")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 16
Serotonin Type 6 and 7 Receptors as a Novel Therapeutic Target for the Treatment of Schizophrenia
Authors Tsegay EW , Demise DG , Hailu NA, Gufue ZH
Received 27 May 2020
Accepted for publication 30 September 2020
Published 28 October 2020 Volume 2020:16 Pages 2499—2509
DOI https://doi.org/10.2147/NDT.S263424
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Etsay Weldekidan Tsegay,1 Desalegn Getnet Demise,1 Nigus Alemu Hailu,2 Zenawi Hagos Gufue3
1Department of Pharmacy, College of Medicine and Health Sciences, Adigrat University, Adigrat, Tigray, Ethiopia; 2Department of Biomedical Science, College of Medicine and Health Sciences, Adigrat University, Adigrat, Tigray, Ethiopia; 3Department of Public Health, College of Medicine and Health Sciences, Adigrat University, Adigrat, Tigray, Ethiopia
Correspondence: Etsay Weldekidan Tsegay
Department of Pharmacy, College of Medicine and Health Sciences, Adigrat University, PO Box 50, Adigrat, Ethiopia
Tel +251-929263029
Email [email protected]
Abstract: Schizophrenia is a serious disease of the central nervous system that affects a person’s ability to think, feel and behave clearly. Even though the pathophysiological hypothesis of the disease is not clearly understood, dysfunction of dopamine, glutamate, serotonin and other neurotransmitters is widely believed to be involved. Serotonin within the synaptic vesicles functions as neurotransmitter and neurohormone in regulation of emotion, learning, memory, hormone release, cognition and motor function. Dysfunction of normal brain activity of serotonin is associated with schizophrenia. The role of serotonin 6 and 7 receptors in schizophrenia, interaction with neurotransmitters and the effect of drugs on those receptors in schizophrenia are the goal of this review. The aim of this review was to provide information for researchers and other scholars to identify the possible intervention points in the management of schizophrenia. The serotonin 6 and 7 receptors are associated with schizophrenia via modulating cyclic adenosine monophosphate, regulation of Fyn kinase and induction of structural plasticity. The above modulatory effects affect cholinergic, dopaminergic, glutamatergic, adrenergic and GABAergic systems. Recently, diverse numbers of selective agonist and antagonist ligands were developed for both receptors. SGS-518, ABT-354, Lu AE58054, SB-742,457, S-518, AVN-211, AVN-322, SYN-114 and SYN-120 are serotonin 6 receptor antagonists and aripiprazole-controlled release serotonin 7 receptor agonists under clinical trial for schizophrenia. Thus, research on novel drugs that act on serotonin 6 and 7 receptors likely facilitates the intervention into schizophrenia patients seeking better quality of life in the future.
Keywords: serotonin 6 receptor, serotonin 7 receptor, agonist, antagonist, schizophrenia
Introduction
Normal and impaired memory uses different signalling mechanisms with a variety of neurotransmitters. One of those is serotonin (5-HT), which is synthesized from tryptophan in the cell bodies of the dorsal raphe nucleus of brain stem.1
Serotonin is transported from the cytoplasm into synaptic vesicles to function as a neurotransmitter and neurohormone2 in regulation of emotion, learning, memory,3 mood, nociception, feeding, hormone release,4 cognition,5 sexual behaviour, circadian rhythm6 and motor function.7 In addition it mediates a variety of peripheral activities like gastrointestinal motility, inflammatory response and nociception modulation.8
Dysfunction of normal brain activity of 5-HT is associated with schizophrenia,9 autism,10 Alzheimer’s diseases,11 depression,12 anxiety,13 drug addiction14 and obesity.15 This paper will try to summarize the possible connection of 5-HT6 and 5-HT7 receptors with schizophrenia, interaction with inhibitory and excitatory neurotransmitters and progress in ligands that targeted those receptors, especially chemicals under preclinical and clinical trials.
Serotonin Receptors
Based on their structural, biochemical and pharmacological differences serotonin receptors (5-HTRs) have seven distinct receptor families with more than twenty subpopulations (Table 1).16 All serotonin receptors are G-protein-coupled receptors (GPCRs) with the exception of 5-HT3R, which is ligand-gated ion channel.2,17
 |
Table 1 Serotonin 6 and 7 Receptors Distribution, Mechanism of Action and Potential Effect80,81 |
Even though 5-HT6 and 5-HT7 receptors are less extensively studied receptors because of their more recent identification, selective agonists and antagonists (Table 2) along with 5-HT 6 and 7 receptor knock-out mice have revealed the involvement of those receptors in central nervous system activities.18 Additionally, their localization in brain areas associated with memory, locomotion and behaviour and their higher affinity to second generation antipsychotics and various antidepressants suggest a possible connection between those receptors and particular psychiatric disorders like schizophrenia.19
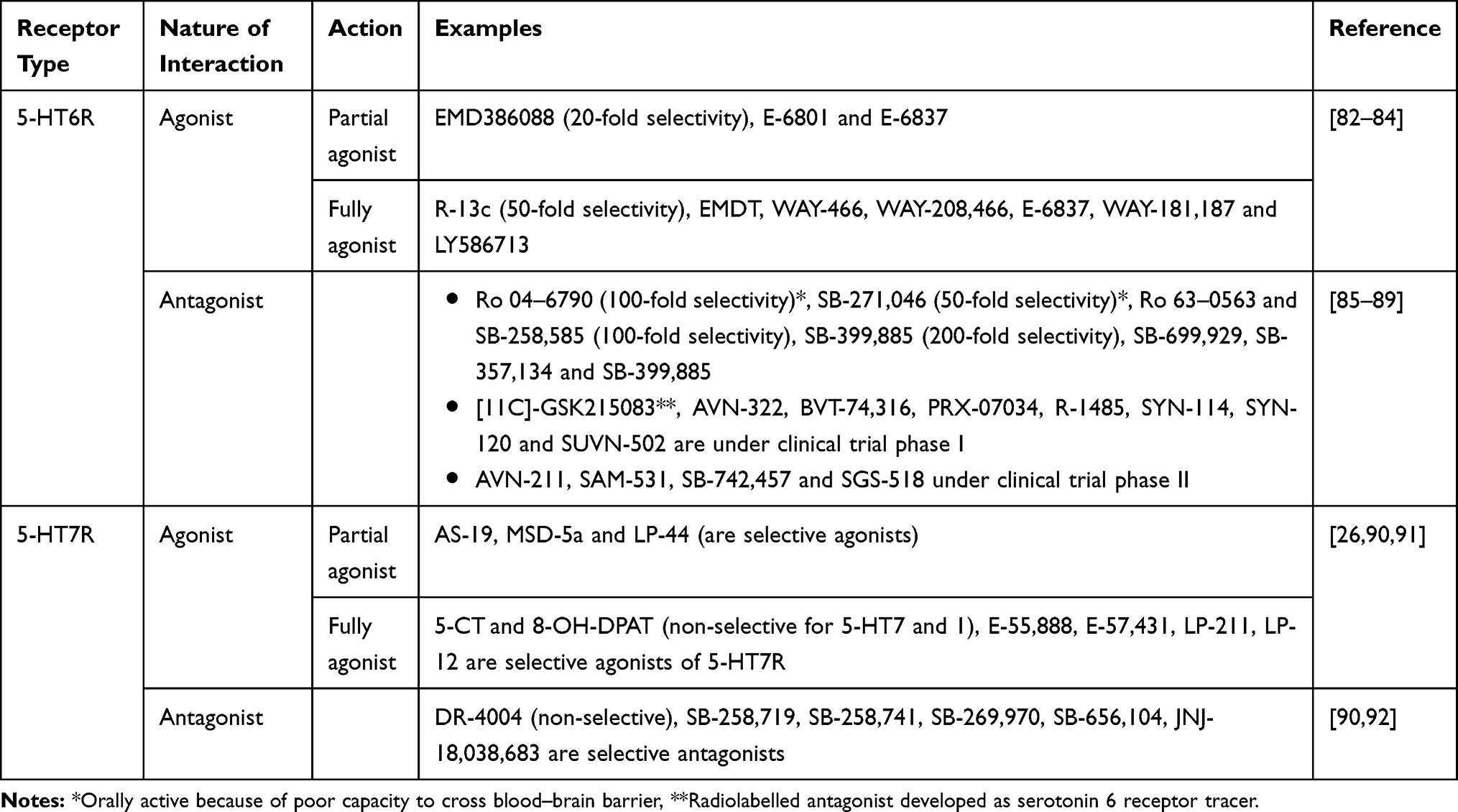 |
Table 2 Agonist and Antagonist Serotonin 6 and 7 Receptor Ligands |
Distribution of Serotonin 6 and 7 Receptors
Data from in situ hybridization indicate exclusive 5-HT6Rs expression within brain mainly in diencephalon, striatum, nucleus accumbens, olfactory tubercle and cortex and moderately expressed in the amygdala, hypothalamus, thalamus and cerebellum.20,21 In situ and northern blot analysis shows 5-HT7Rs distribution in human brain in thalamus, hypothalamus, hippocampus and cortex, and this is in good correlation with distribution in rodents.22
The above brain areas with higher distribution of 5-HT 6 and 7 receptors are associated with schizophrenia, as well as playing an important role in working memory and cognitive flexibility thought to be fundamental in schizophrenia.19
Methods
