")
Back to Journals » Cancer Management and Research » Volume 11
S100A16 regulated by Snail promotes the chemoresistance of nonmuscle invasive bladder cancer through the AKT/Bcl-2 pathway
Authors Wang C, Zhu X, Li A, Yang S, Qiao R, Zhang J
Received 29 November 2018
Accepted for publication 27 February 2019
Published 27 March 2019 Volume 2019:11 Pages 2449—2456
DOI https://doi.org/10.2147/CMAR.S196450
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Antonella D'Anneo
Chanjuan Wang,1 Xi Zhu,2 Aiwei Li,1 Shuo Yang,1 Rui Qiao,1 Jie Zhang1
1Department of Laboratory Medicine, Peking University Third Hospital, Beijing, People’s Republic of China; 2Department of Urology, Beijing Friendship Hospital Affiliated to Capital Medical University, Beijing, People’s Republic of China
Objective: To fully investigate the effect of S100 proteins on the chemoresistance of nonmuscle invasive bladder cancer (NMIBC).
Materials and methods: The mitomycin C-resistant bladder cancer cell line M-RT4 was established and liquid chromatography-tandem mass spectrometry was performed for proteomics analysis. RT-PCR and Western blot were then performed to confirm the findings. To investigate the mechanisms, S100A16 was knocked down by siRNA. Then, the sensitivity of M-RT4 to mitomycin C was analyzed by a cell counting kit-8 (CCK8) assay, and the molecular expression including epithelial-mesenchymal transition (EMT)-related and apoptosis-related markers were also examined by Western blot. Based on the cancer genome atlas (TCGA) data, the prognostic value of S100A16 was also investigated.
Results: There were six S100 proteins with differential expression, among which S100A16 was confirmed to be the only upregulated protein in M-RT4 cells. The expression of S100A16 was regulated by the EMT-related transcription factor Snail. Knockdown of S100A16 suppressed the AKT/Bcl-2 pathway to promote apoptosis, greatly sensitizing M-RT4 cells to mitomycin C. The expression of S100A16 was negatively correlated with the overall survival of bladder cancer patients.
Conclusion: S100A16 contributes to the chemoresistance of NMIBC by promoting the AKT/Bcl-2-mediated anti-apoptosis effect and could be a potential prognostic marker and therapeutic target for NMIBC patients.
Keywords: nonmuscle invasive bladder cancer, chemoresistance, S100A16, apoptosis, Snail
Introduction
Bladder cancer has become the most common carcinoma in the urinary system with a high recurrence rate. Nonmuscle invasive bladder cancer (NMIBC) accounts for approximately 75% of all cases.1,2 Transurethral resection of bladder tumor (TURBT) followed by intravesical chemotherapy is the main treatment for NMIBC patients. However, there are still some patients who hardly benefit from the treatment due to the emergence of chemoresistance, and the mechanisms require further investigation.3 Therefore, understanding how chemoresistance occurs is of great significance for the prognosis of bladder cancer patients.
The S100 protein is the largest subfamily of calcium-binding proteins bearing EF-hand motifs.4 Upon calcium binding, most S100 proteins undergo a conformational change to interact with different targets, which exerts various intracellular regulatory activities. Additionally, S100 proteins can be secreted outside of cells and act in a cytokine-like manner by binding surface receptors, thus allowing them to play a role in extracellular signaling.5–7 Studies have shown that altered expression patterns of S100 genes are associated with many human disorders, including various cancers.8 Agerbaek M reported that focal S100A4 expression is a significant independent predictor of distant metastatic relapse for muscle invasive bladder cancer (MIBC) patients.9 Similarly, Kim JH suggested that S100A4 expression has prognostic value in primary NMIBC.10 In addition, overexpression of S100A8 or S100A11 and inactivation of S100A2 are also positively associated with tumorigenesis and progression of bladder cancer.11,12 Chemoresistance has a close relationship with cancer progression and prognosis. Therefore, whether the S100 family affects the chemotherapeutic response of NMIBC remains to be investigated.
In this study, we performed liquid chromatography-tandem mass spectrometry (LC-MS/MS) to identify which S100 proteins are expressed significantly in mitomycin C-resistant NMIBC cells to select potential markers to predict or reverse the chemoresistance of NMIBC.
