguang Zhang,1 Ying Luan,1 Qi Sun,1 Libo Duo1
1Department of Clinical Laboratory, the Second Hospital Affiliated to Harbin Medical University, Harbin, Heilongjiang, 150086, People’s Republic of China; 2Department of Clinical Laboratory, School of Medicine, Chengdu Women’s and Children’s Central Hospital, University of Electronic Science and Technology of China, Chengdu, Sichuan, 611731, People’s Republic of China
Correspondence: Libo Duo, Department of Clinical Laboratory, the Second Hospital affiliated to Harbin Medical University, No. 246 XueFu Road, NanGang District, Harbin, Heilongjiang, 150086, People’s Republic of China, Tel +86 045186605382, Email [email protected]
Objective: In this study, we constructed ampG knock-out and knock-in strains from a clinically isolated Kp1strain carrying ampR-ampC in its plasmid and compared them with the Kp NTUH-K2044 strain to investigate the relationship between ampG and ampR-ampC-induced expression.
Methods: We created the ampG gene deletion mutant strains Kp1-ΔampG and Kp NTUH-K2044-ΔampG with pKO3-km plasmid using homologous recombination technology. We constructed the Kp NTUH-K2044-RC and Kp NTUH-K2044-ΔampG-RC drug resistance model strains with plasmid pACYC184. We constructed the ampG knock-in strains by introducing the ampG genes of Kp1, Enterobacter cloacae 029M, Pseudomonas aeruginosa PAO1, Escherichia coli ATCC25922, and Salmonella typhimurium LT2 into the ampG gene-deleted strains with carrier pet-30a. Real-time polymerase chain reaction (real-time PCR) was used to detect the relative expressions of ampC and ampG mRNAs.
Results: Compared with Kp1, the induction phenotype of the ampC of Kp1-ΔampG strain disappeared, the ampC expression was reduced, and the minimal inhibitory concentration (MIC) values of cefoxitin and ceftazidime significant decrease from 128 μg/mL to 1 μg/mL. Based on Kp1, five strain were successfully constructed to complement the ampG genes from five knock-in strain, and all of the above complemented strains showed inducible expression of ampC and restored the expression of ampG to varying degrees, as well as restored resistance to the antimicrobial drugs cefoxitin and ceftazidime (P < 0.05). The ampC and ampG genes were barely expressed in Kp NTUH-K2044-ΔampG-RC when compared with Kp NTUH-K2044-RC. The expressions of ampG and ampC in each knock-in strain were recovered, the induction phenotype of ampC was restored, and the MIC values of cefoxitin and ceftazidime were increased. (P < 0.05).
Conclusion: In this study, we found that ampG was an essential regulator for the plasmid-mediated ampC-induced expression in K. pneumoniae.
Keywords: ampC inducible expression, AmpG regulator, gene knockout, gene knock-in, Klebsiella pneumoniae
Introduction
The chromosome ampR-ampC module is required for the β-lactam antibiotic resistance in Enterobacteriaceae. The bla ampC gene is elevated in the presence of an inducer (β-lactam antibiotics). The outer membrane protein is encoded by ampG, which is a transmembrane protein that is involved in promoting the recycling of murein degradation products. It influences the production of ampC by transmitting information about changes in biological mediators to the cytoplasm and regulating the expression of related genes, both of which are critical in ampC expression.1,2 Although the ampR-ampC gene is naturally absent on the Klebsiella pneumoniae chromosome, K. pneumoniae Kp1, which carries ampR-ampC in plasmids, was clinically isolated. It exhibits high levels of ampC induced expression (DHA-1 type), which has emerged as a critical element of bacterial drug resistance.3
The Klebsiella pneumoniae plasmid-mediated extended-spectrum β-lactamases (ESBLs) and AmpC are the primary contributors to its multi-drug resistance. The ampC gene is naturally absent from its chromosomes. However, the ampC gene can be introduced via plasmid so as to achieve sustained high expression of ampC, which has become a crucial component of bacterial drug resistance. Without enzyme inhibitors, the production of AmpC results in the hydrolysis of the third-generation cephalosporins, monobactams, and cephamycins antibiotics. The main regulators of chromosome-mediated ampC expression are ampC, ampD, ampR, ampG, and ampE. The transmembrane protein AmpG, located on the outer membrane of the cell, plays a key role in the production of ampC by transmitting information about changes in biological mediators to the cytoplasm and regulating the expression of related genes. However, the role of AmpG in plasmid-mediated ampC-induced expression is unclear.
The majority of current studies on drug resistance in K. pneumoniae focus on the detection of drug resistance phenotypes and genotypes, while the functional analysis of drug resistance genes is rarely reported. The gene knockout technique is the most effective method for studying gene function.4–6 In this study, we used gene knockout technology based on the temperature-sensitive, high-efficiency suicide vector plasmid pKO3-km to knock out the drug-resistant gene ampG in K. pneumoniae, construct the ampG gene deletion mutant strain, and knock in the ampG genes of five kinds of bacteria into the gene knockout strains. In the same genetic background, we took up a more comprehensive study of the differences between the ampG gene deletion strain and the wild-type strain so as to make a more accurate assessment and gather insights into gene function.
Materials and Methods
Materials
The strains and plasmids used in this experiment are listed in Table 1.
 |
Table 1 Strains and Plasmids Used in This Study |
Methods
Construction of the Kp1-ΔampG, Kp NTUH-K2044-ΔampG, Kp NTUH-K2044-RC, and Kp NTUH-K2044-ΔampG-RC strains.
