Back to Journals » Cancer Management and Research » Volume 13
Silencing of Long Non-Coding RNA HOTAIR Alleviates Epithelial–Mesenchymal Transition in Pancreatic Cancer via the Wnt/β-Catenin Signaling Pathway
Authors Tang Y, Song G, Liu H, Yang S, Yu X, Shi L
Received 1 June 2020
Accepted for publication 18 December 2020
Published 14 April 2021 Volume 2021:13 Pages 3247—3257
DOI https://doi.org/10.2147/CMAR.S265578
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Eileen O'Reilly
Yinhua Tang, Guang Song, Hongcheng Liu, Shuang Yang, Xiaoyi Yu, Lijun Shi
Department of Gastroenterology, The First Affiliated Hospital of Harbin Medical University, Harbin, 150001, People’s Republic of China
Correspondence: Lijun Shi
Department of Gastroenterology, The First Affiliated Hospital of Harbin Medical University, No. 23, Youzheng Street, Harbin, Heilongjiang Province, 150001, People’s Republic of China
Tel +86-0451-85555119
Email [email protected]
Purpose: Pancreatic cancer (PC) is a malignancy with poor prognosis and controversial treatment options. Long non-coding RNA (lncRNA) is a significant factor in the development of PC. In the current study, the possible effects of HOTAIR on the epithelial–mesenchymal transition (EMT) of PC and the related mechanisms were investigated.
Methods: The PC models were induced by 10 mg/100 g dimethylbenzoanthracene (DMBA) in pancreas. Mice were injected with the HOTAIR mimic and HOTAIR shRNA to determine the role of HOTAIR in PC. Subsequently, the expression of HOTAIR in PC cells was assayed. To determine the mechanism of HOTAIR in PC, human PC cell line PANC-1, Miapaca-2 and human normal pancreatic ductal epithelial cell line HPDE6-C7 were transfected with the HOTAIR mimic, the shRNA against HOTAIR, the Wnt/b-catenin activator (LiCl), and the Wnt/b-catenin inhibitor (XAV939), respectively. Moreover, the expressions of the Wnt/β-catenin signaling pathway-related genes (β-catenin, cyclinD1, c-myc, LEF-1 and c-Jun) and the levels of the EMT markers (E-cadherin, N-cadherin and Vimentin) were determined. Finally, the cell biological processes were evaluated by functional experiments.
Results: HOTAIR was found to be highly expressed in the PC cells in mice. The expression of β-catenin, cyclinD1, c-myc, LEF-1 and c-Jun, N-cadherin and Vimentin was found to be decreased, while the expression of E-cadherin was found to be increased subsequent to the silencing of HOTAIR in human PC cell lines PANC-1 and Miapaca-2. Additionally, it was observed that the silencing of HOTAIR could inhibit the Wnt/β-catenin signaling pathway to alleviate EMT of tumor cells and inhibit the capacities of cell proliferation, migration, and invasion.
Conclusion: The key finding of the present study is that the silencing of HOTAIR could potentially inhibit EMT and growth of PC through the Wnt/β-catenin signaling pathway, providing a novel therapy for PC.
Keywords: long non-coding RNA, HOTAIR, Wnt/β-catenin signaling pathway, epithelial–mesenchymal transition, pancreatic cancer
Introduction
Pancreatic cancer (PC) is fatal disease with the reported survival rate of less than 5% in the last 5 years.1 PC patients usually suffer from abdominal and back pain, fatigue, weight loss, cachexia and cholestasis.2 Additionally, obesity is reported to be a significant contributing factor to the development of PC.3 The extensive widespread metastasis is also a significant factor contributing to the poor survival rates of PC.4 Additionally, adjuvant chemotherapy but not chemoradiotherapy was considered to be the only effective treatment in PC.5 The survival of PC patients is low as most PC patients are diagnosed in the advanced stages.6 Considering the poor prognosis and limited treatment of PC, it is imperative to determine novel treatment strategies for PC.
Furthermore, recent studies have displayed that long non-coding RNA (lncRNAs) are enrolled in many biological process and cancers.7 HOTAIR, transcribed from the homeobox C gene locus in chromosome 12, was found to play a significant role in chromatin-modifying enzymes and gene silencing.8 Additionally, HOTAIR has also been established to play an important role in prediction of tumor prognosis and enhancement of oncogenic activity.9 Furthermore, another previously conducted study showed HOTIAR silencing also played an important role in attenuating tumor growth in PC.10 The Wnt/β-catenin signaling pathway was established to be the most conserved intercellular signaling cascade that involved in tumor progression.11 It functioned in regulating cell growth, development and differentiation of normal stem cells.12 Moreover, it has been revealed that Epithelial-to-mesenchymal transition (EMT) produced invasive, migratory cells, therefore involved in cell metastasis.13 It has been found that imidazothiadiazoles exhibits potential antiproliferative function, which is mediated through EMT.14 Additionally, another recent study suggested that EMT promotes the escape of PC cells from the primary tumor site, thereby promoting the cell metastasis in PC,15 and that the Wnt/β-catenin signaling pathway silencing also had close correlation with EMT attenuation in PC,16 while its activation enhanced tumorigenicity of PC.17 Furthermore, the Wnt/β-catenin signaling pathway has been established to contribute to the drug resistance in resistance cancer stem cells.18 On the basis of the abovementioned findings the current study speculated that there may be mechanism of HOTIAR in EMT in PC through the mediation of the Wnt/β-catenin signaling pathway, and it might provide theoretical basis for novel PC treatment. The present study aimed to confirm the effects of HOTAIR on EMT of PC and the related mechanisms.
