")
Back to Journals » Infection and Drug Resistance » Volume 16
Role of Proviral HIV-1 DNA Genotyping for People Living with HIV (PLWH) Who Had Low-Level Viremia While Receiving Antiretroviral Therapy
Authors Lv S, Sun L, Li T, Bai R, Dai M, Wang R, Zhai Y, Hua W, Li A, Xin R, Dai L
Received 28 January 2023
Accepted for publication 10 June 2023
Published 19 July 2023 Volume 2023:16 Pages 4697—4706
DOI https://doi.org/10.2147/IDR.S406188
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Suresh Antony
Shiyun Lv,1,* Lijun Sun,1,* Tongzeng Li,1,* Ruojing Bai,1 Man Dai,2,3 Ran Wang,1 Yuanyi Zhai,1 Wei Hua,1 Aixin Li,1 Ruolei Xin,2 Lili Dai1
1Beijing Youan Hospital, Capital Medical University, Beijing, People’s Republic of China; 2Institute for STD/AIDS Prevention and Treatment, Beijing Center for Disease Prevention and Control, Beijing, People’s Republic of China; 3School of Public Health, Chinese Medical University, Shenyang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lili Dai, Beijing Youan Hospital, Capital Medical University, Beijing, 100069, People’s Republic of China, Tel +86 10 83911069, Email [email protected] Ruolei Xin, Institute for STD/AIDS Prevention and Treatment, Beijing Center for Disease Prevention and Control, Beijing, 100013, People’s Republic of China, Tel +86 10 64407364, Email [email protected]
Objective: To analyze the antiretroviral resistance in people living with HIV (PLWH) who developed low-level viremia (LLV) during antiretroviral therapy (ART) via sequencing of their HIV-1 proviral DNA and RNA and comparisons of their proviral DNA genotyping data with their past and synchronous RNA genotyping data.
Patients and Methods: PLWH with LLV while receiving ART for 6 months or longer from January 2020 to September 2021 were included. HIV-1 proviral DNA and RNA were extracted from white-blood cells and concentrated plasma by ultracentrifugation, respectively, and HIV-1 pol gene fragments were amplified and sequenced. The concordance in the detection of resistance-associated mutations (RAMs) were examined between proviral DNA vs past RNA genotyping and proviral DNA vs synchronous RNA genotyping.
Results: Of the 150 PLWH with LLV, 117 proviral DNA pol sequences detected in 105 PLWH were successfully amplified and RAMs were present in 27.6% and the rate of RAMs conferring low-level or greater resistance to antiretrovirals examined was 17.1%. Fifty-six and 57 PLWH had results of past and synchronous RNA genotyping, respectively, for comparisons with those of proviral DNA genotyping; and the concordance rates were 76.8% and 75.4%, respectively. However, proviral DNA genotyping lost than gained partial information on antiretroviral resistance compared with past or synchronous RNA genotyping.
Conclusion: We found that the concordance between proviral DNA and past and synchronous RNA genotyping was moderate. Proviral DNA genotyping lost than gained more information on antiretroviral resistance compared with past or synchronous RNA genotyping. To optimize ART in PLWH with LLV, antiretroviral resistance profile should be interpreted in combination with proviral DNA and RNA genotyping and a comprehensive review of previous treatment history.
Keywords: genotypic resistance testing, resistance-associated mutation, nucleoside reverse-transcriptase inhibitor, non-nucleoside reverse-transcriptase inhibitor, protease inhibitor
Introduction
Antiretroviral therapy (ART) can effectively suppress HIV-1 replication and maintain plasma viral load (VL) below the detection limit for most of the people living with HIV (PLWH) who adhere to the regimens prescribed,1 which helps reduce morbidity, mortality, and HIV-1 transmission, and consequently result in improving life expectancy of PLWH. However, in approximately 3–10% of PLWH receiving ART, low-level viremia (LLV) may occur.2 LLV usually refers to VL of 50–1000 copies/mL after ART.3 Although LLV is not equivalent to virological failure (VF), it was reported that the presence of LLV may result in emergence of antiretroviral resistance and increased risk for VF.4,5 Therefore, standard genotyping resistance testing (GRT) in PLWH with LLV can provide clinicians with additional valuable information for designing more specific and effective antiretroviral treatment strategies to reduce the risk of VF.
The Department of Health and Human Services (DHHS) guidelines suggest that PLWH with virologic failure (VF) and HIV-RNA levels >200 copies/mL undergo HIV drug-resistance testing to assist the selection of active antiretrovirals.6 The World Health Organization (WHO) guidelines recommend conducting GRT when VL is greater than 1000 copies/mL during ART and adjustment of treatment regimens based on the results of GRT.7 However, when VL is <1000 copies/mL, the low concentration of virus in plasma significantly reduces the success rate with conventional GRT. Therefore, there are challenges to the conventional genotyping in PLWH with LLV.