Materials and methods
Cell lines
The human bladder cancer cell line RT4, which is a good representative cell line of NMIBC, was purchased from ATCC. The corresponding mitomycin C (MMC)-resistant cell line M-RT4 was generated and confirmed according to our previously published study.13 Both cell lines were cultured in DMEM/F12 (1:1) medium supplemented with 10% standard fetal bovine serum (FBS) and 1% penicillin-streptomycin (Gibco, penicillin: 100 IU/ml and streptomycin: 100 mg/ml) at 37℃ in a 5% CO2 atmosphere.
Proteomics
RT4 and M-RT4 cells were harvested and then lysed by RIPA. Protein extraction and SDS-PAGE electrophoresis were performed to ensure the integrity and quality of proteins. For proteomics analysis, LC-MS/MS was carried out by CapitalBio Technology (Beijing, China). The related parameters were provided in
Quantitative real-time PCR
The extraction of total RNA, reverse transcription to cDNA and quantitative real-time PCR (RT-PCR) were performed as described in our previous study.14 The reaction conditions for RT-PCR are shown in Table 1. The primers used for amplification were as follows: S100A16, (sense) 5ʹ-ACTGCTACACGGAGCTGGAGA-3ʹ and (antisense) 5ʹ-GCAAGGGTCAGAGGAAGGTCT-3ʹ; S100A13, (sense) 5ʹ-TCCTAATGGCAGCAGAACCACTGA-3ʹ and (antisense) 5ʹ-TTCTTCCTGATTTCCTTGGCCAGC-3ʹ; GAPDH, (sense) 5ʹ-TGTTCCAATATGATTCCACCC-3ʹ and (antisense) 5ʹ-CTTCTCCATGGTGCGTGAAGA-3ʹ. The relative mRNA expression was normalized to GAPDH, which was calculated based on the Ct value according to the equation: 2^-ΔΔCt. Each sample was analyzed in triplicate.
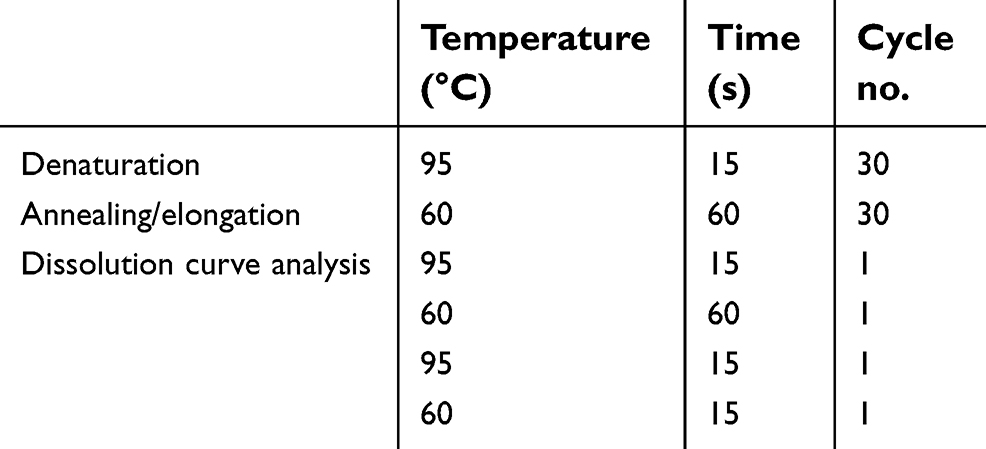 | Table 1 The reaction conditions of RT-PCR |
Western blot analysis
Cells were harvested to extract proteins, followed by Western blot as described previously.14 Briefly, proteins (30 μg each) were loaded onto 10% sodium dodecyl sulfate-polyacrylamide gels, electrophoresed, and transferred onto polyvinylidene difluoride membranes. The membranes were then blocked for 1 h in 5% skim milk followed by incubation with primary antibodies at 4 °C overnight. The primary antibodies used were as follows: S100A16 (1:500 dilution, ab130419, Abcam, US), epithelial-mesenchymal transition (EMT) antibody sampler kit (1:1,000 dilution, #9782, Cell Signaling Technology, US), Phospho-AKT pathway antibody sampler kit (1:1,000 dilution, 9916T, Cell Signaling Technology, US), Bcl-2 (1:500 dilution, #2870S, Cell Signaling Technology, US), Bax (1:1,000 dilution, #2772S, Cell Signaling Technology, US) and GAPDH (1:2,500 dilution, MBL, Japan). After three washes with TBST, the membranes were incubated with secondary antibodies conjugated to LI-COR IRDye for 1 h at room temperature and were then detected using an Odyssey Imager (LI-COR Biosciences, Lincoln, NE).