We used PCR to amplify the upstream and downstream homologous arm fragments of the experimental strain ampG. Next, we used the gene splicing by overlap extension PCR (SOE-PCR) method to obtain the upstream and downstream homologous arm fusion fragments of ampG genes, which were cloned to the temperature-sensitive suicide vector pKO3-km after digestion to construct pKO3-km-ΔampG recombinant plasmids. The plasmids were electroporated into competent K. pneumoniae Kp1 and Kp NTUH-K2044 strains, respectively.
We screened the ampG deletion mutant strains of K. pneumoniae, Kp1-ΔampG and Kp NTUH-K2044-ΔampG using homologous recombination technology and the temperature-sensitive characteristics of pKO3-km. Our laboratory team had earlier constructed the recombinant plasmid pACYC184-ampCR (ampC+, ampR+, CmR).7 We then obtained the K. pneumoniae model strains Kp NTUH-K2044-RC and Kp NTUH-K2044-ΔampG-RC carrying the exogenous ampC gene by electroporating the competent Kp NTUH-K2044 and NTUH-K2044-ΔampG with plasmids carrying the ampC and ampR genes of the Kp1strain.
Construction of ΔampG::ampG Knock-in Strains
Construction of the Pet-30a-ampG Recombinant Plasmid
The five bacterial DNAs were used as templates to amplify the ampG genes of each strain, and the corresponding primers are listed in Table 2. The 50 μL PCR reaction system was as follows: 5×buffer 10 μL, dNTP 4 μL, ampG-F/R 2 μL, bacterial genomic DNA template 2 μL, PrimeSTAR HS DNA Polymerase 1 μL, sterilized deionized water 29 μL. The reaction conditions were as follows: 98°C for 10s, 58°C for 5s, 72°C for 1 min 30s for a total of 30 cycles and then 72°C for 7 min. We used electrophoresis to detect the specificity and concentration of PCR products and the PCR products were purified and recovered with reagent kits for use later. We subjected the above-purified PCR products of the five bacterial ampG genes and plasmid pet-30a to double enzyme digestion. The ampG fragment enzyme digestion system (20 μL) contained 8 μL of gene product, 1 μL of SacI endonuclease, 2 μL of buffer, 1 μL of EcoRI endonuclease, and 8 μL of water, which was incubated at 37°C for 24 hours. The plasmid pet-30a digestion system (50 μL) contained 25 μL of plasmid, 2.5 μL of SacI endonuclease, 5 μL of buffer, 2.5 μL of EcoRI endonuclease, and 15 μL of water, which was incubated at 37°C for 24 hours. The byproduct of the enzyme digestion was isolated and recovered. The digested ampG gene fragment and pet-30a were ligated using T4DNA ligase and transformed into competent E. coli DH5a cells. Then it was coated on LB medium (containing km 50 μg/mL) and cultured overnight at 30°C. The monoclonal colonies were selected, and plasmids were extracted for double enzyme digestion and identification to obtain recombinant plasmid pet-30a-ampG for the construction of complementary strains.
 |
Table 2 Primer Sequences of the Klebsiella Pneumoniae ampG Knock-in Strains |
Construction of ΔampG::ampG Strain
We used the solid plate method to prepare the Kp1-ΔampG, Kp NTUH-K2044-RC, and KpNTUH-K2044-ΔampG-RC competent cells.8 We obtained a total of 100 μL competent cells, which were placed along with 400 ng of pet-30a-ampG plasmid in an electric shock cup with a diameter of 0.2 cm. The cup was chilled on ice for 15 minutes, and electroporated with the parameters of 25 μF, 200 Ω, and 2.5 kV. It was immediately transferred into 800 μL LB liquid medium and incubated at 30°C for 3 hours. The transformed products were all coated onto the LB plates (Km, 50 μg/mL) and incubated overnight at 30°C. Monoclonal colonies were selected, and plasmids were extracted. Each recombinant plasmid was identified with primers as listed in Table 2. Positive clones were determined when the products obtained by PCR amplification were consistent with the ampG fragments of each strain. The obtained knock-in strains were named as:
Kp1-ΔampG knock-in strains: Kp1-ΔampG::Kp1; Kp1-ΔampG:: 029M; Kp1-ΔampG::PAO1; Kp1-ΔampG::25922; Kp1-ΔampG::LT2.
Kp NTUH-K2044-ΔampG-RC knock-in strains: Kp NTUH-K2044-ΔampG-RC::Kp1; KpNTUH-K2044-ΔampG-RC::029M; Kp NTUH-K2044-ΔampG-RC::PAO1; Kp NTUH-K2044-ΔampG-RC::25922;Kp NTUH-K2044-ΔampG-RC:: LT2.