One possible approach to detection of antiretroviral resistance in PLWH with LLV is to sequence proviral HIV-1 DNA. Some studies have shown that proviral DNA sequencing cannot identify all resistance-associated mutations (RAMs) compared with RNA sequencing after ART initiation,8,9 while other studies indicated a good agreement between the two methods and suggested that RAMs detected by proviral DNA GRT could predict VF.10,11 In this study, we aimed to compare the results of proviral DNA genotyping with those of RNA genotyping performed on the same day and those of historical RNA genotyping to investigate the concordance rates between proviral DNA genotyping and RNA genotyping and whether proviral DNA genotyping can provide additional information.
Materials and Methods
Study Participants
PLWH who had received ART for at least 6 months were recruited at Beijing Youan Hospital in January 2020 and were followed until September 2021. The inclusion criteria were as follows: PLWH having at least two documented VL testing; and having at least one VL of 50–1000 copies/mL during follow-up. Exclusion criteria were PLWH without follow-up after ART initiation; and those without important information such as HIV transmission route, baseline CD4+T cell count and VL, history of previous VF, time since ART initiation, number of antiretrovirals since ART initiation, number of past RNA genotyping, and follow-up time et al.
The study was approved by the Research Ethics Committee of Beijing Youan Hospital, Capital Medical University (approval number: LL-2020-127-K) and the study conduct adhered to the Declaration of Helsinki. All PLWH participating in this study provided written informed consent.
Data Collection
Demographic and clinical data, such as age, gender, HIV transmission route, time since ART initiation, VL and CD4+T cell count at baseline and after ART, and the number of antiretrovirals since ART initiation were collected from the database of the National Free Antiretroviral Treatment Program. The results of previous HIV-1 RNA genotyping tests were retrieved from hospital medical records.
LLV was defined as having at least once VL of 50–1000 copies/mL after 6 months of ART. Persistent LLV (pLLV) was defined as two or more consecutive VL of 50–1000 copies/mL, at least 1 month apart, and otherwise intermittent LLV (iLLV/blip).12
Sample Collection and HIV-1 viral load testing
All included PLWH were routinely followed for VL testing every 6 to 12 months after ART initiation. If more than one episode of LLV occurred, only the first episode of LLV was included for analysis. The subsequent follow-up visit after LLV occurred was every 3 months. The observation continued until loss to follow-up, occurrence of VF (defined as VL >1000 copies/mL at least once after 6 months of ART), death or end date of this study on 31 September 2021, whichever occurred earlier.
The peripheral venous blood was collected at each follow-up visit and the sample was centrifuged at 3000 r/min for 10 min at room temperature within 6 hours. Plasma and the concentrated white-blood cells (buffy coat) were frozen at −80°C for further testing. Abbott Real-Time HIV-1 RNA Test was performed with the Abbott molecular m2000 system (Abbott Diagnostics Inc, USA), according to the manufacturer’s instructions.
HIV-1 Pol Gene Amplification
DNA extraction and PCR amplification were performed as follows: HIV-1 DNA was extracted from buffy coat using a DNA Extraction kit (SUPBIO, Guangzhou) for pol gene amplification with the use of nested polymerase chain reaction (PCR). The target pol gene fragments included the entire protease (PR 1–99 aa) and mostly reverse transcriptase (RT, 1–320 aa) regions. The amplified products were detected by 1% agarose gel electrophoresis, after which positive amplicons were sent to Beijing SinoGenMax Limited Company for Sanger sequencing.
RNA extraction and RT-PCR amplification were performed as follows: about 1 mL of plasma was dispensed into 2 mL Eppendorf centrifuge tubes and then centrifuged at 20,000×g for 1h at 4°C. Next, the supernatant was gently removed, leaving about 50 µL of the concentrated solution at the bottom; 120 µL of Abbott m2000 lysis solution was added, lysing for 10 min for RNA extraction. HIV-1 RNA was extracted from plasma using QIAamp Viral RNA Mini Kit (Qiagen, USA) for pol gene amplification using reverse transcription and nested-PCR. The remaining steps were the same as DNA. Sequences of primers for amplification are shown in Table S1.
Analysis of Subtype sequences and Detection of RAMs
Sequence data were edited and assembled with Sequencher version 5.0, and the pol sequences were manually corrected with BioEdit software version 7.2.6.1. HIV-1 subtyping was performed using the COMET HIV-1 (https://comet.lih.lu). RAMs and the levels of resistance were interpreted using the Stanford University HIV Drug-Resistance Database (https://hivdb.stanford.edu). According to the results, HIV-1 drug resistance levels were divided into five categories: susceptible, potential low-level resistance, low-level resistance, intermediate resistance, and high-level resistance. Low-level or above resistance was considered drug resistance in this study.
