")
Back to Journals » OncoTargets and Therapy » Volume 13
Significant Contribution of DNA Repair Human 8-Oxoguanine DNA N-Glycosylase 1 Genotypes to Renal Cell Carcinoma
Authors Chang WS, Shen TC , Liao JM, Tsai YT , Hsia TC, Wu HC, Tsai CW, Bau DT
Received 19 September 2019
Accepted for publication 6 February 2020
Published 20 February 2020 Volume 2020:13 Pages 1583—1591
DOI https://doi.org/10.2147/OTT.S231733
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Takuya Aoki
Wen-Shin Chang,1,* Te-Chun Shen,1– 3,* Jiuan-Miaw Liao,4,* Yueh-Ting Tsai,1 Te-Chun Hsia,2 Hsi-Chin Wu,1,3 Chia-Wen Tsai,1 Da-Tian Bau1,5,6
1Terry Fox Cancer Research Laboratory, Translational Medicine Research Center, China Medical University Hospital, Taichung, Taiwan; 2Division of Pulmonary and Critical Care Medicine, Department of Internal Medicine, China Medical University Hospital, Taichung, Taiwan; 3School of Medicine, China Medical University, Taichung, Taiwan; 4Department of Physiology, Chung Shan Medical University and Chung Shan Medical University Hospital, Taichung, Taiwan; 5Graduate Institute of Biomedical Sciences, China Medical University, Taichung, Taiwan; 6Department of Bioinformatics and Medical Engineering, Asia University, Taichung, Taiwan
*These authors contributed equally to this work
Correspondence: Da-Tian Bau; Chia-Wen Tsai
Terry Fox Cancer Research Laboratory, Translational Medicine Research Center, China Medical University Hospital, No. 2 Yude Road, Taichung 404, Taiwan
Tel +886-4-22052121
Email [email protected]
Introduction: DNA repair systems play essential roles in genomic stability and carcinogenesis. Therefore, genotypes at DNA repair loci may contribute to the determination of personal susceptibility to cancers. The contribution of human 8-oxoguanine DNA N-glycosylase 1 (hOGG1) genotypes to renal cell carcinoma (RCC) is largely unknown. This study aimed to evaluate the contributions of hOGG1 rs1052133 genotypes to the RCC risk.
Methods: We evaluated the contribution of hOGG1 rs1052133 (G/C) genotypes among 118 cases and 590 controls and analyzed the interactions of hOGG1 genotypes with smoking, alcohol drinking, hypertension, and diabetes status.
Results: The hOGG1 rs1052133 CC genotype was significantly associated with a decreased RCC risk compared with that of the GG genotype (odds ratio [OR] = 0.25, 95% confidence interval [CI] = 0.09– 0.72, p = 0.0049). The frequency of the rs1052133 C allele was significantly low in the RCC group (22.5% vs 31.2%; OR = 0.64; 95% CI = 0.46– 0.89, p = 0.0074). Stratifying the analysis according to smoking, alcohol drinking, and diabetes status revealed no difference in the rs1052133 genotype distribution among these subgroups. A significant differential distribution of rs1052133 genotypes was observed among subjects with hypertension.
Conclusion: The CC genotype of rs1052133 may play a role in determining RCC susceptibility among Taiwanese people and may serve as a biomarker of RCC, particularly in patients with hypertension.
Keywords: hOGG1, genotype, polymorphism, renal cell carcinoma
Introduction
Renal cell carcinoma (RCC) imposes a serious disease burden; among the most frequently diagnosed cancers worldwide, RCC is the 6th one in men and the 10th in women.1 Clinically speaking, RCC has been recognized as the most common renal cancer, and its subtypes can be distinguished from one another based on their differences in histology, genetic background, clinical course, and differential responses to treatment.2,3 From a personalized medical viewpoint, several behavioral factors, such as personal physical activity, obesity, fruit and vegetable intake, cigarette smoking, and alcohol consumption, are identified as risk factors of RCC. Some clinical and medical comorbidities, including hypertension, diabetes, urinary stones, other forms of chronic kidney disease, and a family history of cancer, are associated with RCC.3 However, to date, few clinically practical biomarkers in the genome have been identified as early predictors of the RCC risk. Many patients with RCC, even those with advanced stage tumors, are free of obvious symptoms,2,4 and RCC is not detectable at the first visit to doctors. The worst situation is that up to 30% of patients with RCC treated with radical nephrectomy suffer from many adverse effects and face high odds of recurrence soon after their surgery.5 Currently, personal identification as a patient with RCC is based on a series of labor-intensive and time-consuming histological validation processes. Therefore, genomic biomarkers should be elucidated for the precise detection and identification of RCC.
Several gatekeepers, such as DNA repair systems, including base excision repair (BER), mismatch repair, and nucleotide excision repair, which are responsible for maintaining the stability of our genome, have evolved and cooperated to prevent cells from undergoing carcinogenesis.6–8 The most abundant oxidative DNA adduct 8‐oxoguanine (8‐oxoG), mainly produced by reactive oxygen species, is the cause of oxidative damage leading to a transversion from G:C→T:A and causing the development of carcinogenesis.9,10 Human 8-oxoG DNA N-glycosylase 1 (hOGG1) is a pilot enzyme in the BER pathway for the detection and recognition of 8‐oxoG in our genome.6,11 Functional studies have shown that the genotype at rs1052133 (Ser326Cys) single nucleotide polymorphism (SNP) in exon 7 of hOGG1 may determine the glycosylase activity12,13 and serve as a genomic predictor of personal susceptibility to various cancer types.14–19
Therefore, in this study, we aimed to determine whether the hOGG1 rs1052133 polymorphism is associated with the RCC risk in Taiwan and to elucidate the interactions of this SNP with several clinical and behavioral factors.
