")
Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 16
rs217727 of lncRNA H19 is Associated with Cervical Cancer Risk in the Chinese Han Population
Authors Dai J, Zhang S, Shi Y, Xu J, Liu W, Yang J, Shi L, Yan Z, Li C
Received 17 June 2023
Accepted for publication 6 September 2023
Published 30 October 2023 Volume 2023:16 Pages 933—948
DOI https://doi.org/10.2147/PGPM.S422083
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Jie Dai,1,* Shao Zhang,2,* Yuhan Shi,3 Jinmei Xu,2 Weipeng Liu,1 Jia Yang,1 Li Shi,1 Zhiling Yan,2,4 Chuanyin Li1
1Department of Immunogenetics, Institute of Medical Biology, Chinese Academy of Medical Sciences & Peking Union Medical College, Kunming, 650118, People’s Republic of China; 2Department of Gynaecologic Oncology, The No. 3 Affiliated Hospital of Kunming Medical University, Kunming, 650118, People’s Republic of China; 3College of Agronomy and Biotechnology, Yunnan Agricultural University, Kunming, 650041, People’s Republic of China; 4Department of Gynaecologic Oncology, The Hospital of Yuanmou, Yuanmou, 651300, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhiling Yan, Department of Gynaecologic Oncology, The No. 3 Affiliated Hospital of Kunming Medical University, Kunming, 650118, People’s Republic of China, Email [email protected] Chuanyin Li, Department of Immunogenetics, Institute of Medical Biology, Chinese Academy of Medical Sciences & Peking Union Medical College, Kunming, 650118, People’s Republic of China, Email [email protected]
Background: Long noncoding RNAs (LncRNAs) have been revealed to involve in cervical cancer (CC) developing. The current study was designed to explore the association of SNPs (rs217727, rs2366152, rs1859168, rs10505477) located in the lncRNA H19, HOTAIR, HOTTIP and CASC8 genes with the risk of CC in a Chinese Han population.
Methods: Four SNPs were selected and genotyped in 1426 participants (274 CIN patients, 448 CC patients, and 704 healthy control individuals) using MassArray. The association of these SNPs with susceptibility to CC was evaluated.
Results: Significant differences in allelic distribution of rs217727 were observed in the comparison of CC with control (P = 0.001), indicating the risk of rs217727-A allele in CC (OR = 1.33; 95% CI: 1.12– 1.58). The inheritance model analysis revealed that 2AA+GA genotype represented a certain risk of CC (P = 0.001, OR = 1.35; 95% CI: 1.13– 1.62). The stratified analysis revealed a risk of the rs217727-A allele for cervical squamous cell carcinoma (SCC) (P = 0.002, OR = 1.33; 95% CI: 1.11– 1.60).
Conclusion: rs217727 in lncRNA H19 exhibited a significant correlation with CC susceptibility, particularly SCC, and A/A genotype of this SNP might present as a risk in CC.
Keywords: cervical cancer, lncRNA, H19, association, SNPs, CC, long noncoding RNAs
Introduction
Cervical cancer (CC) is a common gynecological cancer in women.1 In 2020, the incidence and mortality of CC ranked fourth in female tumors. There were approximately 604,000 newly diagnosed cases, accounting for approximately 6.5% of all female cancers, and 342,000 deaths from CC, comprising 7.7% of all female cancer deaths globally.2
Generally, persistent infection with high risk human papillomavirus (HR-HPV) is necessary for CC.1,3 However, only a small proportion of women who are infected with HR-HPV ultimately develop CC.1,3,4 Other factors also play important roles in CC developing, and might be the decisive factors for the difference in susceptibility among individuals.5 Several studies reported that long noncoding RNAs (lncRNAs) involved in the development of human cancers, including CC.6–8 LncRNAs are RNA molecules larger than 200 nucleotides in length. Previous data has demonstrated the crucial roles of lncRNA in various cellular processes, such as cell cycle, apoptosis, epigenetics, and regulation of gene expression.9 Furthermore, lncRNAs are known to involve in the susceptibility of human cancers and might be the ideal candidate clinical biomarkers.10 Many lncRNAs were demonstrated to be associated with CC, including lncRNA-H19 and HOTAIR.5,6 Some lncRNAs, such as HOTTIP and CASC8, have been reported to be highly specific for cancer.11,12 With more and more transcriptomes studies of various cancer, more lncRNAs have been identified, indicating the crucial roles of lncRNAs in tumor prognosis and regulating tumorigenesis.13
Genome-wide association studies (GWAS) found the susceptible loci of specific tumor in lncRNAs.12,14,15 These data indicates that single nucleotide polymorphisms (SNPs) in lncRNA play important roles in the development of human cancers. The function of lncRNA SNPs is correlated to their located regions: SNPs located at the exons of lncRNAs are able to alter the folding of lncRNA or modify the binding affinity with their target genes and interacting elements; while SNPs located at the regulatory regions or the introns of lncRNAs might participate in the lncRNA splicing and the transcript regulation.16,17 In previous reports, SNPs in H19, HOTAIR, HOTTIP and CASC8 have been confirmed to be involved in cancer susceptibility.18–21 However, the association of lncRNA SNPs with CC susceptibility was not sufficiently studied. Thus, to investigate the association of the lncRNA SNPs with CC and cervical intraepithelial neoplasia (CIN), two SNPs located at the exon regions of lncRNA (rs217727 in H19 and rs2366152 in HOTAIR) and two SNPs located at the intron regions of lncRNA (rs1859168 in HOTTIP and rs10505477 in CASC8) were selected in the current study.