Statistical Analysis
For continuous variables, descriptive statistics are presented as medians with interquartile ranges (IQRs), and for categorical variables, as counts and proportions. A scoring system was used in order to assess the information gained or lost by proviral DNA genotyping results. The detailed information was as follows: compared with RNA genotyping results, RAM identified in RNA but not in proviral DNA genotyping was defined as a score of −1. Conversely, if RAM found in proviral DNA was not in RNA genotyping, the score was 1. A score of 0 indicated that RAM was consistent both in proviral DNA and RNA genotyping. An increase or decrease in scores indicated the amount of information gained or lost from proviral DNA genotyping compared with RNA genotyping. If the same individual had successful amplification of several sequences at different time points, the resistance analysis would mainly focus on the sequences with more information. All statistical analyses were performed with SPSS (IBM SPSS Statistics, version 26.0).
Results
Study Population
Among 10693 PLWH who were screened, 228 (2.1%) experienced LLV during follow-up; after excluding those who were unable or unwilling to provide informed consent, 150 were included in the analysis (Figure 1). Among them, 49 patients (32.7%) developed pLLV and 101 (67.3%) developed iLLV/blip. Table 1 shows the characteristics of the included PLWH. Of those 150 PLWH, 142 (94.7%) were male and their median age was 37 years (IQR, 32–46). The median ART duration was 3.12 years (IQR 1.10–5.23) and the median follow-up time was 5.13 months (range, 2.97 to 8.12). One hundred and seventeen (78.0%) PLWH had been receiving regimens containing 2 nucleoside reverse-transcriptase inhibitors (NRTIs) plus 1 non-nucleoside reverse-transcriptase inhibitor (NNRTI) when LLV occurred and 22 (14.7%) had experienced VF before developing LLV. PLWH had received a median of 1 (IQR 1–2) RNA genotyping test from ART initiation before proviral DNA genotyping. Finally, 7 PLWH (4.7%) developed VF, and 3 (2.0%) were lost to follow-up or transferred out of care.
 |
Table 1 Characteristics of the included people living with HIV (PLWH) who presented with low-level viremia |
 |
Figure 1 Study flow. Abbreviations: ART, antiretroviral therapy; LLV, low-level viremia; NJ, neighbor-joining; VL, viral load. |
DNA Samples Amplification
A total of 154 blood samples were collected from 150 PLWH, including 137 samples with VL 50–200 copies/mL, 9 with VL 201–400 copies/mL, and 8 with VL 401–1000 copies/mL. In total, 125 proviral DNA sequences and 95 ultracentrifuged synchronous RNA sequences were obtained after amplifying 154 samples with VL of 50–1000 copies/mL. By combining past RNA sequences, ultracentrifuged synchronous RNA sequences, and proviral DNA sequences, the phylogenetic trees were constructed using the neighbor-joining (NJ) method to determine whether different sequences originating from the same individual in order to eliminate potential risk of laboratory contamination. Eight proviral DNA sequences that were not clustered with other sequences in the phylogenetic trees were removed, which left 117 DNA sequences from 105 PLWH for analysis, giving an overall amplification success rate of 76.0%. The entire analysis flow is presented in Figure 1 and the details of amplification are shown in Table 2. Evolutionary tree is presented in Figure S1.
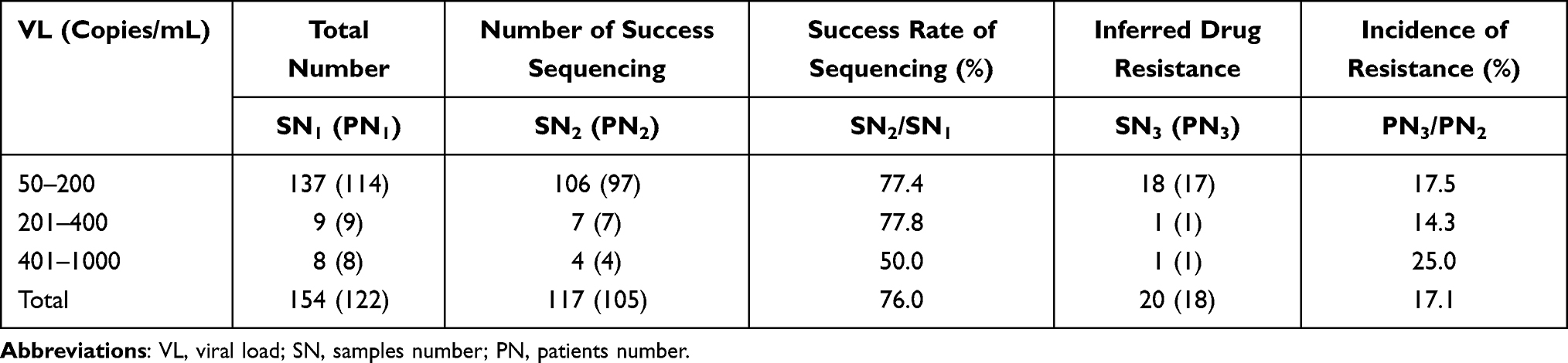 |
Table 2 Sequencing results of DNA samples |
RAMs Detected in Proviral DNA Genotyping
RAMs were identified in 29 out of 105 PLWH, with an overall resistance mutation rate of 27.6%. RAMs to NNRTIs accounted for 22.9% (24/105), with the most common mutations being V106I/M and V179D/E/L, which accounted for 10.5% (11/105) and 11.4% (12/105), respectively. The proportion of RAMs to NRTIs and protease inhibitors (PIs) was 10.5% (11/105) and 4.8% (5/105), respectively, with the most common RAMs to NRTIs being M184V/I (7.6%, 8/105), and those to PIs being M46L and L33F each accounting for 1.9% (2/105).