Immunofluorescence
Cells were seeded on 25 mm diameter slides in a 6-well plate. After 24 h of incubation, the cells were washed with PBS three times and fixed in 4% paraformaldehyde for 20 min at room temperature. After washing three times, the cells were permeated with 0.3% Triton-X. Then, the cells were blocked using 1% bovine serum albumin (BSA) for 1 h at room temperature before staining with an anti-S100A16 primary antibody (1:100 dilution, Abcam, USA) overnight at 4 °C. On the following day, the cells were washed and stained with a FITC-labeled goat anti-rabbit secondary antibody (1:100 dilution, ZSGB-BIO, China) for 2 h at room temperature. After washing with PBS three times, the cells were stained with DAPI and covered with an anti-fluorescence quenching agent (Applygen, China). Finally, the slides were examined under a fluorescence microscope (Olympus).
CCK8 assay
A cell counting kit-8 (CCK8) assay was performed as described previously.13 Briefly, S100A16 or Snail knockdown M-RT4 cells were seeded in triplicate in 96-well plates at a density of 1 × 104 cells per well. After incubation for 24 h, 10 μl of MMC (HZB0149, HARVEY, US) at concentrations ranging from 0.78 to 100 μg/ml was added to each well. After another incubation for 24 h, 10 μl of CCK8 solution was added to each well and incubated for 2 h. The optical density (OD) was measured at a wavelength of 450 nm. Cell viability was calculated as follows: (OD value of experimental groups – OD value of blank groups)/(OD value of control groups – OD value of blank groups).
Knockdown assay
S100A16 and Snail siRNAs were purchased from Santa Cruz Biotechnology (sc-88602 and sc-38398, respectively) and dissolved in DEPC water that was provided by the manufacturer. Transfection medium and transfection reagent were also provided by Santa Cruz Biotechnology (sc-36868 and sc-29528, respectively). M-RT4 cells were cultured in 6-well plates to a density of 60% and a mixture of siRNA was then added using standard procedures. After incubation for 7 h, 1 ml DMEM/F12 medium containing FBS was added followed by another 24 h of incubation. Then, the cells were harvested to perform Western blot, CCK8 assay and flow cytometry.
Flow cytometry
Cell death was analyzed by flow cytometry with an annexin V-FITC/propidium iodide (PI) apoptosis assay kit (Zoman Biotechnology Co., Ltd., Beijing, China). Briefly, cells were harvested and resuspended in PBS at a density of 1×105 cells per tube. Then, the samples were centrifuged, and the supernatant was discarded. Then, 500 μl of 1× binding buffer was added to resuspend the samples. Then, 5 μl annexin V-FITC and 10 μl PI were added, followed by incubation in the dark for 15 min at room temperature. Cells that were positive for annexin V (Q2 and Q4) were considered to be undergoing apoptosis, including early and late apoptosis. In all cases, 10,000 events per tube were acquired. Flow cytometric analysis was performed on a FACS-Canto II flow cytometer (BD Biosciences).
Statistical analysis
Data were analyzed using the GraphPad Prism 5 software (GraphPad Software, Inc., San Diego, CA) and SPSS Statistics 20 (IBM, USA). The results were presented as the mean±SD. Student’s t-test was used for comparison of parental and MMC-resistant cells, and p<0.05 was considered statistically significant.