Materials and Methods
Study Population and Selection Criteria
Our study was approved by the Ethics Committee of Third Affiliated Hospital of Kunming Medical University, and all experiments were performed under the protocol of Helsinki’s Declaration. All individuals participating in the study were informed and signed a written informed consent form. A total of 1426 Chinese women (704 healthy individuals, 274 CIN patients, and 448 CC patients) were recruited at the Third Affiliated Hospital of Kunming Medical University from 2017 to 2019. Patients were diagnosed with CC and CIN based on histological diagnosis and were graded as our previous study.22 Patients who were diagnosed with other cancers before, or had a family history of CC, or were suffering from cardiovascular and infection disease, or had received chemotherapy or radiotherapy were excluded. Healthy individuals who had no family history of cancers and had no pathological changes in cervix were selected as the control of the current study from a physical examinations population at the same time. Afterward, we collected the clinical information of each individual, including age, tumor type, and clinical stage.
DNA Extraction
Five milliliters of blood was collected from the subjects for the extraction of genomic DNA. Then the genomic DNA was extracted using QIAamp DNA Blood Mini Kit (Qiagen, Germany) and finally frozen under −20°C after the concentration and purity was detected using NanoDrop 2000 (Thermo Scientific, USA).
SNP Genotyping
Agena MassArray System (Agena Bioscience, San Diego, CA, USA) was used to genotype of the 4 lncRNA SNPs (rs217727 in H19, rs2366152 in HOTAIR, rs1859168 in HOTTIP, rs10505477 in CASC8), and the primers for genotyping were designed using AssayDesigner 3.1 (Sequenom, USA) (Supplementary Table 1). The PCR conditions and program are described in Supplementary Table 2. Then, the PCR products purified by resin were transferred to a 384-well SpectroCHIP (Sequenom, USA) bioarray in a MassARRAY Nanodispenser RS1000 (Agena Bioscience, USA) as described in our previous study.23 The samples with three different genotypes (wild type homozygote, heterozygote and mutant homozygote) of each SNP were selected for sequencing to confirm the results of SNP genotyping.
Statistical Analysis
One-way ANOVA was performed for the analysis of age differences among different groups using GraphPad Prism 9.4 software. The allele and genotype distribution differences of these four SNPs among CIN, CC and control groups were analyzed by chi-square test using SHEsis online software.24,25 The association of these SNPs with CIN and CC was further evaluated using inheritance model analysis performed by SNPStats software.26 Five inheritance models were analyzed. The determination of the best inheritance models referred to the previous study: the smallest AIC and BIC values represent the best fit model. The sample size and statistical power was calculated using Power and Sample Size software.27 Multiple comparisons in the current study were corrected using Bonferroni correction. The significant different threshold was set at P<0.012 (0.05/4).
Results
Subject Characteristics
Table 1 lists clinical characteristics of the current population. The ages of the three groups exhibited no significant difference (P>0.05, F = 1.640). The ages between subgroups of CC (SCC: 47.47±9.67, AC: 46.96±10.23 and other: 49.87±9.93) were not significantly different (P>0.05, F = 0.5276).