A reference database was collected from PubMed and Google scholar with the following search terms: serotonin 6 receptors AND schizophrenia, serotonin 7 receptors AND schizophrenia, serotonin 6 receptors agonist AND schizophrenia AND clinical trial, serotonin 7 receptors AND schizophrenia AND clinical trial, serotonin 6 receptors agonist AND serotonin 7 receptors schizophrenia agonist AND schizophrenia. Only English language articles from animal studies and clinical human studies were included. We narrowed our results by searching in conjunction with more specific terms, such as definition, mechanisms, agonist, antagonist and clinical trials. The content of the search results was divided into the following sections: ligands, mechanism of action, modulation of neurotransmitters pathway and role in schizophrenia.
Results
Ligands for 5-HT6R
Although several non-selective agonist and antagonist agents that strongly bind to serotonin 6 and 7 receptors were discovered long ago, the availability of selective ligands was increased in number and diversity after the discovery of human serotonin 6 and 7 receptors (Table 2).
Mechanism Action of 5-HT 6 and 5-HT 7 Receptors
Cyclic Adenosine Monophosphate (cAMP) Pathway
Serotonin 6 receptor activates cyclic adenosine monophosphate pathway (cAMP) signalling pathways through adenylate cyclase stimulation. This increases phosphorylation of dopamine and cAMP-regulated phosphoprotein by protein kinase A (PKA).2,23 In addition, the receptor stimulates the extracellular signal regulated kinase (ERK) through PKA and Ras monomeric GTPase-dependent mechanism.24 ERK1/2 activation is PKA and Ras-dependent but Rap1-independent and involved one or more Raf isoforms which is almost similar to 5-HT6 cAMP-mediated signalling mechanism. Activation of 5-HT6R stimulates cAMP-dependent pathways involving PKA then followed by Ras, several Raf, MEK and ERK1/2.25
Serotonin 7 receptor stimulates ERK in two ways. Firstly, using a cAMP-activated guanine nucleotide exchange factor (Epac) which is a PKA-independent pathway, that leads to Ras-dependent activation of the extracellular, signal-regulated kinases. Secondly, 5-HT7R directly stimulates ERK in hippocampal neurons which is important for hippocampal function and mood regulation.26 ERK1/2 activation through 5-HT7Rs was inhibited in cells pre-treated with the PKA inhibitor H89. This indicates the involvement of PKA in 5-HT7R-mediated signalling mechanism.27
In neurons, where 5-HT 6 and 7 receptors mainly operate, the activation of ERK1/2 is involved in cell morphological changes associated with neuroplasticity, such as the formation of synapses or dendritic spine maturation, thus contributing to learning, memory, cognition and other neural activities.28
Regulation of Fyn Kinase
Fyn is a non-receptor protein tyrosine kinase and is expressed to a high degree in neurons, glia and oligodendrocytes.29 5-HT6Rs and Fyn are co-localized on the cell surface and on intracellular membranes.30 The carboxyl terminus of 5-HT6Rs directly interacts with Fyn, leading to Fyn phosphorylation that further activates ERK1/2 through Ras and MEK-dependent pathways.31 5-HT6Rs–Fyn protein complexes play important roles in mammalian neuronal functions such as schizophrenia.25,31
Induction of Structural Plasticity
Neuronal circuit, cyto-architecture and plasticity of the brain neurons during development are shaped by both 5-HT6 and 7 receptors through proliferation, migration, differentiation, neurotic growth and synaptogenesis modulation. The 5-HT6Rs antagonists SB-271,046, SB-399,885 and SB-742,457 have all been shown to induce the expression levels of polysialylated neuronal cell adhesion molecule (PSA-NCAM) in the dentate gyrus of the rat. PSA-NCAM is indicator of neuronal development and synaptogenesis in immature brain and contributes to morpho-functional reorganization of adult brain. PSA creates physical hindrance to provide antiadhesive properties that reduce the binding affinity between opposite membranes of NCAM on neuronal processes, thus promoting structural plasticity.32 Thus, an increment in PSA-NCAM neurons by 5-HT6Rs antagonist supports their involvement in neuronal plasticity regulation.33
Serotonin 7 receptor activation affects the morphology of neurons, excitation of neurons and neuronal plasticity. This strongly suggests its modulator action in remodelling neuronal morphology and circuitry in the mature brain.34 In hippocampus neurons of mouse 5-HT7Rs activation stimulated the small GTP-ases RhoA and Cdc42 and enhanced neurite elongation, dendritic spine density, the number of synaptic contacts and the amount of AMPA receptors expressed at synapses, leading to increased synaptic efficacy.35 This is also supported by recent data that show that 5-HT7Rs agonists 8-OH-DPAT and LP-211 stimulated neurite outgrowth in primary cultures of mouse and rat striatal and cortical neurons by activation of the cyclin-dependent protein-kinase 5 (Cdk5), a kinase playing an important role in microtubule assembly and cytoarchitecture rearrangements.36
Serotonin 6 and 7 Receptor Modulation on Neurotransmitter Pathways
Cholinergic Pathway
Serotonin 6 receptor activation has an inhibitory role on acetylcholine release and is associated with cognitive impairment through cholinergic hypofunction. Selective 5-HT6Rs antagonists have provided direct evidence for the ability of 5-HT6Rs blockade to elevate cholinergic neurotransmission in different brain regions of freely moving adult rats, including the prefrontal cortex (RO-66-0074 and SB-39,988) and the dorsal hippocampus (SB-271,046).32,37,38 Atropine, a cholinergic antagonist, also attenuated behavioural phenomena associated with 5-HT6Rs inhibition including stretching and yawning. Treatment of rat with 5-HT6Rs antagonist antisense oligonucleotides produces a behavioural alteration that alters by atropine but not haloperidol. This shows the modulatory effect of 5-HT6Rs on Ach release. An effect of 5-HT6Rs on the activity of nigrostriatal pathway by modulation of Ach release is also observed in lesioned rats.39 Combined 5-HT6Rs antagonist and cholinesterase inhibitor galantamine improves cognitive symptoms in schizophrenic patients.40 Those findings indicate the positive influence of 5-HT6Rs antagonist in the release of acetylcholine. However, 5-HT7 heteroreceptors have a facilitator role for release of acetylcholine by coupling positively with adenyl cylase.41