Results
There were six S100 proteins with significant differential expression according to LC-MS/MS
We established an MMC-resistant bladder cancer cell line (M-RT4) from parental RT4 cells, and the chemoresistance of M-RT4 cells was verified using a CCK8 assay in our previous study.13 To identify whether the S100 family plays an essential role in chemoresistance, we performed LC-MS/MS for proteomics analysis between M-RT4 and RT4 cells. Prior to the analysis, the concentration and homogeneity of proteins were confirmed by SDS-PAGE electrophoresis (
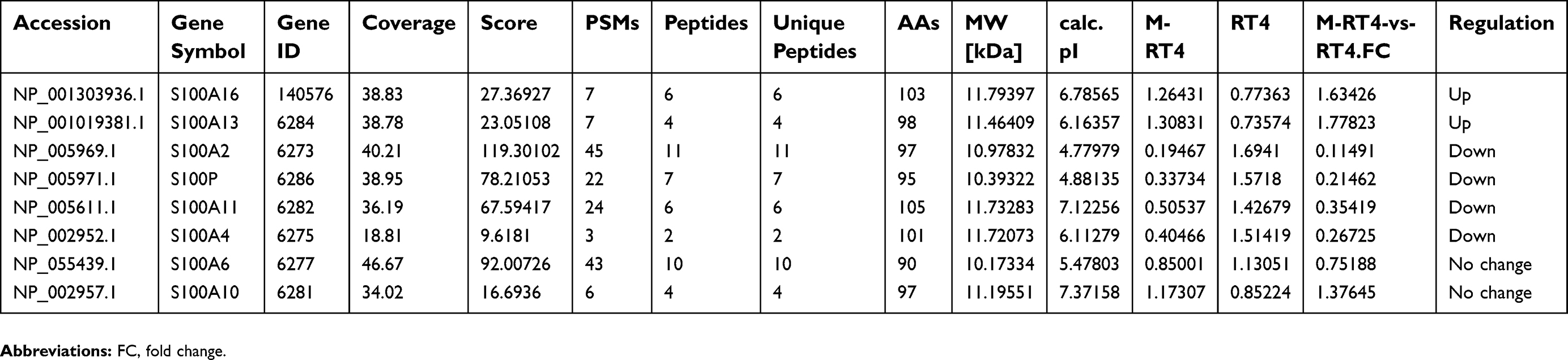 | Table 2 The differential expression of S100 family in M-RT4 and RT4 cells |
S100A16 was significantly upregulated in M-RT4 cells compared with RT4 cells
In this study, we focused on the two upregulated S100 proteins, S100A16 and S100A13.To confirm the LC-MS/MS results, we performed RT-PCR and Western blot to investigate the expression of S100A16 and S100A13 in M-RT4 and RT4 cells. Both the mRNA and protein levels of S100A16 in M-RT4 cells were significantly greater than those in RT4 cells (Figure 1A and B), while there was no detectable expression of S100A13.
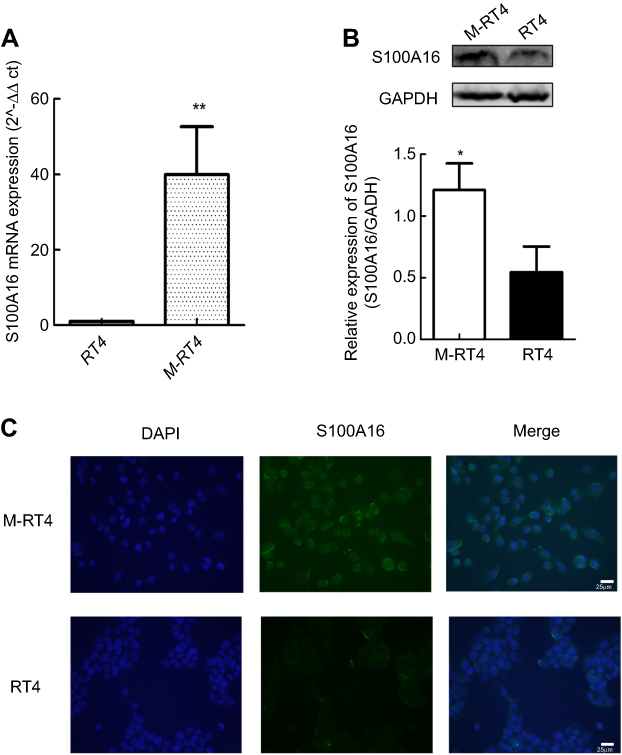 | Figure 1 S100A16 expression was upregulated in M-RT4 cells. (A) RT-PCR results showed the mRNA expression level of S100A16 in M-RT4 and RT4 cells. The results are presented as 2^-ΔΔCt values (ΔCt=Ct(S100A16) – Ct(GAPDH). ΔΔCt=ΔCt(M-RT4) – ΔCt(RT4)). (B) Western blot results showed the protein expression level of S100A16 in M-RT4 and RT4 cells. (C) The expression site of S100A16 in cells indicated by immunofluorescence (20× magnification). The scale bar represents 25 μm. *p<0.05; **p<0.01. |
Next, we also performed immunofluorescence staining to investigate the expression sites of S100A16 in cells. We found that S100A16 was mainly expressed in the nucleus and on the nuclear membrane of M-RT4 cells. Additionally, S100A16 was also expressed in the cytoplasm. However, no specific fluorescent signals were detected in RT4 cells (Figure 1C).