Materials and Methods
Ethics Statement
The study was approved by the ethics committee of The First Affiliated Hospital of Harbin Medical University and carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals. Extensive efforts were made to reduce suffering of all included animals.
Model Establishment
Thirty clean C57BL/6 mice weighted 22–25 g, were obtained from the Shanghai laboratory animal center of Chinese Academy of Sciences, Shanghai, China and were fed under standard feeding conditions. Subsequently, 24 mice were enrolled for the establishment of the PC model and the remaining 6 mice were taken for sham operation. Following 24 h starvation prior to the operation, 2% pentobarbital sodium was injected intraperitoneally into the mice as anesthesia (1.5 mL/kg). A 1.5 cm abdominal incision was subsequently made on all the selected mice prepared for modeling. Subsequent to the full exposure of the pancreas, the capsula and some pancreatic parenchyma were carefully cut from the mouse body and the tail of pancreas (about 2 mm in depth), which were then placed in 10 μL (100 mg/mL) 7.12-dimethylbenzanthracene (DMBA, Sigma, St Louis, MO, USA). Following this, the capsula pancreatitis and the abdominal wall were sutured, and the mice were incubated in normal conditions.19 The capsula pancreatitis, parenchyma incision and the capsula pancreatitis suture were conducted on the 6 sham operated mice. The 24 PC mice were subsequently grouped into the blank control, negative control (NC), HOTAIR overexpression, and shRNA against HOTAIR, with 6 mice in each group. The spirit, diet and activity of the mice were observed, and mice were subsequently euthanized for autopsy at the fifth month following the operation. If any of the mice died naturally during the experiment, autopsy was performed immediately.
Plasmid Transfection
The pc-HOTAIR overexpressed plasmid, sh-HOTAIR plasmid and empty vector were constructed based on the HOTAIR sequence (Gene ID: 83286748). Subsequently, 300 μg plasmid and 600 μg stearylamine (SA) liposomes were mixed and dissolved in 1 mL normal saline, then placed at room temperature for the duration of 30 min. The SA liposome plasmid complex was then injected into the abdominal cavities of the mice.
Hematoxylin-Eosin (HE) Staining
The pancreatic tissues were fixed in 10% formalin (Sigma, St Louis, MO, USA) for the duration of 16 −18 h, and then paraffin-embedded and sliced. Subsequently, the resultant sections were stained with HE and the pathological changes were observed under an optical microscope.
Cell Culture
The human PC cell lines PANC-1, Miapaca-2 and human normal pancreatic ductal epithelial cell line, (Cell bank of Chinese Academy of Sciences, Shanghai, China) were incubated in the Dulbecco’s Modified Eagle Medium (DMEM) (Gibco, Grand Island, NY, USA) which contained 100 U/mL penicillin, 100 U/mL streptomycin and 10% fetal bovine serum (FBS, Gibco, Grand Island, NY, USA) in 5% CO2 at the temperature of 37°C. The subculture was conducted every 2–3 days when the cell confluence reached 70–90%. The cells were then treated with or without the pc-HOTAIR overexpression plasmid, the sh-HOTAIR, the empty vector, 10 mM lithium chloride (Lic1, Wnt/β-catenin activator, Sigma, St Louis, MO, USA) or 1 μM XAV939 (Wnt/β-catenin activator Sigma, St Louis, MO, USA). The Lipofectamine TM 2000 transfection reagent (Invitrogen, Carlsbad, CA, USA) was employed for plasmid transfection.
RNA Isolation and Quantitation
The total RNA in the PC tissues was extracted using the TRIzol reagent (Invitrogen, Carlsbad, California, USA) and 1 μg total RNA was employed for reverse transcription. The reverse transcriptase Moloney Murine Leukemia Virus (M-MLV), ribonuclease (RNAase) inhibitor RNasin and deoxyribonucleoside triphosphates (dNTP) were purchased from Promega (Promega, Madison, WI, USA). Additionally, the Applied Biosystems Step Two Real-time PCRT system (Applied Biosystems Inc., Foster City, CA, USA) was used for the RT-qPCR analysis (SYBR-Green were purchased from Sigma, St Louis, MO, USA). Furthermore, glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was considered as the internal reference. The primer sequences for the RT-qPCR analysis are shown in Table 1. Moreover, the Master Mix (Toyobo, Toyobo Co. Ltd., Japan) was employed for the detection and semi-quantitation of the expression of the target genes. The relative expression of the target genes was calculated by the 2−ΔΔCt formula.
 |
Table 1 Primer Sequences of RT-qPCR |
Western Blot Analysis
The Trizol reagent (Invitrogen, Carlsbad, California, USA) was used to extract the total protein. Subsequently, 1% to 5% sample was collected and transferred onto a polyvinylidene fluoride (PVDF) membrane following a 10% sodium dodecyl sulfate-polyacrylamide-polyacrylamide gel electrophoresis (SDS-PAGE). Following this, the membrane was sealed using the Tris-buffered Saline with Tween 20 (TBST) which contained 5% bovine serum albumin (BSA). Consequently, primary antibodies E-cadherin (1:100, ab76055), N-cadherin (1:500, ab98952), Vimentin (1:100, ab8978) and GAPDH (1:500, ab8245) (Abcam Inc., Cambridge, MA, USA) were added to the membrane at the controlled temperature of 4°C overnight. The unbound primary antibodies were then washed off, following which the secondary antibody (Shanghai Beyotime Biotechnology Co. Ltd., Shanghai, China) marked by horseradish peroxidase was added. Subsequent to incubation, the substrate was added for coloration. The results were expressed by the ratio of the gray level of the target gene band to GAPDH band.
