Back to Journals » Drug Design, Development and Therapy » Volume 16
Shaoyao Gancao Decoction Ameliorates Paclitaxel-Induced Peripheral Neuropathy via Suppressing TRPV1 and TLR4 Signaling Expression in Rats
Authors Chen Y, Lu R, Wang Y ![]() , Gan P
, Gan P
Received 4 February 2022
Accepted for publication 26 May 2022
Published 30 June 2022 Volume 2022:16 Pages 2067—2081
DOI https://doi.org/10.2147/DDDT.S357638
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Georgios Panos
Yu Chen,1 Ruohuang Lu,2 Yang Wang,3 Pingping Gan1
1Department of Oncology, Xiangya Hospital, Central South University, Changsha, People’s Republic of China; 2Department of Stomatology, Third Xiangya Hospital, Central South University, Changsha, People’s Republic of China; 3Department of Integrated Traditional Chinese and Western Medicine, Institute of Integrative Medicine, Xiangya Hospital, Central South University, Changsha, People’s Republic of China
Correspondence: Pingping Gan, Department of Oncology, Xiangya Hospital, Central South University, Changsha, People’s Republic of China, Tel +86 13874975101, Email [email protected]
Purpose: Paclitaxel-induced peripheral neuropathy (PIPN) is increasingly becoming one of the most widespread adverse effects in the treatment of cancer patients, and further precipitate neuroinflammation in the nervous system. Interestingly, Shaoyao Gancao Decoction (SGD), a traditional Chinese analgesic prescription, has emerged as a primary adjuvant to chemotherapy in relieving side effects, especially in the case of PIPN. However, the underlying mechanism of SGD functioning in PIPN remains elusive. Accordingly, the current study set out to explore the potential axis implicated in the functioning of SGD in PIPN.
Methods: First, network pharmacology was adopted to predict the role of the transient receptor potential vanilloid type 1 (TRPV1) protein in treating PIPN with SGD. Subsequently, the effects of SGD treatment on mechanical allodynia and thermal hyperalgesia were evaluated in rat PIPN models. Based on the bioinformatics information and current literature, paclitaxel activates toll-like receptor 4 (TLR4) induces the sensitization of TRPV1 mechanistically. Thereafter, TLR4-myeloid-differentiation response gene 88 (MyD88) signaling and TRPV1 expression patterns in dorsal root ganglias (DRGs) were measured by means of Western blotting, qPCR and immunofluorescence.
Results: Initial bioinformatics reared a total of 105 bioactive compounds and 1075 target genes from SGD. In addition, 40 target genes intersected with PIPN were considered as potential therapeutic genes. Based on the network analysis, SGD was found to exert its analgesic effect by reducing the expression of TRPV1. Further experimentation validated that SGD exerted an analgesic effect on thermal hyperalgesia in PIPN models, such that this protective effect was associated with the suppression of TRPV1 and TLR4-MyD88 Signaling over-expression.
Conclusion: Collectively, our findings indicated that SGD ameliorates PIPN by inhibiting the over-expression of TLR4-MyD88 Signaling and TRPV1, and further highlights the use of SGD as a potential alternative treatment for PIPN.
Keywords: paclitaxel-induced peripheral neuropathy, Shaoyao Gancao Decoction, transient receptor potential vanilloid type 1, network pharmacology, paclitaxel, toll-like receptor 4
Introduction
Cancerous malignant tumors are one of the most prevalent causes of disease and morbidity in urban populations, and further accompanied by an ever-increasing incidence.1 Despite the advent of novel therapies such as immunotherapy and targeted therapies, the use of chemotherapy still remains the gold-standard for most malignancies due to its high efficacy.2,3 Paclitaxel, a well-known anti-cancer agent produced from the Taxus brevifolia’s needles and bark, is extensively applied all over the world to treat a number of malignancies, including ovarian cancer, non-small cell lung cancer, breast cancer and other cancers.4–8 Paclitaxel stabilizes microtubules and reduces their dynamicity via binding to beta-tubulin, and the resulting microtubules exhibit resistance to depolymerization, which leads to mitotic arrest and consequently to tumor cell death.9,10 Although it is highly effective in blocking the proliferation of cancer cells, peripheral neuropathy is encountered as one of the most severe adverse effects of paclitaxel in a large proportion of patients undergoing chemotherapy.11 These patients often present with persistent changes in perception, such as numbness, tingling sensation, and sharp and burning pains.12 Unfortunately, to date, there are no effective regimens to successfully tackle paclitaxel-induced peripheral neuropathy.13 Moreover, single target drugs have previously been applied to treat PIPN, but to no avail. Accordingly, it is imperative to advance the search for novel therapeutic targets to reduce the plight of PIPN.
Shaoyao Gancao Decoction (SGD), one of the most celebrated Chinese herbal prescriptions, also known as Shakuyaku-Kanzo-to in Japan, is composed of Radix Paeoniae Alba (Paeonia lactiflora Pall,root) (PA) and Glycyrrhizae uralensis (Glycyrrhiza uralensis Fisch., root and rhizome, honeyed) (GU) at a ratio of 1:1.8,14,15 Historically, it was first reported in the Treatise on Cold Pathogenic and Miscellaneous Diseases, a traditional Chinese medical classic focusing on exogenous and internal diseases written by Zhang Zhongjing of the Han Dynasty.16 Currently, SGD is often prescribed for the treatment of patients with muscle spasm and gynecological disorders in Asia, owing to its analgesic and anti-inflammatory effects.17–19 Moreover, there is a plethora of evidence to certify the alleviatory effects of SGD on the pain symptoms associated with PIPN with almost negligible side effects.20 Meanwhile, SGD is well established to exert its effects in accordance with the multi-target and multi-pathways principle. However, almost no researchers have explored the underlying mechanism with regard to SGD allieviatory effect on PIPN.
The transient receptor potential vanilloid type 1 (TRPV1), a heat-activated cation channel with high permeability for Ca2+, is primarily expressed in the spinal cord and dorsal root ganglia neurons.21–23 Existing evidence suggests that TRPV1 can be activated by a wide range of mechanisms, such as heat, capsaicin, secondary messengers, reactive oxygen and nitrogen species.22,24 What is noteworthy is that, activation and sensitization of TRPV1 was previously associated with the paclitaxel treatment-induced enhancement of hyperalgesia.25 Moreover, selective blockade of TRPV1 channel with TRPV1 antagonist (AMG9810) is known to completely reverse paclitaxel-induced thermal hyperalgesia.26
The Toll-like receptor 4 protein (TLR4), a type of pattern-recognition receptor (PRR), is known to confer an indispensable role in the mediation of chronic itch and pain.27,28 The initiation and up-regulation of TLR4, combined with its downstream molecule MyD88, have also been previously documented in paclitaxel-related chemotherapy-induced peripheral neuropathy.29,30 Moreover, a prior study revealed that inhibiting or knocking out the expression of TLR4 exerts a diminishing effect on injury-induced neuropathy pain.29,31 Previous studies have also found that TRPV1 was sensitized via TLR4 after utilizing paclitaxel to activate peripheral sensory neurons.32,33 Thus, it could be possible that paclitaxel activates sensory neurons by sensitizing TLR4-MyD88 signaling and TRPV1 channel, inducing nociceptive hypersensitivity and consequently resulting in neuropathic pain. However, there is a lack of reliable evidence to substantiate the treatment functions of SGD on PIPN, and the relevant mechanism of the TRPV1 and TLR4-MyD88 signaling. Accordingly, the current study set sought to explore SGD’s analgesic functions and the underlying biomolecular mechanism in regard to PIPN, with the help of network analyses, animal models and biomolecular experiments (Figure 1).