Resistance to Commonly Used ARVs
According to the Stanford University HIV Drug-Resistance Database, 18 RAMs (17.1%, 18/105) were categorized as conferring low-level or above resistance and 11 (10.5%, 11/105) potential resistance. RAMs conferring low-level or above resistance to zidovudine, lamivudine, and tenofovir disoproxil fumarate accounted for 1.9% (2/105), 7.6% (8/105), and 1.9% (2/105), respectively. RAMs to efavirenz and nevirapine accounted for 9.5% (10/105) and 10.5 (11/105), respectively, while RAMs to ritonavir-boosted lopinavir were not found.
Detection of Antiretroviral Resistance by Proviral DNA and RNA Genotyping
After sequence alignment by phylogenetic analysis, 57 of 105 (54.3%) PLWH with successful proviral DNA genotyping had simultaneously obtained RNA sequences from the same blood sample, and 56 (53.3%) PLWH had past RNA sequences for analysis, which included 35 before initiation of ART, 11 who developed VF during ART, and 10 who developed LLV during ART. The median interval between the performance of past RNA and proviral DNA genotyping was 1.66 years (IQR 0.77–3.02). Therefore, two groups were defined for comparisons: 56 PLWH with both proviral DNA and past RNA genotyping (Group 1); and 57 PLWH with both proviral DNA and RNA genotyping obtained on the same day (Group 2).
As shown in Figure 2, a total score of 0 indicated that, compared with RNA genotyping, proviral DNA genotyping results showed no increase or decrease in RAMs. The concordance rates of RAMs for Groups 1 and 2 were 76.8% (43/56) and 75.4% (43/57), respectively. There were more PLWH with loss than gain in the resistance information (Group 1: score of <0, 16.1% and score of >0, 7.1%; Group 2: score of <0, 21.1% and score of >0, 3.5%) with proviral DNA genotyping compared to compared with past or synchronous RNA genotyping.
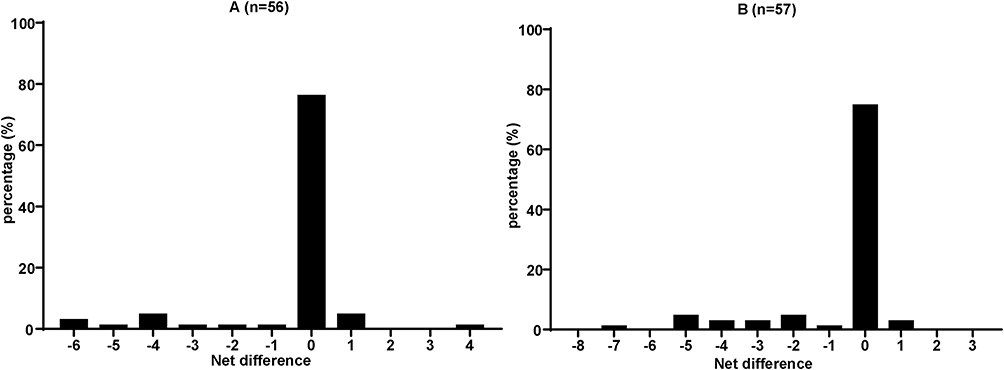 |
Figure 2 Histogram of the net change in resistance information between HIV-1 DNA and RNA genotyping results. (A) Group 1 and (B) Group 2. The score of 0 indicates that drug resistance mutation (DRM) was consistent both in proviral DNA and RNA genotyping results. The score of <0 indicates that, compared with past RNA, proviral DNA genotyping lost information on resistance-associated mutations; conversely, the score of >0 indicates that proviral DNA genotyping gained information. |
Comparisons of RAMs to individual antiretrovirals are summarized in Figure 3A for group 1 and Figure 3B for group 2. As shown in Figure 3A, compared with past RNA genotyping, the loss of information on RAMs in proviral DNA genotyping could be found within all classes of antiretrovirals, which is also shown in Figure 3B.