Cell Counting Kit-8 (CCK-8) Assay
Cells from different treatment groups were placed in each well of the 96-well plate (100 μL, 3.0 × 103 cells) and cultured for the durations of 24, 48 and 72 h, respectively. At each time point, 10 μL CCK-8 (Solarbio, China) was added to each well and incubated in the incubator for an additional 2 h. The absorbance of each well was measured at the wavelength of 450 nm to evaluate the cell proliferation.
Transwell Assay
The Transwell assay was used to detect the invasion and metastasis capacity of the PC cells subsequent to the overexpression and knockdown of HOATIR. For cell invasion experiment, the cells were starved for 4 h, digested with trypsin, washed twice with PBS, and then resuspended in serum-free medium. The cell density was adjusted to 1 × 105 cells/mL. The basement membrane of the upper chamber was pre-coated with Matrigel (Corning Corporation, USA), and then 200 mL of the cell suspension was added, following which 500 mL of medium containing 20% serum was added to the lower chamber. The cells were subsequently cultured for the duration of 24 h. After that, the cells in the inner layer of the microporous membrane were carefully wiped with a cotton swab. Furthermore, the cells were fixed with pre-cooled 4% paraformaldehyde for the duration of 30 min. Subsequent to staining with 0.1% crystal violet solution, the stained cells were counted under an inverted microscope and photographed. The mean values of 5 visual fields were randomly selected to analyze the differences among groups and construct a histogram. The experiment was repeated 3 times to obtain variable data for further analysis. The matrigel coating was not required in the upper chamber of the Transwell chamber to detect cell migration ability, and the other operations were identical to that of the cell invasion experiments.
Statistical Analysis
The statistical analyses were conducted employing the SPSS 21.0 (IBM Corp. Armonk, NY, USA). Measurement data were expressed as the mean ± standard deviation. The comparisons amongst multiple groups were assessed using the one-way analysis of variance (ANOVA) followed by the Tukey’s post hoc test. Additionally, the cell activity at different time points was analyzed using the two-way ANOVA. p < 0.05 was statistically significant.
Results
PC Model Establishment
The successful modeling rate was 75% for the PC models. The mice were euthanized subsequent to the PC model induced by DMBA for 5 months, and HE staining was conducted to observe the histomorphological changes of the pancreas (Figure 1). The size of the PC tissues in mice treated with overexpression HOTAIR plasmid was found to be significantly larger in comparison to the other groups, while that in the mice treated with knockdown HOTAIR plasmid was smaller than that in other groups. The pathological examination of the mice models of PC was identified as pancreatic ductal adenocarcinoma. Furthermore, it was revealed that the tumor cells were adenoid, the size of glandular cavity and PC cells was different while the cytoplasm was eosinophilic. Additionally, the nucleus was located in the basal part of the cell with deeper staining, and the nuclear isomer was obvious (Figure 1). The aforementioned results indicated that PC mouse model was successfully established by the DMBA induction, and that the silencing of HOTAIR suppressed the proliferation of the tumor.
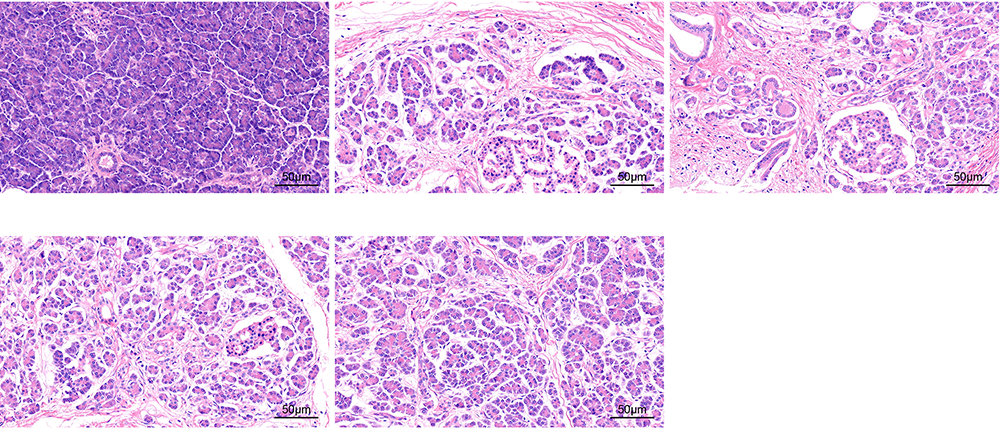 |
Figure 1 The PC model establishment was successful as assessed by HE staining. |
HOTAIR Overexpression Promotes Epithelial–Mesenchymal Transition (EMT) in PC
The gene expression of E-cadherin, N-cadherin, Vimentin was detected using the Western blot analysis. The results (Figure 2) revealed that in comparison to the normal cells and the cells treated with NC plasmids, the expression of E-cadherin was decreased and the expressions of N-cadherin and Vimentin were increased in the cells treated with HOTAIR (p < 0.05). Additionally, the expression of E-cadherin was found to be increased and the expressions of N-cadherin and Vimentin were decreased in the cells treated with shRNA against HOTAIR (p < 0.05). Furthermore, no significant changes were noted in the normal cells and the cells treated with NC plasmids (p > 0.05). Therefore, it was concluded that the overexpression of HOTAIR could promote EMT in PC, while HOTAIR silencing attenuated EMT in PC.