The mRNA Expressions of ampG and ampC Detected by RT-PCR
We carried out RNA extraction as per the instructions of the Shanghai Sangon columnar bacterial total RNA extraction and purification kit. The reverse transcription was done according to the instructions of the Shanghai Sangon M-MuLV first-strand cDNA synthesis kit and stored at −20°C for further use. The real-time PCR reaction system and conditions were readied as per the usage instructions of the SYBR Premix Ex Taq II reagent manufactured by TaKaRa Company of Japan, the primers used are listed in Table 3, and three trials were performed in parallel for each gene. We calculated the average value of the obtained Ct value with 16SrRNA as the internal reference. We used the following formula to calculate the relative expressions of ampC and ampG genes: ΔΔCT value and relative content of 2−ΔΔCt. ΔCt=Ct target gene-Ct16SrRNA. We compared Kp1 and its clones with Kp1-ΔampG, and Kp NTUH-K2044 and its clones with Kp NTUH-K2044-ΔampG-RC.
 |
Table 3 Primer Sequences Used in RT-PCR |
We used analysis of variance (ANOVA) to statistically analyze the relative expressions of ampG and ampC in each experimental strain of K. pneumoniae. P < 0.05 indicated a statistically significant difference.
ampC Induction Experiment
Using the disk diffusion method,9 the experimental strain was turned into a 0.5 M bacterial solution, which was evenly coated on M-H agar plates. Cefoxitin (FOX, 30 μg/tablet) was pasted as an in vitro inducer of ampC in the center of the plate, and Cefotaxime (CTX, 30 μg/tablet), Ceftazidime (CAZ, 30 μg/tablet), Cefepime (FEP, 30 μg/tablet), Piperacillin (PRL, 100 μg/tablet) and Cefoperazone/sulbactam (SCF, 105 μg/tablet) drug-sensitive paper sheets were pasted around it as indicators, with a distance of 20 mm between the center of each paper piece. The plate was incubated at 37°C for 18–20 hours. The result of the ampC induction experiment was deemed positive if any circle on the surrounding disk exhibited a truncation phenomenon on the side close to cefoxitin.
Minimal Inhibitory Concentration (MIC) Detection
The minimal inhibitory concentration (MIC) value was determined by the agar dilution method.10 M-H agar plates containing cefoxitin (FOX), ceftazidime (CAZ), cefuroxime (CXM), and cefazolin (KZ) were prepared, and the detection range of the drug MIC concentration gradient was 1–1024 μg/mL.
Results
Successful Construction of Kp1-ΔampG, Kp NTUH-K2044-ΔampG, Kp NTUH-K2044-RC and Kp NTUH-K2044-ΔampG-RC Strains
The ampG genes in the experimental strain Kp1 and the wild-type strain Kp NTUH-K2044 were successfully knocked out using homologous recombinant technology and the temperature-sensitive suicide vector pKO3-km, and we constructed the ampG gene deletion strains Kp1-ΔampG and Kp NTUH-K2044-ΔampG. The Kp NTUH-K2044-RC and Kp NTUH-K2044-ΔampG-RC clonal strains were obtained by recombinant plasmid pACYC184-ampCR.
Construction of ΔampG::ampG
Construction of Pet-30a-ampG
The genomes of five bacteria were used as templates, and the ampG gene of each strain was amplified by PCR with the primers listed in Table 2. We successfully obtained the ampG gene fragments of Kp1, S. typhimurium LT2, E. cloacae 029M, E. coli ATCC25922 strains with a length of 1476 bp and the ampG gene fragment of PAO1 with a length of 1785 bp. After double enzyme digestion of the PCR products of the ampG gene of the five bacteria, we derived the recombinant plasmid pet-30a-ampG by linking the products to plasmid pet-30a-ampG through T4DNA ligase. The plasmids with positive clones were identified using double enzyme digestion. The recombinant plasmid with ampG fragments of Kp1, E. cloacae 029M, E. coli ATCC25922 and S. typhimurium LT2 had a length of 6898 bp. After digestion by EcoRI and SacI, two clear bands appeared. One was a linear vector with a length of 5422 bp, and the other was a target fragment with a length of 1476 bp. The recombinant plasmid with PAO1 had a length of 7207 bp. As expected, two clear bands appeared after digestion, with one being 5422 bp and the other being 1785 bp. The pet-30a vector was successfully connected to the ampG gene fragment.
Identification of ΔampG::ampG Knock-in Strain
The recombinant plasmids pet-30a-ampG derived from five bacteria were electroporated into competent K. pneumoniae strains Kp1-ΔampG and Kp NTUH-K2044-ΔampG-RC. Positive monoclonal colonies cultured overnight were selected for plasmid extraction. Each recombinant plasmid was identified with the primers listed in Table 2. We found that 1785 bp ampG fragment could be obtained in the strain knocked in with ampG of PAO1, and 1476bp ampG fragment was found in the strain knocked in with ampG of E. coli ATCC25922, E. cloacae 029M, S. typhimurium LT2, and Kp1, which was in line with expectations.
RT-PCR Detection Results for the mRNAs of ampG and ampC
The mRNAs of ampG and ampC were found in each knock-in strain, but the degree of expression was slightly different.
Compared with wild-type Kp1, the expressions of ampC and ampG significantly increased in Kp1-ΔampG that knocked in with the ampG from itself (P < 0.05). In the strain knocked in with ampG of E. cloacae 029M, the expression of ampC increased (P < 0.05), and the level of ampG reached that in Kp1. In the strains knocked in with ampG of PAO1, E. coli ATCC25922, and S. typhimurium LT2, the expression of ampC was lower than that in Kp1 and the expression of ampG decreased as well (P < 0.05) (Figures 1A and B).