Glutamate Pathway
Serotonin 6 receptors activation has inhibitory effect on release of glutamate.42 Chronic treatment with glutamatergic receptor antagonists resulted in a decrease in striatal mRNA levels of 5-HT6Rs. SB-271,046, a 5-HT6Rs antagonist, induces the release of the excitatory neurotransmitter glutamate in the dorsal hippocampus and frontal cortex, as well as aspartate in the frontal cortex.43
Clozapine has high affinity for 5-HT6Rs that enhance glutamate levels in the frontal cortex, and this neurochemical effect contributes to the efficacy of this drug in improving negative symptoms and cognitive dysfunction observed in patients with schizophrenia.40
Acute and chronic activations of 5-HT7Rs have opposite results on glutamate result. The effect of acute activation of 5-HT7Rs in hippocampus is in the post-synaptic terminals and increases the basal glutamate transmission by increasing AMPA receptors expression and NMDA-evoked current.44
However, long-term activation of 5-HT7Rs up-regulates platelet-derived growth factor (PDGF)-β receptor expression and activity which inhibits the membrane expression of NMDA receptors. This activation of PDGFβ receptors selectively inhibits NMDA receptor currents and is involved in neuroprotection by protecting neurons against NMDA-induced excitotoxicity.45
Gamma-Amino Butyric Acid (GABA) Pathway
Within the striatum 5-HT6Rs mRNA is expressed extensively on GABAergic striatopallidal neurons and striatonigral neurons. This localization and effect of 5-HT6R with GABAergic neurons was confirmed by WAY-466 5-HT6Rs agonist.32 Tonic activation of 5-HT6Rs expressed on GABAergic neurons leads to the induction of GABA release and in turn the inhibition of downstream cholinergic and glutamatergic neurons. Blockade of 5-HT6Rs effectively removes this tonic GABAergic inhibition of downstream neurons, resulting in enhanced neurotransmission of Ach and glutamate.32
The carboxyl terminal of the 5-HT6Rs recruits Cdk5, a signaling protein, which is implicated in schizophrenia. This 5-HT6/Cdk5-dependent signalling pathway affects neuronal migration, neurite growth and dendritic structure through phosphorylation of the Cdk5 substrate histone H1.46
Additionally, serotonin 6 receptor forms complexes with proteins that regulate developmental processes including mTOR pathway. The mTOR pathway plays a crucial role in cell proliferation, synaptogenesis and growth of dendrites and axons. Its disturbance in prefrontal and medial frontal cortex results in cognitive symptoms of schizophrenia.47
Modulation of GABAergic transmission in raphe nuclei by 5-HT7Rs activation decreases serotonin release.47 In the raphe nuclei 5-HT7Rs are not localized directly on serotonin cells but rather on GABAergic and glutamatergic neurons. The activation of 5-HT7Rs enhances the excitability of GABAergic neurons in globus pallidus neurons.48 5-HT7Rs-mediated GABA release and activation of spontaneous inhibitory postsynaptic currents frequency is partially due to increase in glutamate release from excitatory terminals located on interneurons through activation of 5-HT7Rs located on glutamatergic neurons and partially due to 5-HT7Rs localized on GABAergic cells.49
Dopamine and Norepinephrine Pathway
Differentiation of serotonin and dopamine neurons during the normal development requires sonic hedgehog (SHH) and fibroblast growth factor 8 (FGF-8) interactions, but serotonin is through a third signal, FGF-4, that modifies SHH and FGF-8 response. Addition of FGF-4 to ventral midbrain increases ectopic serotonin neuron development and inhibits endogenous dopamine neuron development. Therefore, serotonin antagonist increases dopamine synthesis by inhibiting serotonin.50
In schizophrenia, ascending serotoninergic pathways from the dorsal raphe nuclei to the substantia nigra and from the rostral raphe nuclei to the neocortex, limbic regions and basal ganglia are upregulated, leading to dopaminergic hypo-function. It is believed that the symptoms of schizophrenia are at least in part due to this interconnectivity between 5-HTergic and dopaminergic systems.51
Medial prefrontal cortex is abundantly innervated by catecholaminergic neurons, and is associated with cognitive function, temporal organization of behaviour and behavioural flexibility. 5-HT6Rs activation in this brain region results in inhibition of dopamine and norepinephrine release, but selective antagonist increases those neurotransmitters.32 For example, a study conducted on rat using systemic administration of ST1936 5-HT6R agonist shows increasing dopamine and noradrenalin in the prefrontal cortex.25
In addition, generation of dopamine neurons in neurospheres of mesencephalic precursors increases by inhibition of glial 5-HT7Rs. This is confirmed by an increase in generation of dopamine neurons after elimination of the 5-HT neurons or reduction of 5-HT levels using methiothepin (5-HT1,2,5,6,7 receptor antagonist), and the 5-HT7Rs antagonist SB 269,970.52
Blockade of 5-HT7Rs by SB-269,970 increases 5-HT levels in cortical terminals. Since SB-269,970 has no affinity for dopamine and adrenergic receptors, nor does it show any dopamine and noradrenalin reuptake inhibition activity, its effect is through indirect modulating action of released serotonin on dopamine and noradrenalin neurons via 5-HT7 heteroreceptors localized on dopamine and noradrenalin neurons.53
Role of 5-HT 6 and 5-HT 7 Receptors in Schizophrenia
The 5-HT 6 and 7 receptors are associated with a number of psychiatric disorders, including schizophrenia.54 Significant reduction of 5-HT7Rs at prefrontal cortex in post mortem studies of schizophrenic patients and positive genetic correlation between 5-HT7Rs gene with schizophrenia are the suggestive points for the role of 5-HT7Rs in schizophrenia pathophysiology.55 In addition to this, high affinity of multiple antipsychotics drugs like clozapine and risperidone for 5-HT7Rs and their anatomical distribution within the brain increase the motive to study their role in schizophrenia.56