 |
Figure 2 Overexpressed HOTAIR contributed to the EMT in PC tissues. (A) protein bands of E-cadherin, N-cadherin, Vimentin in each group by Western blot analysis; (B) protein levels of E-cadherin, N-cadherin, Vimentin in each group as normalized to GAPDH. *p < 0.05 vs the cells without treatment and cells treated with empty vector. |
HOTAIR Upregulates the Wnt/β-Catenin Signaling Pathway in PC
The Wnt/β-catenin signaling pathway-related gene expression was detected employing the RT-qPCR analysis to determine the role of HOTAIR in PC progression in mice. The results (Figure 3) revealed that, in comparison to the cells without treatment and cells treated with the empty vector, HOTAIR overexpression amplified the expression of β-catenin, cyclinD1, c-myc, LEF-1 and c-Jun, while it was the opposite when HOTAIR was knocked down (p < 0.05). Whereas, there was no significant change noted in the blank and the NC groups (p > 0.05). Cumulatively, the abovementioned results indicated that HOTAIR up-regulated the Wnt/β-catenin signaling pathway-related gene expression.
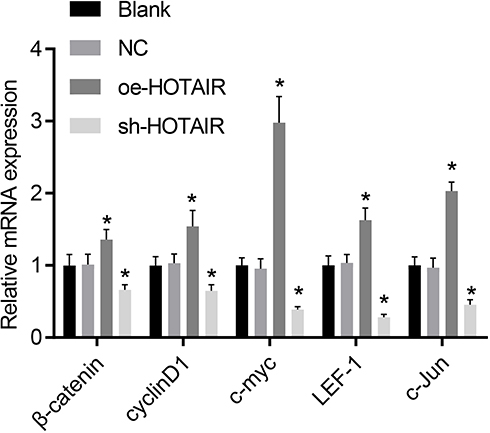 |
Figure 3 The expression of β-catenin, cyclinD1, c-myc, LEF-1 and c-Jun was enhanced by overexpressed HOTAIR as detected by RT-qPCR. *p < 0.05 vs cells without treatment and cells treated with empty vector. |
HOTAIR is Up-Regulated in PC Cells and Tumor Tissues
The HOTAIR expression in PC cell lines and normal pancreatic epithelial cells was confirmed using the RT-qPCR analysis. The results (Figure 4A) established that the expression of HOTAIR was higher in the Miapaca-2 and PANC-1 cell lines in comparison to the cells in normal pancreatic epithelial (p < 0.05). Furthermore, in the PC mouse model, the expression of HOTAIR in the blank control group, the NC group and the overexpression group was found to be significantly higher than that in the sham operation group (Figure 4B). Cumulatively, the present study concluded that HOTAIR was highly expressed in PC cells.
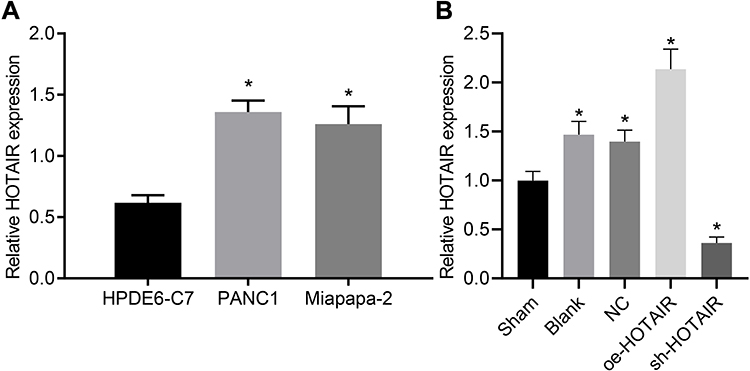 |
Figure 4 HOTAIR was highly expressed in PC cell lines and tissues. (A) HOTAIR was highly expressed in PC cells. (B) HOTAIR was highly expressed in PC mice tissues. In panel A, *p < 0.05 vs cells treated with HPDE6-C7. In panel B, *p < 0.05 vs mice in the sham group. |
HOTAIR Silencing Attenuates EMT in PC
The Western blot analysis was used to detect EMT-related gene expression. The results of the analysis revealed that in comparison to the cells without treatment and cells treated with the empty vector, E-cadherin was increased and N-cadherin and Vimentin were decreased in cells treated with HOTAIR shRNA in PANC-1 (Figure 5A) and Miapaca-2 (Figure 5B) cell lines (p < 0.05). Therefore, it was established that the EMT in PC was inhibited when HOTAIR was knocked down. Subsequently, the cell morphology was observed under a light microscopy, and it was found that the cell lines overexpressing HOTAIR exhibited interstitial cell morphology (Figure 5C). Additionally, the CCK-8 analysis was employed to detect the proliferative capacity of the two types of cells (PANC-1 and Miapaca-2 cell lines). In the cells overexpressing HOTAIR, the OD value was significant higher than that in the other groups (p < 0.05), and it was obvious lower in the cells with HOTAIR knockdown than that in the other groups (p < 0.05, Figure 5D). Moreover, Transwell assay was employed to detect the migratory and invasive abilities of cells (Figure 5E). In cells overexpressing HOTAIR, the migratory and invasive abilities of cells were stronger than those in other treatment groups (p < 0.05). The above experimental results further confirmed our hypothesis that the silencing of HOTAIR could limit the EMT of tumor cells and inhibit the abilities of cell proliferation, migration, and invasion.
