")
Back to Journals » OncoTargets and Therapy » Volume 13
Silencing of Long Noncoding RNA LINC00346 Inhibits the Tumorigenesis of Colorectal Cancer Through Targeting MicroRNA-148b
Authors Li T, Wang B, Zhang L, Cui M, Sun B
Received 17 December 2019
Accepted for publication 19 March 2020
Published 17 April 2020 Volume 2020:13 Pages 3247—3257
DOI https://doi.org/10.2147/OTT.S242715
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Nicola Silvestris
Tao Li,1 Benjun Wang,2 Lianxiang Zhang,3 Meng Cui,1 Bing Sun1
1Department of General Surgery, The First Affiliated Hospital of Shandong First Medical University, Jinan City, Shandong Province, 250014, People’s Republic of China; 2Department of Anorectal, Shandong Provincial Hospital of Traditional Chinese Medicine, Jinan City, Shandong Province 250014, People’s Republic of China; 3Department of Anorectal, Huantai County Hospital of Traditional Chinese Medicine, Huantai County, Shandong Province 256400, People’s Republic of China
Correspondence: Bing Sun
Department of General Surgery, The First Affiliated Hospital of Shandong First Medical University, No. 16766, Jingshi Road, Jinan City, Shandong Province 250014, People’s Republic of China
Tel +86- 531- 89268503
Email [email protected]
Purpose: This study was aimed to explore the regulatory effect of long noncoding RNA LINC00346 (LINC00346) on colorectal cancer (CRC) and the potential molecular mechanisms.
Methods: The expression of LINC00346 and microRNA-148b (miR-148b) in CRC tissues and cells was detected by qRT-PCR. LINC00346 was overexpressed and silenced in HT29 and HCT116 cells by the transfection of pcDNA-LINC00346 and si-LINC00346, respectively. The cell proliferation, migration, invasion, and apoptosis were analyzed by cell counting kit-8 (CCK-8), wound-healing, transwell, and flow cytometry assay, respectively. The targeting relationship between LINC00346 and miR-148b was predicted by TargetScan and determined by dual-luciferase reporter assay. A tumor xenograft model was established in mice to evaluate the tumor growth in vivo.
Results: The expression of LINC00346 was up-regulated in CRC tissues and cells. The expression of LINC00346 was positively associated with the TNM stage, lymphoma metastasis and histological grade. Overexpression of LINC00346 promoted the proliferation, migration and invasion and inhibited the apoptosis of HT29 and HCT116 cells. MiR-148b was a target of LINC00346. Silencing of miR-148b reversed the anti-tumor effect of si-LINC00346 on CRC cells. Furthermore, silencing of LINC00346 inhibited the tumor growth in mice through up-regulating miR-148b.
Conclusion: Silencing of LINC00346 inhibited the proliferation, migration and invasion, and promoted the apoptosis of CRC cells through targeting miR-148b.
Keywords: LINC00346, colorectal cancer, proliferation, apoptosis, miR-148b
Introduction
Colorectal cancer (CRC) is a common cancer that originates in the colon.1 A recent study has shown that the annual incidence rate of CRC is increasing worldwide.2 Currently, traditional Chinese medicines, chemotherapy, surgery, targeted therapy, and radiotherapy are used for the treatment of CRC.3 CRC can be cured in 95% of cases if detected in the early stage.4 Exploring the potential targets for the diagnosis and treatment of CRC is urgently needed.
Long non-coding RNAs (lncRNAs) are non-coding RNA transcripts that involved in the pathogenesis of many cancers, such as bladder cancer, pancreatic cancer and breast cancer.5–7 LncRNA LINC00346 (LINC00346) belongs to the intragenic lncRNAs is located on the chromosome 13q34 with a total length of 6322 bp.8 Recently, some studies have proved that LINC00346 is involved in cancer progression. Ye et al have demonstrated that silencing of LINC00346 attenuates the proliferation and migration, and promotes the apoptosis of bladder cancer cells.9 Shi et al have suggested that LINC00346 shows the ability to promote the growth of pancreatic cancer.10 Liu et al have reported that the LINC00346 promotes the proliferation of breast cancer cells.11 However, the specific regulatory role of LINC00346 on the occurrence and progress of CRC remains unclear.
MicroRNAs (miRNAs) are a class of small noncoding RNAs of about 22 nucleotides, which are involved in the regulation of cell proliferation and apoptosis.12,13 MiR-148b is a specific miRNAs located on chromosome 12q13.14 Song et al have shown that miR-148b attenuates the tumor growth of CRC through regulating cholecystokinin-2 receptor.14 Wang et al have proved that miR-148b inhibits the cell cycle progression in CRC.15 Researchers have indicated that lncRNAs act as molecular sponges of miRNAs to affect the expression of target mRNA and ultimately influence the development of CRC.5,16 However, the specific regulatory relationship between LINC00346 and miR-148b in CRC remains undefined.
Here, we investigated the effects of LINC00346 on the proliferation, migration, invasion and apoptosis of CRC cells and on the tumor growth in mice. The regulatory relationship between LINC00346 and miR-148b was further evaluated. Our research may discover a hopeful therapeutic target for CRC, and reveal the underlying mechanisms for CRC treatment.
Patients and Methods
Tissue Samples
In total, 60 CRC patients (32 males and 28 females, 53.46 ± 10.50 years old) were collected from our hospital between January 2017 and December 2018. CRC tissues and adjacent non-cancerous tissues were collected by surgical resection. This study was approved by the ethical committee of Huantai County Hospital of Traditional Chinese Medicine, and was conducted in accordance with the principles of the Declaration of Helsinki. All patients signed informed consent.
Cell Culture
Five human CRC cell lines (HT29, HCT8, LoVo, SW480 and HCT116) and human normal colonic cell line FHC were obtained from the American Type Culture Collection (ATCC). Cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM, Invitrogen, Carlsbad, CA, USA) containing 10% fetal bovine serum (FBS, Invitrogen) at 37°C containing 5% CO2. Cells were randomly divided into three groups and used in three independent experiments. All experiments were repeated three times.