Antipsychotics (risperidone, ziprasidone, pimozide, sertindole, zotepine, clozapine and olanzapine), antidepressants (amitriptyline) and 5-HT2 receptor antagonists (mesulergine, ritanserin and LY215840) have a high affinity for 5-HT 6 and 7 receptors.28
In animal models of psychosis, SB-271,406, a 5-HT6Rs antagonist, selectively increases extracellular concentrations of both dopamine and epinephrine,38 hippocampus glutamate and aspartate within the frontal cortex,57 PSA-NCAM in hippocampus and dose-dependently reversed amphetamine-induced prepulse inhibition disruption.58 SB-399,885, a 5-HT6Rs antagonist, increases dopamine, norepinephrine, acetylcholine concentration and hippocampus PSA-NCAM.55 On top of this, enhancement of GABA and reduction of glutamate in hippocampus by way-466, a 5-HT6Rs antagonist, in combination with increasing of DRAPP-2 phosphorylation by EMDT, suggest that 5-HT6Rs antagonists may have some therapeutic utility in the treatment of schizophrenia.59
The distribution of 5-HT6Rs mRNA in the limbic and cortical regions of the human brain together with the high affinity of many antipsychotic agents could suggest a role in the development of schizophrenia. It may be possible that SB271046, a selective 5-HT6Rs antagonist, can improve cognition through facilitation of cortical and hippocampal glutamatergic activity and acetylcholine release. Although 5-HT6Rs distribute most densely in the STR and NAC, SB271046, a selective 5-HT6Rs antagonist, has been reported to increase glutamate release but not dopamine release in the mPFC and dorsal hippocampus without an effect in the NAC or STR.60
Besides the effects on cognition, 5-HT6Rs antagonists have potential effects on other comorbid conditions in schizophrenia, such as reduction of alcohol and nicotine addiction61 and cocaine abuse62 and had anxiolytic-like and antidepressant-like effects in rodents.63
Serotonin 6 receptor stimulation is associated with activation of mammalian target of rapamycin (mTOR) signalling. 5-HT6Rs-induced cognitive defects are inhibited by mTOR inhibitor drugs like raphamycin. This mTOR inhibition is associated with improvement in social cognation, working memory, executive function and episodic memory.64 Additionally, combination of prazosin with 5-HT6R antagonist PRX-07034 enhances memory, which indicates its use in schizophrenia.65
Atypical antipsychotic, typical antipsychotic and antidepressant drugs show high affinity for 5-HT7Rs. For example, amisulpride is an effective antidepressant drug effective against schizophrenia, but it also shows modest 5-HT7Rs antagonist potency.66 Additionally, postpartum studies from schizophrenic individuals show deficiency of 5-HT7Rs expression in cortex.67 A genomewide study on schizophrenia also indicates linkage of 10q22 gene in close proximity with human 5-HT7Rs 10q21–24 gene.68
Animal models predictive of antipsychotic-like activity are used to evaluate 5-HT7Rs antagonists, through administration of amphetamine or antagonists of NMDA receptors like phencyclidine, ketamine or dizocilpine. Antagonists of the NMDA receptors evoke behaviours that reflect both positive symptoms and negative symptoms and cognitive impairments.69
The 5-HT7Rs antagonists SB-258,741 and SB-269,970 significantly blocked NMDA receptor antagonist amphetamine-induced hyperactivity. SB-269,970 decreases amphetamine and ketamine-induced hyperactivity and reverses amphetamine-induced prepulse inhibition disruption in mice, without changing startle amplitude.70 It also reverses phencyclidine-induced deficits in the novel object recognition test in rats. 5-HT7 receptor antagonists ameliorate behavioural abnormalities in PACAP-deficient mice.71 Together, these observations indicate that pharmacological blockade of the 5-HT7Rs has anti-schizophrenic effects.
Different 5-HT7Rs antagonists show precognitive effect (anticipation of future events after forced exposure during trial) in animals. This supports the possible use 5-HT7Rs antagonism as a target in the management of cognitive symptoms in schizophrenic patients. For example, scopolamine, MK-801 and PCP-induced learning and memory impairment is attenuated by lurasidone, and scopolamine and PCP-induced learning defect is attenuated by SB-269,970.72 Additionally, LP-211, a selective 5-HT7Rs antagonist, improves spatial reference memory, motor coordination, synaptic plasticity and overall emotional memory.73
Dizocilpine-induced learning and memory impairments in the passive avoidance and Morris water maze tests in rats are reversed by 5-HT7Rs antagonist SB-656,104-A. This implies a role of 5-HT7Rs antagonist in cognitive defect of schizophrenia.74 SB-258,741 ameliorates the PCP but not amphetamine-induced disruption of prepulse inhibition. This indicates that 5-HT7Rs affects the glutaminergic but not dopaminergic path of PCP. These data are in conflict with results in apomorphine and amphetamine-treated 5-HT7R knock-out mice.56
The pharmacological blockade of 5-HT7Rs also has therapeutic implications for the treatment of negative symptoms in schizophrenia. Although SB-25,874 had no beneficial effects on PCP-induced deficits in social interactions, SB-269,970 ameliorated ketamine-induced social withdrawal in rats.75
Several 5-HT 6 and 7 receptor ligands are under clinical trials (Table 3), and multiple mechanisms of action have been proposed for the different effects of 5-HT 6 and 7 receptor antagonists: Firstly, by enhancing dopamine, glutamate and Ach efflux76 and decreasing GABA levels in the prefrontal cortex, 5-HT 6 and 7 receptor antagonists may decrease the release of GABA in the prefrontal cortex, which subsequently disinhibits glutamate and/or Ach release.
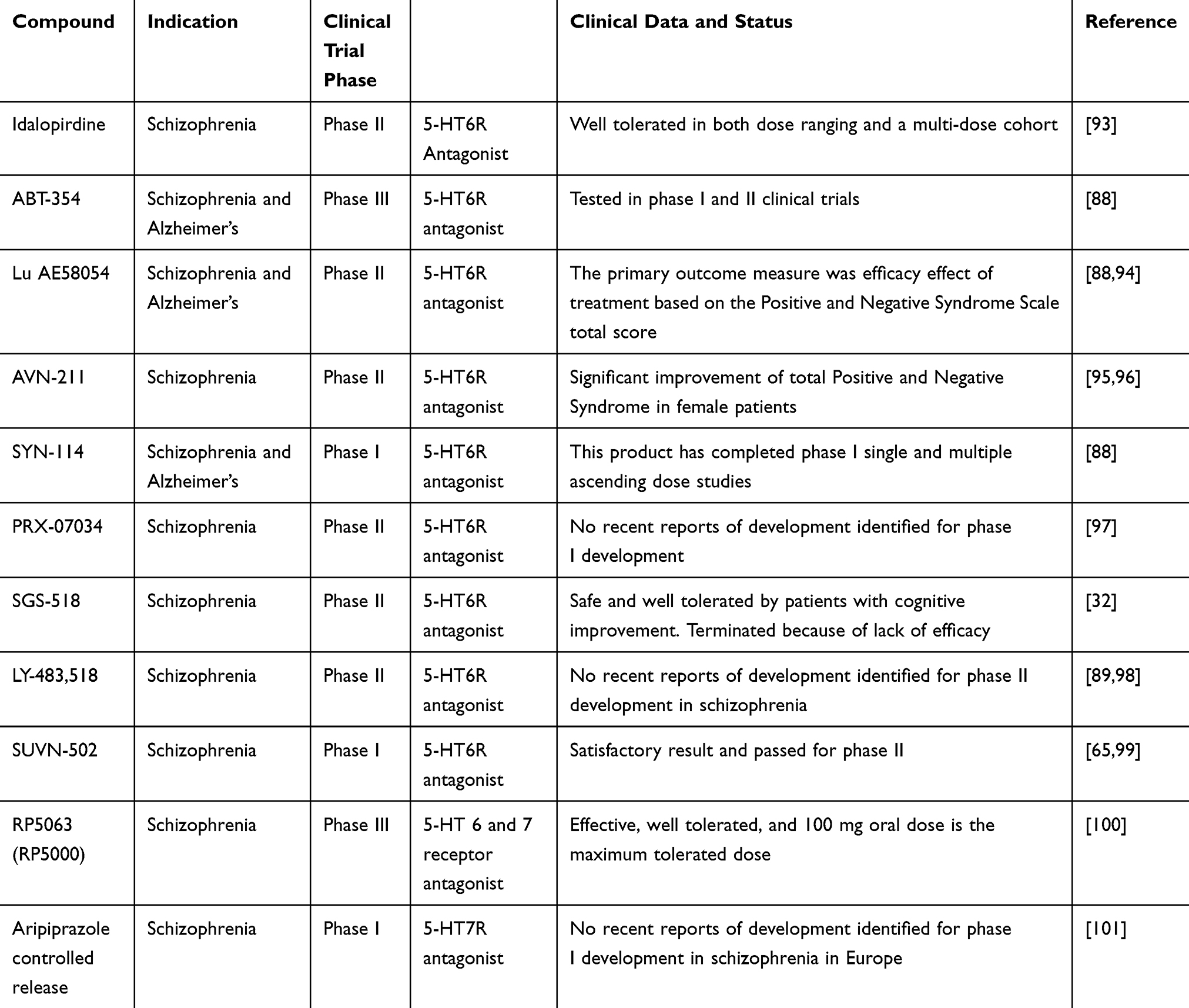 |
Table 3 Serotonin 6 and 7 Receptors Antagonist Under Clinical Trial |
Secondly, 5-HT 6 and 7 receptor antagonists may decrease GABAergic interneuron excitability, leading to disinhibition and a subsequent enhancement of synaptic plasticity in synapses and brain regions where 5-HT6Rs are expressed.77
Thirdly, chronic administration of 5-HT 6 and 7 receptor antagonists increased the number of NCAM PSA-immunoreactive neurons in the dentate gyrus and in the entorhinal and perirhinal regions of the cortex, which affects synaptic plasticity, possibly enhancing the ability of neurons to remodel their connections and contribute to ongoing adaptations in neuronal circuits.78