 |
Figure 3 The percentage of proviral HIV-1 DNA genotyping yielding resistance-associated mutations conferring resistance to individual antiretroviral is compared with that past RNA genotyping or synchronous RNA genotyping. (A) Group 1 and (B) Group 2. Abbreviations: ABC, abacavir; AZT, zidovudine; FTC, emtricitabine; 3TC, lamivudine; TDF, tenofovir disoproxil fumarate; DOR, doravirine; EFV, efavirenz; ETR, etravirine; NVP, nevirapine; RPV, rilpivirine; ATV/r, ritonavir-boosted atazanavir; DRV/r, ritonavir-boosted darunavir; IDV/r, ritonavir-boosted indinavir; LPV/r, ritonavir-boosted lopinavir. |
Discussion
Currently, two hypotheses have been proposed to explain the underlying mechanisms of LLV. One suggests that low-level viral replication is probably derived from resting CD4+T lymphocytes as the HIV-1 reservoir. These latently infected CD4+T lymphocytes become activated, resulting in clonal expansion to produce virus particles.13,14 Another hypothesis is that ART may not completely suppress viral replication, and the insufficient antiretroviral concentrations in the sanctuary site may lead to active virus replication.15,16 At the same time, studies have also confirmed that the emergence of new RAMs in PLWH with lower VL might be caused by continuous viral replication,17 and RAMs detected in PLWH with LLV could be a risk factor for VF.4,5 Therefore, understanding the characteristics of antiretrovrial resistance in PLWH with LLV will be beneficial in guiding the clinicians in optimizing treatment regimens to reduce the risk of subsequent VF.
The incidence of LLV varies from region to region. In a retrospective study of 2795 PLWH in the United States from 2005 to 2015,18 9.4% of PLWH experienced LLV defined as VL of 51–500 copies/mL. In a national observational study of 6956 PLWH in Sweden, 9% experienced LLV during the follow-up for an average of 5.7 years.19 The discrepancies in incidence may be related to the different clinical characteristics of the study populations, ART used, definitions of LLV used and VL testing frequency. The definitions used by major international HIV treatment guidelines vary, which should be considered when interpreting the LLV data presented. For example, the WHO guidelines,7 International Antiviral Society (IAS USA) guidelines,20 and DHHS guidelines6 define LLV as VL of 50–1000 copies/mL, 50–200 copies/mL, and less than 200 copies/mL, respectively, when PLWH continue to receive ART.
When PLWH develop LLV, conventional genotyping methods are often unsuccessful because the viral RNA levels are often too low for detection of RAMs. The proviral DNA can be viewed as an archive of viral quasispecies in PLWH.21 Previous studies have found a good agreement between past RNA and proviral DNA genotyping results in PLWH who did not develop VF,11,22,23 and some additional information can be obtained from proviral DNA genotyping.24 However, research suggesting the lack of sensitivity of proviral DNA genotyping compared with historical cumulative RNA genotyping remains limited.8,9
In this study, we used ultracentrifugation of blood samples for RNA genotyping and, at the same time, we combined the sequences obtained from past RNA genotyping to evaluate the concordance of proviral DNA genotyping with RNA genotyping. We found that the rate of RAMs detected by proviral DNA genotyping was present in 27.6% of the included PLWH with LLV, and the rate of RAMs conferring low-level resistance or greater to antiretrovirals was 17.1%. Compared with the results of synchronous RNA or with past RNA genotyping, we also found that more PLWH had a loss than gain of information on RAMs provided by proviral DNA genotyping, and the loss of information on RAMs in proviral DNA genotyping could be found for all classes of antiretrovirals.