Cell Transfection
The pcDNA3.1 LINC00346 (pcDNA-LINC00346), pcDNA3.1 negative control (pcDNA-NC), LINC00346 siRNA (si-LINC00346), siRNA negative control (si-NC), miR-148b mimic, miR-148b negative control (miR-NC) and miR-148b inhibitor were purchased from Ruibo (Shanghai, China). HT29 and HCT116 cells grown to 80% confluence were transfected with these above agents using Lipofectamine 3000 reagent (Invitrogen). The HT29 and HCT 116 cells were divided into 4 groups: pcDNA-NC group, pcDNA-LINC00346 group, si-NC group and si-LINC00346 group. In addition, the HCT116 cells were further divided into 4 groups: si-NC + miR-NC group, si-NC + miR-148b inhibitor group, si-LINC00346 + miR-NC group and si-LINC00346 + miR-148b inhibitor group. Cells only transfected with Lipofectamine 3000 without the above agents were considered as the BLANK (BLANK group).
Quantitative Real Time Polymerase Chain Reaction (qRT-PCR)
The total RNA was extracted from cells and tissues by using TRIZOL regent (Invitrogen), and was reverse-transcribed into cDNA by using Prime Script RT reagent kit (Takara, Dalian, China). qRT-PCR was performed by using SYBR Green PCR kit (Takara). Amplification conditions were as follows: 95°C for 10 min, 40 cycles of 95°C for 10 s, 60°C for 20 s, and 72°C for 34 s. Relative expression of genes was calculated by the 2−ΔΔCt method. GAPDH was used for the normalization of LINC00346. U6 was used for the normalization of miR-148b. The primer sequences (Bioengineering in Shanghai, China) were shown in Table 1.
 |
Table 1 Primer Sequences Used in Quantitative Real Time Polymerase Chain Reaction (qRT-PCR) |
Cell Counting Kit-8 (CCK-8) Assay
Cells were seeded into 96-well plates (2 × 104 cells/well) and cultured for 24, 48, 72 and 96 h, respectively. Then 10 μL CCK-8 solution (BD Biosciences, San Jose, CA, USA) was added into each well. After incubated for 2 h at 37°C, the absorbance at 450 nm was measured using a microplate reader (Bio-Tek Instruments, Winooski, VT, USA).
Flow Cytometry
Cells were suspended and adjusted to 1 × 106 cells/mL. Then 500 µL cells were stained with 5 μL Annexin V-fluorescein isothiocyanate (V-FITC) and 10 μL propidium iodide (PI) for 20 min in the dark. Cell apoptosis were analyzed by a FACScan flow cytometer (BD Biosciences).
Wound Healing Assay
When cells were cultured at 90% confluence, an artificial scratch was created using a 10 μL pipette tip. Cells were then incubated for 48 h and observed under an inverted microscope (Olympus Co., Tokyo, Japan). Wound healing rate was calculated according to the fraction of cell coverage across the line.
Transwell Assay
The transwell assay was used to determine the cell invasion using transwell chambers (8 nm pore size, Corning Inc., Corning, NY, USA). Cells (2 × 105) in serum free medium were added to upper chambers pre-coated with matrigel (BD Biosciences). DMEM containing 10% FBS (Invitrogen) was added to the lower chambers. After 48 h of incubation at 37°C, cells were removed from the upper chambers with a cotton swab. Cells in lower chambers were fixed in methanol and stained with 0.5% crystal violet for 2 min. The stained cells were photographed and counted at five randomly selected fields.
Dual-Luciferase Reporter Assay
The potential binding site between miR-148b and LINC00346 was predicted by TargetScan. The LINC00346-Wt and LINC00346-Mut were cloned and combined with psiCHECK-2 vector (Promega, Madison, WI, USA). LINC00346-Wt or LINC00346-Mut was co-transfected with miR-148b mimics or miR-NC (Genepharma, Shanghai) into HT29 and HCT116 cells with Lipofectamine 3000 (Invitrogen). After 48 h of transfection, the luciferase activity was detected by dual-luciferase reporter gene assay system (Promega).
RNA-Pull Down Assay
Bio-LINC00346-Wt, Bio-LINC00346-Mut and Bio-NC (GenePharma) were transfected into CRC cells (HT29 and HCT116) using Lipofectamine 3000 (Invitrogen). After cultured for 48 h, cells were incubated with Dynabeads M-280 Streptavidin beads (Invitrogen) for 1 h. The RNAs were detected by qRT-PCR.
Tumor Formation in Mice
Thirty male BALB/c nude mice (30 ± 3.4 g) were obtained from Huafukang (Beijing, China). To construct tumor xenograft model, HCT116 cells (1 × 106) co-transfected with si-NC + miR-NC (si-NC + miR-NC group), si-NC + miR-148b inhibitor (si-NC + miR-148b inhibitor group), si-LINC00346 + miR-NC (si-LINC00346 + miR-NC group) or si-LINC00346 + miR-148b inhibitor (si-LINC00346 + miR-148b inhibitor group) (n = 6, per group) were subcutaneously injected into mice. Mice in the Blank group (n = 6) were subcutaneously injected with HCT116 cells without transfection. The tumor volume was measured every week with calipers, and calculated as (L⋅W2)/2 (L, the largest diameter in millimeters; W, the smallest diameter in millimeters). After the last measurement (the 4th week), mice were anesthetized with 50 mg/kg pentobarbital sodium and sacrificed by neck-lifting method. The tumor was isolated and weighed. All animal experiments were approved by the Animal Care and Use Committee of our hospital, and were performed in accordance with the Guide for the Care and Use of Laboratory Animals (eighth edition, 2011, National Institutes of Health, USA).
Statistical Analysis
Statistical analysis was performed using SPSS 23.0 (SPSS Inc., Chicago, IL, USA) and GraphPad Prism software 7.0. Data were presented as mean ± standard deviation (SD). The differences among multi-groups were analyzed by one-way ANOVA followed by the multiple comparisons test. The differences between two groups were assessed by Student’s t-test. A P value < 0.05 was considered statistically significant.
Results
The Expression of LINC00346 Is Up-Regulated in CRC Tissues and Cells
The expression of LINC00346 in CRC tissues was significantly higher than that in adjacent normal tissues (P < 0.001) (Figure 1A). The expression of LINC00346 in CRC cell lines (HT29, HCT8, LoVo, SW480 and HCT116) was significantly higher than that in normal human colonic cell line FHC (P < 0.05) (Figure 1B). ROC curve showed that LINC00346 exhibited a high diagnostic value on CRC [area under the curve (AUC), 0.9586; confidence intervals (CIs), 0.9248–0.9924; sensitivity, 86.67%; specificity, 96.67%; P < 0.05] (Figure 1C). The relationship between the expression of LINC00346 and the clinical characteristics of CRC patients was analyzed. The expression of LINC00346 in tumors at TNM stage I–II was significantly lower than that in tumors at TNM stage III–IV (P < 0.01) (Figure 1D). In addition, the expression of LINC00346 was significantly higher in poorly differentiated tumors than that in well and moderate differentiated tumors, and was significantly in higher in tumors with lymphoma metastasis than that in tumors without lymphoma metastasis (P < 0.001) (Figure 1E and F) (Table 2).