Knockdown of S100A16 could sensitize M-RT4 cells to MMC
To further confirm the effect of S100A16 on chemoresistance, we decreased the expression of S100A16 in M-RT4 cells by siRNA. First, we used four different concentrations of siRNA. Western blot confirmed the success of the knockdown assay, especially for 60 and 80 pmol siRNA (Figure 2A). After adding 60 pmol siRNA to knock down S100A16 in M-RT4 cells, a CCK8 assay was performed. The CCK8 assay showed that the IC50 decreased significantly in S100A16-knockdown M-RT4 cells compared with M-RT4 cells without siRNA (Figure 2B and C), suggesting that the sensitivity of M-RT4 cells to MMC was greatly restored by suppressing S100A16 expression.
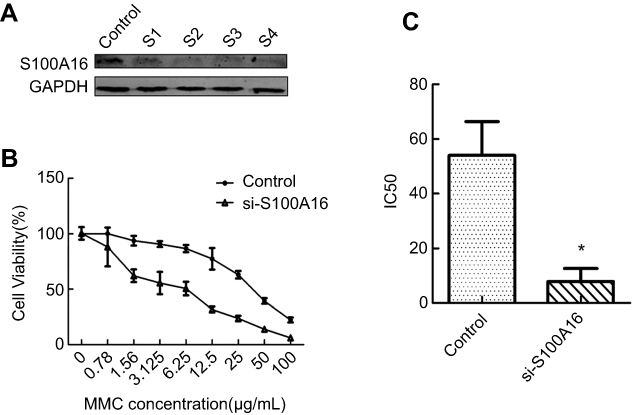 | Figure 2 Knockdown of S100A16 could restore the sensitivity of M-RT4 cells to MMC. (A) The success of the knockdown assay was confirmed by Western blot. Control: transfection reagent and transfection medium (without siRNA). S1: 20 pmol siRNA. S2: 40 pmol siRNA. S3: 60 pmol siRNA. S4: 80 pmol siRNA. (B) Cell viability after MMC addition analyzed by a CCK8 assay. The concentration of MMC ranged from 0.78 to 100 μg/mL. si-S100A16: 60 pmol siRNA. (C) The IC50 of MMC for M-RT4 and RT4 cells. IC50: half maximal inhibitory concentration. MMC: mitomycin C. *p<0.05. |
S100A16 regulated by Snail could regulate apoptosis through the AKT/Bcl-2 pathway
To elucidate the mechanisms by which S100A16 exerts an effect on chemoresistance, we performed Western blot to compare the molecular changes after S100A16 knockdown, including EMT-related and apoptosis-related molecules. We found that EMT markers, including N-cad, E-cad, Vimentin and Claudin-1, were not significantly affected by the knockdown of S100A16 (Figure 3A), while Bcl-2 and pAKT/AKT were significantly suppressed in S100A16-downregulated M-RT4 cells (Figure 3C).