Materials and Methods
Investigated Population
This case–control study was conducted in China Medical University Hospital. RCC was diagnosed by Dr. Wu’s surgical team, while the grades and types of each patient with RCC were histopathologically confirmed by well-trained pathologists. Among 139 patients interviewed, 118 (85%) agreed to participate and were included in this study. For each patient with RCC, five controls were frequency-matched and recruited in the Health Examination Center of China Medical University Hospital. They had the same gender as the patients and ± 2 years of his/her age. None of the cancer-free controls had any biological relationship with one another. The inclusion criteria of the controls were the citizens of Taiwan without any history of any cancer type. All the controls were within the normal range of carcinoembryonic antigen (CEA) and identified as cancer-free. After the pre-screen first-term matching process, patients with incomplete demographic data about smoking, alcohol drinking status, hypertension, diabetes, or family history of cancer were excluded. Then, a further exclusion criterion among the control candidates was set as any symptom suggestive of RCC, such as hematuria. A five-fold number (590) of controls were retained for genotyping experiments and analytical processes. Each participant completed a written informed consent and provided 3–5 mL of his/her venous blood for the genotyping experiments under the guidance and supervision of the Institutional Review Board of China Medical University Hospital (DMR98-IRB-209). All the clinical investigations and records in this study were restrictively performed in accordance with the principles expressed in the Declaration of Helsinki. The selected demographic characteristics of all the participants are summarized and compared in Table 1.
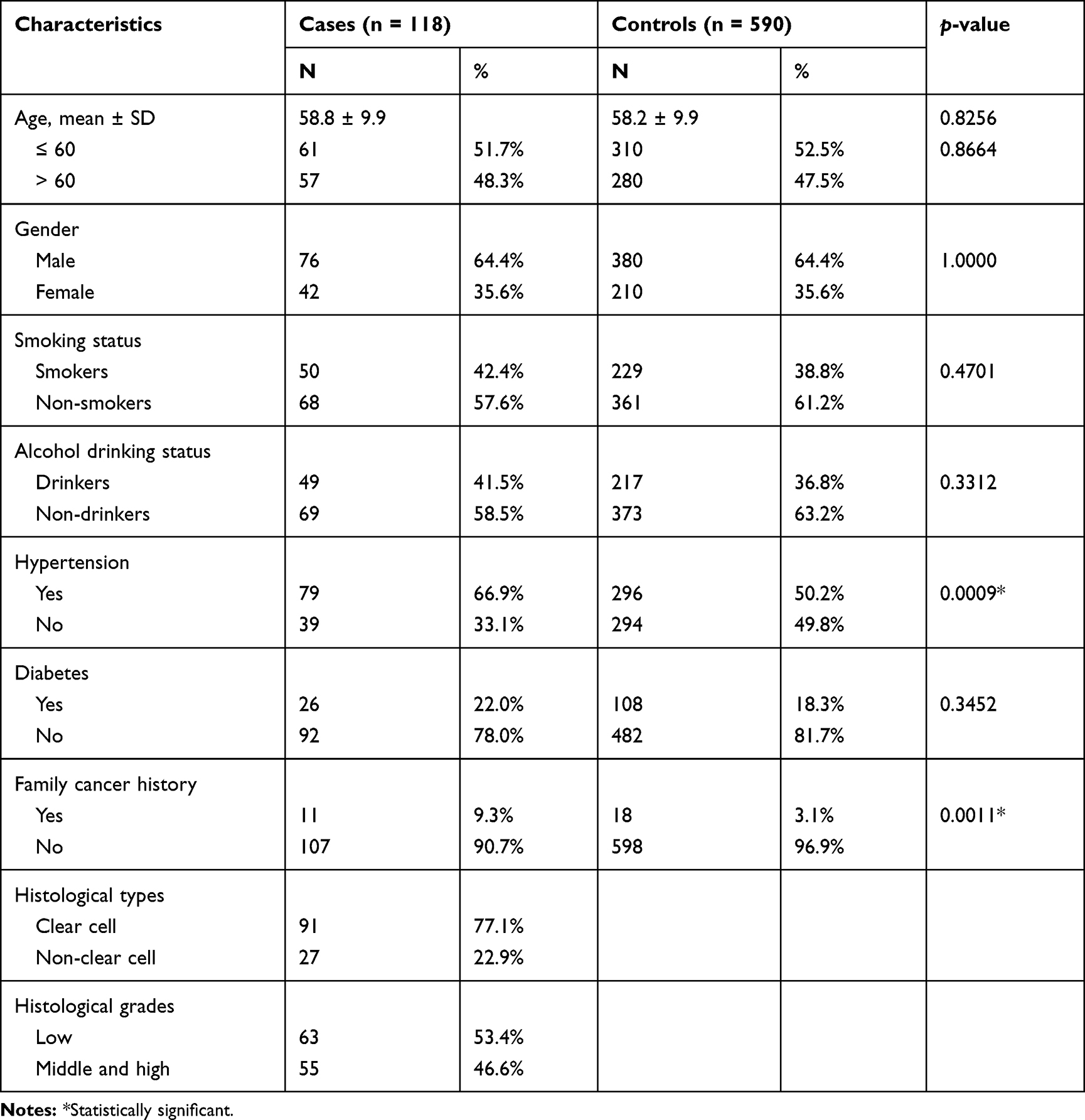 |
Table 1 Analysis of the Distributions of Demographic Characteristics Among the RCC Cases and Healthy Controls |
DNA Preparation and Storage
Genomic DNA from the leukocytes of each study subject was extracted within 24 h after collection by using a QIAamp blood mini kit (Qiagen, Valencia, CA, USA), stored at −80°C, simultaneously diluted, aliquoted, and stored for genotyping as a working stock at −20°C as we previously reported.20–24
hOGG1 Genotype Discrimination Methodology