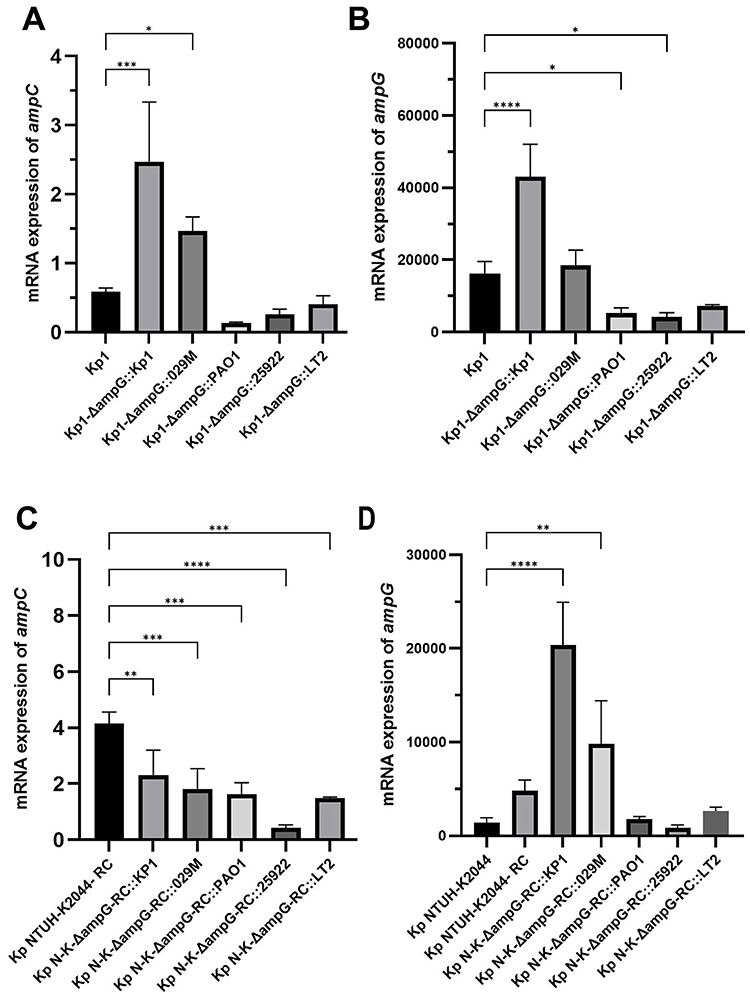 |
Figure 1 mRNA expression of experimental strains ampG and ampC. (A) mRNA expression of ampC in Kp1 and Kp1-ΔampG::ampG. (B) mRNA expression of ampG in Kp1 and Kp1-ΔampG::ampG. (C) mRNA expression of ampC in Kp NTUH-K2044 and Kp NTUH-K2044-ΔampG::ampG. (D) mRNA expression of ampG in Kp NTUH-K2044 and Kp NTUH-K2044-ΔampG::ampG. Notes: ****Represents the comparison with the wild-type strain, P<0.0001; *** represents the comparison with the wild-type strain, P<0.001; **Represents the comparison with the wild-type strain, P<0.01; * represents the comparison with the wild-type strain, P<0.05. |
Compared with Kp NTUH-K2044-RC, the expression of ampC was slightly reduced in Kp NTUH-K2044-ΔampG-RC knocked in with ampG of Kp1 and E. cloacae 029M, while the expression of ampG was significantly increased (P < 0.05). In the strains knocked in with ampG of PAO1, E. coli ATCC25922, and S. typhimurium LT2, the expression of ampG was also correspondingly reduced, and the expression of ampC decreased significantly (P < 0.05) (Figure 1C and D).
ampC Induction Experiment
The Kp1 strain is a DHA-1-producing K. pneumoniae with plasmid-mediated ampC expression, carrying the ampR-ampC gene. The drug susceptibility results showed that Kp1was resistant to cefoxitin, exhibiting an ampC-mediated resistance phenotype (Figure 2A). When the ampG gene was knocked out, Kp1-ΔampG showed sensitivity to cefoxitin, and the ampC-induced phenotype disappeared (Figure 2B). The Kp1-ΔampG, which knocked in with the ampG genes of Kp1 and E. cloacus 029M, showed drug resistance to cefoxitin, recovered induced phenotype, and truncation phenomenon (Figure 2C and D). In the strains knocked in with the ampG genes of P. aeruginosa PAO1, E. coli ATCC25922, and S. typhimurium LT2, the induced phenotype was not recovered, but they all showed drug resistance to cefoxitin (Figure 2E-G).