Fourthly, 5-HT6Rs antagonists may have pro-cognitive effects through the decrease in mTOR signalling.79 Fifthly, 5-HT 6 and 7 receptor antagonists may affect cognition through interaction of the 5-HT 6 and 7 receptors with the Fyn-tyrosine kinase: 5-HT6Rs agonists were shown to increase Fyn kinase activity, and 5-HT6Rs antagonists79 antagonized effects. Finally, 5-HT 6 and 7 receptors interact with DARPP-32, which affects cognitive function and pathogenesis of schizophrenia.55
Conclusion
Recent neurohumoral study indicates that there is involvement of not only dopamine but also serotonin through its serotonin 6 and 7 receptors in the pathogenesis of schizophrenia. The different effects of 5-HT6 and 7 receptor antagonists on schizophrenia are by enhancing dopamine, glutamate and Ach efflux and decreasing GABA levels in the prefrontal cortex, increasing the number of NCAM PSA-immunoreactive neurons in the dentate gyrus and in the entorhinal and perirhinal regions of the cortex, which affects synaptic plasticity, possibly enhancing the ability of neurons to remodel their connections and contribute to on-going adaptations in neuronal circuits, through the decrease in mTOR signalling. Finally, serotonin 6 and 7 receptors antagonist plays a role in cognitive function and possibly in the pathogenesis of schizophrenia via interaction with DARPP-32.
Acknowledgment
Writing and editorial assistance was given by medicinal chemistry department, School of Pharmacy, Mekelle University.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in conception, study design, execution, acquisition of data, analysis and interpretation, or in all those areas; took part in drafting, revising or critical reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
There is no competing interest among the authors.
References
1. McCorvy JD, Roth BL. Structure and function of serotonin G protein-coupled receptors. Pharmacol Ther. 2015;150:129–142. doi:10.1016/j.pharmthera.2015.01.009
2. Yun HM, Rhim H. The serotonin-6 receptor as a novel therapeutic target. Exp Neurobiol. 2011;20(4):159–168.
3. Meneses A, Liy-Salmeron G. Serotonin and emotion, learning and memory. Rev Neurosci. 2012;23(5–6):543–553. doi:10.1515/revneuro-2012-0060
4. Young SN, Leyton M. The role of serotonin in human mood and social interaction. Insight from altered tryptophan levels. Pharmacol Biochem Behav. 2002;71(4):857–865. doi:10.1016/S0091-3057(01)00670-0
5. Schmitt JA, Wingen M, Ramaekers JG, Evers EA, Riedel WJ. Serotonin and human cognitive performance. Curr Pharm Des. 2006;12(20):2473–2486. doi:10.2174/138161206777698909
6. Morin LP. Serotonin and the regulation of mammalian circadian rhythmicity. Ann Med. 1999;31(1):12–33. doi:10.3109/07853899909019259
7. Gerin CG, Hill A, Hill S, Smith K, Privat A. Serotonin release variations during recovery of motor function after a spinal cord injury in rats. Synapse (New York, NY). 2010;64(11):855–861. doi:10.1002/syn.20802
8. Ciranna L, Catania MV. 5-HT7 receptors as modulators of neuronal excitability, synaptic transmission and plasticity: physiological role and possible implications in autism spectrum disorders. Front Cell Neurosci. 2014;8:250. doi:10.3389/fncel.2014.00250
9. Bleich A, Brown SL, Kahn R, van Praag HM. The role of serotonin in schizophrenia. Schizophr Bull. 1988;14(2):297–315. doi:10.1093/schbul/14.2.297
10. Muller CL, Anacker AMJ, Veenstra-VanderWeele J. The serotonin system in autism spectrum disorder: from biomarker to animal models. Neuroscience. 2016;321:24–41. doi:10.1016/j.neuroscience.2015.11.010
11. Geldenhuys WJ, Van der Schyf CJ. Role of serotonin in Alzheimer’s disease: a new therapeutic target? CNS Drugs. 2011;25(9):765–781. doi:10.2165/11590190-000000000-00000
12. Cowen PJ, Browning M. What has serotonin to do with depression? World Psychiatr. 2015;14(2):158–160. doi:10.1002/wps.20229
13. Baldwin D, Rudge S. The role of serotonin in depression and anxiety. Int Clin Psychopharmacol. 1995;9(Suppl 4):41–45. doi:10.1097/00004850-199501004-00006
14. Muller CP, Homberg JR. The role of serotonin in drug use and addiction. Behav Brain Res. 2015;277:146–192. doi:10.1016/j.bbr.2014.04.007
15. Wurtman RJ, Wurtman JJ. Carbohydrate craving, obesity and brain serotonin. Appetite. 1986;7:99–103. doi:10.1016/S0195-6663(86)80055-1
16. Pytliak M, Vargova V, Mechirova V, Felsoci M. Serotonin receptors - from molecular biology to clinical applications. Pharmacol Res. 2011;60(1):15–25. doi:10.33549/physiolres.931903
17. Naughton M, Mulrooney JB, Leonard BE. A review of the role of serotonin receptors in psychiatric disorders. Hum Psychopharmacol. 2000;15(6):397–415. doi:10.1002/1099-1077(200008)15:6<397::AID-HUP212>3.0.CO;2-L
18. Nikiforuk A. Targeting the serotonin 5-HT7 receptor in the search for treatments for CNS disorders: rationale and progress to date. CNS Drugs. 2015;29(4):265–275. doi:10.1007/s40263-015-0236-0
19. Mohler EG, Baker PM, Gannon KS, et al. The effects of PRX-07034, a novel 5-HT6 antagonist, on cognitive flexibility and working memory in rats. Psychopharmacology. 2012;220(4):687–696. doi:10.1007/s00213-011-2518-7
20. Frederick JA, Meador-Woodruff JH. Effects of clozapine and haloperidol on 5-HT6 receptor mRNA levels in rat brain. Schizophr Res. 1999;38(1):7–12. doi:10.1016/S0920-9964(99)00002-X
21. Perez-Garcia G, Gonzalez-Espinosa C, Meneses A. An mRNA expression analysis of stimulation and blockade of 5-HT7 receptors during memory consolidation. Behav Brain Res. 2006;169(1):83–92. doi:10.1016/j.bbr.2005.12.013
22. Ruat M, Traiffort E, Leurs R, et al. Molecular cloning, characterization, and localization of a high-affinity serotonin receptor (5-HT7) activating cAMP formation. Proc Natl Acad Sci. 1993;90(18):8547–8551. doi:10.1073/pnas.90.18.8547
23. Yger M, Girault JA. DARPP-32, jack of all trades … master of which? Front Behav Neurosci. 2011;5:56. doi:10.3389/fnbeh.2011.00056
24. Hoyer D, Hannon JP, Martin GR. Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol Biochem Behav. 2002;71(4):533–554.