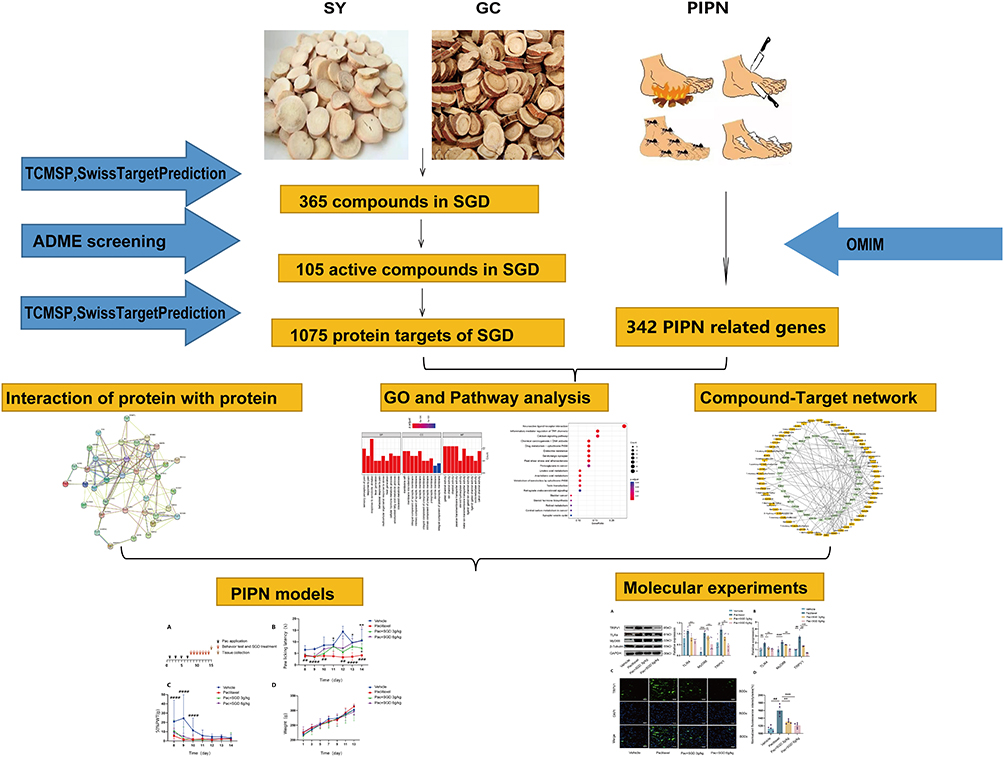 |
Figure 1 The workflow of the current study. Abbreviations: SY, Shaoyao; GC, Gancao; PIPN, paclitaxel-induced peripheral neuropathy; SGD, Shaoyao Gancao Decoction; TCMSP, praditional Chinese medicine systems pharmacology database; ADME, absorption, distribution, metabolism, and excretion; OMIM, Online Mendelian Inheritance in Man. |
Materials and Methods
Network Target Analysis
Searching Bioactive Ingredients in SGD
The traditional Chinese medicine systems pharmacology (TCMSP) database (http://tcmspw.com/) was adopted to retrieve and collect bioactive elements of SGD.34 Ingredients were then filtered using the following selection criteria: oral bioavailability (OB) threshold ≥30% and drug-likeness (DL) threshold ≥0.18.
Protein Target Genes in Databases
The SwissTargetPrediction database (http://www.swisstargetprediction.ch/) and TCMSP database (http://tcmspw.com/) were utilized to screen SGD protein targets. Related target genes of PIPN were additionally obtained from the OMIM therapeutic database (http://www.ncbi.nlm.nih.gov/omim/). In addition, pain-related target genes were collected from the GeneMANIA database (https://genemania.org/), using the following keywords: paclitaxel-induced peripheral neuropathy, paclitaxel-induced peripheral neuropathic pain and paclitaxel-induced neuropathic pain. The UniProt database (https://www.uniprot.org/) was then adopted to standardize all the aforementioned target names. Intersection genes (ie repetitive target genes) regarding SGD treatment for PIPN were finally obtained using the Venny 2.1 tool (https://bioinfogp.cnb.csic.es/tools/venny/). Venn diagram was established by Microsoft Office PowerPoint.
Construction of Protein–Protein Interaction (PPI) Network
In order to establish a protein–protein interaction (PPI) network, the STRING10.5 online database (https://string-db.org/cgi/help.pl?UserId=PWTj1MTAhQKH&sessionId=8wlBKy7kNz5I) was utilized to analyze PIPN-SGD-related target proteins. In the PPI network map, each node symbolized a protein gene and each edge denoted a functional link between potential target genes. The Cytoscape software was then adopted to analyze the network after importing target genes into it.
Establishment of Compound-Target Network
To explore the pharmacological mechanisms, a compound-target network was constructed between the 40 potential target genes and bioactive ingredients of SGD using the Cytoscape 3.7.2 software. Each node symbolized the target genes and compounds, and similarly, lines denoted the interactive association between the target genes and compounds in the compound-target network. Key compounds and target genes were subsequently selected using the network topology parameters.
GO and KEGG Pathway Enrichment Analysis
The R package (org.Hs.eg.db, version 3.8) was utilized to further explore the Gene Ontology (GO) and KEGG (Kyoto Encyclopedia of Genes and Genomes) databases. Terms presented with thresholds Count ≥2 and Expression Analysis Systematic Explorer (EASE) scores ≤0.05 were selected for further functional clustering.
Next, in order to further investigate the relative signaling pathway regarding TRPV1 and the target genes of pain, the GeneMANIA database was utilized for prediction. Additionally, KEGG pathway enrichment analysis was also performed.
Experiment for Verification
Chemical Procurement
Paclitaxel was procured from Hospira Australia Pty. Ltd. (Australia). Saline (0.9% NaCl injectable solution) was purchased from Kelun Pharm Co. Ltd. (Chengdu, China). Shaoyao (Paeoniae Radix Alba, the root of Paeonia lactiflora Pall.) and Gancao (Glycyrrhizae Radix et Rhizoma, the root and rhizome of Glycyrrhiza uralensis Fisch.) were obtained from the Xiangya Hospital of Central South University (Changsha, China). Prof. SY Hu from the department of Chinese Herbal Medicine of Central South University carried out the authentication of the acquired crude herbs. The voucher specimens (No. 21040908 and No. 21041608) were deposited at the Ethnopharmacology Laboratory of Xiangya Hospital.
Ethics Concerns
The current study was approved by the Medical Ethics Committee of Central South University (approval no. 2021sydw0146), and in accordance with guidance from Central South University for the use and care of animals. Extensive efforts were undertaken to minimize both the number and suffering from the experimental animals.
Animal Usage
Total 20 male Sprague–Dawley rats (5–8 weeks, weighing 270–300 g) were acquired from the Department of Laboratory Animal Research at Central South University. Every 3 rats were housed in laminar flow cages and sustained in conditions of 50%–60% relative humidity, 22±2°C temperature, under a 12-h light/12-h dark cycle as previously described.8 The rats were allowed ad libitum access to a standard chow diet and tap water. The rats were housed in the aforementioned cages for at least 3 days for acclimatization prior to experimentation.
PIPN Rat Model Establishment
Firstly, 6 mg/mL concentrated paclitaxel (Hospira Australia Pty. Ltd, Australia) was diluted by a concentration of 1 mg/mL with the addition of 0.9% saline. Subsequently, the rats were intraperitoneally administered diluted paclitaxel (a dosage of 2 mg/kg) every other day on days 1, 3, 5, and 7. Meanwhile, rats in the control group received only saline treatment of the same volumes.
Drug Administration
SGD preparation was carried out in accordance with previously published literature.8,35 Briefly, Baishao (100 g) and Gancao (100 g) were blended and distilled twice in water (at a ratio of 1:10, g/mL) for 1 h. The combination included the above two filtrates, and the concentration of SGD was determined to be 1g/mL. The decoction was stored at 4°C for subsequent oral gavage to rats. A total of 4 groups of rats (n = 5 for each treatment) were established using the random number table method. Two rat groups were administered SGD gavage (dosages of 3g/kg and 6g/kg) from day 8 to day 14, and for the control group and paclitaxel group, rats were allowed access to distilled water as control.
Mechanical Allodynia
The above-mentioned rats were habituated in acrylic transparent chambers one by one on a mesh floor, for at least 15 minutes prior to any experimentation until they were adapted to the new environment and stopped cage exploration. Next, to measure the mechanical allodynia, the mid-plantar surface of the hind paws of rats was prickled with von Frey filaments (Stoelting, IL, USA), with appropriate power bending the filament for 6–8 s slightly. Active responses included the following: a sharp withdrawal of the paw, licking and immediate flinching after stimulation.36 In order to calculate the threshold, the up-down paradigm was adopted, and based on the nonparametric Dixon test, 50% paw withdrawal threshold (PWT) was measured and recorded.36 During the whole behavior test, the experimenter was blinded to all of the rat groupings.