In theory, compared to the information on RAMs obtained by RNA genotyping, that obtained by proviral DNA genotyping will be more comprehensive. The HIV-1 continuously accumulates secondary RAMs when it replicates under the selective pressure of antiretrovirals. After treatment interruption, replicating HIV-1 may gradually switch to more adaptable wild strains, which results in the continuous attenuation of drug-resistant quasispecies into minor drug-resistant strains.25 As the conventional Sanger sequencing can only detect major HIV-1 strains, the information on RAMs of the minor virus strains with RAMs, which account for less than 15–25%, are often missed.26 Some minor RAMs may be found using deep sequencing with higher sensitivity; however, this approach may not be routinely available in clinical laboratories due to the high cost and operational complexity.27 As proviral DNA is an archive of HIV quasispecies,21 it is often considered to provide more information on RAMs. Moreover, APOBEC3 cytidine deaminases are known to induce G to A hypermutation of GG or GA dinucleotides in DNA,28 which might also result in proviral DNA genotyping to gain more mutation information.29,30 These mutations are frequently associated with hypermutated sequences and represent the majority of defective proviruses in HIV-1-infected host,31 while a previous study has found that the concordance in detecting RAMs between proviral DNA and RNA genotyping is compromised in the presence of defective proviruses.32
In this study, we found that in 10 individuals, RAMs were detected in the proviral DNA genotyping but not in the previous and current RNA genotyping. At the same time, three individuals in our study showed polymorphic accessory NNRTI-selected mutation V179D, and the other three showed non-polymorphic mutation I47L, K70T, and M46L, which contributes to the decreases in antiretroviral susceptibility; however, it remains unclear the extent to which these mutations might affect the effectiveness of future treatment options. Compared to RNA genotyping, we found more resistance information was lost by proviral DNA genotyping. Longitudinal analyses have also found that RAMs in plasma HIV-1 emerge more than a year earlier before being found in peripheral blood mononuclear cells (PBMCs).33 In contrast, because genetic changes in proviral DNA occur more slowly than HIV-1 in plasma, plasma HIV-1 is more sensitive to changes in antiretroviral selection pressure.34 Therefore, emerging RAMs may be present in plasma HIV-1 but not yet expressed in proviral DNA.
Furthermore, as the drug-resistant quasispecies transfer to minor drug-resistant strains in plasma, the storage capacity of RAMs in proviral DNA tends to decrease gradually.35 It has been hypothesized that archived resistant provirus-infected cells could be diluted by more recently uninfected cells in PLWH who were on effective ART, thus making RAMs less easily detectable in proviral DNA. In the present study, the incidence of RAMs detected in proviral DNA genotyping was low, and there was a certain degree of loss of information on antiretroviral resistance when compared with past RNA and current RNA genotyping results as 90.8% of the samples with successful amplification in our study had VL of 50–200 copies/mL.
It was reported that a longer duration of ART and the number of cumulative antiretrovirals used could account for the loss of information on RAMs by proviral DNA genotyping.29 In the present study, when comparing the results of proviral DNA with those of past and ultracentrifuged RNA genotyping by multivariable logistic regression analysis, we were not able to identify any factors associated with the loss of information on RAMs by proviral DNA genotyping (P>0.05). We speculate that this could be contributed to the smaller sample size of our study.
In this study, we also found that the information obtained by synchronous RNA genotyping was greater than that by past RNA genotyping. It can be speculated that continuous active viral replication in PLWH with LLV might cause new RAMs to emerge during ART, which was found to be a risk factor for VF in previous studies.4,5
The present study has several limitations. Firstly, because of a relatively small sample size, the power in the detection of factors associated with the difference in proviral DNA and RNA genotyping results is significantly limited. Secondly, we did not obtain the blood samples when the included PLWH first developed VL 50–1000 copies/mL, but instead, we obtained the blood samples after 3–6 months of follow-up; the RAMs detected might not fully reflect the drug resistance characteristics associated with LLV in real time. Thirdly, limited by the sample size in this study, no separate analysis was conducted for PLWH with pLLV that only accounted for 32.7% (n = 49), although pLLV might have a greater impact on clinical management. Fourthly, we did not adopt some technologies such as single-genome amplification (SGA) to minimize the loss of information on RAMs by the population sequencing. Finally, detection of RAMs to integrase strand-transfer inhibitors (INSTIs) was not performed in this study because only a small proportion of included PLWH were receiving INSTI-based regimens. Additional studies are warranted to investigate the concordance between proviral DNA and RNA genotyping as INSTI-based regimens have become the recommended first-line therapy in major HIV treatment guidelines.6
Conclusion
We conclude that while proviral DNA genotyping may provide some more information on RAMs that were not detected by RNA genotyping, the concordance between proviral DNA and past and synchronous RNA genotyping was moderate. Proviral DNA genotyping lost more information on antiretroviral resistance compared with past or synchronous RNA genotyping. To optimize treatment strategies for PLWH with LLV, comprehensive evaluation of the data obtained by proviral DNA and RNA genotyping and antiretroviral treatment history is needed.
Acknowledgment
We thank Supriya Mahajan for designing our subject and revising the manuscript.
Funding
This work was supported by the Beijing Municipal of Science and Technology Major Project (Z211100002921003 to L.D.), Beijing Natural Science Foundation (7222092 to L.D.), Capital’s Funds for Health Improvement and Research (CFH2022-1G-3015 to R.X.) and Fengtai District Health System Research Project (2020-106 to A.L.). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Disclosure
The authors report no conflicts of interest in this work. The abstract of this paper was presented at the HIV Glasgow 2022 as a poster with interim findings. The poster’s abstract was published in “Poster Abstracts” in Journal of the International AIDS society: https://hivglasgow.org/wp-content/uploads/2023/01/P125_Lv_Final-Poster.pdf.