25. Riccioni T, Bordi F, Minetti P, et al. ST1936 stimulates cAMP, Ca2+, ERK1/2 and Fyn kinase through a full activation of cloned human 5-HT6 receptors. Eur J Pharmacol. 2011;661(1–3):8–14. doi:10.1016/j.ejphar.2011.04.028
26. Hedlund PB, Sutcliffe JG. Functional, molecular and pharmacological advances in 5-HT7 receptor research. Trends Pharmacol Sci. 2004;25(9):481–486. doi:10.1016/j.tips.2004.07.002
27. Norum JH, Hart K, Levy FO. Ras-dependent ERK activation by the human G(s)-coupled serotonin receptors 5-HT4(b) and 5-HT7(a). J Biol Chem. 2003;278(5):3098–3104. doi:10.1074/jbc.M206237200
28. Yun H-M, Rhim H. The serotonin-6 receptor as a novel therapeutic target. Exp Neurobiol. 2011;20(4):159–168.
29. Sperber BR, McMorris FA. Fyn tyrosine kinase regulates oligodendroglial cell development but is not required for morphological differentiation of oligodendrocytes. J Neurosci Res. 2001;63(4):303–312. doi:10.1002/1097-4547(20010215)63:4<303::AID-JNR1024>3.0.CO;2-A
30. Kim SH, Kim DH, Lee KH, et al. Direct interaction and functional coupling between human 5-HT6 receptor and the light chain 1 subunit of the microtubule-associated protein 1B (MAP1B-LC1). PLoS One. 2014;9(3):e91402. doi:10.1371/journal.pone.0091402
31. Yun HM, Kim S, Kim HJ, et al. The novel cellular mechanism of human 5-HT6 receptor through an interaction with Fyn. J Biol Chem. 2007;282(8):5496–5505. doi:10.1074/jbc.M606215200
32. Upton N, Chuang TT, Hunter AJ, Virley DJ. 5-HT6 receptor antagonists as novel cognitive enhancing agents for Alzheimer’s disease. NeuroRx. 2008;5(3):458–469. doi:10.1016/j.nurt.2008.05.008
33. van Loevezijn A, Venhorst J, Bakker WII, et al. Optimization of N′-(arylsulfonyl) pyrazoline-1-carboxamidines by exploiting a novel interaction site in the 5-HT6 antagonistic binding pocket. Bioorg Med Chem Lett. 2016;26(6):1605–1611. doi:10.1016/j.bmcl.2016.02.001
34. Volpicelli F, Speranza L, Di Porzio U, Crispino M, Perrone-Capano C. The serotonin receptor 7 and the structural plasticity of brain circuits. Front Behav Neurosci. 2014;8. doi:10.3389/fnbeh.2014.00318
35. Kobe F, Guseva D, Jensen TP, et al. 5-HT7R/G12 signaling regulates neuronal morphology and function in an age-dependent manner. J Neurosci. 2012;32(9):2915–2930. doi:10.1523/JNEUROSCI.2765-11.2012
36. Speranza L, Giuliano T, Volpicelli F, et al. Activation of 5-HT7 receptor stimulates neurite elongation through mTOR, Cdc42 and actin filaments dynamics. Front Behav Neurosci. 2015;9:62. doi:10.3389/fnbeh.2015.00062
37. East SZ, Burnet PW, Kerwin RW, Harrison PJ. An RT-PCR study of 5-HT(6) and 5-HT(7) receptor mRNAs in the hippocampal formation and prefrontal cortex in schizophrenia. Schizophr Res. 2002;57(1):15–26. doi:10.1016/S0920-9964(01)00323-1
38. Lacroix LP, Dawson LA, Hagan JJ, Heidbreder CA. 5-HT6 receptor antagonist SB-271046 enhances extracellular levels of monoamines in the rat medial prefrontal cortex. Synapse (New York, NY). 2004;51(2):158–164. doi:10.1002/syn.10288
39. Sleight AJ, Boess FG, Bös M, Levet‐Trafit B, Riemer C, Bourson A. Characterization of Ro 04‐6790 and Ro 63‐0563: potent and selective antagonists at human and rat 5‐HT6 receptors. Br J Pharmacol. 1998;124(3):556–562.
40. Marcos B, Chuang TT, Gil-Bea FJ, Ramirez MJ. Effects of 5-HT6 receptor antagonism and cholinesterase inhibition in models of cognitive impairment in the rat. Br J Pharmacol. 2008;155(3):434–440. doi:10.1038/bjp.2008.281
41. D’Agostino G, Condino AM, Gallinari P, Franceschetti GP, Tonini M. Characterization of prejunctional serotonin receptors modulating [3H]acetylcholine release in the human detrusor. J Pharmacol Exp Ther. 2006;316(1):129–135. doi:10.1124/jpet.105.092551
42. Xie G, Zuo W, Wu L, et al. Serotonin modulates glutamatergic transmission to neurons in the lateral habenula. Sci Rep. 2016;6.
43. Ciranna L. Serotonin as a modulator of glutamate- and GABA-mediated neurotransmission: implications in physiological functions and in pathology. Curr Neuropharmacol. 2006;4(2):101–114. doi:10.2174/157015906776359540
44. Vasefi MS, Yang K, Li J, et al. Acute 5-HT7 receptor activation increases NMDA-evoked currents and differentially alters NMDA receptor subunit phosphorylation and trafficking in hippocampal neurons. Mol Brain. 2013;6:24. doi:10.1186/1756-6606-6-24
45. Vasefi MS, Kruk JS, Liu H, Heikkila JJ, Beazely MA. Activation of 5-HT7 receptors increases neuronal platelet-derived growth factor beta receptor expression. Neurosci Lett. 2012;511(2):65–69. doi:10.1016/j.neulet.2012.01.016
46. Nikiforuk A. The procognitive effects of 5-HT6 receptor ligands in animal models of schizophrenia. Rev Neurosci. 2014;25(3):367–382. doi:10.1515/revneuro-2014-0005
47. Meffre J, Chaumont-Dubel S, Mannoury la Cour C. 5-HT 6 receptor recruitment of mTOR as a mechanism for perturbed cognition in schizophrenia. EMBO Mol Med. 2012;4(10):1043–1056.