Thermal Hyperalgesia
The above-mentioned rats were habituated in acrylic transparent chambers one by one for at least 15 minutes prior to any experimentation. An Incremental Hot Cold Plate (IITC Life Science, Woodland Hills, CA, USA) was utilized to assess thermal hyperalgesia. Briefly, the rats were placed on the surface of the hot plate (heated to a temperature of 55 ± 2°C) one at a time. Positive responses included the following: jumping or licking the hind-paw. A 20-s cutoff threshold was set up to prevent tissue damage. A significant drop in paw licking latency was regarded as heat hyperalgesia. During the whole behavior test, the experimenter was blinded to all of the rat groupings. Each rat’s thermal latency was measured 3 times to obtain the mean value.
Tissue Collection
After the completion of behavioral testing, the rats were anesthetized with the administration of sodium pentobarbital (Sigma-Aldrich Co, St Louis, Mo, USA). Next, the rats were intracardially perfused with 4% paraformaldehyde. DRGs of Left and right L4-6 and segments of L4-6 spinal cord were immediately excised and stored in liquid nitrogen. The obtained tissue samples were maintained at −80°C prior to further experimentation. Later, the tissues were cryoprotected in 20% sucrose and 30% sucrose at 4°C for at least 48 h.
Quantitative RT-PCR Analysis
The TRIzol reagent (Invitrogen, Waltham, MA) was adopted for the extraction of total RNA content, and PrimeScript™ kits were (TaKaRa Bio Inc, Otsu, Japan) utilized for the synthesis of cDNA, in accordance with the manufacturer’s protocols. A SYBR Green fluorescence-based assay (TaKaRa Bio Inc, Otsu, Japan) was adopted to perform qRT-PCR on a ViiA™ 7 RT-PCR system (Applied Biosystems, Carlsbad, CA). The primers for real-time PCR were as follows: TRPV1: forward: CCAGTCAAGCCCCACATCTT; reverse: CCAGCTCCTGGCAGTTACTC. GAPDH: forward: TGTGGGCATCAATGGATTTGG; reverse: ACACCATGATGGGGGGGGGATCAAT. TLR4: forward: CCGCTCTGGCATCATCTTCA; reverse: TCCCACTCGAGGTAGGTGTT. MyD88: forward: TGTCTCCCCTGACATGCCTA; reverse: CTGGGGGCGGAATGTTTTTGGAPDH. GAPDH was adopted as the reference control. The relative mRNA expressions were calculated using the 2-ΔΔCt method.
Western Blot Assay
SGD and spinal cord samples were centrifuged and immersed in RIPA buffer (P0013B, Beyotime, Shanghai, China) containing protease inhibitor (Thermo Fisher Scientific, Waltham, MA, United States) for the extraction of total protein content. The preparations were collected and stored at −80°C. The equal amounts of protein (10–50 μg) were separated by means of 10% SDS-PAGE, and then transferred onto polyvinylidene fluoride (PVDF) membranes (Millipore, Bedford, MA). The membranes were subsequently blocked with 5% skim milk for 2 h at 37°C, and incubated with primary antibodies against TRPV1 (ab6166, Abcam, Cambridge, UK), TLR4 (66350, abclonal, Wuhan, China) and MyD88 (ab219413, Abcam, Cambridge, UK) overnight at 4°C. The following day, immunoreactive bands were incubated with a horseradish peroxidase (HRP)-conjugated secondary antibody (#7074 CST, MA, USA) for 30 minutes at room temperature. The ChemiDocXRS+ System (Bio-Rad, Hercules, CA) was utilized for imaging the bands, which were analyzed with the ImageJ software (NIH). The relative expression was calculated as the ratio of the intensity of the gene of interest to that of GAPDH (abs132004, absin, China) and β-Tubulin (#2148S CST, MA, USA).
Immunofluorescence Staining
A frozen microtome (Thermo NX50, MA, USA) was utilized to cut the spinal cord cross sections (thickness of 20 µm), and these sections were transferred onto gelatin-coated glass slides to prepare immunofluorescence staining. Following pre-incubation with 10% normal goat serum and 0.3% Triton X-100 in PBS for 1 h at room temperature, a primary antibody (rabbit anti-TRPV1, ab203103, Abcam, Cambridge, UK) was applied for 18 h at 4°C. Subsequently, the sections were rinsed with PBS thrice, and then incubated with a secondary antibody conjugated to Alexa fluor 488 (488 goat anti-rabbit antibody ab150077, Abcam, Cambridge, UK) for 1 h at 37 ◦C. Afterwards, the nuclei were stained with DAPI and visualized under a Leica DM5000 B microscope (Leica Microsystems, Buffalo Grove, IL, USA).
Statistical Analysis
Statistical analyses were performed using the GraphPad Prism 7.0 software. Non-normal data were presented as median and range (min–max). Normal data were presented in the form of mean ± SD. At first, Mauchly’s test of sphericity and QQ plots together with Shapiro–Wilk test were applied to assess data distribution. Behavioral data were analyzed with two-way ANOVA followed by Tukey’s post hoc test. Biomolecular experiment data were analyzed by Student’s t-test or Mann–Whitney U-test accordingly. A value of p < 0.05 was regarded as statistically significant.
Results
Network Target Analysis
Identification of the Active Compounds in SGD
A total of 365 compounds were obtained from the TCMSP and SwissTargetPrediction database, which included 85 compounds in BS and 280 compounds in GC after excluding duplicates. Among the 85 ingredients in BS, 13 active compounds met the selection criteria of OB ≥30% and DL ≥0.18. Meanwhile, among the 280 compounds in GC, 92 active compounds met the selection criteria of OB ≥30% and DL ≥0.18. Following ADME screening, altogether 105 components were selected as candidate bioactive components for further analyses.
Target Genes and PPI Network Map
A total of 342 PIPN target genes were obtained from the OMIM databases. Similarly, following analyses of the 105 active ingredients of SGD from the TCMSP and SwissTargetPrediction database, 1435 target genes were acquired. Subsequently, 1075 target genes were screened after duplicate removal. Next, a sum of 40 potential targets (Figure 2A, Table 1) were found at the intersection of PIPN target genes and SGD target genes using the Venny 2.1 software. Thereafter, a protein–protein interaction network was constructed (Figure 2B) to explore the relationship between the 342 PIPN target genes and 1075 target genes, which revealed that six genes are located in the core area. These genes include albumin (ALB), estrogen receptor (ESR1), interleukin-1 beta (IL1B), transient receptor potential cation channel subfamily V member 1 (TRPV1), prostaglandin E synthase 2 (PTGS2), as well as the high-affinity nerve growth factor receptor (NTRK1) gene.