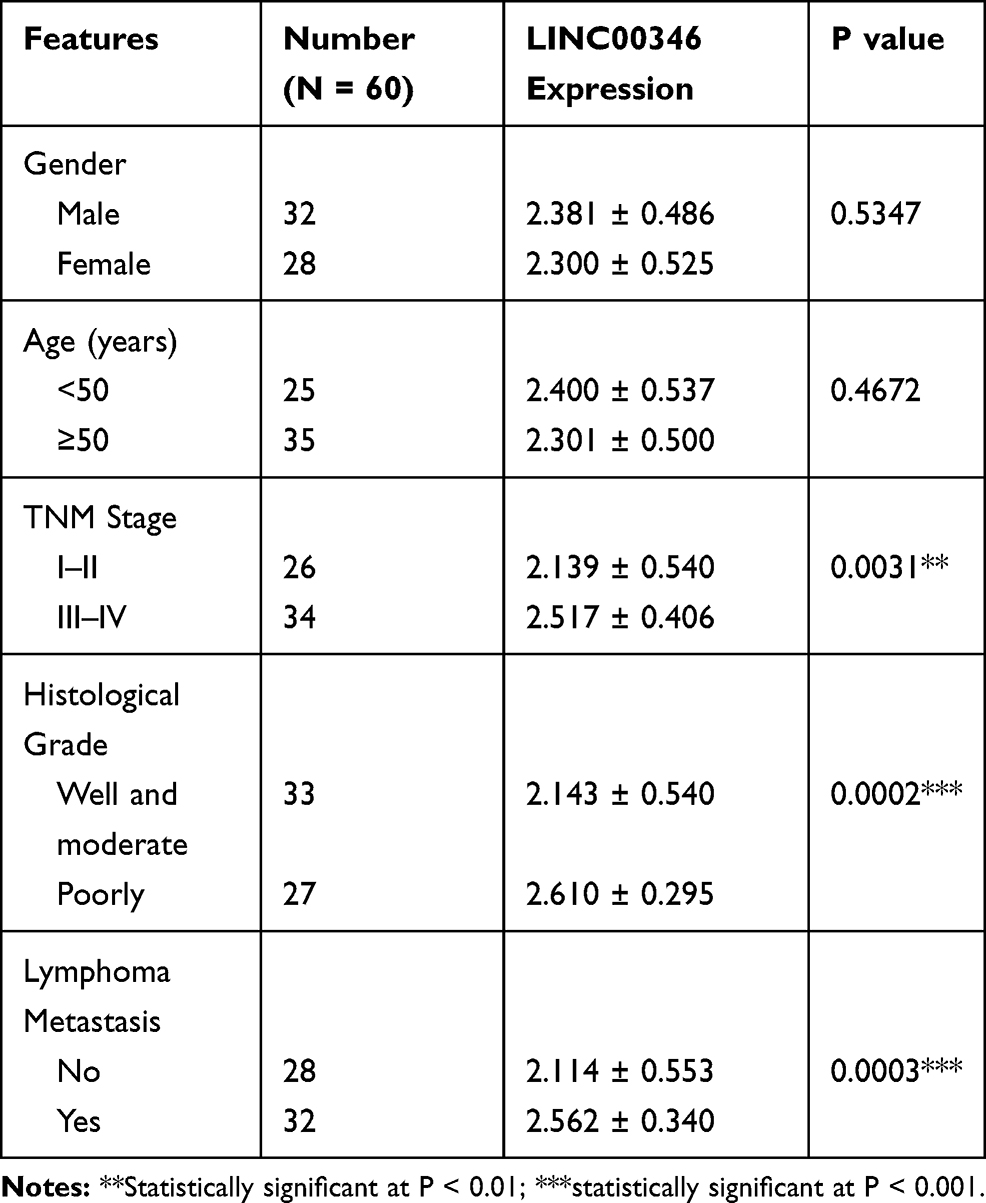 |
Table 2 The Relationship Between the Expression of LINC00346 and Clinical Characteristics in CRC Patients |
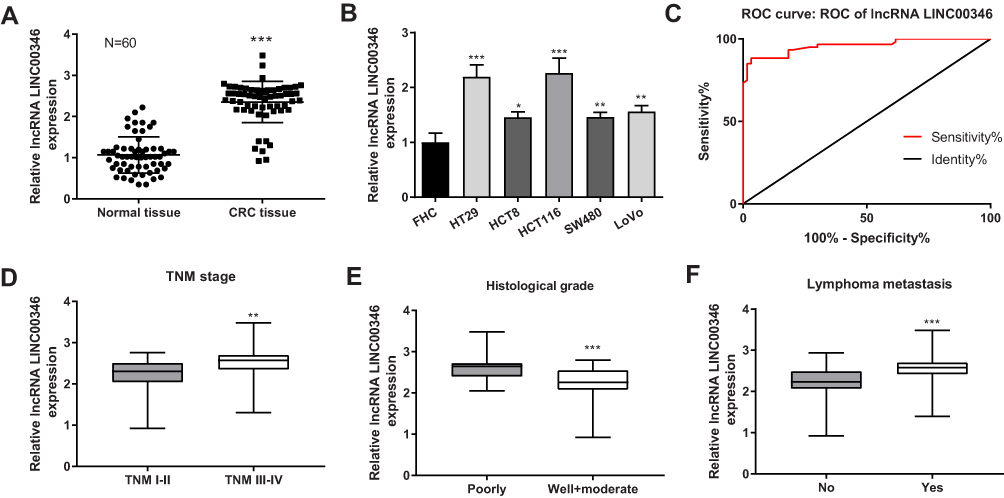 |
Figure 1 The expression of LINC00346 in CRC tissues and cells. (A) The expression of LINC00346 in normal tissues and CRC tissues was detected by qRT-PCR (N = 60). ***P < 0.001 vs Normal tissue; (B) the expression of LINC00346 in human normal colonic cell line FHC and CRC cell lines (HT29, HCT8, LoVo, SW480 and HCT116) was detected by qRT-PCR (N = 3). *P < 0.05, **P < 0.01, ***P < 0.001 vs FHC; (C) the diagnostic value of LINC00346 on CRC was analyzed by a ROC curve; (D) the expression of LINC00346 in CRC tissues at different TNM stages (I–II, N = 26; III–IV, N = 34). **P < 0.01 vs TNM I–II; (E) the expression of LINC00346 in CRC tissues at different histological grades (Well and moderate, N = 33; Poorly, N = 27). ***P < 0.001 vs Poorly; (F) the expression of LINC00346 in CRC tissues with or without lymphoma metastasis (No, N = 28; Yes, N = 32). ***P < 0.001 vs No. N represented biological replicates. All experiments were repeated three times. |
Overexpression of LINC00346 Promotes the Proliferation and Inhibits the Apoptosis of CRC Cells
To evaluate the regulatory effects of LINC00346 on the proliferation and apoptosis of CRC cells, LINC00346 was overexpressed and silenced in CRC cells (HT29 and HCT116 cells) by the transfection of pcDNA-LINC00346 and si-LINC00346, respectively. qRT-PCR showed that the expression of LINC00346 was significantly up-regulated in the pcDNA-LINC00346 group and was significantly down-regulated in the si-LINC00346 group compared with the BLANK group (P < 0.001) (Figure 2A). CCK-8 assay showed that overexpression and silencing of LINC00346 significantly increased and decreased the OD450 values at 48, 72, and 96 h post-culturing, respectively (P < 0.05) (Figure 2B and C). Flow cytometry showed that overexpression and silencing of LINC00346 significantly inhibited and promoted the apoptosis of HT29 and HCT116 cells, respectively (P < 0.01) (Figure 2D).