48. Racca C, Catania MV, Monyer H, Sakmann B. Expression of AMPA-glutamate receptor B subunit in rat hippocampal GABAergic neurons. Eur J Neurosci. 1996;8(8):1580–1590. doi:10.1111/j.1460-9568.1996.tb01303.x
49. Tokarski K, Kusek M, Hess G. 5HT-receptors modulate GABAergic transmission in rat hippocampal CA1 area. J Physiol Pharmacol. 2011;62(5):535.
50. Rodriguez-Pallares J, Guerra MJ, Labandeira-Garcia JL. Elimination of serotonergic cells induces a marked increase in generation of dopaminergic neurons from mesencephalic precursors. Eur J Neurosci. 2003;18(8):2166–2174. doi:10.1046/j.1460-9568.2003.02949.x
51. Svob Strac D, Pivac N, Muck-Seler D. The serotonergic system and cognitive function. Transl Neurosci. 2016;7(1):35–49. doi:10.1515/tnsci-2016-0007
52. Parga J, Rodriguez-Pallares J, Munoz A, Guerra MJ, Labandeira-Garcia JL. Serotonin decreases generation of dopaminergic neurons from mesencephalic precursors via serotonin type 7 and type 4 receptors. Dev Neurobiol. 2007;67(1):10–22. doi:10.1002/dneu.20306
53. Wesolowska A, Kowalska M. Influence of serotonin 5-HT(7) receptor blockade on the behavioral and neurochemical effects of imipramine in rats. Pharmacol Rep. 2008;60(4):464–474.
54. Vogt IR, Shimron-Abarbanell D, Neidt H, et al. Investigation of the human serotonin 6 [5-HT6] receptor gene in bipolar affective disorder and schizophrenia. Am J Med Genet. 2000;96(2):217–221. doi:10.1002/(SICI)1096-8628(20000403)96:2<217::AID-AJMG17>3.0.CO;2-0
55. Nikiforuk A. Serotonergic and cholinergic strategies as potential targets for the treatment of schizophrenia. Curr Pharm Des. 2016;22(14):2093–2116. doi:10.2174/1381612822666160127113758
56. Hedlund PB. The 5-HT 7 receptor and disorders of the nervous system: an overview. Psychopharmacology. 2009;206(3):345–354.
57. Dawson LA. The central role of 5-HT6 receptors in modulating brain neurochemistry. Int Rev Neurobiol. 2011;96:1–26.
58. Pouzet B, Didriksen M, Arnt J. Effects of the 5-HT7 receptor antagonist SB-258741 in animal models for schizophrenia. Pharmacol Biochem Behav. 2002;71(4):655–665. doi:10.1016/S0091-3057(01)00744-4
59. Schechter I, Butler PD, Jalbrzikowski M, Pasternak R, Saperstein AM, Javitt DC. A new dimension of sensory dysfunction: stereopsis deficits in schizophrenia. Biol Psychiatry. 2006;60(11):1282–1284. doi:10.1016/j.biopsych.2006.03.064
60. Meltzer HY, Li Z, Kaneda Y, Ichikawa J. Serotonin receptors: their key role in drugs to treat schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry. 2003;27(7):1159–1172. doi:10.1016/j.pnpbp.2003.09.010
61. De Bruin N, McCreary A, Van Loevezijn A, et al. A novel highly selective 5-HT6 receptor antagonist attenuates ethanol and nicotine seeking but does not affect inhibitory response control in Wistar rats. Behav Brain Res. 2013;236:157–165. doi:10.1016/j.bbr.2012.08.048
62. Valentini V, Piras G, De Luca MA, et al. Evidence for a role of a dopamine/5-HT6 receptor interaction in cocaine reinforcement. Neuropharmacology. 2013;65:58–64. doi:10.1016/j.neuropharm.2012.08.025
63. Wesołowska A, Partyka A, Jastrzębska-Więsek M, Kołaczkowski M. The preclinical discovery and development of cariprazine for the treatment of schizophrenia. Expert Opin Drug Discov. 2018;13(8):779–790. doi:10.1080/17460441.2018.1471057
64. Oguro-Ando A, Rosensweig C, Herman E, et al. Increased CYFIP1 dosage alters cellular and dendritic morphology and dysregulates mTOR. Mol Psychiatry. 2015;20(9):1069–1078.
65. de Bruin MWJ, Kruse C. 5-HT6 receptor antagonists: potential efficacy for the treatment of cognitive impairment in schizophrenia. Curr Pharm Des. 2015;21(26):3739–3759.
66. Kampf P, Agelink MW, Maß R, Jahn H, Schafer I, Naber D. Amisulpride in addition to clozapine: a retrospective study indicates improved efficacy and good tolerability. German J Psychiatr. 2003;6(3):64–68.
67. East SZ, Burnet PW, Leslie RA, Roberts JC, Harrison PJ. 5‐HT6 receptor binding sites in schizophrenia and following antipsychotic drug administration: autoradiographic studies with [125I] SB‐258585. Synapse. 2002;45(3):191–199. doi:10.1002/syn.10097
68. Ikeda M, Iwata N, Kitajima T, et al. Positive association of the serotonin 5-HT 7 receptor gene with schizophrenia in a Japanese population. Neuropsychopharmacology. 2006;31(4):866–871. doi:10.1038/sj.npp.1300901
69. Neill JC, Barnes S, Cook S, et al. Animal models of cognitive dysfunction and negative symptoms of schizophrenia: focus on NMDA receptor antagonism. Pharmacol Ther. 2010;128(3):419–432. doi:10.1016/j.pharmthera.2010.07.004
70. Galici R, Boggs JD, Miller KL, Bonaventure P, Atack JR. Effects of SB-269970, a 5-HT7 receptor antagonist, in mouse models predictive of antipsychotic-like activity. Behav Pharmacol. 2008;19(2):153–159. doi:10.1097/FBP.0b013e3282f62d8c
71. Horiguchi M, Huang M, Meltzer HY. The role of 5-hydroxytryptamine 7 receptors in the phencyclidine-induced novel object recognition deficit in rats. J Pharmacol Exp Ther. 2011;338(2):605–614. doi:10.1124/jpet.111.180638
72. De Filippis B, Nativio P, Fabbri A, et al. Pharmacological stimulation of the brain serotonin receptor 7 as a novel therapeutic approach for Rett syndrome. Neuropsychopharmacology. 2014;39(11):2506–2518.
73. Horisawa T, Nishikawa H, Toma S, et al. The role of 5-HT7 receptor antagonism in the amelioration of MK-801-induced learning and memory deficits by the novel atypical antipsychotic drug lurasidone. Behav Brain Res. 2013;244:66–69. doi:10.1016/j.bbr.2013.01.026
74. Matthys A, Haegeman G, Van Craenenbroeck K, Vanhoenacker P. Role of the 5-HT7 receptor in the central nervous system: from current status to future perspectives. Mol Neurobiol. 2011;43(3):228–253. doi:10.1007/s12035-011-8175-3
75. Nikiforuk A, Kos T, Fijał K, Hołuj M, Rafa D, Popik P. Effects of the selective 5-HT7 receptor antagonist SB-269970 and amisulpride on ketamine-induced schizophrenia-like deficits in rats. PLoS One. 2013;8(6):e66695. doi:10.1371/journal.pone.0066695
76. Dawson LA. The Central Role of 5-HT6 Receptors in Modulating Brain Neurochemistry. International Review of Neurobiology. Vol. 96. Elsevier; 2011:1–26.