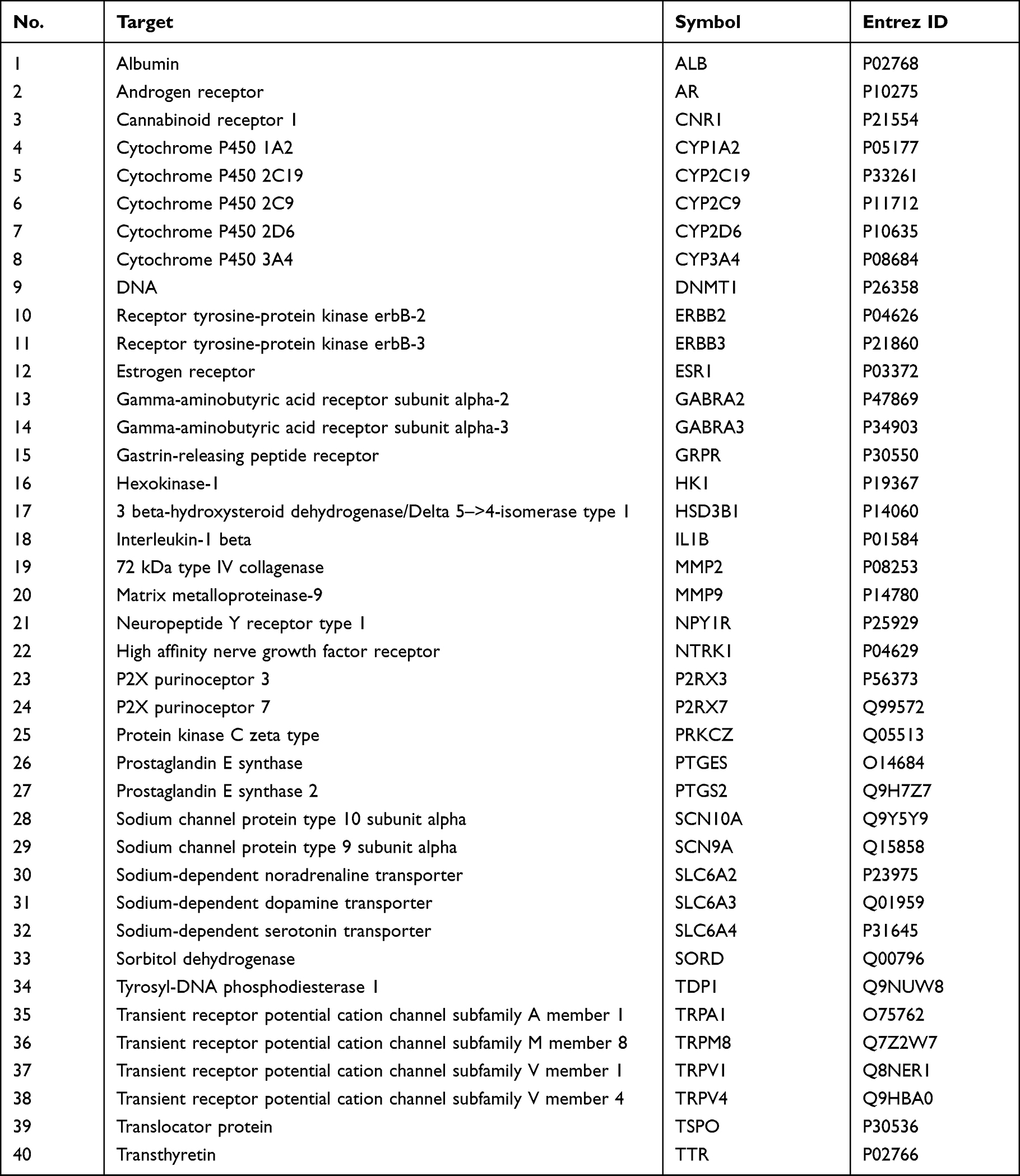 |
Table 1 40 Potential Target Gene of SGD Treatment for PIPN |
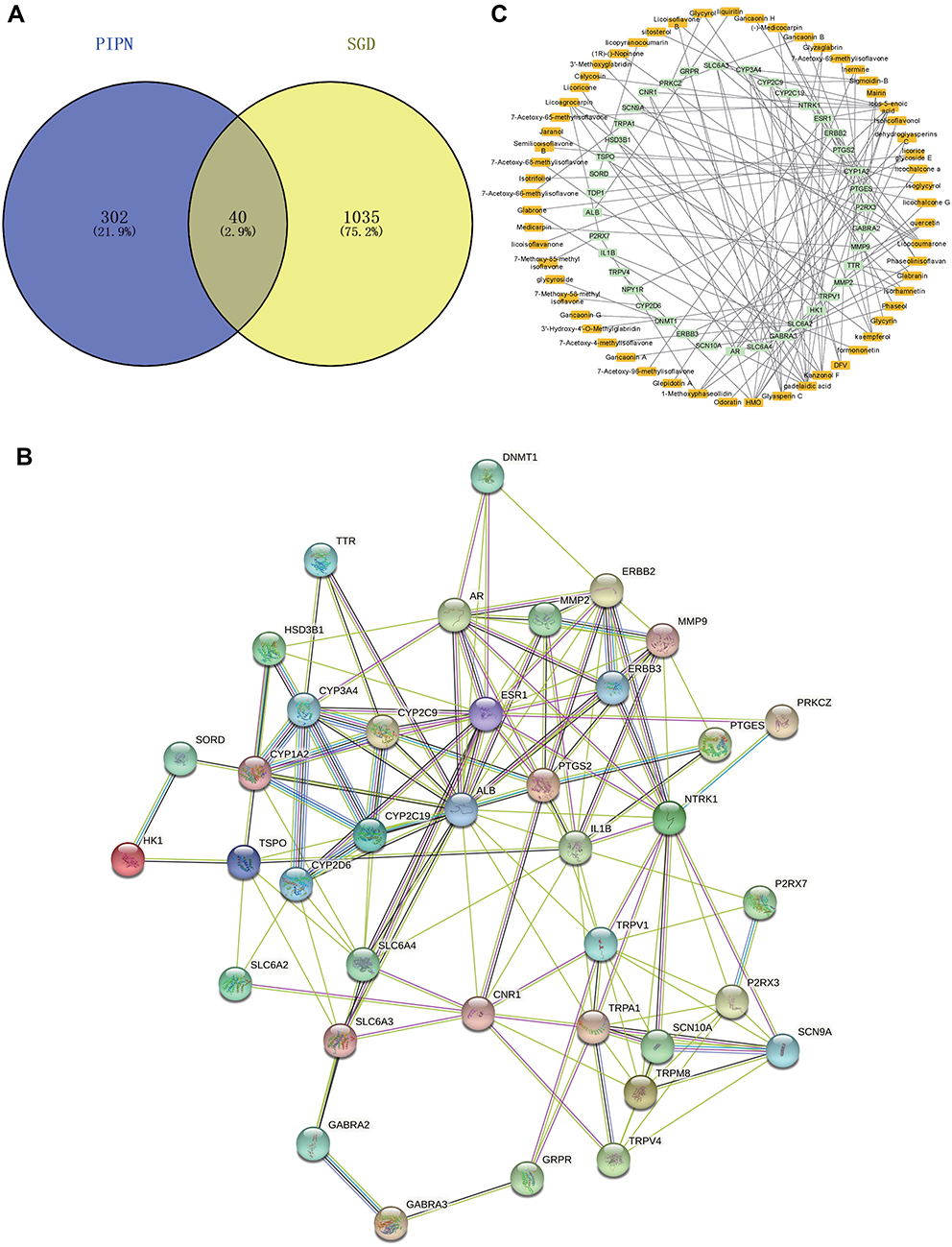 |
Figure 2 Potential target genes, compound-target network and PPI network map of SGD therapy for PIPN. (A) The venny results of potential target genes of SGD therapy for PIPN. (B) The PPI network map of 40 target genes. (C) The SGD-PIPN-potential target gene network. The orange nodes represent candidate active compounds and the green nodes represent potential protein targets. The edges indicate that each node is interconnected. Abbreviations: PIPN, paclitaxel-induced peripheral neuropathy; SGD, Shaoyao Gancao Decoction; PPI, protein–protein interaction. |
Compound-Target Network and Analysis
The 40 intersected genes and bioactive compounds were carried into the Cytoscape software (version 3.7.2) to acquire a compound-target network of SGD on PIPN (Figure 2C), which included 98 nodes and 129 edges. A single compound of SGD may interrelate with multiple targets, and a single target may relate to multiple compounds. There were a total of 14 high-degree target genes which were correlated with multiple compounds (degree >4) among the potential target genes, namely, ERBB2, CYP1A2, AR, ESR1, GABRA2, GABRA3, HK1, MMP2, MMP9, P2RX3, PTGES, PTGS2, SLC6A2, SLC6A4, TTR and TRPV1. In addition, among the 105 bioactive compounds, 4 high-degree compounds were found to be localized at the core position (degree >4), which included icos-5-enoic acid, gadelaidic acid, glyasperin C and kanzonol F.
GO and KEGG Pathway Enrichment Analysis
The R package for statistical computing was utilized to clarify the biological processes, cell composition and molecular function of the 40 intersected target proteins (Table 1). Subsequent results showed that there were 386 biological processes (BP), 40 cellular components (CC), and 84 molecular functions (MF) terms. The top 10 categories enriched from BP, CC, and MF are shown in Figure 3A, which implied that SGD may alleviate PIPN through sensory perception of pain, membrane raft, and gated channel activity.