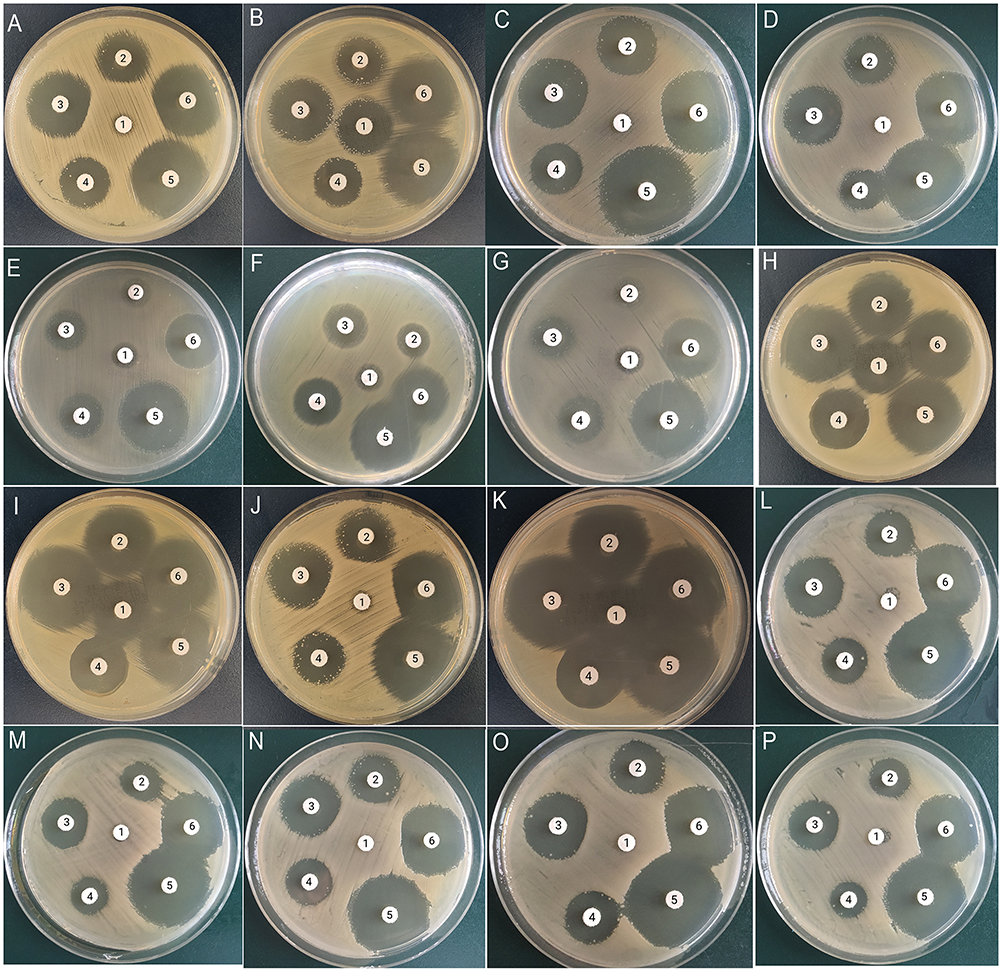 |
Figure 2 ampC induction experiment. 1: cefoxitin; 2: ceftazidime; 3: cefotaxime; 4: piperacillin; 5: cefepime; 6: cefoperazone/sulbactam; (A) Kp1; (B) Kp1-ΔampG strain; (C) Kp1-ΔampG::Kp1strain; (D) Kp1-ΔampG::029M strain; (E) Kp1-ΔampG::P AO1 strain; (F) Kp1-ΔampG::25922 strain (G) Kp1-ΔampG::LT2 strain; (H) Kp NTUH-K2044 strain; (I) Kp NTUH-K2044-ΔampG strain; (J) Kp NTUH-K2044-RC strain; (K) Kp NTUH-K2044-ΔampG-RC strain; (L) Kp NTUH-K2044-ΔampG-RC::Kp1strain; (M) Kp NTUH-K2044-ΔampG-RC::029M strain; (N) Kp NTUH-K2044-ΔampG-RC::PAO1 ampG strain; (O) Kp NTUH-K2044-ΔampG-RC::25922 strain; (P) Kp NTUH-K2044-ΔampG-RC::LT2. Notes: Using disk 1 as the inducer, if the induced phenotype exists in the experimental strain, there is a circle truncation phenomenon between disk 1 and the rest of the disks. |
The Kp NTUH-K2044 is a non-ampC-producing strain that does not have the ampR-ampC gene. It was sensitive to all the drugs that were tested, and it did not exhibit the ampC induction phenomenon (Figure 2H). Also, we did not observe induction phenomenon in the ampG-knockout Kp NTUH-K2044 strain (Figure 2I). When the exogenous plasmid pACYC184-ampRC was introduced into the Kp NTUH-K2044 strain, it showed drug resistance to cefoxitin, and the ampC induction experiment was positive (Figure 2J). When ampR-ampC was introduced into its ampG knockout strain, the strain regained sensitivity to cefoxitin, and the ampC induction experiment was negative (Figure 2K). In Kp NTUH-K2044-ΔampG-RC that knocked in with the ampG genes of Kp1, E. cloacae 029M, P. aeruginosa PAO1, E. coli ATCC25922, and S. typhimurium LT2, they all showed drug resistance to cefoxitin, recovered ampC-induced phenotypes, and truncation phenomena (Figure 2L-P).
MIC Values of Each Experimental Strain
Kp1was resistant to cefoxitin, ceftazidime, cefuroxime, and cefazoline. When the ampG gene was knocked out, Kp1-ΔampG became sensitive to the above antibacterial drugs, and the MIC value decreased, cefuroxime was reduced from 512 μg/mL to 4 μg/mL, cefazolin was reduced from 512 μg/mL to 2 μg/mL, and ceftazidime and cefoxitin were both reduced from 128 μg/mL to 1 μg/mL. When the ampG genes of the five strains were knocked in, all Kp1-ΔampG::ampG strains showed drug resistance and increased MIC values, resistance to all four antimicrobials returned to the Kp1 resistance level, with resistance to cefazolin increasing to 1024 μg/mL, twice the Kp1 resistance level.