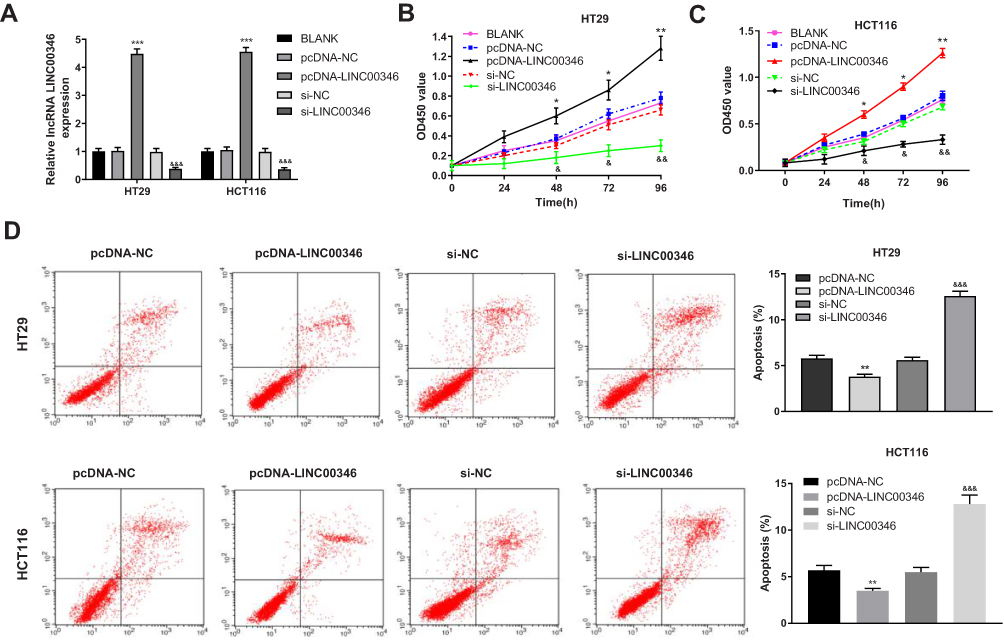 |
Figure 2 Overexpression of LINC00346 promoted the proliferation and inhibited the apoptosis of CRC cells. (A) The expression of LINC00346 in HT29 and HCT116 cells was detected by qRT-PCR (N = 3); (B and C) the OD450 values of HT29 and HCT116 cells were detected by CCK-8 assay (N = 3); (D) the apoptosis of HT29 and HCT116 cells was detected by flow cytometry (N = 3). *P < 0.05, **P < 0.01, ***P < 0.001 vs BLANK or pcDNA-NC; &P < 0.05, &&P < 0.01, &&&P < 0.001 vs BLANK or si-NC. N represented biological replicates. All experiments were repeated three times. |
Overexpression of LINC00346 Enhances the Migration and Invasion of CRC Cells
Wound healing and transwell assay were used to detect the migration and invasion of CRC cells. The relative migrated rates of HT29 and HCT116 cells were significantly increased in the pcDNA-LINC00346 group compared with the pcDNA-NC group (P < 0.01) (Figure 3A). The relative number of invasive cells in the pcDNA-LINC00346 group was significantly higher than that in the pcDNA-NC group (P < 0.001) (Figure 3B). The transfection of si-LINC00346 significantly decreased the relative migrated rate and relative number of invasive cells (P < 0.01) (Figure 3A and B).