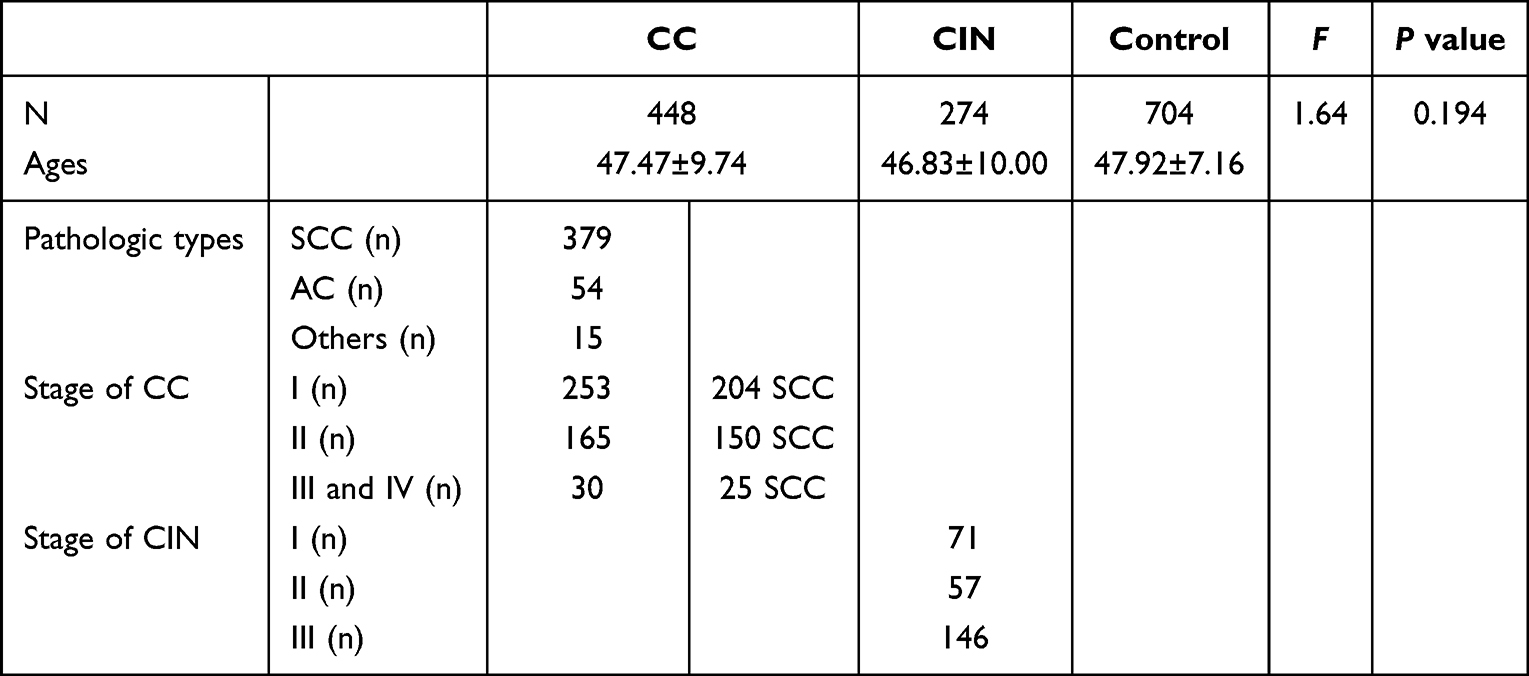 |
Table 1 The Clinical Characteristics of the Subjects Enrolled in the Current Study |
Distribution Analysis of the Four SNPs in CIN, CC and Control Groups
These four SNPs were all in HWE in three groups, except for rs1859168 in CIN group (Supplementary Table 3). The allelic distribution characteristics of these SNPs in the three groups were displayed in Table 2. Rs217727-A allele was markedly lower in control group compared with CC group after Bonferroni correction (P = 0.001, OR = 1.33; 95% CI: 1.12–1.58). While significant differences in distribution of this SNP were not observed in the comparison of CIN group with CC or Control groups.
 |
Table 2 The Allelic Distribution of the Four SNPs in the Three Groups |
The results of inheritance model analysis were displayed in Table 3. In the comparison of CC with control group, the log-additive model was the best fit model for rs217727. In this model, 2AA+GA genotype represented a certain risk of CC (P = 0.001, OR = 1.35; 95% CI: 1.13–1.62). However, no association of other SNPs with CIN or CC was found (P>0.012).
 |
Table 3 Inheritance Model Analysis of the Four SNPs Among CIN, CC and Control Groups |
Association of the Four SNPs with CC Pathological Types
Table 4 presents the distribution characteristics of the SNPs in different pathological type groups and control group. The results showed a significant difference in allelic frequencies of rs217727 between SCC and control groups (P = 0.002), indicating a risk role of allele A for SCC (OR = 1.33; 95% CI: 1.11–1.60). However, no SNPs showed significant association with AC (P>0.012).
 |
Table 4 The Allelic Distribution of the Four SNPs in Different Pathologic Types of CC and Control Groups |
The inheritance analysis results of different pathological types of CC and these SNPs are presented in Table 5. The 2AA + GA genotype of rs217727 significantly increased the risk of SCC (P = 0.002, OR = 1.35; 95% CI: 1.12–1.63) in log-additive model (best fitting model). However, no SNPs showed significant association with AC (P>0.012).
 |
Table 5 Inheritance Model Analysis of Four SNPs Between Different Pathologic Types of CC and Control Groups |
Association of Four SNPs with CC Clinical Stages
The SNPs distribution characteristics in different stage groups and control group are shown in Table 6. The allelic distribution of rs1859168 showed significantly different between the stage II and control groups (P = 0.003), indicating the protective effect of A allele of this SNP on CC (OR = 0.70; 95% CI: 0.54–0.89). A similar result for rs1859168 was also found between the stage II of SCC patients and controls (P = 0.005, OR = 0.69; 95% CI: 0.54–0.90) (Supplementary Table 4). The frequency of rs217727 allele A in stage I group was markedly higher than that in the control group (P = 0.005, OR = 1.35; 95% CI: 1.09–1.66). After Bonferroni correction, no other SNPs showed significant association with different stages of CC (P>0.012 Table 6 and Supplementary Table 4).