Kp NTUH-K2044 and Kp NTUH-K2044-ΔampG strains were sensitive to cefoxitin, ceftazidime, cefuroxime, and cefazoline. When ampR-ampC was introduced, Kp NTUH-K2044-RC showed resistance to various antibacterial drugs with an increased MIC value, MIC values consistent with Kp1 resistance levels, while Kp NTUH-K2044-ΔampG-RC was sensitive with a decreased MIC value. The NTUH-K2044-ΔampG-RC::ampG strains that were knocked in with the ampG genes of five bacteria showed resistance to drugs, and the MIC value was increased, cefuroxime increased from 2 μg/mL to 512 μg/mL, cefazolin increased from 1 μg/mL to >1024 μg/mL, and ceftazidime and cefoxitin both increased from <1μg/mL to 128μg/mL (Table 4).
 |
Table 4 Phenotype Detection of AmpC in the Experimental Strain |
Discussion
Nosocomial infections are frequently caused by the gram-negative bacillus K. pneumoniae. The primary source of drug resistance in K. pneumoniae is its ability to produce ESBLs and AmpC. Drug resistance in gram-negative bacilli against new broad-spectrum β-lactam antibiotics is mostly attributable to AmpC β-lactamases, commonly known as cephalosporinase.11 AmpC expression was previously thought to be minimal and to be mediated only by the chromosome. However, plasmid-mediated ampC has increased, most often with sustained high expression, in recent years. Clinical anti-infection treatment has been complicated by the detection of K. pneumoniae strains with ampC-induced expression.12,13
AmpC encoded by structural gene ampC, and its expression is controlled by the amp complex operons ampC, ampR, ampD, ampG, and ampE.14 There is an activator, ampR, and an inhibitor, ampD. When no β-lactam antibiotic is present to function as an inducer, ampD blocks the production of ampR, resulting in low expression of ampC. In the presence of the inducer, ampR may no longer be inhibited by ampD, resulting in high production of ampC. This kind of expression is linked to the synthesis and degradation of peptidoglycan in the cell wall and requires the involvement of the ampR regulator. Furthermore, ampG plays a vital role as a transporter in the murein cycle.15 In earlier experiments, the AmpG topology of 20 strains of DHA-1-producing K. pneumoniae with plasmid-mediated ampC expression was predicted using the Expasy online software SOSUI, and it was determined that AmpG is a transmembrane protein with 12 transmembrane structures, with its N and C terminals exposed outside the membrane. However, more research and confirmation are required to establish the role of ampG in the control of plasmid-mediated ampC-induced expression.
Knocking out the target gene and investigating its role by comparing pre- and post-knockout strain phenotypic alterations is the most direct method for studying gene function. To test whether the AmpG regulator can control the plasmid-mediated ampC-induced expression in K. pneumoniae, we chose two different K. pneumoniae strains and knocked out the ampG gene. The ampRC gene was found on the plasmid of the DHA-1-producing clinical strain Kp1, which had been previously isolated in the laboratory. Professor Wang Jintang of the National Taiwan University generously provided the other strain, known as Kp NTUH-K2044. No ampRC gene was located on any of the chromosomes or plasmids in the genome of the strain, making it a perfect model strain for studies on the mechanism of ampC-mediated drug resistance in K. pneumoniae.
The research on gene knockouts now relies heavily on homologous recombination strategies based on effective suicide vector systems. In this study, we selected a temperature-sensitive suicide vector, pKO3-km. The first round of recombination occurred in culture at 43°C, and the second round of recombination occurred at 30°C. Finally, the positive clonal strains were screened. The positive strains were screened out by resistance, with continuous bacterial recombination induced in fresh medium. The purpose was clear, and the positive rate was high. The unique capsule structure of K. pneumoniae results in a low transformation efficiency of plasmids. To address this issue, we used clones in solid medium to directly prepare competent cells, which increased the transformation efficiency of K. pneumoniae. The gene knockout strains constructed with this method do not leave any traces, which lays a foundation for further double and multiple gene knockouts and the construction of knock-in strains.
The Kp1strain carries the ampC gene and has a positive ampC-induced phenotype. When its ampG gene is knocked out, ampC is not expressed, and the ampC-induced phenotype is negative. The MIC of β-lactam antibiotics is significantly reduced. (Table 4) It can be seen that the ampG gene plays an important role in the expression of ampC. This conclusion is consistent with the findings of Daniel et al in 2008 that the loss of ampG in N. gonorrhoeae caused a significant increase in the release of peptidoglycan fragments,16 and the reports of Schmidt et al, in which nitrosoguanidine was used for chemical-induced mutation of E. coli SN0301, so as to obtain an ampG mutant strain, which led to a loss of strain inductivity.17
The ampRC gene was absent in the Kp NTUH-K2044 wild-type strain, with no expression of ampC. The ampC induced phenotype was negative, and the strain was sensitive to cephalosporin antibiotics. When ampG gene was knocked out, the above indicators of the strain Kp NTUH-K2044-ΔampG did not change. The pACYC184-ampRC expression vector that was constructed earlier in the laboratory was introduced into KpNTUH-K2044-ΔampG. Although the recombinant Kp NTUH-K2044-ΔampG-RC obtained the exogenous ampRC gene, no ampC expression was found, the ampC induction experiment was negative, due to the knockout of ampG. Nevertheless, when the recombinant strain Kp NTUH-K2044-RC obtained the exogenous ampRC gene, the mRNA expression of ampC was 4.145 ± 0.415 (P < 0.05), and the ampC induction experiment was positive. Antibiotic resistance also increased significantly.(Table 4) It may be due to the high copy number of the pACYC184-ampRC plasmid and the resulting high expression of the ampRC gene, which significantly increases bacterial drug resistance. The above findings show that ampG is a necessary regulator for plasmid-mediated ampC-induced expression.