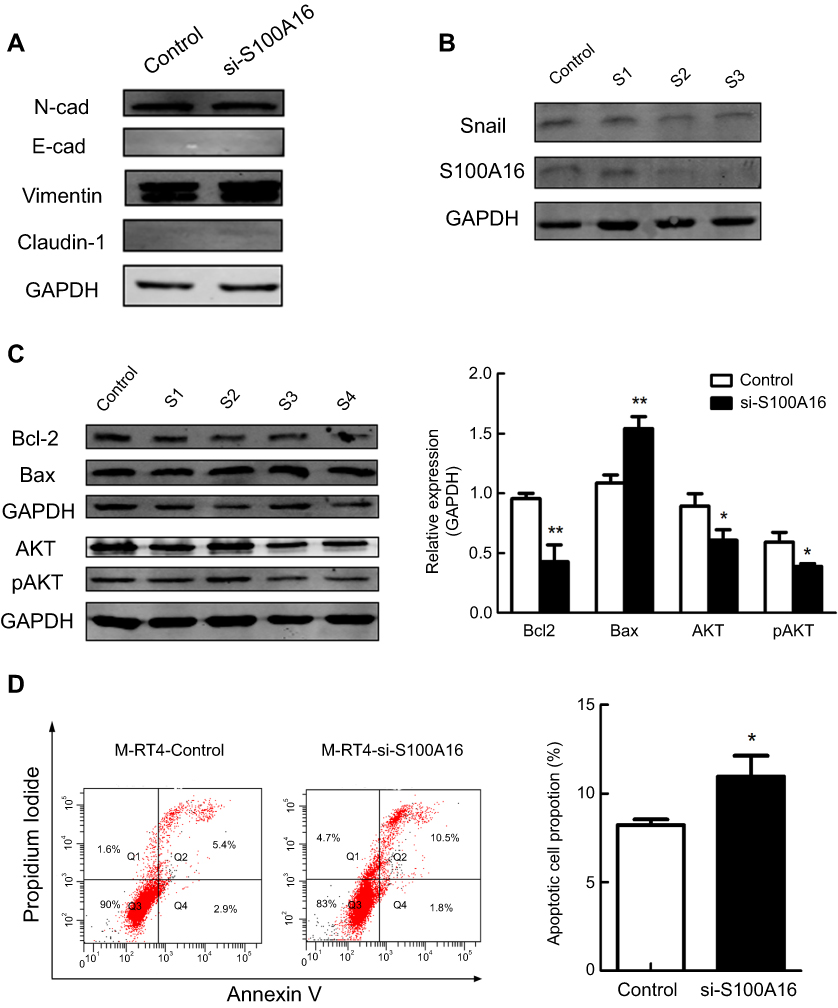 | Figure 3 Knockdown of S100A16 could suppress the AKT/Bcl-2 pathway to promote apoptosis in M-RT4 cells. (A) The effect of S100A16 knockdown on EMT-related molecules in M-RT4 cells. Control: transfection reagent and transfection medium (without siRNA). si-S100A16: 60 pmol siRNA. (B) The effect of Snail knockdown on S100A16 expression. Control: transfection reagent and transfection medium (without siRNA). S1: 20 pmol Snail siRNA. S2: 40 pmol Snail siRNA. S3: 60 pmol Snail siRNA. (C) The effect of S100A16 knockdown on apoptosis-related molecules in M-RT4 cells. Control: transfection reagent and transfection medium (without siRNA). S1: 20 pmol S100A16 siRNA. S2: 40 pmol S100A16 siRNA. S3: 60 pmol S100A16 siRNA. S4: 80 pmol S100A16 siRNA. pAKT: phospho-AKT (S473). (D) Apoptosis of M-RT4 cells after S100A16 knockdown. Q4: early apoptosis. Q2: late apoptosis. Q2+Q4: total apoptosis. Control: transfection reagent and transfection medium (without siRNA). si-S100A16: 60 pmol siRNA. *p<0.05; **p<0.01. |
Because S100A16 did not affect EMT-related molecules, we speculated whether the expression of S100A16 is affected by the EMT process. We suppressed the expression of Snail, an important transcription factor of EMT, by siRNA in M-RT4 cells. Western blot confirmed the success of the knockdown assay. Indeed, S100A16 expression was significantly downregulated after the knockdown of Snail (Figure 3B), confirming our speculation.