The hOGG1 rs1052133 polymorphic site was genotyped as we previously described in 2013.18,25 Polymerase chain reaction (PCR)–restriction fragment length polymorphism analysis was performed. The sequences of forward and reverse primers for hOGG1 rs1052133 genotyping were 5ʹ-ACTGTCACTAGTCTCACCAG-3ʹ and 5ʹ-GGAAGGTGGGAAGGTG-3ʹ, respectively. In detail, 100 ng of the genomic DNA of each sample was subjected to a typical PCR. In this procedure, a 25 μL reaction mixture contained 300 μM dNTP, 2 U Taq DNA polymerase, 1× PCR buffer, 1.5 mM MgCl2, and 0.8 μM of each designed primer. After the mixture was thoroughly mixed, the overall reaction mixture was heated to 94°C for 4 mins and steadily amplified with My Cycler (Bio-Rad, Hercules, CA, USA) with 30 cycles of denaturation at 94°C for 60 s, annealing at 60°C for 60 s, and extension at 72°C for 60 s. A final extension step at 72°C for 5 mins was conducted. The volume of the restriction enzyme digestion was set at 12.5 μL that contained 8 μL of PCR products, 2 U Fnu4H I restriction enzyme, and 1× buffer. The reaction mixture was then incubated at 60°C for 16 hrs or overnight to allow complete digestion. The resultant DNA fragments were then subjected to 3.0% agarose gel electrophoresis at a constant voltage of 100 V for 30 mins. After electrophoresis, the gels were immediately stained with ethidium bromide and imaged under UV (260 nm) light to observe DNA fragments. For the C allele of hOGG1 rs1052133, the single 200 bp fragment was no longer digested. For the G allele, two (100 and 100 bp) fragments were observed as a single band after gel electrophoresis. Any result with both types of bands was identified as the heterovariant CG genotype of hOGG1 rs1052133.
Statistical Analysis
In this study, 590 cancer-free healthy controls and 118 patients with RCC were analyzed for their genotypic and clinical details as shown in Tables 1 and 2 and Figures 1–4. The deviation of the genotype frequencies of hOGG1 SNPs in the control subjects from those expected under Hardy–Weinberg equilibrium was assessed using a goodness-of-fit test to ensure that the control subjects in this study were representative of the general Taiwanese population and to exclude the possibility of genotyping error (Table 2). Pearson’s chi-square test and Fisher’s exact test (when any cell was less than 5 persons) were adopted to compare the distribution of hOGG1 genotypes between the cases and control groups and conduct stratification analysis. The difference in