In order to further explore the function of ampG in ampC induced expression, we introduced the ampG genes of K. pneumoniae Kp1, E. cloacae 029M, P. aeruginosa PAO1, E. coli ATCC25922, and S. typhimurium LT2 into Kp1-ΔampG and Kp NTUH-K2044-ΔampG-RC strains, respectively. By detecting the differences in levels of ampC and ampG in each strain, the induced phenotype of ampC, and the change in MIC values, we analyzed the compensatory effect of ampG derived from different bacterial sources on ampG of K. pneumoniae. The expressions of ampC was observed in each knock-in strain of Kp1-ΔampG::ampG, and the expression level was slightly different compared with that of the Kp1 wild-type strain. The expression of ampC in Kp1-ΔampG::Kp1 increased most significantly (P < 0.05), when its own ampG gene was knocked into Kp1-ΔampG, the expression of ampC was significantly increased to 1.47 ± 0.203 in Kp1-ΔampG::029M (P < 0.05).
The corresponding ampC induction phenotype of the above two knock-in strains recovered, and these strains changed from being sensitive to being resistant to cephalosporins. The MIC values significantly increased, which returned to the resistance level of Kp1.
There was a statistically significant (P<0.05) reduction in ampG expression compared to Kp1 in the knock-in strains Kp1-ampG::P AO1, Kp1-ampG::25922, and Kp1-ampG::LT2. The induction of ampC failed to occur, whereas cefoxitin resistance was seen. After gene knock-in, previously sensitive bacteria become resistant to cephalosporin drugs. These results indicate that these strains produced ampC, but it was a low output. The yielded ampC was not enough to produce an induced phenotype. Therefore, it manifested as cefoxitin resistance.
We performed a Basic Local Alignment Search Tool (BLAST) comparison between the ampG gene in the above strains and the ampG gene in K. pneumoniae to further explore the reasons for the above induction phenomenon. We found that the ampG gene of K. pneumoniae had 84% homology with that of E. cloacae, 82% homology with S. typhimurium LT2, and no homology with that of E. coli ATCC25922 and P. aeruginosa PAO1. Although the ampG gene of S. typhimurium LT2 had 82% homology with that of K. pneumoniae, the expressions of ampC and ampG still did not reach the expression level of wild-type, and the induced phenotype was not restored.
The ampG genes of Kp1 and E. cloacae 029M could completely compensate for the deletion of the ampG gene in Kp1, while the ampG genes of E. coli ATCC25922, S. typhimurium LT2, and P. aeruginosa PAO1 could partially make up for the deficiency. We speculate that this discrepancy may be due to the difference between the ampG origins of K. pneumoniae and the above strains. Therefore, it only partially compensated for the ampG gene deletion background of K. pneumoniae, manifesting as resistance to cefoxitin without the corresponding induction phenotype. In 2010, Zhang et al confirmed that ampG was an essential gene for ampC induced expression in P. aeruginosa, and the ampG gene of E. coli only partially compensated for the production of ampC.18 In their study in 2012, using Xanthomonas str. 17, Yang et al reported that ampG was a key gene required for ampC-induced expression in Xanthomonas.19 The overexpression of ampG gene could lead to an increase in ampC expression, and the ampG gene of E. coli only partially compensated for its ampG deletion. Our findings are consistent with the above conclusions that the ampG gene of E. coli can only partially compensate for the ampG gene of K. pneumoniae.
When the ampG genes of five strains were knocked into the Kp NTUH-K2044-ΔampG-RC strain, RT-PCR showed ampC expression in each strain. Compared with Kp NTUH-K2044-RC, the ampC and ampG expression levels were significantly increased when the ampG of Kp1 and E. cloacae 029M were knocked in (P < 0.05). In contrast, when the ampG of P. aeruginosa PAO1, E. coli ATCC25922, and S. typhimurium LT2 were knocked in, the ampC expression decreased (P < 0.05) when compared with Kp NTUH-K2044-RC. The corresponding ampG expression was lower as well. However, despite the ampC expression of the above knock-in strains being lower than that of Kp NTUH-K2044-RC, it was sufficient to restore the ampC induced phenotype, and these strains showed resistance to cefoxitin. The MICs of cefuroxime and cefazolin increased to 512 ug/mL and > 1024 ug/mL, and the MICs of ceftazidime and cefoxitin both reached 128 ug/mL.
Since plasmid-mediated ampC has an inducible mechanism, certain powerful cephalosporin antibiotics regularly used in clinical practice frequently promote high expression of ampC, leading to the development of drug-resistant strains. Additional drug-resistance genes, such as extended-spectrum β-lactamases or drug-resistant integrators, may be expressed alongside ampC in clinical strains, making their genetic environment even more complicated.20
Conclusion
In summary, the role of ampG in plasmid-mediated ampC induced expression was validated by the knockout of the ampG gene in this experiment. The study of the ampG gene will help provide new insights for the development of novel gene therapies. In the future, knockout of the ampG gene in ampC-producing strains will result in drug sensitivity to antibiotics and undoubtedly become a new target for drugs.
Abbreviations
ATCC, American Type Culture Collection; CAZ, Ceftazidime; CLSI, Clinical and Laboratory Standards Institute; CXM, Cefuroxime; ESBLs, Extended-spectrum β-lactamases; FEP, Cefepime; FOX, Cefoxitin; I, Intermediate; Km, Kanamycin; Kp, Klebsiella pneumoniae; KZ, Cefazolin; MIC, minimum inhibitory concentration; PCR, polymerase chain reaction; R, resistant; RT-PCR, Real-time PCR; S, Susceptible; SCF, Cefoperazone/sulbactam; PIP, Piperacillin; CTX, Cefotaxime; E.cloa 029M, E.cloacae 029M; S.ty LT2, S.typhimurium LT2; E.coli 25922, E.coli ATCC25922; PA O1, P.aeruginosa O1.