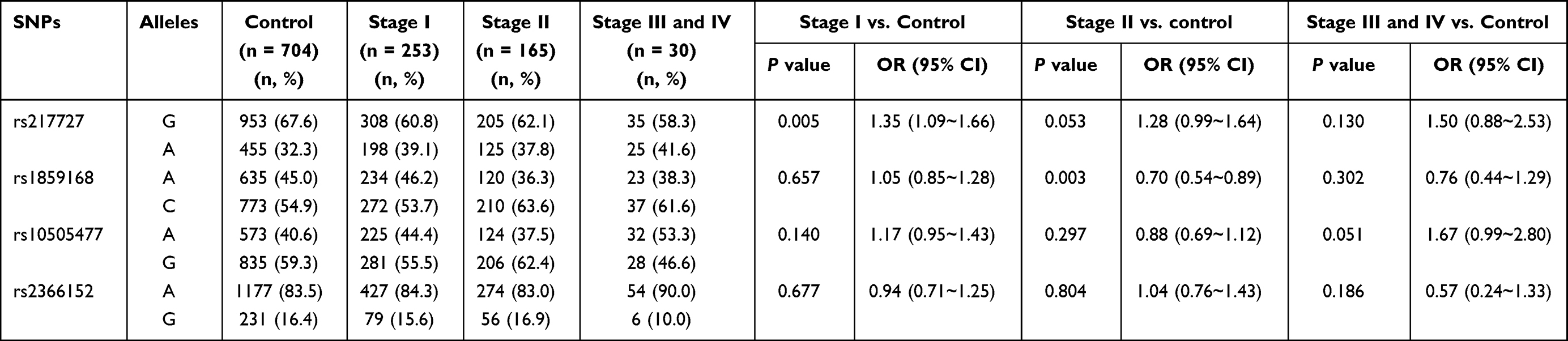 |
Table 6 The Allelic Distribution of SNPs in Different Stages of CC and Control Groups |
Inheritance model analysis of four SNPs in comparison of different stages of CC and SCC with control group was displayed in Table 7 and Supplementary Table 5. The results indicated that genotype 2AA+CA of rs1859168 was associated with lower risk of stage II CC (P = 0.004, OR = 0.68; 95% CI: 0.53–0.88). While, the 2AA+GA genotype of rs217727 was associated with higher risk of stage I CC (P = 0.005, OR = 1.36; 95% CI: 1.10–1.69).
 |
Table 7 Inheritance Model Analysis of SNPs Between Different Stages of CC and Control Groups |
Discussion
LncRNAs exhibit diverse expression patterns in various types of cancer.28 LncRNAs not only function as the key regulator of cell biological progress, but also exhibit a strong association with tumor cell phenotypes, and are involved in cancer initiation and progression.29–31
Paternally imprinted H19 gene, neighboring to IGF2 coding region, is located close to the 11p15.5 telomeric sequence.6,32 H19 was reported to be up-regulated in many human tumor types but lower in some normal adult tissues.33 Roychowdhury et al reported that H19 down-regulation may result in the dysregulation of numerous miRNAs and prompt cancer progress.34 H19 has been detected in most of CC tissues, and might involve in the proliferation and migration in CC35,36 through the interplay with miRNAs and other molecular.37 The association of SNPs in the H19 gene, among which rs217727 was the most frequently assessed, with the risk of human cancer has become a focus,38 which revealed the risk role of rs217727-AA or AA+GA genotype in lung cancer, bladder cancer and gastric cancer.39–41 Similarly, we demonstrated that the risk role of rs217727-A allele in CC, especially SCC in the current study. And the current results were also consistent with the study of Jin et al which reported that the minor allele (A) of this loci increases CC risk.42 However, Huang et al reported a contradictory results that rs217727 was not associated to CC risk.43 The inconsistent roles of rs217727 in different studies might be because of the different sample size of the different studies. Thus, to clarify the relationship of this SNP with CC, more association studies with larger sample size still need to be carried out in the future. Previous studies showed that rs217727 could influence the expression of a downstream cancer-related gene MRPL23‐AS1.44,45 Another study revealed that rs217727 could involve in the regulation of H19/miR-675/FADD/caspase-8/caspase-3/apoptosis signaling pathway, which makes it be associated with hepatic cell carcinoma.46 Furthermore, rs217727 may affect miRNA‒lncRNA interactions, leading to alteration of the miRNA (hsa-miR-4804-5p, hsa-miR-8071, hsa-miR-3960 and hsa-miR-8071) binding site on H19.47 Based on these data, it could be deduced that rs217727 may be associated with CC susceptibility through altering the related gene expression level and the interaction of H19 with relative miRNAs.42,48 However, functional studies still need to be carried out to clarify the association mechanisms of rs217727 with CC in the future.