 |
Figure 5 HOTAIR silencing suppressed EMT in PC. (A) protein levels of E-cadherin, N-cadherin, Vimentin in PANC-1 cells treated with oe-HOTAIR mimic or sh-HOTAIR by Western blot analysis as normalized to GAPDH; (B) protein levels of E-cadherin, N-cadherin, Vimentin in Miapaca-2 cells treated with oe-HOTAIR or sh-HOTAIR as normalized to GAPDH; (C) cell morphology of differently treated cells; (D) CCK-8 proliferation result of PANC-1 and Miapaca-2 cells; (E) migration and invasion results of PANC-1 and Miapaca-2 cells. *p < 0.05 vs cells without treatment and cells treated with empty vector. |
HOTAIR Upregulates the Wnt/β-Catenin Signaling Pathway in PC Cells
The expression of genes related to the Wnt/β-catenin signaling pathway was determined using the RT-qPCR to confirm to correlation between HOTAI and the Wnt/β-catenin signaling pathway in PC cells. The results of the analysis confirmed that in contrast to the cells without treatment and the cells treated with the empty vector, the overexpression of HOTAIR increased β-catenin, cyclinD1, c-myc, LEF-1 and c-Jun expression, while the β-catenin, cyclinD1, c-myc, LEF-1 and c-Jun expression was inhibited when HOTAIR was knocked down (p < 0.05) (Figure 6). According to the aforementioned results, HOTAIR up-regulated the Wnt/β-catenin signaling pathway in PC cells cultured in vitro.
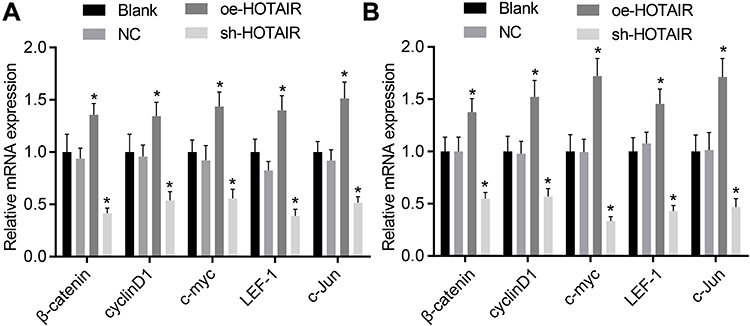 |
Figure 6 HOTAIR silencing inhibited the Wnt/β-catenin signaling pathway in PC cells. (A) mRNA levels of Wnt/β-catenin signaling pathway-related genes in PANC-1 cell line; (B) mRNA levels of Wnt/β-catenin signaling pathway-related genes in Miapaca-2 cell line. *p < 0.05 vs cells without treatment and cells treated with empty vector. |
Wnt/β-Catenin Signaling Pathway Inhibition Attenuates EMT in PC
Western blot analysis was conducted to determine the role of the Wnt/β-catenin signaling pathway played in EMT in PC. The results (Figure 7) suggested that in comparison to the cells without treatment, the expression of E-cadherin was decreased and the N-cadherin and Vimentin expression was found to be increased in the cells treated with LiC1. Additionally, the expression of E-cadherin was found to be increased and the expressions of N-cadherin and Vimentin were found to be decreased in the cells treated with XAV939 (all p < 0.05). Cumulatively, the abovementioned results indicated that the Wnt/β-catenin signaling pathway inhibition suppressed EMT in PC.
 |
Figure 7 Suppressed Wnt/β-catenin signaling pathway inhibited EMT in PC. (A) protein bands of E-cadherin, N-cadherin, Vimentin in each group by Western blot analysis; (B) protein levels of E-cadherin, N-cadherin, Vimentin in each group as normalized to GAPDH. *p < 0.05 vs cells without treatment. |
Discussion
PC is a fatal disease with a high mortality rate and very few effective therapies.20 A previous study has displayed that the acquisition EMT had close correlation with drug resistance in pancreatic cancer cells21 The current study further investigated the role of LincRNA HOTAIR in EMT in PC through the mediation of the Wnt/β-catenin signaling pathway. The findings of the present study cumulatively suggested that the silencing of HOTAIR attenuates EMT in PC by suppressing the Wnt/β-catenin signaling pathway.
In the current study, HOTAIR was highly expressed in PC cell lines. Recent studies have also revealed that lncRNAs are significant factors in the development of PC such as lncRNA H19 and lncRNA HOTTIP.22,23 HOTAIR has been considered as a poor prognostic biomarker in malignant tumors.24 Additionally, the overexpression of HOTAIR was regarded as a biomarker in the prediction of tumor development in hepatocellular carcinoma.25 Furthermore, another recent study showed that HOTAIR promoted the cell proliferation by up-regulating the insulin-like growth factor 2 expression in PC.26 Another previously conducted study has also established that HOTAIR functioned as a negative factor in PC, thereby enhancing the vitality of PC cells.10 Moreover, it was found that the TNF-related apoptosis-inducing ligand resistance in PC was inhibited by overexpressed HOTAIR.27 Therefore, the current study considered that HOTAIR participated in PC development.