Considering Bcl-2 and AKT as anti-apoptosis markers, we then performed flow cytometry for apoptosis analysis to confirm the Western blot results. Annexin V+/PI– indicates early apoptosis, and annexin V+/PI+ indicates late apoptosis. Total apoptosis increased significantly after S100A16 knockdown (
S100A16 could be a prognostic marker for bladder cancer patients
Since chemoresistance has a close relationship with patient prognosis, whether the expression of S100A16 can predict the prognosis of bladder cancer patients remains to be investigated. Therefore, we analyzed survival data on the OncoLnc website (
Considering the regulatory effect of Snail on S100A16, we also explored whether Snail has prognostic value for bladder cancer. There also was a significantly negative correlation between overall survival and the expression of SNAI1 (the Snail gene), which was consistent with S100A16 (
Discussion
Similar to the other 17 kinds of S100 genes, S100A16 is located on chromosome 1q21, a region that frequently exhibits rearrangements during tumor development.15 Upregulation of S100A16 expression has been observed in many different cancers, including bladder, lung, thyroid gland, pancreas, and ovarian cancers.16 In our study, we discovered that S100A16 is the only significantly upregulated S100 protein in the MMC-resistant cell line. Additionally, decreasing the expression level of S100A16 was notably shown to sensitize M-RT4 cells to MMC, which was mediated by the suppression of the AKT/Bcl-2 pathway to promote apoptosis. In accordance with our study, Mani J also demonstrated that chemoresistance in bladder cancer decreases significantly after suppressing the Bcl-2-mediated anti-apoptosis effect.17 In Emmanuel’s study, S100A16 was found to accumulate within nucleoli and to translocate to the cytoplasm in response to Ca2+ stimulation.18 Similarly, we observed a wide distribution of S100A16 with main expression in the nucleus and on the nuclear membrane. In addition, a study on colorectal cancer conducted by Sun X demonstrated a rich S100A16 expression on the cell membrane, which might be associated with the extracellular function of S100A16.19
EMT has been reported to participate in the chemoresistance of cancers in addition to tumor cell migration and metastasis, especially due to the close relationship between EMT and cancer stem cells.20 Our previous study showed that epithelial markers such as Claudin-1 and E-cad were downregulated in M-RT4 cells compared with RT4 cells, while mesenchymal markers, including Snail, N-cad and Vimentin, were upregulated.13 Zhou et al showed that upregulation of S100A16 expression promotes EMT via the Notch1 pathway in breast cancer.21 However, our study showed that knockdown of S100A16 is not sufficient to suppress EMT. The reason may be due to the multiple regulatory pathways involved in EMT. Interestingly, we found that the expression of S100A16 was regulated by the EMT-related transcription factor Snail. Therefore, the complicated regulatory network between EMT and S100A16 remains to be elucidated in future studies.
S100A16 has been speculated to be a prognostic marker for colorectal cancer, lung adenocarcinoma and breast cancer.19,22,23 In this study, we also confirmed the prognostic value of S100A16 in bladder cancer by analyzing TCGA data. Since the onset of chemoresistance is one of main causes of poor prognosis for patients, it is reasonable that there is a negative correlation between S100A16 and survival.
In addition to the S100 family, proteomics analysis showed that more than 500 proteins or peptides were differentially expressed. In addition, we further performed gene ontology (GO) and pathway enrichment analysis (
In conclusion, our study suggested that S100A16 contributes to the chemoresistance of NMIBC by suppressing apoptosis, which is mediated by the AKT/Bcl-2 pathway. This study might provide a new strategy to predict response to chemotherapy and overcome chemoresistance in bladder cancer.
Acknowledgments
We are grateful to CapitalBio Co., Ltd. (Beijing, China) for technical support and proteomics analysis. This work was supported by the National Natural Science Foundation of China (61401290), the Beijing Municipal Administration of Hospitals’ Youth Programme (QML20160104) and the Administration of Hospitals Clinical Medicine Development of Special Funding Support (ZYLX201604).
Disclosure
Miss Chanjuan Wang reports non-financial support from CapitalBio Co., Ltd (Beijing, China), outside the submitted work; Dr Xi Zhu reports grants from National Nature Science Foundation of China, Administration of Hospitals Clinical Medicine Development of Special Funding Support, Beijing Municipal Administration of Hospitals’ Youth programme, during the conduct of the study; non-financial support from CapitalBio Co., Ltd (Beijing, China), outside the submitted work; Miss Aiwei Li reports non-financial support from CapitalBio Co., Ltd (Beijing, China), outside the submitted work; Dr Shuo Yang reports non-financial support from CapitalBio Co., Ltd (Beijing, China), outside the submitted work; Dr Rui Qiao reports non-financial support from CapitalBio Co., Ltd (Beijing, China), outside the submitted work; Dr Jie Zhang reports non-financial support from CapitalBio Co., Ltd (Beijing, China), outside the submitted work. The authors report no other conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2017. CA Cancer J Clin. 2017;67(1):7–30. doi:10.3322/caac.21387
2. Kamat AM, Hahn NM, Efstathiou JA, et al. Bladder cancer. Lancet. 2016;388(10061):2796–2810. doi:10.1016/S0140-6736(16)30512-8
3. Huncharek M, McGarry R, Kupelnick B. Impact of intravesical chemotherapy on recurrence rate of recurrent superficial transitional cell carcinoma of the bladder: results of a meta-analysis. Anticancer Res. 2001;21(1b):765–769.