HOTTIP, HOXA transcript at the distal tip, is localized at the 5′-end of the HOXA cluster on 7p15.2.11 Studies revealed roles of HOTTIP in human cancers (e.g., breast cancer and tongue squamous cell carcinoma).49–52 Recently, the association of rs1859168 located in HOTTIP gene regulation region with human cancers (pancreatic and gastric cancers) was studied, and the allele C of this SNP was related to lower risk of pancreatic cancer.53,54 In addition, rs1859168 was found to be associated with lung cancer through affecting HOTTIP folding.55 However, the opposite effect of this SNP was found in breast cancer, that the C allele functions as the risk factor.56 Similarly, in the current study, the A allele was identified with a protective role of CC in the stratified analysis implying the risk role of allele C. The inconsistent association results might be because of the complex role of rs1859168 in different types of cancer. And it also might be due to the different study population of these studies. Therefore, to clarify the role of rs1859168 in human cancers, more association studies of different cancers in different population with suitable sample size are needed in the future.
HOX transcript antisense RNA (HOTAIR) is the transcript of homeobox gene C cluster antisense strand.57–59 It is a general consensus that HOTAIR plays key roles in human cancers.13 S. Sharma et al found a significant association of rs2366152-C with CC cases with low HOTAIR levels in India,60 however, we did not find any association with CC in the current population. Cancer susceptibility candidate 8 (CASC8) is reported to be important in the development of various human cancers.13,61,62 GWAS and other association studies have reported that rs10505477 has an association with human cancers risk.63–65 However, our results suggest that rs10505477 was not associated with CIN and CC in the Chinese Han population. The reason for these inconsistent could be the different genetic background of the different studied population. For example, the frequency of rs2366152-C is 15% in Chinese Beijing Han people, whereas it is 34.5~37.7% in Indian people (data from 1000 Genomes Projects Phase 3).
In conclusion, rs217727 in H19 gene was associated with the susceptibility of CC risk in a Chinese Han population. The allele A of the loci was associated with a higher risk of CC, and the statistical power is 0.891, which indicated the credibility of the current results. There was a limitation of the current study that the clinical data of the current population was not incomplete, because of which the current study could not analyze the interplay of the genetic factor with other risk factors. The molecular mechanisms of the association between rs217727 and CC are still not clarified. Therefore, it is necessary to study the role of rs217727 in CC in the future, and further validation of the relationship with different pathological types is needed.
Data Sharing Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author Chuanyin Li.
Acknowledgments
Thanks are due to Dr. Yufeng Yao for the valuable work in formal analysis and writing review and editing. The funding projects that supported the current study are gratefully acknowledged: the CAMS Innovation Fund for Medical Sciences (CIFMS) (2021-I2M-2-004), National Science Foundation of China (82103190); Association Foundation Program of Yunnan Provincial Science and Technology Department and Kunming Medical University (202201AY070001-139); Yunnan Province Clinical Research Center for Gynecological and Obstetric Disease (2022YJZX-FC04/05/06). The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Cohen PA, Jhingran A, Oaknin A, Denny L. Cervical cancer. Lancet. 2019;393(10167):169–182. doi:10.1016/S0140-6736(18)32470-X
2. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA a Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
3. Xu H, Lin A, Shao X, Shi W, Zhang Y, Yan W. Diagnostic accuracy of high-risk HPV genotyping in women with high-grade cervical lesions: evidence for improving the cervical cancer screening strategy in China. Oncotarget. 2016;7(50):83775–83783. doi:10.18632/oncotarget.11959