age, which is a continuous factor, was evaluated via Student’s t-test. The association of hOGG1 genotypes with the risk of RCC was estimated in terms of odds ratios (ORs) and their counterpart 95% confidence intervals (CIs) through logistic regression analysis with or without adjustment for possible confounders as indicated in table footnotes. Any p < 0.05 was considered to indicate a statistically significant result.
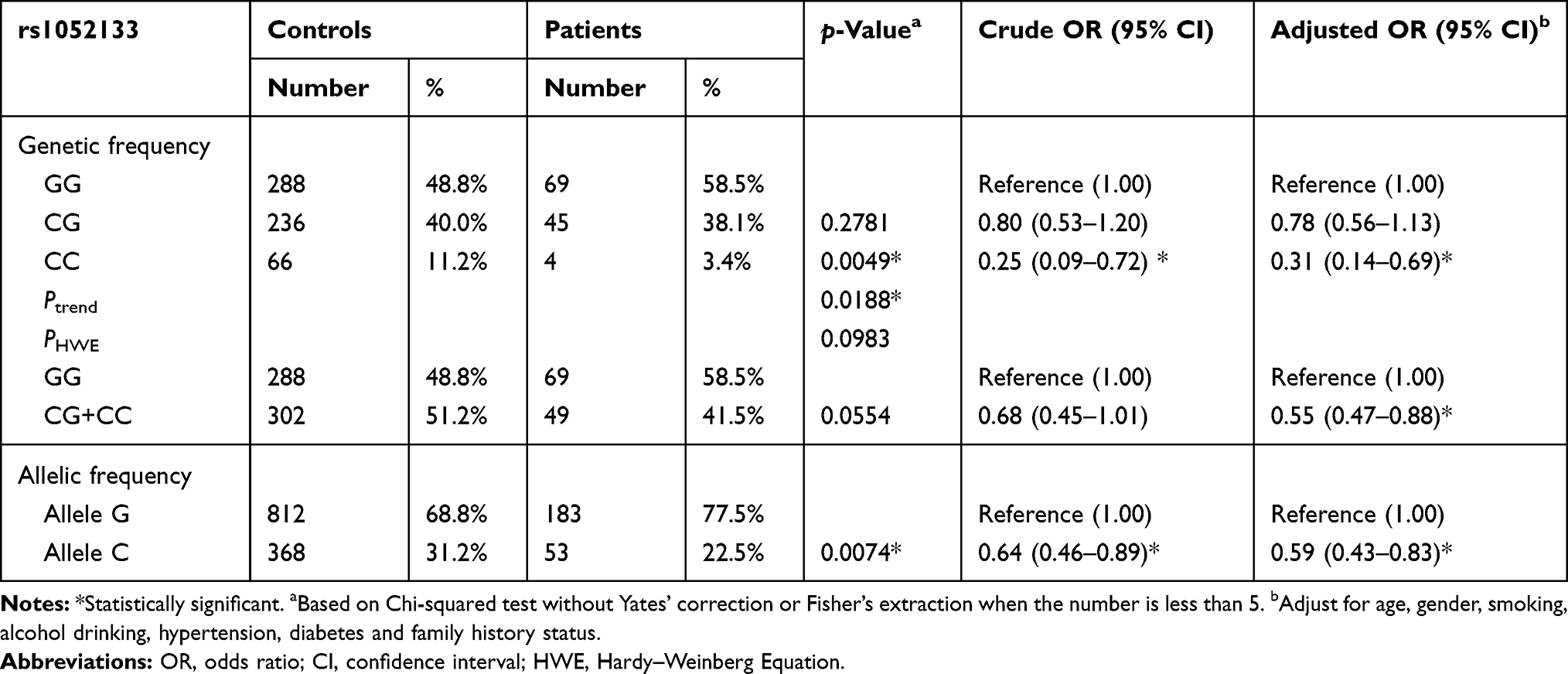 |
Table 2 Analysis of hOGG1 Rs1052133 Genotypic and Allelic Frequency Distributions Among the Renal Cell Carcinoma Patients and Healthy Controls |
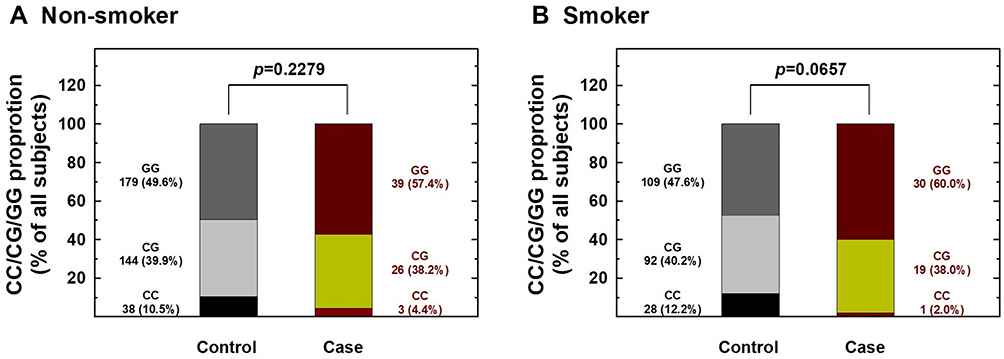 |
Figure 1 Contribution of hOGG1 rs1052133 genotype to the risk of renal cell carcinoma after stratification by smoking status. The distributions of GG, CG, and CC genotypes at hOGG1 rs1052133 among non-smokers (A) and smokers (B). |
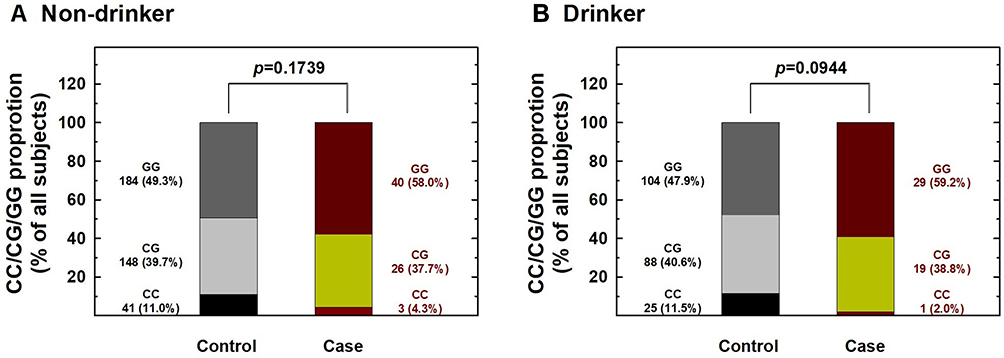 |
Figure 2 Contribution of hOGG1 rs1052133 genotype to the risk of renal cell carcinoma after stratification by alcohol consumption status. The distributions of GG, CG, and CC genotypes at hOGG1 rs1052133 among non-drinkers (A) and drinkers (B). |
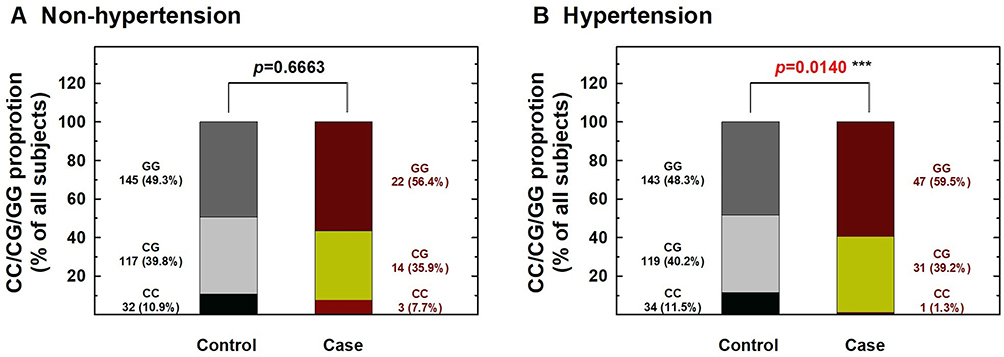 |
Figure 3 Contribution of hOGG1 rs1052133 genotype to the risk of renal cell carcinoma after stratification by hypertension status. The distributions of GG, CG, and CC genotypes at hOGG1 rs1052133 among individuals without (A) and with (B) hypertension. (*** Statistically significant between case and control groups). |
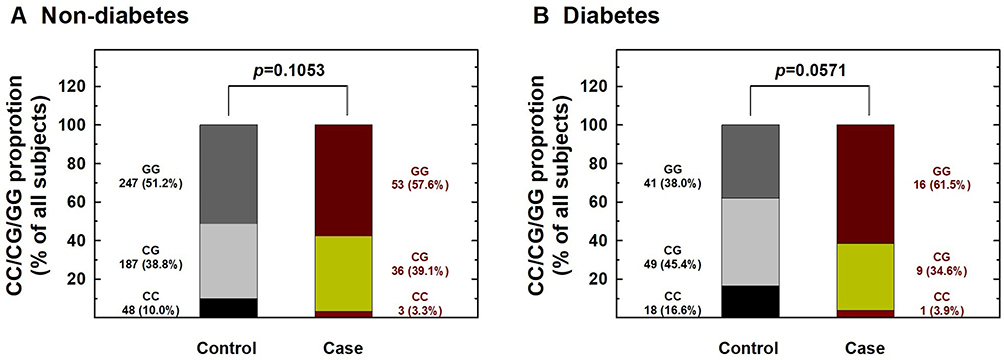 |
Figure 4 Contribution of hOGG1 rs1052133 genotype to the risk of renal cell carcinoma after stratification by diabetes status. The distributions of GG, CG, and CC genotypes at hOGG1 rs1052133 among individuals without (A) and with (B) diabetes. |
Results
Comparisons of Demographic Characteristics Among the Investigated Subjects
The frequency distributions of age, gender, and behavioral habits, such as smoking and alcohol drinking status, of the 118 patients with RCC and the 590 cancer-free control subjects were compared (Table 1). The control subjects were matched with the patients with RCC for age and gender during subject selection, so no difference was observed between the two groups in terms of age and gender (p > 0.05). No preferential difference was found in the frequencies of diabetes status or personal behavioral habits, such as smoking or