Furthermore, it was confirmed that the expression of β-catenin, cyclinD1, c-myc, LEF-1 and c-Jun was decreased when HOTAIR was silenced in PC mice. It was further suggested that cyclinD1, c-Myc, and LEF-1 were downstream genes of the Wnt/β-catenin signaling pathway.28 Additionally, abnormal β-catenin expression was found commonly in PC, and it was also established that it participated in the organogenesis and oncogenesis.29 Moreover, a previous study revealed that the overexpression of β-catenin contributed to the EMT in breast cancer.30 Another study has also suggested that β-catenin and Wnt were inhibited when HOTAIR was knocked down in hepatocellular carcinoma.31 Furthermore, a previously conducted study revealed that cyclinD1 was a key regulator in the cell cycle, and that it participated in many cancers, such as esophageal squamous cell carcinoma and colorectal cancer.32 C-myc was a transcription factor, and the silencing of HOTAIR attenuated c-myc induced cancer development.33 Additionally, it was established that HOTAIR activated the Wnt signaling pathway, and thereby participated in the progression of cell metastasis in esophageal squamous cell carcinoma.34 HOTAIR activation was also found to enhance the initiation and chemoresistance of ovarian cancer by activating the Wnt/β-catenin signaling in human ovarian cancer.35 Therefore, the current study assumed that silenced HOTAIR inhibited the Wnt/β-catenin signaling pathway.
Moreover, E-cadherin upregulation and N-cadherin and Vimentin downregulation were found in PC when HOTAIR or Wnt/β-catenin signaling pathway was inhibited which suggested that HOTAIR silencing attenuated the EMT development. The EMT was considered a key event in cancer metastasis, with increased cell motility and invasiveness, and loss of epithelial marker E-cadherin and gain of mesenchymal marker N-cadherin.36 Additionally, EMT was characterized by the up-regulated mesenchymal cell markers like Vimentin.37 Vimentin was confirmed to be a significant marker of EMT in regulating mesenchymal cell migration.38 A previously conducted study has established that decreased E-cadherin and increased Vimentin were closely positively correlated with diseases severity.39 Another recent study displayed that HOTAIR played a significant role in promoting tumor cell invasion and metastasis by regulating EZH2 and suppressing E-cadherin in oral squamous cell carcinoma.40 Furthermore, a previous study revealed that HOTAIR increased the expression of E-cadherin and decreased the expression of Vimentin, thereby attenuating cell proliferation and cell invasion in human hypopharyngeal carcinoma cells.41 Additionally, it has been established that the Wnt/β-catenin signaling pathway played an important role in regulating cell fate and behavior during embryogenesis, adult tissue homeostasis, and regeneration when it was inappropriately activated.42 The activation of the Wnt/β-catenin signaling pathway promoted tumor growth and cell metastasis and EMT in non-small cell lung cancer.43 Moreover, the enhancement of the Wnt/β-catenin signaling pathway has also been shown to contribute to the EMT in breast cancer.44 Herein, we considered HOTAIR silencing inhibited EMT in PC by suppressing Wnt/β-catenin signaling pathway.
Conclusion
Conclusively, the current study found that HOTAIR was overexpressed in PC, and it promoted EMT in PC through the Wnt/β-catenin signaling pathway. Additionally, the upregulation of E-cadherin and the downregulation of N-cadherin and Vimentin were found in PC when HOTAIR or the Wnt/β-catenin signaling pathway was inhibited which suggested that HOTAIR silencing attenuated the EMT development, thereby suppressing PC development. The study provided theoretical basis in PC and it may open a novel therapeutic target in PC treatment. However, more statistics are necessary to provide more credible results, and a more specific mechanism of HOTAIR in PC is waiting to be discovered.
Abbreviations
PC, pancreatic cancer; lncRNA, long non-coding RNA; EMT, epithelial–mesenchymal transition; DMBA, dimethylbenzoanthracene; NC, negative control; SA, stearylamine; HE, Hematoxylin-eosin; DMEM, Dulbecco’s Modified Eagle Medium; FBS, fetal bovine serum; Lic1, lithium chloride; M-MLV, Moloney Murine Leukemia Virus; RNAase, ribonuclease; dNTP, deoxyribonucleoside triphosphates; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; PVDF, polyvinylidene fluoride; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide-polyacrylamide gel electrophoresis; TBST, tris-buffered Saline with Tween 20; BSA, bovine serum albumin; ANOVA, analysis of variance.
Data Sharing Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
We would like to give our sincere appreciation to the reviewers for their helpful comments on this article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by General program of Natural Science Foundation of Heilongjiang Province (No. D200826); General Projects of Scientific Research and Innovation Fund of the First Affiliated Hospital of Harbin Medical University (No. 2017Y006).
Disclosure
The authors declare that they have no competing interests.
References
1. Neoptolemos JP, Stocken DD, Bassi C, et al. Adjuvant chemotherapy with fluorouracil plus folinic acid vs gemcitabine following pancreatic cancer resection: a randomized controlled trial. JAMA. 2010;304(10):1073–1081. doi:10.1001/jama.2010.1275
2. Fric P, Sedo A, Skrha J, et al. Early detection of sporadic pancreatic cancer: time for change. Eur J Gastroenterol Hepatol. 2017;29(8):885–891. doi:10.1097/MEG.0000000000000904
3. Arslan AA. Anthropometric measures, body mass index, and pancreatic cancer: a pooled analysis from the Pancreatic Cancer Cohort Consortium (PanScan). Arch Intern Med. 2010;170(9):791–802. doi:10.1001/archinternmed.2010.63
4. Campbell PJ, Yachida S, Mudie LJ, et al. The patterns and dynamics of genomic instability in metastatic pancreatic cancer. Nature. 2010;467(7319):1109–1113. doi:10.1038/nature09460
5. Butturini G, et al. Influence of resection margins and treatment on survival in patients with pancreatic cancer: meta-analysis of randomized controlled trials. Arch Surg. 2008;143(1):75–83. doi:10.1001/archsurg.2007.17