4. Huang J, Deng Y, Boakye D, et al. Global distribution, risk factors, and recent trends for cervical cancer: a worldwide country-level analysis. Gynecol Oncol. 2022;164(1):85–92. doi:10.1016/j.ygyno.2021.11.005
5. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi:10.1158/0008-5472.CAN-16-2634
6. Aalijahan H, Ghorbian S. Long non-coding RNAs and cervical cancer. Exp Mol Pathol. 2019;106:7–16. doi:10.1016/j.yexmp.2018.11.010
7. Liu SJ, Dang HX, Lim DA, Feng FY, Maher CA. Long noncoding RNAs in cancer metastasis. Nat Rev Cancer. 2021;21(7):446–460. doi:10.1038/s41568-021-00353-1
8. Yao Z, Yang Y, Sun M, et al. New insights into the interplay between long non‐coding RNAs and RNA‐binding proteins in cancer. Cancer Commun. 2022;42(2):117–140. doi:10.1002/cac2.12254
9. Vitiello M, Tuccoli A, Poliseno L. Long non-coding RNAs in cancer: implications for personalized therapy. Cell Oncol. 2015;38(1):17–28. doi:10.1007/s13402-014-0180-x
10. Bartonicek N, Maag JLV, Dinger ME. Long noncoding RNAs in cancer: mechanisms of action and technological advancements. Mol Cancer. 2016;15(1):43. doi:10.1186/s12943-016-0530-6
11. Lian Y, Cai Z, Gong H, Xue S, Wu D, Wang K. HOTTIP: a critical oncogenic long non-coding RNA in human cancers. Mol BioSyst. 2016;12(11):3247–3253. doi:10.1039/C6MB00475J
12. Zhu Z, Gao X, He Y, et al. An insertion/deletion polymorphism within RERT-lncRNA modulates hepatocellular carcinoma risk. Cancer Res. 2012;72(23):6163–6172. doi:10.1158/0008-5472.CAN-12-0010
13. Cui Z, Gao M, Yin Z, Yan L, Cui L. Association between lncRNA CASC8 polymorphisms and the risk of cancer: a meta-analysis. CMAR. 2018;10:3141–3148. doi:10.2147/CMAR.S170783
14. Zheng Y, Lei T, Jin G, et al. LncPSCA in the 8q24.3 risk locus drives gastric cancer through destabilizing DDX5. EMBO Rep. 2021;22(11):e52707. doi:10.15252/embr.202152707
15. Betts JA, Moradi Marjaneh M, Al-Ejeh F, et al. Long noncoding RNAs CUPID1 and CUPID2 mediate breast cancer risk at 11q13 by modulating the response to DNA damage. Am J Hum Genet. 2017;101(2):255–266. doi:10.1016/j.ajhg.2017.07.007
16. McGeary SE, Lin KS, Shi CY, et al. The biochemical basis of microRNA targeting efficacy. Science. 2019;366(6472):eaav1741. doi:10.1126/science.aav1741
17. Novikova IV, Hennelly SP, Sanbonmatsu KY. Structural architecture of the human long non-coding RNA, steroid receptor RNA activator. Nucleic Acids Res. 2012;40(11):5034–5051. doi:10.1093/nar/gks071
18. Alzeer HS, Shaik JP, Reddy Parine N, et al. Genetic variants of HOTAIR associated with colorectal cancer: a case-control study in the Saudi Population. Genes. 2023;14(3):592. doi:10.3390/genes14030592
19. Cui P, Zhao Y, Chu X, et al. SNP rs2071095 in LincRNA H19 is associated with breast cancer risk. Breast Cancer Res Treat. 2018;171(1):161–171. doi:10.1007/s10549-018-4814-y
20. Abdi E, Latifi-Navid S, Zahri S, et al. SNP-SNP interactions of oncogenic long non-coding RNAs HOTAIR and HOTTIP on gastric cancer susceptibility. Sci Rep. 2020;10(1):16763. doi:10.1038/s41598-020-73682-0
21. Hu L, Chen S-H, Q-L L, et al. Clinical significance of long non-coding RNA CASC8 rs10505477 polymorphism in lung cancer susceptibility, platinum-based chemotherapy response, and toxicity. IJERPH. 2016;13(6):545. doi:10.3390/ijerph13060545
22. Li C, Li Y, Yan Z, et al. Polymorphisms in endoplasmic reticulum aminopeptidase genes are associated with cervical cancer risk in a Chinese Han population. BMC Cancer. 2020;20(1):341. doi:10.1186/s12885-020-06832-2
23. Chen K, Yan Z, Dong X, et al. Genetic polymorphisms in microRNA Genes Targeting PI3K/Akt signal pathway modulate cervical cancer susceptibility in a Chinese Population. Front Genet. 2022;13:856505. doi:10.3389/fgene.2022.856505
24. Yang J, Wang Y, Zhang S, et al. The association of TNF-α promoter polymorphisms with genetic susceptibility to cervical cancer in a Chinese Han Population. IJGM. 2022;15:417–427. doi:10.2147/IJGM.S350263
25. Li Z, Zhang Z, He Z, et al. A partition-ligation-combination-subdivision EM algorithm for haplotype inference with multiallelic markers: update of the SHEsis (http://analysis.bio-x.cn). Cell Res. 2009;19(4):519–523.