alcohol consumption (p > 0.05) between the two groups (Table 1). Interestingly, the rate of family history of cancer of patients with RCC was higher than that of the controls (9.3% versus 3.1%, p = 0.0011). This result indicated that RCC is an inherited disease. The results also showed that the percentage of subjects with hypertension in the RCC group (66.9%) was higher than that of the control group (50.2%; p = 0.0009). From a histological angle, 77.1% of patients had the most frequently occurring RCC subtype, namely, clear cell RCC (ccRCC). The proportions of patients with low-grade and middle- and high-grade RCC were 53.4% and 46.6%, respectively (Table 1).
Analysis of the Association of hOGG1 Genotypes with RCC Risk in Taiwan
The observed genotypic and allelic frequencies of hOGG1 rs1052133 among RCC cases and controls and their associations with the risk of RCC are summarized in Table 2. The hOGG1 rs1052133 genotypes among healthy controls were in a Hardy–Weinberg equilibrium (p = 0.0983). In the trend analysis, the distributions of hOGG1 rs1052133 genotypes significantly differed between the control and case groups (p = 0.0188). In detail, the hOGG1 rs1052133 CG and CC variant genotypes were present at frequencies of 40.0% and 11.2% in the control group and 38.1% and 3.4% in the case group, respectively (Table 2, top panel). In multivariate logistic regression analysis, after adjustments were made for age, gender, smoking, alcohol drinking, hypertension, diabetes, and family history status, the hOGG1 rs1052133 homovariant CC was associated with an altered RCC risk (OR = 0.31 and 0.78, 95% CI = 0.14–0.69 and 0.56–1.13, for hOGG1 rs1052133 CC homozygotes and CG heterozygotes, respectively; Table 2, top panel). The hOGG1 rs1052133 variant CG and CC genotypes were subsequently combined to construct a dominant genetic model, and our results revealed that the combined genotypes conferred a significantly reduced RCC risk (OR = 0.55, 95% CI = 0.47–0.88; Table 2; middle panel). We examined the distributions of the allelic frequencies of hOGG1 rs1052133 among the cases and controls and found a significant association between the hOGG1 rs1052133 C allele and a decreased RCC risk in Taiwan (OR = 0.59, 95% CI = 0.43–0.83). Considering that 77.1% of patients had ccRCC, we found that the risk estimates were similar to the overall analysis when we restricted our analysis to patients with ccRCC (data not shown).