4. Donato R. S100: a multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int J Biochem Cell Biol. 2001;33:637–668.
5. Heizmann CW, Fritz G, Schafer BW. S100 proteins: structure, functions and pathology. Front Biosci. 2002;7:d1356–1368.
6. Donato R. Intracellular and extracellular roles of S100 proteins. Microsc Res Tech. 2003;60(6):540–551. doi:10.1002/jemt.10296
7. Donato R, Sorci G, Giambanco I. S100A6 protein: functional roles. Cell Mol Life Sci. 2017;74(15):2749–2760. doi:10.1007/s00018-017-2526-9
8. Hongyan Chen CX, Qing’e Jin ZL. S100 protein family in human cancer. Am J Cancer Res. 2014;4(2):89–115.
9. Agerbaek M, Alsner J, Marcussen N, et al. Focal S100A4 protein expression is an independent predictor of development of metastatic disease in cystectomized bladder cancer patients. Eur Urol. 2006;50(4):777–785. doi:10.1016/j.eururo.2006.02.027
10. Kim JH, Park J. Prognostic significance of heme oxygenase-1, S100 calcium-binding protein A4, and syndecan-1 expression in primary non-muscle-invasive bladder cancer. Hum Pathol. 2014;45(9):1830–1838. doi:10.1016/j.humpath.2014.04.020
11. Yao R, Davidson DD, Lopez-Beltran A, et al. The S100 proteins for screening and prognostic grading of bladder cancer. Histol Histopathol. 2007;22(9):1025–1032. doi:10.14670/HH-22.1025
12. Lee J, Wysocki PT, Topaloglu O, et al. Epigenetic silencing of S100A2 in bladder and head and neck cancers. Oncoscience. 2015;2(4):410–418. doi:10.18632/oncoscience.140
13. Wang C, Li A, Yang S, et al. CXCL5 promotes mitomycin C resistance in non-muscle invasive bladder cancer by activating EMT and NF-kappaB pathway. Biochem Biophys Res Commun. 2018;498(4):862–868. doi:10.1016/j.bbrc.2018.03.071
14. Zheng J, Zhu X, Zhang J. CXCL5 knockdown expression inhibits human bladder cancer T24 cells proliferation and migration. Biochem Biophys Res Commun. 2014;446(1):18–24. doi:10.1016/j.bbrc.2014.01.172
15. Ridinger K, Ilg EC, Niggli FK, et al. Clustered organization of S100 genes in human and mouse. Biochim Biophys Acta. 1998;1448(2):254–263.
16. Marenholz I, Heizmann CW. S100A16, a ubiquitously expressed EF-hand protein which is up-regulated in tumors. Biochem Biophys Res Commun. 2004;313(2):237–244.
17. Mani J, Vallo S, Rakel S, et al. Chemoresistance is associated with increased cytoprotective autophagy and diminished apoptosis in bladder cancer cells treated with the BH3 mimetic (-)-Gossypol (AT-101). BMC Cancer. 2015;15:224. doi:10.1186/s12885-015-1584-3
18. Sturchler E, Cox JA, Durussel I, et al. S100A16, a novel calcium-binding protein of the EF-hand superfamily. J Biol Chem. 2006;281(50):38905–38917. doi:10.1074/jbc.M605798200
19. Sun X, Wang T, Zhang C, et al. S100A16 is a prognostic marker for colorectal cancer. J Surg Oncol. 2018;117(2):275–283. doi:10.1002/jso.24822
20. Saxena M, Stephens MA, Pathak H, et al. Transcription factors that mediate epithelial–mesenchymal transition lead to multidrug resistance by upregulating ABC transporters. Cell Death Dis. 2011;2(7):e179. doi:10.1038/cddis.2011.82
21. Zhou W, Pan H, Xia T, et al. Up-regulation of S100A16 expression promotes epithelial-mesenchymal transition via Notch1 pathway in breast cancer. J Biomed Sci. 2014;21:97. doi:10.1186/s12929-014-0097-8
22. Katono K, Sato Y, Kobayashi M, et al. S100A16, a promising candidate as a prognostic marker for platinum-based adjuvant chemotherapy in resected lung adenocarcinoma. Onco Targets Ther. 2017;10:5273–5279. doi:10.2147/OTT.S145072
23. Tanaka M, Ichikawa-Tomikawa N, Shishito N, et al. Co-expression of S100A14 and S100A16 correlates with a poor prognosis in human breast cancer and promotes cancer cell invasion. BMC Cancer. 2015;15:53. doi:10.1186/s12885-015-1584-3
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.