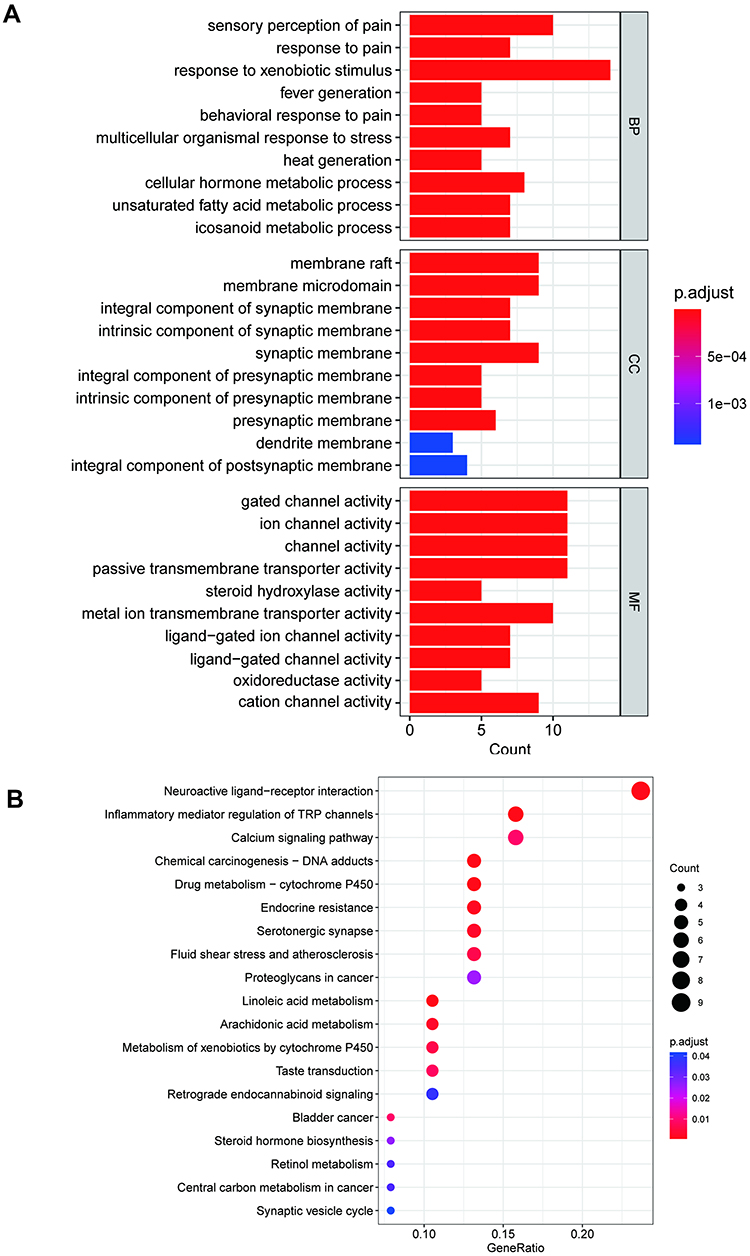 |
Figure 3 The 10 most significance genes ontology (A) and 20 most significance of pathway enrichment (B) analysis of therapy target genes of SGD on PIPN. Abbreviations: PIPN, paclitaxel-induced peripheral neuropathy; SGD, Shaoyao Gancao Decoction. |
KEGG pathway enrichment analysis was performed to further investigate the potential target genes and significant pathways. As shown in Figure 3B, 20 pathways correlated with 40 protein targets were enriched, including inflammatory mediator regulation of TRP channels, neuroactive ligand–receptor interaction, calcium signaling pathway, chemical carcinogenesis – DNA adducts and so on. The key translational implications indicated that the active ingredients of SGD may be useful in the prevention and treatment of PIPN via acting on these signaling pathways.
An additional KEGG pathway enrichment assay was carried out to further explore the relationship between TRPV1 and the target genes of pain (Supplementary Table 1). Among the 95 pathways, TLRs signaling pathway was found to be enriched with a relatively high gene count and P value.
Experiment for Verification
Shaoyao-Gancao Decoction Attenuated Thermal Hyperalgesia in PIPN Model
Paclitaxel treated rat models were first established to investigate the functional relationship between Shaoyao-gancao decoction and PIPN, following the protocols illustrated in Figure 4A. Rats were intraperitoneally administered with paclitaxel (at a dosage of 2 mg/kg for every two days) or vehicle (saline, same volumes and intervals as paclitaxel-injected rats). Prior to any Shaoyao-gancao treatments, the paclitaxel-treated rats exhibited a significant drop in the paw licking latency on the hot plate test (p < 0.05) and paw withdrawal threshold based on the Von Frey test (p < 0.05), relative to the vehicle-treated rats (Figure 4B and C). These findings substantiated the significant development of peripheral neuropathic hypersensitivity in paclitaxel-induced rat models.
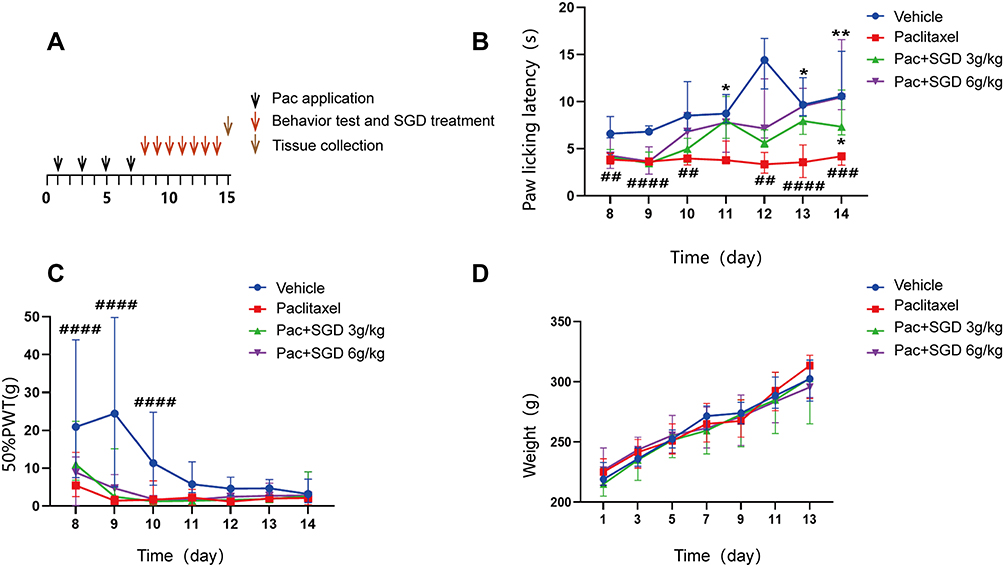 |
Figure 4 SGD attenuates thermal hyperalgesia in a PIPN model. (A) Experiment design for the construction of PIPN model and SGD treatment. (B) Reduction of paclitaxel-induced thermal hyperalgesia by administrating SGD for 7 consequent days. (C) Effects of SGD treatment on mechanical allodynia of PIPN rats. (D) Effects of paclitaxel injection and SGD treatment on body weight of rat models. Data were expressed as median and range (min - max). ## p < 0.01, ### p < 0.001, #### p < 0.0001 versus the vehicle group. * p < 0.05, ** p < 0.01 versus the paclitaxel group. n = 5 rats/group. Abbreviations: PIPN, paclitaxel-induced peripheral neuropathy; SGD, Shaoyao Gancao Decoction. |
Different concentrations of Shaoyao-gancao decoction were administered to the modelled rats starting from day 8, precisely, the day after the last injection of paclitaxel. SGD elicited significant and persistent relief from thermal hyperalgesia from day 8 compared to that produced by single paclitaxel treatment. As for the SGD 6g/kg group, the paw licking latency was increased from 3.59 ± 0.36 s at day 9 to 9.37 ± 0.77 s at day 14, compared with the SGD 3g/kg group, while the extent of increase was much lower relative to the high concentration of SGD group, suggesting that higher concentration of SGD exerted a more profound effect of relief from thermal hyperalgesia. However, the SGD’s effect of anti-allodynia was not mechanically observed until the end of the observation time after applying SGD for a week. Moreover, compared with the vehicle group, there was no change in the body weight of rats undergoing paclitaxel treatment (Figure 4D), and consecutive SGD administration exerted no influence on the body weight of rats either.