Funding
National Natural Science Foundation of China (No. 30672005); Natural Science Foundation of Heilongjiang Province of China (No. H2013100).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Li GL, Duo LB, Luan Y, et al. Identification of genotypes of plasmid-encoded AmpC beta-lactamases from clinical isolates and characterization of mutations in their promoter and attenuator regions. Gene Expr. 2012;15(5–6):215–223. doi:10.3727/105221613x13571653093204. PMID: 23539899; PMCID: PMC6043836.
2. Luo DT, Duo LB. Research progress and regulation of AmpG in AmpC enzyme expression. Chine J Lab Diagnosis. 2012;16(1):176–179. doi:10.3969/j.issn.1007-4287.2012.01.077
3. Duo LB, Liu J, Yang DX, et al. Cloning and sequencing of the plasmid-mediated DHA-1 gene of AmpC β-lactamase in Klebsiella pnuemoniae. Chin J Microbiol Immunol. 2005;25(8):683–686.
4. Wang L, Duo LB. Application of gene knockout technique in gene function of Klebsiella pneumoniae. Chine J Clin Lab Sci. 2015;5:376–377. doi:10.13602/j.cnki.jcls.2015.05.15
5. Yacyshyn BR, Crooke ST. The concept and application of antisense oligonucleotides. Dis Colon Rectum. 2001;44(9):1241–1243. doi:10.1007/BF02234777. PMID: 11584192.
6. Lee SH, Sinko PJ. siRNA--getting the message out. Eur J Pharm Sci. 2006;27(5):401–410. doi:10.1016/j.ejps.2005.12.002. Epub 2006 Jan 25. PMID: 16442784.
7. Luan Y, Li GL, Duo LB, et al. DHA-1 plasmid-mediated AmpC β-lactamase expression and regulation of Klebsiella pnuemoniae isolates. Mol Med Rep. 2015;11(4):3069–3077 . doi:10.3892/mmr.2014.3054
8. Zheng Y, Hong XP, Liu JH. Highly efficient transformation with plasmid DNA in Klebsiella pneumoniae. Acta Microbiologica Sinica. 2007;47(4):721–724.
9. Clinical and Laboratory Standards Institute. Performance standards for antimicrobial disk susceptibility tests: Approved standard-eleventh edition. 2012; 32(1):M02–A11.
10. Clinical and Laboratory Standads Institute. Methods for dilution antimicrobial susceptibility tests for bacteria that grow Aerobically. Approved standard-Ninthedition. 2012;32(1):M07–A9.
11. Philippon A, Arlet G, Jacoby GA. Plasmid-mediated AmpC-type beta-lactamases. Antimierob Apents Chemother. 2002;46(1):1–11 . doi:10.1128/AAC.46.1.1-11.2002
12. Joanna E, Jaroslav H, Aleksandra K, et al. DHA-1-Producing Klebsiella pneumoniae in a teaching hospital in the Czech Republic. Microbial Drug Resistance. 2010;16(4):291–295. doi:10.1089/mdr.2010.0030
13. Gupta V, Kumarasamy K, Gulati N, et al. AmpC β-lactamases in nosocomial isolates of Klebsiella pneumoniae from India. Indian J Med Res. 2012;136(2):237–241
14. Tuomanen E, Lindquist S, Sande S, et al. Coordinate regulation of beta-lactamase induction and peptidogly can composition by the amp operon. Science. 1991;251(4990):201–204 . doi:10.1126/science
15. Chahboune A, Decaffmeyer M, Brasseur R, Joris B. Membrane topology of the Escherichia coli AmpG permease required for recycling of cell wall anhydromuropeptides and AmpC beta-lactamase induction. Antimicrob Agents Chemother. 2005;49(3):1145–1149. doi:10.1128/AAC.49.3.1145-1149.2005. PMID: 15728916; PMCID: PMC549251.
16. Garcia DL, Dillard JP. Mutations in ampG or ampD affect peptidoglycan fragment release from Neisseria gonorrhoeae. Journal of Bacteriology. 2008;190(11):3799–3807. doi:10.1128/JB.01194-07
17. Schmidt H, Korfmann G, Barth H, Martin HH. The signal transducer encoded by ampG is essential for induction of chromosomal AmpC beta-lactamase in Escherichia coli by beta-lactam antibiotics and ‘unspecific’ inducers. Microbiology. 1995;141(Pt 5):1085–1092. doi:10.1099/13500872-141-5-1085. PMID: 7773404.
18. Zhang Y, Bao Q, Gagnon LA, et al. ampG gene of Pseudomonas aeruginosa and its role in β-lactamase expression. Antimicrob Agents Chemother. 2010;54(11):4772–4779. doi:10.1128/AAC.00009-10. Epub 2010 Aug 16. PMID: 20713660; PMCID: PMC2976151.
19. Yang TC, Chen TF, Tsai JJ, Hu RM. AmpG is required for BlaXc beta-lactamase expression in Xanthomonas campestris pv. campestris str. 17. FEMS Microbiol Lett. 2013;340(2):101–108. doi:10.1111/1574-6968.12071. Epub 2013 Jan 31. PMID: 23278458.
20. Rowe-Magnus DA, Guerout AM, Mazel D. Bacterial resistance evolution by recruitment of super-integron gene cassettes. Mol Microbiol. 2002;43(6):1657–1669. doi:10.1046/j.1365-2958.2002.02861.x. PMID: 11952913.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.