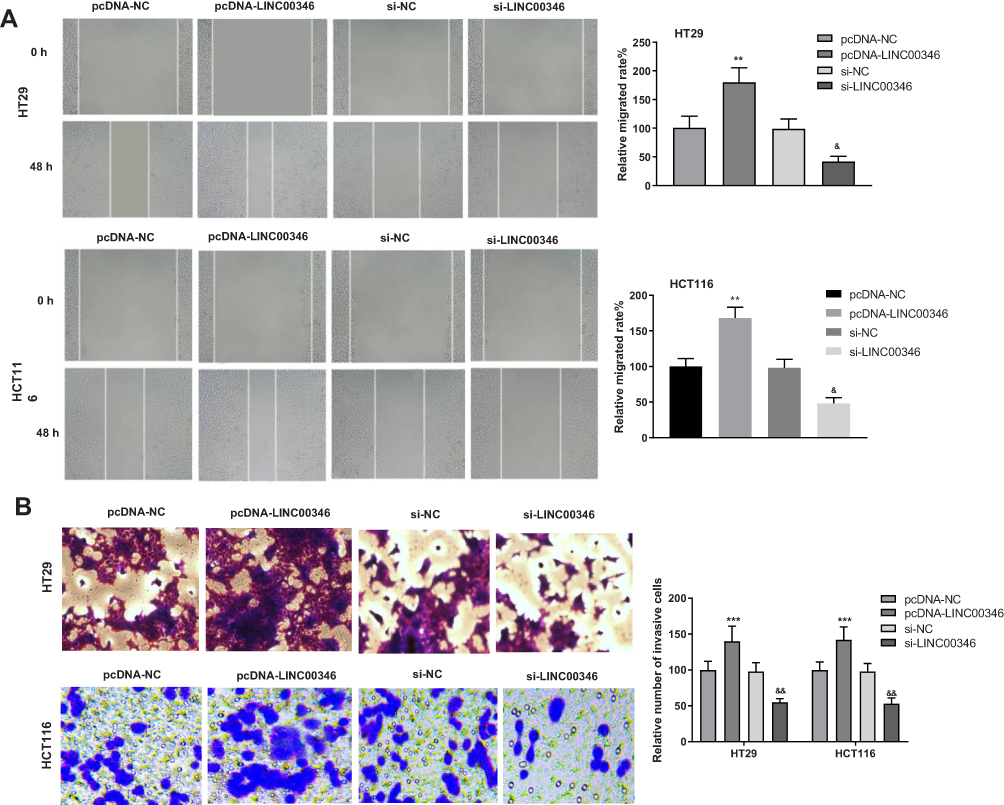 |
Figure 3 Overexpression of LINC00346 enhanced the migration and invasion of CRC cells. (A) The relative migrated rates of HT29 and HCT116 cells were detected by wound healing assay (N = 3); (B) relative numbers of invasive cells of HT29 and HCT116 cells were detected by transwell assay (N = 3). ***P < 0.001, **P < 0.01 vs pcDNA-NC; &&P < 0.01, &P < 0.05 vs si-NC. N represented biological replicates. All experiments were repeated three times. |
MiR-148b Is a Target of LINC00346 in CRC Cells
The expression of miR-148b in CRC tissues and cells was detected by qRT-PCR. As showed in Figure 4A and B, the expression of miR-148b was significantly lower in CRC tissues than that in adjacent normal tissues (P < 0.001), and was significantly lower in HT29 and HCT116 than that in FHC cells (P < 0.05). There was a negative correlation between the expression of LINC00346 and miR-148b in CRC tissues (N = 60, r = −0.3139, P = 0.0146) (Figure 4C). TargetScan predicted a binding site of miR-148b on LINC00346 (Figure 4D). Dual-luciferase reporter assay showed that the relative luciferase activity of cells co-transfected with miR-148b mimics and LINC00346-Wt was significantly decreased compared with those co-transfected with miR-NC and LINC00346-Wt (P < 0.001) (Figure 4E). RNA-pull down assay showed that the expression of miR-148b in the Bio-LINC00346-Wt group was significantly increased compared with the Bio-LINC00346-Mut group (P < 0.001) (Figure 4F). Furthermore, the expression of miR-148b was significantly decreased by the transfection of pcDNA-LINC00346 and was significantly increased by the transfection of si-LINC00346 (P < 0.01) (Figure 4G).
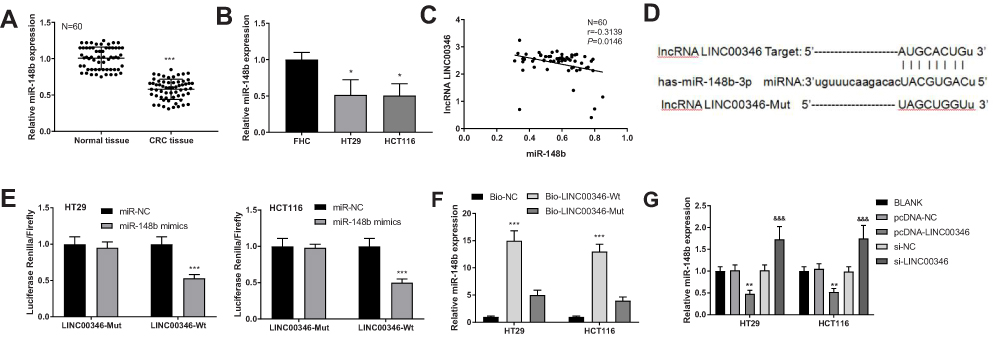 |
Figure 4 MiR-148b was a target of LINC00346 in CRC. (A) The expression of miR-148b in normal tissues and CRC tissues was detected by qRT-PCR (N = 60). ***P < 0.001 vs Normal tissue; (B) the expression of miR-148b in FHC, HT29 and HCT116 cells was detected by qRT-PCR (N = 3). *P < 0.05 vs FHC; (C) the correlation between the expression of LINC00346 and miR-148b in CRC tissues (N = 60); (D) the target site of miR-148b on LINC00346 was predicated by TargetScan; (E) relative luciferase activity of HT29 and HCT116 cells was measured by dual-luciferase reporter assay (N = 3). ***P < 0.001 vs miR-NC; (F) the interaction between miR-148b and LINC00346 in HT29 and HCT116 cells was assessed by RNA pull down assay (N = 3). ***P < 0.001 vs Bio-NC; (G) the expression of miR-148b in HT29 and HCT116 cells was detected by qRT-PCR (N = 3). **P < 0.01 vs pcDNA-NC or BLANK; &&&P < 0.001 vs si-NC and BLANK. N represented biological replicates. All experiments were repeated three times. |
Silencing of miR-148b Eliminates the Anti-Tumor Role of Si-LINC00346 in CRC Cells
To verify the regulatory effect of miR-148b on CRC cells, miR-148b was silenced in CRC cells (HT29 and HCT116) by the transfection of miR-148b inhibitor. The expression of miR-148b in the miR-148b inhibitor group was significantly decreased compared with the BLANK group (P < 0.001) (Figure 5A). CCK-8 assay revealed that the OD450 value of HCT116 cells was significantly decreased in the si-LINC00346 + miR-NC group and was significantly increased in the si-NC + miR-148b inhibitor group compared with the si-NC + miR-NC group at 48, 72, and 96 h post-culturing (P < 0.05). The transfection of miR-148b inhibitor significantly reversed the inhibiting effect of si-LINC00346 on the proliferation of HCT116 cells (P < 0.05) (Figure 5B). The results of cell apoptosis were opposite to the OD450 value in HCT116 cells (P < 0.05) (Figure 5C). In addition, the relative migrated rate and relative number of invasive cells were significantly decreased in the si-LINC00346 + miR-NC group and were significantly increased in the si-NC + miR-148b inhibitor group compared with the si-NC + miR-NC group (P < 0.01). The transfection of miR-148b inhibitor significantly reversed the inhibiting effects of si-LINC00346 on the migration and invasion of HCT116 cells (P < 0.05) (Figure 5D and E).