26. Sole X, Guino E, Valls J, Iniesta R, Moreno V. SNPStats: a web tool for the analysis of association studies. Bioinformatics. 2006;22(15):1928–1929. doi:10.1093/bioinformatics/btl268
27. Dupont WD, Plummer WD. PS power and sample size program available for free on the internet. Control Clin Trials. 1997;18(3):274. doi:10.1016/S0197-2456(97)00074-3
28. Qiu Y, Xu M, Huang S. Long noncoding RNAs: emerging regulators of normal and malignant hematopoiesis. Blood. 2021;138(23):2327–2336. doi:10.1182/blood.2021011992
29. Alvarez-Dominguez JR, Lodish HF. Emerging mechanisms of long noncoding RNA function during normal and malignant hematopoiesis. Blood. 2017;130(18):1965–1975. doi:10.1182/blood-2017-06-788695
30. Alvarez-Dominguez JR, Hu W, Gromatzky AA, Lodish HF. Long noncoding RNAs during normal and malignant hematopoiesis. Int J Hematol. 2014;99(5):531–541. doi:10.1007/s12185-014-1552-8
31. Alvarez-Dominguez JR, Hu W, Yuan B, et al. Global discovery of erythroid long noncoding RNAs reveals novel regulators of red cell maturation. Blood. 2014;123(4):570–581. doi:10.1182/blood-2013-10-530683
32. Cai X, Cullen BR. The imprinted H19 noncoding RNA is a primary microRNA precursor. RNA. 2007;13(3):313–316. doi:10.1261/rna.351707
33. Ghafouri-Fard S, Esmaeili M, Taheri M. H19 lncRNA: roles in tumorigenesis. Biomed Pharmacother. 2020;123:109774. doi:10.1016/j.biopha.2019.109774
34. Roychowdhury A, Samadder S, Das P, et al. Deregulation of H19 is associated with cervical carcinoma. Genomics. 2020;112(1):961–970. doi:10.1016/j.ygeno.2019.06.012
35. Feigenberg T, Gofrit ON, Pizov G, Hochberg A, Benshushan A. Expression of the H19 oncofetal gene in premalignant lesions of cervical cancer: a potential targeting approach for development of nonsurgical treatment of high-risk lesions. ISRN Obstet Gynecol. 2013;2013:1–7. doi:10.1155/2013/137509
36. Iempridee T. Long non-coding RNA H19 enhances cell proliferation and Anchorage-independent growth of cervical cancer cell lines. Exp Biol Med. 2017;242(2):184–193. doi:10.1177/1535370216670542
37. Cáceres-Durán MÁ, Ribeiro-dos-Santos Â, Vidal AF. Roles and mechanisms of the long noncoding RNAs in cervical cancer. IJMS. 2020;21(24):9742. doi:10.3390/ijms21249742
38. Wang X, Zhong J, Chen F, et al. Association between lncRNA H19 rs217727 polymorphism and the risk of cancer: an updated meta-analysis. BMC Med Genet. 2019;20(1):186. doi:10.1186/s12881-019-0904-x
39. Li L. Association between H19 SNP rs217727 and lung cancer risk in a Chinese population: a case control study. BMC Med Genet. 2018;19(1):1–6.
40. Yang C, Tang R, Ma X, et al. Tag SNPs in long non-coding RNA H19 contribute to susceptibility to gastric cancer in the Chinese Han population. Oncotarget. 2015;6:15311–15320.
41. Hua Q, Lv X, Gu X, et al. Genetic variants in lncRNA H19 are associated with the risk of bladder cancer in a Chinese population. Mutagenesis. 2016;31(5):531–538. doi:10.1093/mutage/gew018
42. Jin T, Wu X, Yang H, et al. Association of the miR-17-5p variants with susceptibility to cervical cancer in a Chinese population. Oncotarget. 2016;7(47):76647–76655. doi:10.18632/oncotarget.12299
43. Huang M-C, Chou Y-H, Shen H-P, et al. The clinicopathological characteristic associations of long non-coding RNA gene H19 polymorphisms with uterine cervical cancer. J Cancer. 2019;10(25):6191–6198. doi:10.7150/jca.36707
44. Tan T, Li J, Wen Y, et al. Association between lncRNA- H19 polymorphisms and hepatoblastoma risk in an ethic Chinese population. J Cell Mol Med. 2021;25(2):742–750. doi:10.1111/jcmm.16124
45. Yuan Z, Yu Y, Zhang B, et al. Genetic variants in lncRNA H19 are associated with the risk of oral squamous cell carcinoma in a Chinese population. Oncotarget. 2018;9(35):23915–23922. doi:10.18632/oncotarget.23673
46. Ge L, Wang Q, Hu S, Yang X. Rs217727 polymorphism in H19 promotes cell apoptosis by regulating the expressions of H19 and the activation of its downstream signaling pathway. J Cell Physiol. 2019;234(5):7279–7291. doi:10.1002/jcp.27485
47. Deng Y, Zhou L, Yao J, et al. Associations of lncRNA H19 polymorphisms at MicroRNA binding sites with glioma susceptibility and prognosis. Mol Ther Nucleic Acids. 2020;20:86–96. doi:10.1016/j.omtn.2020.02.003
48. Matouk IJ, DeGroot N, Mezan S, et al. The H19 non-coding RNA is essential for human tumor growth. PLoS One. 2007;2(9):e845. doi:10.1371/journal.pone.0000845
49. Ghafouri-Fard S, Dashti S, Taheri M. The HOTTIP (HOXA transcript at the distal tip) lncRNA: review of oncogenic roles in human. Biomed Pharmacother. 2020;127:110158. doi:10.1016/j.biopha.2020.110158
50. Yang Y, Qian J, Xiang Y, Chen Y, Qu J. The prognostic value of long noncoding RNA HOTTIP on clinical outcomes in breast cancer. Oncotarget. 2017;8(4):6833–6844. doi:10.18632/oncotarget.14304
51. Sun Y, Zeng C, Gan S, et al. LncRNA HOTTIP-mediated HOXA11 expression promotes cell growth, migration and inhibits cell apoptosis in breast cancer. IJMS. 2018;19(2):472. doi:10.3390/ijms19020472
52. Zhang H, Zhao L, Wang Y-X, Xi M, Liu S-L, Luo -L-L. Long non-coding RNA HOTTIP is correlated with progression and prognosis in tongue squamous cell carcinoma. Tumor Biol. 2015;36:8805–8809.