References
1. Lohse N, Obel N. Update of Survival for Persons With HIV Infection in Denmark. Ann Intern Med. 2016;165(10):749–750. doi:10.7326/L16-0091
2. Li Q, Chen M, Zhao H, et al. Persistent Low-Level Viremia is an Independent Risk Factor for Virologic Failure: a Retrospective Cohort Study in China. Infect Drug Resist. 2021;14:4529–4537. doi:10.2147/IDR.S332924.eCollection2021
3. Cohen C. Low-level viremia in HIV-1 infection: consequences and implications for switching to a new regimen. HIV Clin Trials. 2009;10(2):116–124. doi:10.1310/hct1002-116
4. Mackie NE, Phillips AN, Kaye S, et al. Antiretroviral drug resistance in HIV-1-infected patients with low-level viremia. J Infect Dis. 2010;201(9):1303–1307. doi:10.1086/651618
5. Delaugerre C, Gallien S, Flandre P, et al. Impact of low-level-viremia on HIV-1 drug-resistance evolution among antiretroviral treated-patients. PLoS One. 2012;7(5):e36673. doi:10.1371/journal.pone.0036673
6. DHHS Panel on Antiretroviral Guidelines for Adults and Adolescents. Guidelines for the use of antiretroviral agents in adults and adolescents with HIV; 2021. Available from: https://clinicalinfo.hiv.gov/sites/default/files/guidelines/documents/AdultandAdolescentGL.pdf.
7. WHO. Update of recommendations on first-and second-line antiretroviral regimens: HIV treatment; 2021. Available from: https://apps.who.int/iris/bitstream/handle/10665/325892/WHOCDS-HIV-19.15-eng.pdf.
8. Delaugerre C, Braun J, Charreau I, et al. Comparison of resistance mutation patterns in historical plasma HIV RNA genotypes with those in current proviral HIV DNA genotypes among extensively treated patients with suppressed replication. HIV Med. 2012;13(9):517–525. doi:10.1111/j.1468-1293.2012.01002.x
9. Wirden M, Soulie C, Valantin MA, et al. Historical HIV-RNA resistance test results are more informative than proviral DNA genotyping in cases of suppressed or residual viraemia. J Antimicrob Chemother. 2011;66(4):709–712. doi:10.1093/jac/dkq544
10. Derache A, Shin HS, Balamane M, et al. HIV drug resistance mutations in proviral DNA from a community treatment program. PLoS One. 2015;10(1):e0117430. doi:10.1371/journal.pone.0117430
11. Lambert-Niclot S, Allavena C, Grude M, et al. Usefulness of an HIV DNA resistance genotypic test in patients who are candidates for a switch to the rilpivirine/emtricitabine/tenofovir disoproxil fumarate combination. J Antimicrob Chemother. 2016;71(8):2248–2251. doi:10.1093/jac/dkw146
12. Joya C, Won SH, Schofield C, et al. Persistent low-level viremia while on antiretroviral therapy is an independent risk factor for virologic failure. Clin Infect Dis. 2019;69(12):2145–2152. doi:10.1093/cid/ciz129
13. Hosmane NN, Kwon KJ, Bruner KM, et al. Proliferation of latently infected CD4(+) T cells carrying replication-competent HIV-1: potential role in latent reservoir dynamics. J Exp Med. 2017;214(4):959–972. doi:10.1084/jem.20170193
14. Simonetti FR, Sobolewski MD, Fyne E, et al. Clonally expanded CD4+ T cells can produce infectious HIV-1 in vivo. Proc Natl Acad Sci U S A. 2016;113(7):1883–1888. doi:10.1073/pnas.1522675113
15. Lorenzo-Redondo R, Fryer HR, Bedford T, et al. Persistent HIV-1 replication maintains the tissue reservoir during therapy. Nature. 2016;530(7588):51–56. doi:10.1038/nature16933
16. Fletcher CV, Staskus K, Wietgrefe SW, et al. Persistent HIV-1 replication is associated with lower antiretroviral drug concentrations in lymphatic tissues. Proc Natl Acad Sci U S A. 2014;111(6):2307–2312. doi:10.1073/pnas.1318249111
17. Clutter DS, Jordan MR, Bertagnolio S, et al. HIV-1 drug resistance and resistance testing. Infect Genet Evol. 2016;46:292–307. doi:10.1016/j.meegid.2016.08.031
18. Fleming J, Mathews WC, Rutstein RM, et al. Low-level viremia and virologic failure in persons with HIV infection treated with antiretroviral therapy. Aids. 2019;33(13):2005–2012. doi:10.1097/QAD.0000000000002306