SGD Reversed the Up-Regulation of TLR4, MyD88 and TRPV1 in DRGs and Spinal Cord of PINP
The top three pathways in the KEGG analysis, namely, neuroactive ligand–receptor interaction, inflammatory mediator regulation of TRP channels, and calcium signaling pathway, are all associated with TRPV1. In addition, Yukako Kamata et al provided convincing evidence that increased expression levels of TRPV1 channel serve as a key factor in regulating paclitaxel-treated rodents.37 Additional KEGG analysis regarding TRPV1 and the target genes of pain revealed the involvement of the toll-like receptor signaling pathway. Moreover, Yan Li et al have previously demonstrated that for PIPN rats, the expression levels of TLR4 and its downstream signaling MyD88 and TRPV1 are both up-regulated in DRGs and spinal cord tissues.30,33 Furthermore, the peripheral neuropathy of PIPN rats is known to be reduced following blockade of TLR4-MyD88 signaling.31 Accordingly, we speculated whether SGD relieved PIPN by means of augmenting TRPV1 and TLR4-MyD88 signaling.
We further explored the molecular mechanisms of SGD’s pain-relief function in paclitaxel-induced peripheral neuropathic pain. First, we collected the L4-6 DRGs of rats, given the fact that paclitaxel accumulates in the DRGs of animal models because DRGs lack an effective blood–brain barrier.38 The results of Western blot and Quantitative Real-time PCR demonstrated the presence of increased expressions of TLR4, MyD88 and TRPV1 at the protein and mRNA levels (Figure 5A and B). These findings validated that TLR4, MyD88 and TRPV1 were all up-regulated in L4-6 DRGs following intraperitoneal paclitaxel administration in rats model. On the other hand, regular administration of SGD for one week exerted a counter effect on the aforementioned over-expression, wherein the SGD 6g/kg group exhibited better reversal effects compared to the SGD 3g/kg group.
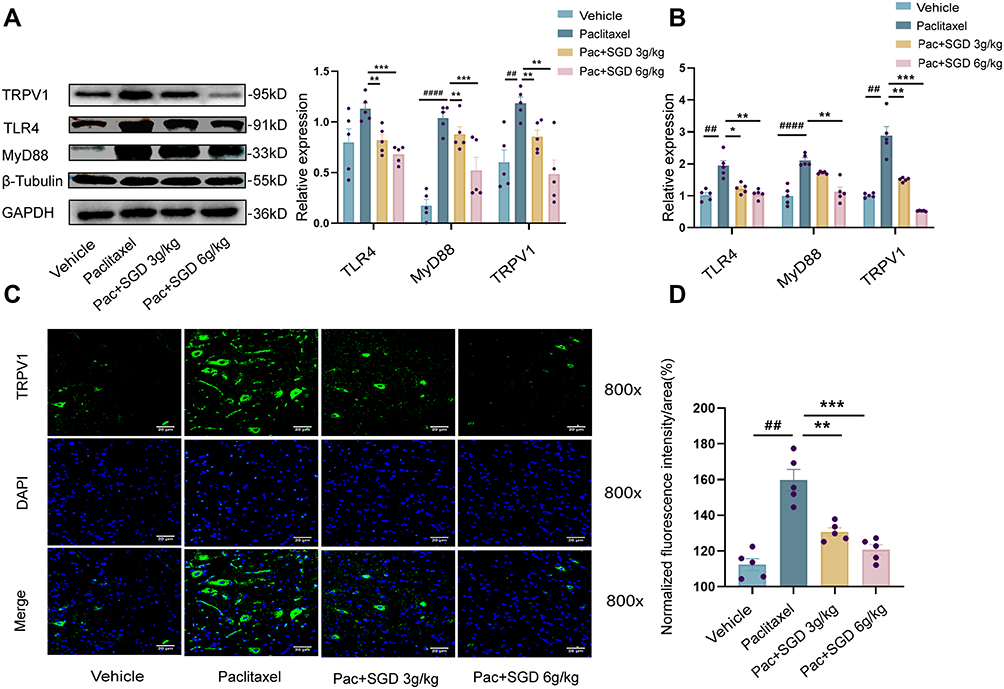 |
Figure 5 SGD reduced the over-expression of TLR4, MyD88 and TRPV1 in dorsal root ganglion (DRG) neurons or spinal cord from PIPN rats. (A) Proteins expression after repeat SGD treatment in PIPN model. The right bar graph shows the proteins expression normalized to GAPDH or β-tubulin. Representative protein images are presented on the left side. (B) mRNA expression after repeat SGD treatment in PIPN model. (C) Representative immunofluorescence image of TRPV1 and DAPI in vehicle, paclitaxel, paclitaxel + SGD 3g/kg, paclitaxel + SGD 6g/kg. The mean fluorescence intensity (D) was also measured. Data were expressed as mean ± SD. ## p < 0.01, ####p < 0.0001 versus the vehicle group. *p < 0.05, **p < 0.01, ***p < 0.001 versus the paclitaxel group. n = 5 rats/group. Abbreviations: PIPN, Paclitaxel-induced peripheral neuropathy; SGD, Shaoyao Gancao Decoction; TRPV1, transient receptor potential vanilloid type 1; TLR4, toll-like receptor 4; MyD88, myeloid-differentiation response gene 88; DRG, dorsal root ganglia; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; DAPI, 4’,6-diamidino-2-phenylindole. |
Existing literature further indicates that chemotherapies can also bring about the activation of astrocyte and microglia in the spinal cord dorsal horn.39–42 Consequently, immunofluorescence was carried out to observe the mean fluorescence intensity of TRPV1 immune positive (TRPV1+) neurons in the spinal cord. Subsequent results demonstrated that the TRPV1 protein was primarily expressed in the cytoplasm and cell membrane, while there was almost no distribution in the nucleus. As shown in Figure 5C and D, the mean fluorescence intensity of neurons was significantly extended in the paclitaxel-treated group. Importantly, the over-expression of TRPV1 induced by paclitaxel in spinal cord neurons exhibited a deficit response following oral administration of SGD (from 159.74 ± 11.96 to 130.71 ± 4.56, P < 0.01). Furthermore, the mean fluorescent intensity of TRPV1+ neurons showed a significant decrease in SGD 6g/kg compared to SGD 3g/kg group. In summary, SGD alleviates PIPN by reducing the expression and activity of TRPV1 and TLR4-MyD88 signaling. The application of SGD may be a promising therapeutic agent for pain control.
Discussion
Paclitaxel-induced peripheral neuropathy (PIPN) is one of the major side effects faced in the treatment of tumor patients.38 A traditional Chinese medicine concoction, SGD exerts a number of polypharmacological effects based on the ‘multi-component, multi-target and multi-pathways’ principle,43 while exhibiting better clinical efficacy and negligible side effects compared to traditional analgesic medications.17 In an effort to further elucidate the analgesic mechanism of SGD, we employed a series of network pharmacology approaches to predict the mechanism of SGD in alleviation of PIPN.
First, in the complex system of SGD, a total of 105 bioactive compounds and 40 potential protein targets of SGD on PIPN were analyzed based on network pharmacology. Subsequently, SGD-PIPN-potential target gene interaction and compound-target networks were constructed using 40 protein targets retrieved from three databases. As documented in the SGD-PIPN interaction network, the 40 potential target genes were not independent, but exhibited linkage and interaction with each other. Additionally, we came across potential protein targets with high degree nodes occupying hub positions in the network. Subsequent analyses of the compound-target network illustrated that compounds with higher-degree were localized at the core sites. These targets and compounds exhibited a higher degree in this process, which suggested that they may confer an essential role in the meditation and treatment of PIPN, and potentially account for the therapeutic effects of SGD treatment on PIPN. Thereafter, 40 target genes were screened with the help of GO and KEGG enrichment analyses. Moreover, subsequent results from the GO and KEGG analyses indicated that the occurrence and development of PIPN were directly associated with 386 biological functions and 20 signaling pathways, underscoring that the bioactive functions and signaling pathways of these genes may function as the potential mechanism of SGD in treating PIPN.
The hard-done work of our peers has further shown that the up-regulation and sensitization of TRPV1 in peripheral sensory neurons serve as the mechanism of chronic pain, including the pain induced by paclitaxel.26,33 Herein, our network analyses revealed that TRPV1 occupied the core position in the PPI and compound-target network. Meanwhile, the results of KEGG analysis demonstrated that all the top three pathways, namely neuroactive ligand–receptor interaction, inflammatory mediator regulation of TRP channels, and calcium signaling pathway, were associated with TRPV1. In lieu of these findings, we speculated that SGD may exert its analgesic effect on PIPN via regulation of the over-expression of TRPV1.