53. Hu P, Qiao O, Wang J, et al. rs1859168 A > C polymorphism regulates HOTTIP expression and reduces risk of pancreatic cancer in a Chinese population. World J Surg Onc. 2017;15(1):155. doi:10.1186/s12957-017-1218-0
54. Duan F, Huang X, Shuang Z. Functional long non-coding RNAs associated with gastric cancer susceptibility and evaluation of the epidemiological efficacy in a central Chinese population. Cancer Medicine. 2018;7(4):1285–1296. doi:10.1002/cam4.1318
55. Gong W-J, Yin J-Y, X-P L, et al. Association of well-characterized lung cancer lncRNA polymorphisms with lung cancer susceptibility and platinum-based chemotherapy response. Tumor Biol. 2016;37(6):8349–8358. doi:10.1007/s13277-015-4497-5
56. Abdelaleem OO, Shaker OG, AbdelHafez MN, et al. The influence of rs1859168 polymorphism on serum expression of HOTTIP and its target miR-615-3p in Egyptian patients with breast cancer. Biomolecules. 2021;11(5):733. doi:10.3390/biom11050733
57. Qu X, Alsager S, Zhuo Y, Shan B. HOX transcript antisense RNA (HOTAIR) in cancer. Cancer Lett. 2019;454:90–97. doi:10.1016/j.canlet.2019.04.016
58. Silva RA, Ferreira MR, Gomes AM, Zambuzzi WF. LncRNA HOTAIR is a novel endothelial mechanosensitive gene. J Cell Physiol. 2020;235(5):4631–4642. doi:10.1002/jcp.29340
59. Xin X, Li Q, Fang J, Zhao T. LncRNA HOTAIR: a potential prognostic factor and therapeutic target in human cancers. Front Oncol. 2021;11:679244. doi:10.3389/fonc.2021.679244
60. Sharma Saha S, Roy Chowdhury R, Mondal NR, et al. Identification of genetic variation in the lncRNA HOTAIR associated with HPV16-related cervical cancer pathogenesis. Cell Oncol. 2016;39(6):559–572. doi:10.1007/s13402-016-0298-0
61. Yang B, Gu B, Zhang J, Xu L, Sun Y. CASC8 lncRNA promotes the proliferation of retinoblastoma cells through downregulating miR34a methylation. CMAR. 2020;12:13461–13467. doi:10.2147/CMAR.S268380
62. Wu Q, Zhang H, Yang D, et al. The m6A-induced lncRNA CASC8 promotes proliferation and chemoresistance via upregulation of hnRNPL in esophageal squamous cell carcinoma. Int J Biol Sci. 2022;18(13):4824–4836. doi:10.7150/ijbs.71234
63. Kim T, Cui R, Jeon Y-J, et al. Long-range interaction and correlation between MYC enhancer and oncogenic long noncoding RNA CARLo-5. Proc Natl Acad Sci USA. 2014;111(11):4173–4178. doi:10.1073/pnas.1400350111
64. Haerian M Sadat, Baum L, Haerian B Sadat. (2011). Association of 8q24.21 loci with the risk of colorectal cancer: a systematic review and meta-analysis. J Gastroenterol Hepatol, 26(10), 1475–84. 10.1111/j.1440-1746.2011.06831.x
65. Ma G et al . (2015). Genetic variant in 8q24 is associated with prognosis for gastric cancer in a Chinese population. J Gastroenterol Hepatol, 30(4), 689–95. 10.1111/jgh.12801
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.