Subgroup Stratification Analysis of hOGG1 Rs1052133 Genotypes According to Personal Behavioral and Clinical Factors
We further performed stratification analysis to investigate the association between hOGG1 rs1052133 genotypes and the risk of RCC based on potential Taiwanese-specific personal behavioral and clinical factors, such as cigarette smoking, alcohol consumption, hypertension, and diabetes status, which are listed in Table 1. First, the distributions of genotype frequencies between the RCC case and control groups were similar among nonsmokers and smokers (p > 0.05; Figure 1). The adjusted ORs of the carriers with the CG and CC genotypes at hOGG1 rs1052133 were 0.81 and 0.38 for nonsmokers (95% CI = 0.44–1.37 and 0.14–1.22, respectively) and 0.81 and 0.24 for smokers (95% CI = 0.42–1.38 and 0.09–1.02, respectively; Figure 1), respectively. The results showed no obvious protective effect of hOGG1 rs1052133 genotype on the risk of RCC in nonsmokers or smokers (Figure 1). Second, the distributions of the genotypic frequencies were similar among nondrinkers and alcohol drinkers between the case and control groups (p > 0.05; Figure 2). The adjusted ORs of the carriers with genotypes CG and CC at hOGG1 rs1052133 were 0.82 and 0.37 among nondrinkers (95% CI = 0.51–1.33 and 0.18–1.26, respectively) and 0.76 and 0.18 among alcohol drinkers (95% CI = 0.42–1.53 and 0.09–1.17, respectively; Figure 2), respectively. The protective effects of hOGG1 rs1052133 genotypes on the risk of RCC appeared to be nonsignificant among nondrinkers and alcohol drinkers (Figure 2). Interestingly, the distributions of the genotypic frequencies between the case and control groups were significantly different only in subjects with hypertension but not in subjects without hypertension (Figure 3). The adjusted ORs of the carriers with CG and CC at hOGG1 rs1052133 were 0.80 and 0.67 among the subjects without hypertension (95% CI = 0.41–1.54 and 0.22–2.03, respectively) and 0.81 and 0.09 among the patients with hypertension (95% CI = 0.54–1.28 and 0.01–0.57, respectively; Figure 3). Notably, the protective effects of hOGG1 rs1052133 genotype on the risk of RCC were obvious among people with hypertension, and only the genotype of the homovariant CC was protective (Figure 3). The distributions of hOGG1 rs1052133 genotype frequencies were not significantly different between the case and control groups among the subjects in the subpopulations without or with diabetes (Figure 4). The adjusted ORs of carriers with CG and CC at hOGG1 rs1052133 were 0.91 and 0.16 among the subjects without diabetes (95% CI = 0.61–1.33 and 0.18–1.07, respectively) and 0.66 and 0.31 among those with diabetes (95% CI = 0.24–1.19 and 0.07–1.36, respectively; Figure 4), respectively. The effect of hOGG1 rs1052133 genotype on the RCC risk appeared to be nonprotective regardless of the diabetes status (Figure 4). We also performed a stratified analysis in study participants without a family history and found that the results were similar to the overall analysis (data not shown). This result suggested that the effect of rs1052133 is independent of family history.
Discussion
hOGG1 encodes an 8-oxoguanine DNA glycosylase in charge of the recognition of the most common oxidative DNA adducts, namely, 8‐oxoGs, and this gene acts as an AP-lyase to remove these adducts from the genome via the BER machinery.9,10 As a glycosylase in the first step, hOGG1 not only recognizes 8‐oxoGs but also cleaves the glycosylic bond between the modified base and the sugar moiety. Then, hOGG1 cleaves 3ʹ to the AP site, leaving 5ʹ-phosphate, 3ʹ-phospho-α, β-unsaturated aldehyde, and an apurinic/apyrimidinic site for further actions of DNA polymerase β and DNA ligases I and III.6 The most recognizable polymorphic site of hOGG1 is the well-known rs1052133 (Ser326Cys, C to G), and several genotype–phenotype studies have provided evidence that the glycosylase activity of the “G” variant of the hOGG1 enzyme is more sensitive to the inactivating influence of oxidizing agents than that of the “C” wild type; cells carrying “G” alleles can accumulate mutations more readily under the same challenges of oxidative stress.12,26,27
To our knowledge, only one study has investigated the contribution of hOGG1 to RCC and focused on rs1052133 as we did.28 The G allele at hOGG1 rs1052133 is associated with a 1.4-fold increased risk of RCC in a Chinese population. They found that the contribution is extremely significant in the subgroups of subjects with overweight (defined as a body mass index >24 kg/m2) and nonsmokers.28 This conclusion is consistent with our results that the CC genotype at hOGG1 rs1052133 was associated with a lower risk of RCC than that of the GG genotype (Table 2). In the current study, we further found that the determinant value of hOGG1 rs1052133 of the RCC risk was high among patients with hypertension. Hypertension is a common risk factor of RCC, so this SNP could represent a potential prognosis biomarker of patients with RCC. However, the sample size was limited, and the detailed underlying mechanisms should be further investigated. Their study and ours are both valuable because we have provided genomic information from Eastern populations with relatively representative sample sizes. Studies have rarely investigated the genomic factors contributing to the RCC risk possibly because of the low prevalence of RCC relative to other cancers worldwide. As such, collecting enough samples from patients with RCC for case–control studies is difficult. Additional studies involving larger sample sizes and focusing on various populations are necessary to validate the present findings. To the best of our knowledge, this study was the first to show an association between the hOGG1 rs1052133 polymorphism and the RCC risk in the Taiwanese population. Our novel finding was that hOGG1 genotypes might interact with hypertension to determine the risk of RCC.