To ascertain our speculation, we further established paclitaxel-related PIPN rat models and investigated the pain-reliving function of SGD on paclitaxel-related hypersensitivities. Subsequent experimentation revealed that SGD exerted a diminishing effect on thermal hyperalgesia in PINP models. Compared with the SGD 3g/kg dosage group, the high concentration SGD group (6 g/kg) achieved a better effect in relieving thermal hyperalgesia. Interestingly, the effect of SGD on alleviating mechanical hypersensitivities was not observed. This can be explained by the fact that TRPV1 is a detector of noxious heat.44 Thus, it does not have high sensitivity to mechanical pain.45 Moreover, repeat applications of paclitaxel and SGD are invasive stimulation, which may construct conditioned reflex regarding painful stimulation in rats.
Existing evidence indicates that TRPV1 is regulated by a number of mechanisms.46,47 To further investigate the underlying mechanism of TRPV1 regulation, we performed another KEGG pathway enrichment to analyze TRPV1 and the target genes of pain. We observed that TLRs signaling was enriched with a relatively high gene count and P value. Interestingly, toll-like receptors (TLRs) have been previously shown to be associated with neuropathic pain.48,49 Yan Li et al further illustrated that administration of paclitaxel can bring about an abrupt sensitization of TRPV1 via the TLR4-MyD88 signaling pathway.33 Also, in accordance with our findings, the study carried out by Yuanyuan Li et al reported that EA alleviates PIPN via mechanisms involving suppressing the over-expression of TLR4 signaling and TRPV1.50 In light of the above-mentioned findings, we chose to further explore the significance of TLR4 and MyD88 expressions in regard to SGD’ allieviation of PIPN.
Additional results of molecular biological validation demonstrated that TRPV1 and TLR4-MyD88 signaling were both highly expressed in DRGs of PIPN rats, whereas SGD administration brought about marked attenuation of the over-expression of TRPV1 and TLR4-MyD88 signaling, wherein high concentration of SGD (6g/kg) achieved a better effect compared with low concentration SGD (3g/kg). Moreover, recent findings have come to light illustrating the involvement of the up-regulated TLR4/PI3K/Akt/TRPV1 signalling pathway in paclitaxel-mediated adverse effects and PIPN in rodents.51,52 Meanwhile, existing evidence further suggests that peripheral neuropathy can be reversed with the use of a selective PI3K-signalling inhibitor.51 In addition, the study performed by Zhuangpeng Chang et al discovered that SGD administration could reduce the inflammation state by down-regulating TLR4/PI3K/Akt signalling, which is in line with our findings.53 Taking these findings and data into consideration, it would be plausible to suggest that there are certain correlations between TLR4-PI3K signaling and TRPV1, and TLR4-PI3K signaling maybe the upstream of TRPV1.
Our study was the first-of-its-kind to explore the potential mechanism of SGD in treating PIPN and unveiled the mechanism wherein SGD exerts its analgesic effect by inhibiting the high expression of TRPV1 and TLR4-MyD88 signaling. However, how SGD regulates the TLR4/PI3K/Akt/TRPV1 signalling is still unclear, and warrants further exploration.
Conclusions
Collectively, our findings highlighted that SGD treatment relieves paclitaxel-induced chronic pain in rat models. Mechanistic data further indicated that SGD reverses the expressions of TRPV1 and TLR4-MyD88 signaling in DRGs of paclitaxel-treated rats, underscoring the use of SGD as a potential treatment for PIPN patients in clinics.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
2. Pilkington G, Boland A, Brown T, Oyee J, Bagust A, Dickson R. A systematic review of the clinical effectiveness of first-line chemotherapy for adult patients with locally advanced or metastatic non-small cell lung cancer. Thorax. 2015;70(4):359–367. doi:10.1136/thoraxjnl-2014-205914
3. Atkins S, He F. Chemotherapy and beyond infections in the era of old and new treatments’ for hematologic malignancies. Infect Dis Clin North Am. 2019;33(2):289. doi:10.1016/j.idc.2019.01.001
4. Crown J, Dieras V, Kaufmann M, et al. Chemotherapy for metastatic breast cancer - report of a European expert panel. Lancet Oncol. 2002;3(12):719–727. doi:10.1016/S1470-2045(02)00927-0
5. Zhu L, Chen L. Progress in research on paclitaxel and tumor immunotherapy. Cell Mol Biol Lett. 2019;24(1):40. doi:10.1186/s11658-019-0164-y
6. Hanna NH, Temin S, Masters G. Therapy for stage IV non-small-cell lung cancer without driver alterations: ASCO and OH (CCO) joint guideline update summary. J Oncol Pract. 2020;16(8):E844–E848. doi:10.1200/JOP.19.00770
7. Yang YH, Mao JW, Tan XL. Research progress on the source, production, and anti-cancer mechanisms of paclitaxel. Chin J Nat Med. 2020;18(12):890–897. doi:10.1016/S1875-5364(20)60032-2
8. Wang Y, Huang X, Zhong MZ, et al. Pretreatment of shaoyao gancao decoction () alters pharmacokinetics of intravenous paclitaxel in rats. Chin J Integr Med. 2017;23(1):70–75. doi:10.1007/s11655-016-2619-6
9. Xiao H, Verdier-Pinard P, Fernandez-Fuentes N, et al. Insights into the mechanism of microtubule stabilization by taxol. Proc Natl Acad Sci USA. 2006;103(27):10166–10173. doi:10.1073/pnas.0603704103
10. Weaver BA. How taxol/paclitaxel kills cancer cells. Mol Biol Cell. 2014;25(18):2677–2681. doi:10.1091/mbc.e14-04-0916
11. Argyriou AA, Bruna J, Mantovani E, Tamburin S. Neuromuscular complications of cancer therapy. Curr Opin Neurol. 2021;34(5):658–668. doi:10.1097/WCO.0000000000000969
12. Boyette-Davis J, Xin W, Zhang H, Dougherty PM. Intraepidermal nerve fiber loss corresponds to the development of taxol-induced hyperalgesia and can be prevented by treatment with minocycline. Pain. 2011;152(2):308–313. doi:10.1016/j.pain.2010.10.030
13. St Germain DC, O’Mara AM, Robinson JL, Torres AD, Minasian LM. Chemotherapy-induced peripheral neuropathy: identifying the research gaps and associated changes to clinical trial design. Cancer. 2020;126(20):4602–4613. doi:10.1002/cncr.33108
14. Shao YY, Chang ZP, Cheng Y, et al. Shaoyao-gancao decoction alleviated hyperandrogenism in a letrozole-induced rat model of polycystic ovary syndrome by inhibition of NF-κB activation. Biosci Rep. 2019;39(1). doi:10.1042/BSR20181877.
15. Shao YY, Guo YT, Gao JP, et al. Shaoyao-gancao decoction relieves visceral hyperalgesia in TNBS-induced postinflammatory irritable bowel syndrome via inactivating transient receptor potential vanilloid type 1 and reducing serotonin synthesis. Evid Based Complement Alternat Med. 2020;2020:7830280. doi:10.1155/2020/7830280
16. Li M, Dan W, Zhang H, Yun Y, He Q. Xiao Chai Hu Tang for peptic ulcers: a systematic review and meta-analysis of randomized controlled trials. Evid Based Complement Alternat Med. 2021;2021:6693677. doi:10.1155/2021/6693677
17. Bi XL, Gong MR, Di LQ. Review on prescription compatibility of shaoyao gancao decoction and reflection on pharmacokinetic compatibility mechanism of traditional Chinese medicine prescription based on in vivo drug interaction of main efficacious components. Evid-Based Complement Altern Med. 2014;2014:1–8. doi:10.1155/2014/208129
18. Tao WY, Wang LY. Mechanism of shaoyao gancao decoction on anti-inflammatory and analgesic effects by molecular docking.