19. Elvstam O, Marrone G, Medstrand P, et al. All-Cause Mortality and Serious Non-AIDS Events in Adults With Low-level Human Immunodeficiency Virus Viremia During Combination Antiretroviral Therapy: results From a Swedish Nationwide Observational Study. Clin Infect Dis. 2021;72(12):2079–2086. doi:10.1093/cid/ciaa413
20. Saag MS, Gandhi RT, Hoy JF, et al. Antiretroviral Drugs for Treatment and Prevention of HIV Infection in Adults: 2020 Recommendations of the International Antiviral Society-USA Panel. JAMA. 2020;324(16):1651–1669. doi:10.1001/jama.2020.17025
21. Brodin J, Zanini F, Thebo L, et al. Establishment and stability of the latent HIV-1 DNA reservoir. Elife. 2016;5. doi:10.7554/eLife.18889
22. Arias A, López P, Sánchez R, et al. Sanger and Next Generation Sequencing Approaches to Evaluate HIV-1 Virus in Blood Compartments. Int J Environ Res Public Health. 2018;15(8). doi:10.3390/ijerph15081697
23. Huruy K, Mulu A, Liebert UG, et al. HIV-1C proviral DNA for detection of drug resistance mutations. PLoS One. 2018;13(10):e0205119. doi:10.1371/journal.pone.0205119
24. Zaccarelli M, Santoro MM, Armenia D, et al. Genotypic resistance test in proviral DNA can identify resistance mutations never detected in historical genotypic test in patients with low level or undetectable HIV-RNA. J Clin Virol. 2016;82:94–100. doi:10.1016/j.jcv.2016.07.007
25. Metzner KJ, Giulieri SG, Knoepfel SA, et al. Minority quasispecies of drug-resistant HIV-1 that lead to early therapy failure in treatment-naive and -adherent patients. Clin Infect Dis. 2009;48(2):239–247. doi:10.1086/595703
26. Palmer S, Kearney M, Maldarelli F, et al. Multiple, linked human immunodeficiency virus type 1 drug resistance mutations in treatment-experienced patients are missed by standard genotype analysis. J Clin Microbiol. 2005;43(1):406–413. doi:10.1128/JCM.43.1.406-413.2005
27. Wang C, Mitsuya Y, Gharizadeh B, et al. Characterization of mutation spectra with ultra-deep pyrosequencing: application to HIV-1 drug resistance. Genome Res. 2007;17(8):1195–1201. doi:10.1101/gr.6468307
28. Kieffer TL, Kwon P, Nettles RE, et al. G-->A hypermutation in protease and reverse transcriptase regions of human immunodeficiency virus type 1 residing in resting CD4+ T cells in vivo. J Virol. 2005;79(3):1975–1980. doi:10.1128/JVI.79.3.1975-1980.2005
29. Boukli N, Boyd A, Collot M, et al. Utility of HIV-1 DNA genotype in determining antiretroviral resistance in patients with low or undetectable HIV RNA viral loads. J Antimicrob Chemother. 2018;73(11):3129–3136. doi:10.1093/jac/dky316
30. Charpentier C, Montes B, Perrier M, et al. HIV-1 DNA ultra-deep sequencing analysis at initiation of the dual therapy dolutegravir + lamivudine in the maintenance DOLULAM pilot study. J Antimicrob Chemother. 2017;72(10):2831–2836. doi:10.1093/jac/dkx233
31. Fourati S, Malet I, Lambert S, et al. E138K and M184I mutations in HIV-1 reverse transcriptase coemerge as a result of APOBEC3 editing in the absence of drug exposure. Aids. 2012;26(13):1619–1624. doi:10.1097/QAD.0b013e3283560703
32. Allavena C, Rodallec A, Leplat A, et al. Interest of proviral HIV-1 DNA genotypic resistance testing in virologically suppressed patients candidate for maintenance therapy. J Virol Methods. 2018;251:106–110. doi:10.1016/j.jviromet.2017.10.016
33. Poveda E, Briz V, Paraskevis D, et al. Dynamics of drug-resistant HIV-1 in plasma and peripheral blood cells in patients during and after enfuvirtide therapy. AIDS Res Hum Retroviruses. 2007;23(9):1078–1082. doi:10.1089/aid.2006.0287
34. Turriziani O, Bucci M, Stano A, et al. Genotypic resistance of archived and circulating viral strains in the blood of treated HIV-infected individuals. J Acquir Immune Defic Syndr. 2007;44(5):518–524. doi:10.1097/QAI.0b013e3180315515
35. Kabamba-Mukadi B, Duquenne A, Henrivaux P, et al. HIV-1 proviral resistance mutations: usefulness in clinical practice. HIV Med. 2010;11(8):483–492. doi:10.1111/j.1468-1293.2009.00814.x
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.