The majority of RRCC is ccRCC, and other minor subtypes include papillary, chromophobe, and Bellini collecting duct types. Among our RCC cases, 91 (77.1%) patients had ccRCC, 15 (12.7%) had a papillary type, 9 (7.6%) had a chromophobe type, and 3 (2.5%) had a Bellini collecting duct type. In our stratified analyses, the association of rs1052133 with ccRCC was similar to the overall RCC, whereas the numbers of other non-ccRCC subtypes were too small for meaningful analysis. Although the prognosis of these different subtypes varies, their etiology and risk factors are similar. Genetic susceptibility may also be similar. Nevertheless, future studies with a sufficient number of minor subtypes are warranted to clarify the associations in minor subtypes.
CG and GG at hOGG1 rs1052133 are associated with an increased risk of various cancer types, including oral cancer,19 lung adenocarcinoma,15 breast cancer,29 laryngeal cancer,30 esophageal cancer,31 colorectal cancer,14 gallbladder cancer,16 prostate cancer,32,33 and leukemia.34 Few negative findings have shown no association between hOGG1 rs1052133 genotypes and several cancer types.25,35 Some reports have described controversial findings that the G allele may cause a reduced glycosylase hOGG1 activity, leading to an overall downregulation of the BER capacity.12,13 The link between hOGG1 genotype, DNA repair capacity, and cancer risk still requires further investigations, especially those focusing on genotype–phenotype correlations and measuring the DNA repair capacity of cells with various genotypes. These inconsistencies may be attributed to differences in genomic background, sampling methodology, and small sample sizes. The role of hOGG1 in carcinogenesis is much more complicated than that in BER because few alternative mechanisms can take over the function of hOGG1 when its capacity is downregulated.
In conclusion, our data suggested that hOGG1 rs1052133 genotype is associated with the RCC risk in Taiwan. More functional examinations will contribute to studies on genotype–phenotype correlation. Larger sample sizes with more detailed information about environmental exposure for precise stratification analysis will help reveal the etiology of RCC.
Acknowledgments
We thank the Tissue Bank and colleagues of the Department of Urology in China Medical University Hospital for their excellent technical assistance. The genotyping and analyzing work are partially helped by Yun-Chi Wang, Hsin-Ting Li, and Huai-Mei Hsu. The statistical analyses were kindly double-checked by Cheng-Li Lin (analyzing manpower under MOHW109-TDU-B-212-114004).
Funding
This study was supported mainly by the grant from the Taiwan Ministry of Science and Technology (MOST-107-2320-B-040-028) to Dr. Liao and China Medical University Hospital (DMR108-110) to Dr. Shen.
Disclosure
The authors declare no conflicts of interest related to this study.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68:7–30. doi:10.3322/caac.21442
2. Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 2010;127:2893–2917. doi:10.1002/ijc.25516
3. Klaassen Z, Sayyid RK, Wallis CJD. Lessons learned from the global epidemiology of kidney cancer: a refresher in epidemiology 101. Eur Urol. 2019;75:85–87. doi:10.1016/j.eururo.2018.09.035
4. Niedworok C, Dörrenhaus B, Vom Dorp F, et al. Renal cell carcinoma and tumour thrombus in the inferior vena cava: clinical outcome of 98 consecutive patients and the prognostic value of preoperative parameters. World J Urol. 2015;33:1541–1552. doi:10.1007/s00345-014-1449-4
5. Cohen HT, McGovern FJ. Renal-cell carcinoma. N Engl J Med. 2005;353:2477–2490. doi:10.1056/NEJMra043172
6. Christmann M, Tomicic MT, Roos WP, Kaina B. Mechanisms of human DNA repair: an update. Toxicology. 2003;193:3–34. doi:10.1016/S0300-483X(03)00287-7
7. Dahle J, Brunborg G, Svendsrud DH, Stokke T, Kvam E. Overexpression of human OGG1 in mammalian cells decreases ultraviolet A induced mutagenesis. Cancer Lett. 2008;267:18–25. doi:10.1016/j.canlet.2008.03.002
8. Hung RJ, Hall J, Brennan P, Boffetta P. Genetic polymorphisms in the base excision repair pathway and cancer risk: a HuGE review. Am J Epidemiol. 2005;162:925–942. doi:10.1093/aje/kwi318
9. Cheng KC, Cahill DS, Kasai H, Nishimura S, Loeb LA. 8-Hydroxyguanine, an abundant form of oxidative DNA damage, causes G—T and A—C substitutions. J Biol Chem. 1992;267:166–172.
10. Luna L, Rolseth V, Hildrestrand GA, et al. Dynamic relocalization of hOGG1 during the cell cycle is disrupted in cells harbouring the hOGG1-Cys326 polymorphic variant. Nucleic Acids Res. 2005;33:1813–1824. doi:10.1093/nar/gki325
11. Hill JW, Evans MK. Dimerization and opposite base-dependent catalytic impairment of polymorphic S326C OGG1 glycosylase. Nucleic Acids Res. 2006;34:1620–1632. doi:10.1093/nar/gkl060
12. Yamane A, Kohno T, Ito K, et al. Differential ability of polymorphic OGG1 proteins to suppress mutagenesis induced by 8-hydroxyguanine in human cell in vivo. Carcinogenesis. 2004;25:1689–1694. doi:10.1093/carcin/bgh166
13. Collins AR, Gaivao I. DNA base excision repair as a biomarker in molecular epidemiology studies. Mol Aspects Med. 2007;28:307–322. doi:10.1016/j.mam.2007.05.005
14. Kim JI, Park YJ, Kim KH, et al. hOGG1 Ser326Cys polymorphism modifies the significance of the environmental risk factor for colon cancer. World J Gastroenterol. 2003;9:956–960. doi:10.3748/wjg.v9.i5.956
15. Okasaka T, Matsuo K, Suzuki T, et al. hOGG1 Ser326Cys polymorphism and risk of lung cancer by histological type. J Hum Genet. 2009;54:739–745. doi:10.1038/jhg.2009.108
16. Jiao X, Huang J, Wu S, et al. hOGG1 Ser326Cys polymorphism and susceptibility to gallbladder cancer in a Chinese population. Int J Cancer. 2007;121:501–505. doi:10.1002/ijc.22748
17. Weiss JM, Goode EL, Ladiges WC, Ulrich CM. Polymorphic variation in hOGG1 and risk of cancer: a review of the functional and epidemiologic literature. Mol Carcinog. 2005;42:127–141. doi:10.1002/(ISSN)1098-2744
18. Tsai CW, Ho CY, Shih LC, et al. The joint effect of hOGG1 genotype and smoking habit on endometriosis in Taiwan. Chin J Physiol. 2013;56:263–268. doi:10.4077/CJP.2013.BAB142
19. Tsai CW, Tsai MH, Tsou YA, et al. The joint effect of smoking and hOGG1 genotype on oral cancer in Taiwan. Anticancer Res. 2012;32:3799–3803.