77. West P, Marcy V, Marino M, Schaffhauser H. Activation of the 5-HT6 receptor attenuates long-term potentiation and facilitates GABAergic neurotransmission in rat hippocampus. Neuroscience. 2009;164(2):692–701. doi:10.1016/j.neuroscience.2009.07.061
78. Foley AG, Hirst WD, Gallagher HC, et al. The selective 5-HT6 receptor antagonists SB-271046 and SB-399885 potentiate NCAM PSA immunolabeling of dentate granule cells, but not neurogenesis, in the hippocampal formation of mature Wistar rats. Neuropharmacology. 2008;54(8):1166–1174. doi:10.1016/j.neuropharm.2008.03.012
79. Meffre J, Chaumont‐Dubel S, la Cour CM, et al. 5‐HT6 receptor recruitment of mTOR as a mechanism for perturbed cognition in schizophrenia. EMBO Mol Med. 2012;4(10):1043–1056.
80. Gérard C, Mestikawy SE, Lebrand C, et al. Quantitative RT‐PCR distribution of serotonin 5‐HT6 receptor mRNA in the central nervous system of control or 5, 7‐dihydroxytryptamine‐treated rats. Synapse. 1996;23(3):164–173. doi:10.1002/(SICI)1098-2396(199607)23:3<164::AID-SYN5>3.0.CO;2-6
81. Gellynck E, Heyninck K, Andressen KW, et al. The serotonin 5-HT 7 receptors: two decades of research. Exp Brain Res. 2013;230(4):555–568. doi:10.1007/s00221-013-3694-y
82. Nikiforuk A, Kos T, Wesołowska A. The 5-HT6 receptor agonist EMD 386088 produces antidepressant and anxiolytic effects in rats after intrahippocampal administration. Psychopharmacology. 2011;217(3):411–418. doi:10.1007/s00213-011-2297-1
83. Cole DC, Lennox WJ, Lombardi S, et al. Discovery of 5-arylsulfonamido-3-(pyrrolidin-2-ylmethyl)-1 H-indole derivatives as potent, selective 5-HT6 receptor agonists and antagonists. J Med Chem. 2005;48(2):353–356. doi:10.1021/jm049243i
84. Heal D, Smith S, Fisas A, Codony X, Buschmann H. Selective 5-HT6 receptor ligands: progress in the development of a novel pharmacological approach to the treatment of obesity and related metabolic disorders. Pharmacol Ther. 2008;117(2):207–231. doi:10.1016/j.pharmthera.2007.08.006
85. Mitchell ES, Neumaier JF. 5-HT6 receptors: a novel target for cognitive enhancement. Pharmacol Ther. 2005;108(3):320–333. doi:10.1016/j.pharmthera.2005.05.001
86. Schechter LE, Lin Q, Smith DL, et al. Neuropharmacological profile of novel and selective 5-HT 6 receptor agonists: WAY-181187 and WAY-208466. Neuropsychopharmacology. 2008;33(6):1323–1335. doi:10.1038/sj.npp.1301503
87. Fijał K, Pachuta A, McCreary AC, et al. Effects of serotonin (5-HT) 6 receptor ligands on responding for cocaine reward and seeking in rats. Pharmacol Rep. 2010;62(6):1005–1014. doi:10.1016/S1734-1140(10)70362-5
88. Froestl W, Muhs A, Pfeifer A. Cognitive enhancers (nootropics). Part 1: drugs interacting with receptors. Update 2014. J Alzheimers Dis. 2014;41(4):961–1019. doi:10.3233/JAD-140228
89. Ohno Y, Mizuguchi Y, Ohno Y. Improving the treatment of schizophrenia: role of 5-HT receptors in modulating cognitive and extrapyramidal motor. CNS Neurol Disord Drug Targets. 2013;12:861–869. doi:10.2174/18715273113129990088
90. Pittala V, Salerno L, Modica M, Siracusa MA, Romeo G. 5-HT7 receptor ligands: recent developments and potential therapeutic applications. Mini Rev Med Chem. 2007;7(9):945–960. doi:10.2174/138955707781662663
91. Perrone R, Berardi F, Colabufo NA, Lacivita E, Leopoldo M, Tortorella V. Synthesis and structure− affinity relationships of 1-[ω-(4-aryl-1-piperazinyl) alkyl]-1-aryl ketones as 5-HT7 receptor ligands. J Med Chem. 2003;46(4):646–649. doi:10.1021/jm020994z
92. Medina RA, Sallander J, Benhamú B, et al. Synthesis of new serotonin 5-HT7 receptor ligands. Determinants of 5-HT7/5-HT1A receptor selectivity. J Med Chem. 2009;52(8):2384–2392. doi:10.1021/jm8014553
93. Codony X, Vela JM, Ramírez MJ. 5-HT6 receptor and cognition. Curr Opin Pharmacol. 2011;11(1):94–100. doi:10.1016/j.coph.2011.01.004
94. Arnt J, Bang-Andersen B, Grayson B, et al. Lu AE58054, a 5-HT6 antagonist, reverses cognitive impairment induced by subchronic phencyclidine in a novel object recognition test in rats. Int J Neuropsychopharmacol. 2010;13(8):1021–1033. doi:10.1017/S1461145710000659
95. Morozova M, Burminskiy D, Rupchev G, et al. 5-HT6 receptor antagonist as an adjunct treatment targeting residual symptoms in patients with schizophrenia: unexpected sex-related effects (double-blind placebo-controlled trial). J Clin Psychopharmacol. 2017;37(2):169–175. doi:10.1097/JCP.0000000000000673
96. Morozova MA, Lepilkina TA, Rupchev GE, et al. Add-on clinical effects of selective antagonist of 5HT6 receptors AVN-211 (CD-008-0173) in patients with schizophrenia stabilized on antipsychotic treatment: pilot study. CNS Spectr. 2014;19(4):316–323. doi:10.1017/S1092852913000394
97. Abraham R, Nirogi R, Shinde A, Irupannanavar S. Low-dose prazosin in combination with 5-HT6 antagonist PRX-07034 has antipsychotic effects. Can J Physiol Pharmacol. 2015;93(1):13–21. doi:10.1139/cjpp-2014-0254
98. Shimizu S, Mizuguchi Y, Ohno Y. Improving the treatment of schizophrenia: role of 5-HT receptors in modulating cognitive and extrapyramidal motor functions. CNS Neurol Disord Drug Targets. 2013;12(6):861–869.
99. Nirogi R, Abraham R, Benade V, et al. SUVN-502, a novel, potent, pure, and orally active 5-HT6 receptor antagonist: pharmacological, behavioral, and neurochemical characterization. Behav Pharmacol. 2019;30(1):16–35. doi:10.1097/FBP.0000000000000414
100. Cantillon M, Bhat L. Response to concerns over Cantillon et al. dopamine serotonin stabilizer RP5063 clinical trial’s design, analyses and findings (SCHRES-D-17-00455) by Ahmed S Aboraya, MD, DrPh. Schizophr Res. 2018;195:581–582. doi:10.1016/j.schres.2017.09.010
101. Meltzer HY, Risinger R, Nasrallah HA, et al. A randomized, double-blind, placebo-controlled trial of aripiprazole lauroxil in acute exacerbation of schizophrenia. J Clin Psychiatry. 2015;76(8):1085–1090. doi:10.4088/JCP.14m09741
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.