6. Nentwich MF, Konig A, Izbicki JR. Limits of surgery for pancreatic cancer. Rozhl Chir. 2014;93(9):445–449.
7. Jayakodi M, Jung JW, Park D, et al. Genome-wide characterization of long intergenic non-coding RNAs (lincRNAs) provides new insight into viral diseases in honey bees Apis cerana and Apis mellifera. BMC Genomics. 2015;16(1):680. doi:10.1186/s12864-015-1868-7
8. Bhan A, Hussain I, Ansari KI, Kasiri S, Bashyal A, Mandal SS. Antisense transcript long noncoding RNA (lncRNA) HOTAIR is transcriptionally induced by estradiol. J Mol Biol. 2013;425(19):3707–3722. doi:10.1016/j.jmb.2013.01.022
9. Li H, An J, Wu M, et al. LncRNA HOTAIR promotes human liver cancer stem cell malignant growth through downregulation of SETD2. Oncotarget. 2015;6(29):27847–27864. doi:10.18632/oncotarget.4443
10. Kim K, Jutooru I, Chadalapaka G, et al. HOTAIR is a negative prognostic factor and exhibits pro-oncogenic activity in pancreatic cancer. Oncogene. 2013;32(13):1616–1625. doi:10.1038/onc.2012.193
11. Tarapore RS, Siddiqui IA, Mukhtar H. Modulation of Wnt/ -catenin signaling pathway by bioactive food components. Carcinogenesis. 2012;33(3):483–491. doi:10.1093/carcin/bgr305
12. Yao H, Ashihara E, Maekawa T. Targeting the Wnt/β-catenin signaling pathway in human cancers. Expert Opin Ther Targets. 2011;15(7):873–887. doi:10.1517/14728222.2011.577418
13. Taube JH, Herschkowitz JI, Komurov K, et al. Core epithelial-to-mesenchymal transition interactome gene-expression signature is associated with claudin-low and metaplastic breast cancer subtypes. Proc Natl Acad Sci U S A. 2010;107(35):15449–15454. doi:10.1073/pnas.1004900107
14. Cascioferro S, Petri GL, Parrino B, et al. Imidazo[2,1-b] [1,3,4]thiadiazoles with antiproliferative activity against primary and gemcitabine-resistant pancreatic cancer cells. Eur J Med Chem. 2020;189:112088. doi:10.1016/j.ejmech.2020.112088
15. Zhu H, Wang D, Zhang L, et al. Upregulation of autophagy by hypoxia-inducible factor-1α promotes EMT and metastatic ability of CD133+ pancreatic cancer stem-like cells during intermittent hypoxia. Oncol Rep. 2014;32(3):935–942. doi:10.3892/or.2014.3298
16. Liu -Q-Q, Chen K, Ye Q, Jiang X-H, Sun Y-W. Oridonin inhibits pancreatic cancer cell migration and epithelial-mesenchymal transition by suppressing Wnt/β-catenin signaling pathway. Cancer Cell Int. 2016;16(1):57. doi:10.1186/s12935-016-0336-z
17. Zhou W, Li Y, Gou S, et al. MiR-744 increases tumorigenicity of pancreatic cancer by activating Wnt/β-catenin pathway. Oncotarget. 2015;6(35):37557–37569. doi:10.18632/oncotarget.5317
18. Cui J, Jiang W, Wang S, Wang L, Xie K. Role of Wnt/β-catenin Signaling in Drug Resistance of Pancreatic Cancer. Curr Pharm Des. 2012;18(17):2464–2471. doi:10.2174/13816128112092464
19. Khalaileh A, Dreazen A, Khatib A, et al. Phosphorylation of ribosomal protein S6 attenuates DNA damage and tumor suppression during development of pancreatic cancer. Cancer Res. 2013;73(6):1811–1820. doi:10.1158/0008-5472.CAN-12-2014
20. Zelinkova Z, van der Ent C, Bruin KF, et al. Effects of Discontinuing Anti–Tumor Necrosis Factor Therapy During Pregnancy on the Course of Inflammatory Bowel Disease and Neonatal Exposure. Clin Gastroenterol Hepatol. 2013;11(3):318–321. doi:10.1016/j.cgh.2012.10.024
21. Ma J, Fang B, Zeng F, et al. Down-regulation of miR-223 reverses epithelial-mesenchymal transition in gemcitabine-resistant pancreatic cancer cells. Oncotarget. 2015;6(3):1740–1749. doi:10.18632/oncotarget.2714
22. Li Z, Zhao X, Zhou Y, et al. The long non-coding RNA HOTTIP promotes progression and gemcitabine resistance by regulating HOXA13 in pancreatic cancer. J Transl Med. 2015;13(1):84. doi:10.1186/s12967-015-0442-z
23. Yoshimura H, Matsuda Y, Yamamoto M, et al. Reduced expression of the H19 long non-coding RNA inhibits pancreatic cancer metastasis. Lab Invest. 2018;98(6):814–824. doi:10.1038/s41374-018-0048-1
24. Endo H, Shiroki T, Nakagawa T, et al. Enhanced expression of long non-coding RNA HOTAIR is associated with the development of gastric cancer. PLoS One. 2013;8(10):e77070. doi:10.1371/journal.pone.0077070
25. Yang Z, Zhou L, Wu L-M, et al. Overexpression of long non-coding RNA HOTAIR predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Ann Surg Oncol. 2011;18(5):1243–1250. doi:10.1245/s10434-011-1581-y