20. Yueh TC, Wu CN, Hung YW, et al. The contribution of MMP-7 genotypes to colorectal cancer susceptibility in Taiwan. Cancer Genomics Proteomics. 2018;15:207–212. doi:10.21873/cgp.20099
21. Chang WS, Shen TC, Yeh WL, et al. Contribution of inflammatory cytokine interleukin-18 genotypes to renal cell carcinoma. Int J Mol Sci. 2019;20:1563. doi:10.3390/ijms20071563
22. Hsu PC, Pei JS, Chen CC, et al. Association of matrix metallopeptidase-2 promoter polymorphisms with the risk of childhood leukemia. Anticancer Res. 2019;39:1185–1190. doi:10.21873/anticanres.13228
23. Tsai YY, Bau DT, Chiang CC, Cheng YW, Tseng SH, Tsai FJ. Pterygium and genetic polymorphism of DNA double strand break repair gene Ku70. Mol Vis. 2007;13:1436–1440.
24. Hsu CF, Tseng HC, Chiu CF, et al. Association between DNA double strand break gene Ku80 polymorphisms and oral cancer susceptibility. Oral Oncol. 2009;45:789–793. doi:10.1016/j.oraloncology.2008.12.002
25. Liu CJ, Hsia TC, Tsai RY, et al. The joint effect of hOGG1 single nucleotide polymorphism and smoking habit on lung cancer in Taiwan. Anticancer Res. 2010;30:4141–4145.
26. Bravard A, Vacher M, Moritz E, et al. Oxidation status of human OGG1-S326C polymorphic variant determines cellular DNA repair capacity. Cancer Res. 2009;69:3642–3649. doi:10.1158/0008-5472.CAN-08-3943
27. Zielinska A, Davies OT, Meldrum RA, Hodges NJ. Direct visualization of repair of oxidative damage by OGG1 in the nuclei of live cells. J Biochem Mol Toxicol. 2011;25:1–7. doi:10.1002/jbt.v25.1
28. Zhao H, Qin C, Yan F, et al. hOGG1 Ser326Cys polymorphism and renal cell carcinoma risk in a Chinese population. DNA Cell Biol. 2011;30:317–321. doi:10.1089/dna.2010.1135
29. Yuan W, Xu L, Feng Y, et al. The hOGG1 Ser326Cys polymorphism and breast cancer risk: a meta-analysis. Breast Cancer Res Treat. 2010;122:835–842. doi:10.1007/s10549-009-0722-5
30. Pawlowska E, Janik-Papis K, Rydzanicz M, et al. The Cys326 allele of the 8-oxoguanine DNA N-glycosylase 1 gene as a risk factor in smoking- and drinking-associated larynx cancer. Tohoku J Exp Med. 2009;219:269–275. doi:10.1620/tjem.219.269
31. Xing DY, Tan W, Song N, Lin DX. Ser326Cys polymorphism in hOGG1 gene and risk of esophageal cancer in a Chinese population. Int J Cancer. 2001;95:140–143. doi:10.1002/1097-0215(20010520)95:3<140:AID-IJC1024>3.0.CO;2-2
32. Nam RK, Zhang WW, Jewett MA, et al. The use of genetic markers to determine risk for prostate cancer at prostate biopsy. Clin Cancer Res. 2005;11:8391–8397. doi:10.1158/1078-0432.CCR-05-1226
33. Dhillon VS, Yeoh E, Fenech M. DNA repair gene polymorphisms and prostate cancer risk in South Australia–results of a pilot study. Urol Oncol. 2011;29:641–646. doi:10.1016/j.urolonc.2009.08.013
34. Li Q, Huang L, Rong L, et al. hOGG1 Ser326Cys polymorphism and risk of childhood acute lymphoblastic leukemia in a Chinese population. Cancer Sci. 2011;102:1123–1127. doi:10.1111/j.1349-7006.2011.01928.x
35. Ferguson HR, Wild CP, Anderson LA, et al. No association between hOGG1, XRCC1, and XPD polymorphisms and risk of reflux esophagitis, Barrett’s esophagus, or esophageal adenocarcinoma: results from the factors influencing the Barrett’s adenocarcinoma relationship case-control study. Cancer Epidemiol Biomarkers Prev. 2008;17:736–739. doi:10.1158/1055-9965.EPI-07-2832
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.