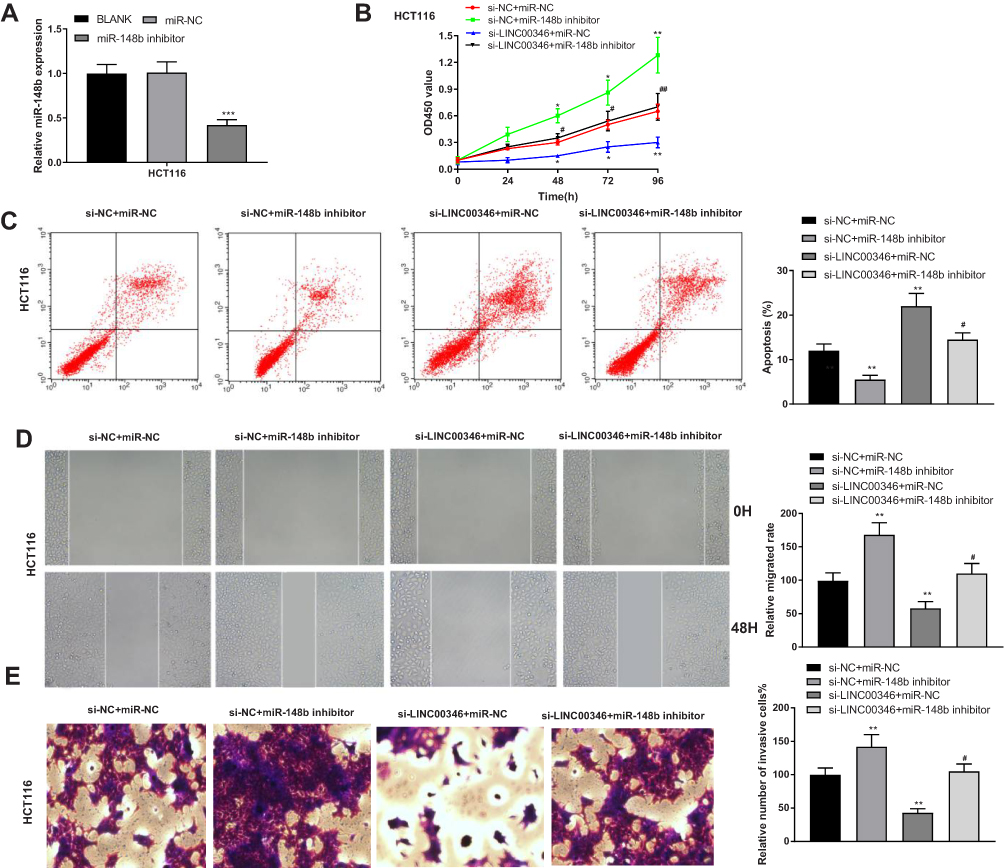 |
Figure 5 Silencing of miR-148b eliminated the anti-tumor effect of si-LINC00346 on CRC cells. (A) The expression of miR-148b in HCT116 cells was detected by qRT-PCR (N = 3); (B) the OD450 value of HCT116 cells was detected by CCK-8 assay (N = 3); (C) the apoptosis of HCT116 cells was detected by flow cytometry (N = 3). (D) The relative migrated rate in HCT116 cells was detected by wound healing assay (N = 3). (E) The relative number of invasive cells inHCT116 cells was detected by transwell assay (N = 3). *P < 0.05, **P < 0.01, ***P < 0.001 vs BLANK, si-NC+miR-NC or miR-NC; #P < 0.05, ##P < 0.01 vs si-LINC00346 + miR-NC. N represented biological replicates. All experiments were repeated three times. |
Silencing of LINC00346 Inhibits the Tumor Growth in Mice
To further verify the anti-tumor effect of si-LINC00346 in vivo, HCT116 cells were subcutaneously injected into mice. As shown in Figure 6A and B, the tumor volume and weight were significantly increased in the si-NC + miR-148b inhibitor group and were significantly decreased in the si-LINC00346 + miR-NC group compared with the si-NC + miR-NC or BLANK group (P < 0.01). The transfection of miR-148b inhibitor significantly reversed the inhibiting effect of si-LINC00346 on the tumor growth in mice (P < 0.05). In addition, the expression of LINC00346 was significantly increased in the si-NC + miR-148b inhibitor group and was significantly decreased in the si-LINC00346 + miR-NC group compared with the si-NC + miR-NC or BLANK group (P < 0.05). The expression changes of miR-148b were contrary to those of LINC00346 (P < 0.01) (Figure 6C and D).
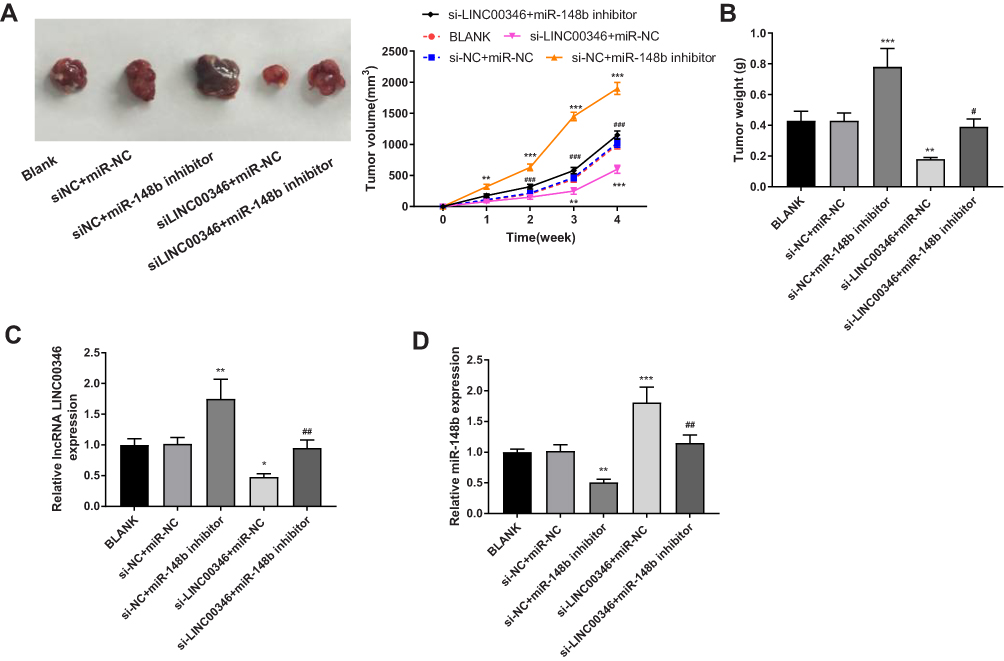 |
Figure 6 Silencing of LINC00346 inhibited the tumor growth in mice. (A) The tumor volume was measured every week (N = 6); (B) the tumor weight was measured at the 4th week post-injection (N = 6). (C) The expression of LINC00346 in tumor tissues was detected by qRT-PCR (N = 6); (D) the expression of miR-148b in tumor tissues was detected by qRT-PCR (N = 6). *P < 0.05, **P < 0.01, ***P < 0.001 vs BLANK or si-NC + miR-NC; #P < 0.05, ##P < 0.01, ###P < 0.001 vs miR-NC + si-LINC00346. N represented biological replicates. All experiments were repeated three times. |
Discussion
CRC has a high mortality and poor prognosis all over the world.17,18 Recently, some lncRNAs have been identified as potential biomarkers for the prognosis of CRC.19 For instances, overexpression of lncRNA HOTTPI is a biomarker for the unfavorable prognosis of CRC.20 High expression of lncRNA HOTTIP serves as a valuable biomarker for CRC.21 In this study, the expression of LINC00346 was up-regulated in CRC tissues and cells. These results indicate that LINC00346 may be a tumor promoter in CRC. In addition, we found that overexpression of LINC00346 was positively associated with the TNM stage, lymphoma metastasis and histological grade in patients with CRC. Yi et al have shown that overexpression of LncRNA FTX was positively related to the TNM stage, lymph node metastasis and tumor diameter in patients with CRC.22 The predictive value of LINC00346 was similar to that of LncRNA FTX in CRC. LINC00346 may be a valuable diagnostic and prognostic factor for CRC.