26. Cai H, An Y, Chen X, et al. Epigenetic inhibition of miR-663b by long non-coding RNA HOTAIR promotes pancreatic cancer cell proliferation via up-regulation of insulin-like growth factor 2. Oncotarget. 2016;7(52):86857–86870. doi:10.18632/oncotarget.13490
27. Yang S-Z, Xu F, Zhou T, Zhao X, McDonald JM, Chen Y. The long non-coding RNA HOTAIR enhances pancreatic cancer resistance to TNF-related apoptosis-inducing ligand. J Biol Chem. 2017;292(25):10390–10397. doi:10.1074/jbc.M117.786830
28. Wang Q, Cai J, Cai X-H, Chen L, Shi X-M. miR-346 Regulates Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells by Targeting the Wnt/β-Catenin Pathway. PLoS One. 2013;8(9):e72266. doi:10.1371/journal.pone.0072266
29. Sarkar S, Mandal C, Sangwan R, Mandal C. Coupling G2/M arrest to the Wnt/β-catenin pathway restrains pancreatic adenocarcinoma. Endocr Relat Cancer. 2014;21(1):113–125. doi:10.1530/ERC-13-0315
30. Yan D, Avtanski D, Saxena NK, Sharma D. Leptin-induced Epithelial-Mesenchymal Transition in Breast Cancer Cells Requires β-Catenin Activation via Akt/GSK3- and MTA1/Wnt1 Protein-dependent Pathways. J Biol Chem. 2012;287(11):8598–8612. doi:10.1074/jbc.M111.322800
31. Gao J-Z, Li J, Du J-L, Li X-L. Long non-coding RNA HOTAIR is a marker for hepatocellular carcinoma progression and tumor recurrence. Oncol Lett. 2016;11(3):1791–1798. doi:10.3892/ol.2016.4130
32. Wang M-T, Chen G, An S-J, et al. Prognostic significance of cyclinD1 amplification and the co-alteration of cyclinD1/pRb/ppRb in patients with esophageal squamous cell carcinoma. Dis Esophagus. 2012;25(7):664–670. doi:10.1111/j.1442-2050.2011.01291.x
33. Huang L, Li F, Deng P, Hu C. [ARTICLE WITHDRAWN] MicroRNA-223 Promotes Tumor Progression in Lung Cancer A549 Cells via Activation of the NF-κB Signaling Pathway. Oncol Res. 2016;24(6):405–413. doi:10.3727/096504016X14685034103437
34. Ge X-S, Ma H-J, Zheng X-H, et al. HOTAIR, a prognostic factor in esophageal squamous cell carcinoma, inhibits WIF-1 expression and activates Wnt pathway. Cancer Sci. 2013;104(12):1675–1682. doi:10.1111/cas.12296
35. Li J, Yang S, Su N, et al. Erratum to: overexpression of long non-coding RNA HOTAIR leads to chemoresistance by activating the Wnt/β-catenin pathway in human ovarian cancer. Tumour Biol. 2015;36(11):9093–9094. doi:10.1007/s13277-015-4210-8
36. Yang H, Zhan L, Yang T, et al. Ski prevents TGF-β-induced EMT and cell invasion by repressing SMAD-dependent signaling in non-small cell lung cancer. Oncol Rep. 2015;34(1):87–94. doi:10.3892/or.2015.3961
37. Han M, Wang Y, Liu M, et al. MiR-21 regulates epithelial-mesenchymal transition phenotype and hypoxia-inducible factor-1α expression in third-sphere forming breast cancer stem cell-like cells. Cancer Sci. 2012;103(6):1058–1064. doi:10.1111/j.1349-7006.2012.02281.x
38. Vuoriluoto K, Haugen H, Kiviluoto S, et al. Vimentin regulates EMT induction by Slug and oncogenic H-Ras and migration by governing Axl expression in breast cancer. Oncogene. 2011;30(12):1436–1448. doi:10.1038/onc.2010.509
39. Chaw SY, Abdul Majeed A, Dalley AJ, Chan A, Stein S, Farah CS. Epithelial to mesenchymal transition (EMT) biomarkers – E-cadherin, beta-catenin, APC and Vimentin – in oral squamous cell carcinogenesis and transformation. Oral Oncol. 2012;48(10):997–1006. doi:10.1016/j.oraloncology.2012.05.011
40. Wu Y, Zhang L, Zhang L, et al. Long non-coding RNA HOTAIR promotes tumor cell invasion and metastasis by recruiting EZH2 and repressing E-cadherin in oral squamous cell carcinoma. Int J Oncol. 2015;46(6):2586–2594. doi:10.3892/ijo.2015.2976
41. Cheng H, Cui X, Guo Y, et al. The central role of HOTAIR in the malignancy of CD44+ human hypopharyngeal carcinoma cells. Am J Transl Res. 2016;8(11):5052–5058.
42. Zhang Q, Major MB, Takanashi S, et al. Small-molecule synergist of the Wnt/β-catenin signaling pathway. Proc Natl Acad Sci U S A. 2007;104(18):7444–7448. doi:10.1073/pnas.0702136104
43. Yang S, Liu Y, Li M-Y, et al. FOXP3 promotes tumor growth and metastasis by activating Wnt/β-catenin signaling pathway and EMT in non-small cell lung cancer. Mol Cancer. 2017;16(1):124. doi:10.1186/s12943-017-0700-1
44. Cai J, Guan H, Fang L, et al. MicroRNA-374a activates Wnt/β-catenin signaling to promote breast cancer metastasis.. J Clin Invest. 2013;123(2):566–579. doi:10.1172/JCI65871
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.