19. Zhu N, Hou J, Ma G, Liu J. Network pharmacology identifies the mechanisms of action of shaoyao gancao decoction in the treatment of osteoarthritis. Med Sci Monit. 2019;25:6051–6073. doi:10.12659/MSM.915821
20. Staff NP, Grisold A, Grisold W, Windebank AJ. Chemotherapy-induced peripheral neuropathy: a current review. Ann Neurol. 2017;81(6):772–781. doi:10.1002/ana.24951
21. Basbaum AI, Bautista DM, Scherrer G, Julius D. Cellular and molecular mechanisms of pain. Cell. 2009;139(2):267–284. doi:10.1016/j.cell.2009.09.028
22. Davis JB, Gray J, Gunthorpe MJ, et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature. 2000;405(6783):183–187. doi:10.1038/35012076
23. Woodbury CJ, Zwick M, Wang SY, et al. Nociceptors lacking TRPV1 and TRPV2 have normal heat responses. J Neurosci. 2004;24(28):6410–6415. doi:10.1523/JNEUROSCI.1421-04.2004
24. Duitama M, Vargas-López V, Casas Z, Albarracin SL, Sutachan JJ, Torres YP. TRP channels role in pain associated with neurodegenerative diseases. Front Neurosci. 2020;14:782. doi:10.3389/fnins.2020.00782
25. Morelli MB, Amantini C, Liberati S, Santoni M, Nabissi M. TRP channels: new potential therapeutic approaches in CNS neuropathies. CNS Neurol Disord Drug Targets. 2013;12(2):274–293. doi:10.2174/18715273113129990056
26. Kawashiri T, Inoue M, Mori K, et al. Preclinical and clinical evidence of therapeutic agents for paclitaxel-induced peripheral neuropathy. Int J Mol Sci. 2021;22(16):8733.
27. Liu T, Gao YJ, Ji RR. Emerging role of Toll-like receptors in the control of pain and itch. Neurosci Bull. 2012;28(2):131–144. doi:10.1007/s12264-012-1219-5
28. Donnelly CR, Chen O, Ji RR. How do sensory neurons sense danger signals? Trends Neurosci. 2020;43(10):822–838. doi:10.1016/j.tins.2020.07.008
29. Li Y, Zhang HJ, Zhang HM, Kosturakis AK, Jawad AB, Dougherty PM. Toll-like receptor 4 signaling contributes to paclitaxel-induced peripheral neuropathy. J Pain. 2014;15(7):712–725. doi:10.1016/j.jpain.2014.04.001
30. Li Y, Zhang HM, Kosturakis AK, et al. MAPK signaling downstream to TLR4 contributes to paclitaxel-induced peripheral neuropathy. Brain Behav Immun. 2015;49:255–266. doi:10.1016/j.bbi.2015.06.003
31. Liu XJ, Zhang Y, Liu T, et al. Nociceptive neurons regulate innate and adaptive immunity and neuropathic pain through MyD88 adapter. Cell Res. 2014;24(11):1374–1377. doi:10.1038/cr.2014.106
32. Yu L, Yang F, Luo H, et al. The role of TRPV1 in different subtypes of dorsal root ganglion neurons in rat chronic inflammatory nociception induced by complete Freund’s adjuvant. Mol Pain. 2008;4:1744–8069. doi:10.1186/1744-8069-4-61.
33. Li Y, Adamek P, Zhang HJ, et al. The cancer chemotherapeutic paclitaxel increases human and rodent sensory neuron responses to TRPV1 by activation of TLR4. J Neurosci. 2015;35(39):13487–13500. doi:10.1523/JNEUROSCI.1956-15.2015
34. Ru JL, Li P, Wang JN, et al. TCMSP: a database of systems pharmacology for drug discovery from herbal medicines. J Cheminform. 2014;6(1):6. doi:10.1186/1758-2946-6-6
35. Feng D, Tang T, Fan R, et al. Gancao (Glycyrrhizae radix) provides the main contribution to shaoyao-gancao decoction on enhancements of CYP3A4 and MDR1 expression via pregnane X receptor pathway in vitro. BMC Complement Altern Med. 2018;18(1):345. doi:10.1186/s12906-018-2402-7
36. Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods. 1994;53(1):55–63. doi:10.1016/0165-0270(94)90144-9
37. Kamata Y, Kambe T, Chiba T, et al. Paclitaxel induces upregulation of transient receptor potential vanilloid 1 expression in the rat spinal cord. Int J Mol Sci. 2020;21(12):4341. doi:10.3390/ijms21124341
38. Yamamoto S, Egashira N. Drug repositioning for the prevention and treatment of chemotherapy-induced peripheral neuropathy: a mechanism- and screening-based strategy. Front Pharmacol. 2021;11. doi:10.3389/fphar.2020.607780
39. Peters CM, Jimenez-Andrade JM, Kuskowski MA, Ghilardi JR, Mantyh PW. An evolving cellular pathology occurs in dorsal root ganglia, peripheral nerve and spinal cord following intravenous administration of paclitaxel in the rat. Brain Res. 2007;1168:46–59. doi:10.1016/j.brainres.2007.06.066
40. Allen DT, Kiernan JA. Permeation of proteins from the blood into peripheral nerves and ganglia. Neuroscience. 1994;59(3):755–764. doi:10.1016/0306-4522(94)90192-9
41. Strumberg D, Brugge S, Korn MW, et al. Evaluation of long-term toxicity in patients after cisplatin-based chemotherapy for non-seminomatous testicular cancer. Ann Oncol. 2002;13(2):229–236. doi:10.1093/annonc/mdf058
42. Jaggi AS, Singh N. Mechanisms in cancer-chemotherapeutic drugs-induced peripheral neuropathy. Toxicology. 2012;291(1–3):1–9. doi:10.1016/j.tox.2011.10.019
43. Kuenzi BM, Rix LLR, Stewart PA, et al. Polypharmacology-based ceritinib repurposing using integrated functional proteomics. Nat Chem Biol. 2017;13(12):
44. Joseph J, Wang S, Lee J, Ro JY, Chung MK. Carboxyl-terminal domain of transient receptor potential vanilloid 1 contains distinct segments differentially involved in capsaicin- and heat-induced desensitization. J Biol Chem. 2013;288(50):35690–35702. doi:10.1074/jbc.M113.513374
45. Ma WY, Quirion R. Inflammatory mediators modulating the transient receptor potential vanilloid 1 receptor: therapeutic targets to treat inflammatory and neuropathic pain. Expert Opin Ther Targets. 2007;11(3):307–320. doi:10.1517/14728222.11.3.307
46. Jeske NA, Diogenes A, Ruparel NB, et al. A-kinase anchoring protein mediates TRPV1 thermal hyperalgesia through PKA phosphorylation of TRPV1. Pain. 2008;138(3):604–616. doi:10.1016/j.pain.2008.02.022
47. Zheng W, Wen H. Heat activation mechanism of TRPV1: new insights from molecular dynamics simulation. Temperature. 2019;6(2):120–131. doi:10.1080/23328940.2019.1578634
48. Jia Z-J, Wu F-X, Huang Q-H, Liu J-M. Toll-like receptor 4: the potential therapeutic target for neuropathic pain. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2012;34(2):168–173. doi:10.3881/j.issn.1000-503X.2012.02.013
49. Nicotra L, Loram LC, Watkins LR, Hutchinson MR. Toll-like receptors in chronic pain. Exp Neurol. 2012;234(2):316–329. doi:10.1016/j.expneurol.2011.09.038
50. Li YY, Yin CY, Li XJ, et al. Electroacupuncture alleviates paclitaxel-induced peripheral neuropathic pain in rats via suppressing TLR4 signaling and TRPV1 upregulation in sensory neurons. Int J Mol Sci. 2019;20(23):5917. doi:10.3390/ijms20235917
51. Adamek P, Heles M, Bhattacharyya A, Pontearso M, Slepicka J, Palecek J. Dual PI3Kδ/γ inhibitor duvelisib prevents development of neuropathic pain in model of paclitaxel-induced peripheral neuropathy. J Neurosci. 2022;42(9):1864–1881. doi:10.1523/JNEUROSCI.1324-21.2021
52. Adamek P, Heles M, Palecek J. Mechanical allodynia and enhanced responses to capsaicin are mediated by PI3K in a paclitaxel model of peripheral neuropathy. Neuropharmacology. 2019;146:163–174. doi:10.1016/j.neuropharm.2018.11.027
53. Chang ZP, Deng GF, Shao YY, et al. Shaoyao-gancao decoction ameliorates the inflammation state in polycystic ovary syndrome rats via remodeling gut microbiota and suppressing the TLR4/NF-κB pathway. Front Pharmacol. 2021;12:670054. doi:10.3389/fphar.2021.670054
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.