LINC00346 is involved in the tumorigenesis of different types of cancers by regulating cell proliferation, migration, invasion and apoptosis. Ye et al have found that knockdown of LINC00346 inhibits the proliferation and migration, and induces the cell cycle arrest and apoptosis in bladder cancer cells.9 Wang et al have proved that silencing of LINC00346 in non-small cell lung cancer cells promotes cell apoptosis, inhibits cell proliferation, and arrests cell cycle in G1-G0 phase.23 Shi et al have shown that knockdown of LINC00346 suppresses cell proliferation and causes a cell-cycle arrest at the G2/M-phase in pancreatic cancer cells.24 These findings indicate that LINC00346 is a tumor promoter in bladder cancer, non-small cell lung cancer, and pancreatic cancer. In this study, silencing of LINC00346 significantly inhibits the proliferation, migration, and invasion, and promotes the apoptosis of CRC cells. Our results are consistent with previous studies, and further illustrate that LINC00346 acts as a tumor promoter in CRC. LINC00346 may be a promising therapeutic target for CRC. In order to verify the anti-tumor effect of LINC00346 silencing in vivo, a tumor xenograft model was established in mice. We found that silencing of LINC00346 significantly inhibits the tumor growth in mice. This result further illustrates that silencing of LINC00346 can inhibit the tumorigenesis of CRC in vivo.
LncRNAs act as competing endogenous RNAs or as sponges of miRNAs to regulate the expression of target mRNAs. Many regulatory interactions between lncRNAs and miRNAs have been revealed in CRC by previous studies, such as lncRNA DANCR and miR-577,5 lncRNA PART1 and miR-143,25 lncRNA CRNDE and miR-181a-5p,26 and lncRNA DORAD and miR-202-5P.27 In this study, miR-148b was confirmed as a target of LINC00346. MiR-148b is a tumor inhibitor that down-regulated in hepatocellular carcinoma,28 pancreatic cancer,29 gastric cancer,30 and CRC.31 Song et al have shown that overexpression of miR-148b in CRC cells inhibits cell proliferation in vitro and suppresses tumorigenicity in vivo.31 Wang et al have proved that overexpression of miR-148b blocks cell cycle progression and inhibits cell proliferation in CRC cells.15 Here, a negative regulatory relationship between LINC00346 and miR-148b was revealed in CRC cells. We speculate that the tumor promoting role of LINC00346 may be closely associated with the interaction with miR-148p. This speculation was further verified by the following feedback experiments. We found that silencing of miR-148b eliminates the anti-tumor effect of si-LINC00346 on CRC cells. In addition, silencing of miR-148b also reversed the inhibiting effect of si-LINC00346 on the tumor growth in mice. These results illustrate that silencing of LINC00346 can inhibit the tumorigenesis of CRC through up-regulating miR-148b both in vitro and in vivo.
This study exhibits some limitations. First, the association between the expression of LINC00346 and the prognosis of CRC patients was not evaluated. Second, the diagnostic value of LINC00346 on CRC needs to be verified in a large population. Third, the target genes of miR-148b and LINC00346 were not analyzed. Further researches on these fields are still needed.
Conclusions
In conclusion, the expression of LINC00346 was up-regulated in CRC tissues and cells. MiR-148b was a target of LINC00346. Silencing of LINC00346 inhibits the proliferation, migration and invasion, and promotes the apoptosis of CRC cells through up-regulating miR-148b. Silencing of LINC00346 also inhibited the tumor growth in mice through up-regulating miR-148b. LINC00346 may be a promising therapeutic target for CRC.
Ethics and Consent Statement
This study was conducted after obtaining local ethical committee approval of The First Affiliated Hospital of Shandong First Medical University. All patients signed informed consent, and this was conducted in accordance with the Declaration of Helsinki.
All animal experiments were approved by the Animal Care and Use Committee of The First Affiliated Hospital of Shandong First Medical University, and were performed in accordance with the Guide for the Care and Use of Laboratory Animals (eighth edition, 2011, National Institutes of Health, USA).
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
There is no funding in the research.
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Brenner H, Kloor M, Pox CP. Colorectal cancer. Lancet. 1490-1502;383(9927).
2. Blum〣arnett E, Madrid S, Burnett〩artman A, Mueller SR, Feigelson HS. Financial burden and quality of life among early﹐nset colorectal cancer survivors: a qualitative analysis. Health Expectations. 2019;22(5):1050–1057.
3. Mody K, Baldeo C, Bekaii-Saab T. Antiangiogenic therapy in colorectal cancer. Cancer J. 2018;24(4):165–170. doi:10.1097/PPO.0000000000000328
4. Pawa N, Arulampalam T, Norton JD. Screening for colorectal cancer: established and emerging modalities. Nat Rev Gastroenterol Hepatol. 2011;8(12):711–722. doi:10.1038/nrgastro.2011.205
5. Wang Y, Lu Z, Wang N, et al. Long noncoding RNA DANCR promotes colorectal cancer proliferation and metastasis via miR-577 sponging. Exp Mol Med. 2018;50(5):57.
6. Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):0–641.
7. Huarte M. The emerging role of lncRNAs in cancer. Nature Med. 2015;21(11):1253.
8. Wang F, Chen JG, Wang LL, Yan ZZ, Wang XG. Up-regulation of LINC00346 inhibits proliferation of non-small cell lung cancer cells through mediating JAK-STAT3 signaling pathway. Eur Rev Med Pharmacol Sci. 2017;21(22):5135–5142.
9. Ye T, Ding W, Wang N, Huang H, Pan Y, Wei A. Long noncoding RNA linc00346 promotes the malignant phenotypes of bladder cancer. Biochem Biophys Res Commun. 2017;491(1):79–84. doi:10.1016/j.bbrc.2017.07.045
10. Shi W, Zhang C, Ning Z, et al. Long non-coding RNA LINC00346 promotes pancreatic cancer growth and gemcitabine resistance by sponging miR-188-3p to derepress BRD4 expression. J Exp Clin Cancer Res. 2019;38(1):60.
11. Liu H, Juan L, Pratirodh K, et al. Long non-coding RNAs as prognostic markers in human breast cancer. Oncotarget. 2016;7(15):20584.
12. Mach CM, Kim J, Soibam B, et al. Novel MicroRNAs regulating proliferation and apoptosis in uterine papillary serous carcinomas. Cancer Lett. 2013;335(2):314–322. doi:10.1016/j.canlet.2013.02.043
13. Harapan H, Andalas M. The role of microRNAs in the proliferation, differentiation, invasion, and apoptosis of trophoblasts during the occurrence of preeclampsia—a systematic review. Tzu Chi Med J. 2015;27(2):54–64. doi:10.1016/j.tcmj.2015.05.001
14. Song Y, Xu Y, Wang Z, et al. MicroRNA-148b suppresses cell growth by targeting cholecystokinin-2 receptor in colorectal cancer. Int J Cancer. 2012;131(5):1042–1051.
15. Wang G, Cao X, Lai S, et al. Altered p53 regulation of miR-148b and p55PIK contributes to tumor progression in colorectal cancer. Oncogene. 2015;34(7):912–921. doi:10.1038/onc.2014.30
16. Anna SC, Yumi K, Yusuke Y, Fumitaka T, Takahiro O. Emerging roles of long non-coding RNA in cancer. Cancer Sci. 2018;109(7):2093–2100.
17. Guinney J, Dienstmann R, Wang X, et al. The consensus molecular subtypes of colorectal cancer. Nature Med. 2015;21(11):1350–1356. doi:10.1038/nm.3967
18. Miller KD, Siegel RL, Lin CC, et al. Cancer treatment and survivorship statistics, 2016. Cancer J Clin. 2016;66(4):271–289. doi:10.3322/caac.21349
19. Smolle M, Uranitsch S, Gerger A, Pichler M, Haybaeck J. Current status of long non-coding RNAs in human cancer with specific focus on colorectal cancer. Int J Mol Sci. 2014;15(8):13993–14013. doi:10.3390/ijms150813993
20. Ren YK, Xiao Y, Wan XB, Zhao YZ, Wang YC. Association of long non-coding RNA HOTTIP with progression and prognosis in colorectal cancer. Int J Clin Exp Pathol. 2015;8(9):11458–11463.
21. Niu H, Hu Z, Liu H, et al. Long non-coding RNA AK027294 involves in the process of proliferation, migration, and apoptosis of colorectal cancer cells. Tumor Biol. 2016;37(8):10097–10105. doi:10.1007/s13277-015-4350-x
22. Yi Y, Jinpei Z, Xi C, et al. LncRNA FTX sponges miR-215 and inhibits phosphorylation of vimentin for promoting colorectal cancer progression. Gene Ther. 2018;25(5):321–330.
23. Wang F, Chen JG, Wang LL, Yan ZZ, Chen SP, Wang XG. Up-regulation of LINC00346 inhibits proliferation of non-small cell lung cancer cells through mediating JAK-STAT3 signaling pathway. Eur Rev Med Pharmacol Sci. 2017;21(22):5135–5142. doi:10.26355/eurrev_201711_13830
24. Shi W, Zhang C, Ning Z, et al. Long non-coding RNA LINC00346 promotes pancreatic cancer growth and gemcitabine resistance by sponging miR-188-3p to derepress BRD4 expression. J Exp Clin Cancer Res. 2019;38(1):60. doi:10.1186/s13046-019-1055-9
25. Hu Y, Ma Z, He Y, Liu W, Su Y, Tang Z. PART-1 functions as a competitive endogenous RNA for promoting tumor progression by sponging miR-143 in colorectal cancer. Biochem Biophys Res Commun. 2017;490(2):317–323. doi:10.1016/j.bbrc.2017.06.042
26. Han P, Li JW, Zhang BM, et al. The lncRNA CRNDE promotes colorectal cancer cell proliferation and chemoresistance via miR-181a-5p-mediated regulation of Wnt/beta-catenin signaling. Mol Cancer. 2017;16(1):9. doi:10.1186/s12943-017-0583-1
27. Zhang J, Li X-Y, Hu P, Ding Y-S. LncRNA NORAD contributes to colorectal cancer progression by inhibition of miR-202-5p. Oncol Res Featuring Preclinical Clin Cancer Ther. 2018;26(9):1411–1418.
28. Sadeghian Y, Kamyabi-Moghaddam Z, Nodushan SMT, Khoshbakht S, Mohebbi M. Profiles of tissue microRNAs; miR-148b and miR-25 serve as potential prognostic biomarkers for hepatocellular carcinoma. Tumor Biol. 2015;37(12):16379–16380. doi:10.1007/s13277-015-3799-y
29. Zhao G, Zhang JG, Liu Y, et al. miR-148b functions as a tumor suppressor in pancreatic cancer by;targeting AMPK alpha 1. Mol Cancer Ther. 2013;12(1):83–93.
30. Li X, Jiang M, Chen D, et al. miR-148b-3p inhibits gastric cancer metastasis by inhibiting the Dock6/Rac1/Cdc42 axis. J Exp Clin Cancer Res. 2018;37(1):71. doi:10.1186/s13046-018-0729-z
31. Song Y, Xu Y, Wang Z, et al. MicroRNA-148b suppresses cell growth by targeting cholecystokinin-2 receptor in colorectal cancer. Int J Cancer. 2012;131(5):1042–1051. doi:10.1002/ijc.26485
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.