")
Back to Journals » OncoTargets and Therapy » Volume 13
Role of αVβ3 in Prostate Cancer: Metastasis Initiator and Important Therapeutic Target
Authors Tang L, Xu M, Zhang L, Qu L, Liu X
Received 22 April 2020
Accepted for publication 19 June 2020
Published 28 July 2020 Volume 2020:13 Pages 7411—7422
DOI https://doi.org/10.2147/OTT.S258252
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Leo Jen-Liang Su
Lin Tang,1 Meng Xu,2,3 Long Zhang,4 Lin Qu,5 Xiaoyan Liu6
1College of Mathematics and Computer Science, Chifeng University, Chifeng, The Inner Mongol Autonomous Region 024005, People’s Republic of China; 2Department of Urology, The First Affiliated Hospital of Jinzhou Medical University, Jinzhou, Liaoning 121000, People’s Republic of China; 3R&D Department, Seekgene Technology Co., Ltd, Beijing 100000, People’s Republic of China; 4Department of Hepatobiliary Surgery, Yidu Central Hospital, Weifang, Shandong 262500, People’s Republic of China; 5Department of Orthopaedic Surgery, Anshan Hospital of the First Hospital of China Medical University, Anshan, Liaoning 114000, People’s Republic of China; 6Department of Pathology, The Fifth Medical Center of Chinese PLA General Hospital, Beijing 100000, People’s Republic of China
Correspondence: Meng Xu
Jinzhou Medical University, Songpo Road, Jinzhou, Liaoning 121000, People’s Republic of China
Email [email protected]
Abstract: In prostate cancer, distant organ metastasis is the leading cause of patient death. Although the mechanism of malignant tumor metastasis is unclear, studies have confirmed that integrin αVβ 3 plays an important role in this process. In prostate cancer, αVβ 3 mediates adhesion, invasion, immune escape and neovascularization through interactions with different ligands. Among these ligands and in addition to proteins that are directly related to tumor invasion, other proteins that contain the RGD structure could also bind to αVβ 3 and cause a number of biological effects. In this article, we summarized the ligand and downstream proteins related to αVβ 3-mediated prostate cancer metastasis as well as some diagnostic and therapeutic measures targeting αVβ 3.
Keywords: prostate cancer, metastasis, αVβ 3, signal pathway
Introduction
According to Cancer Statistics, 2020, the incidence of prostate cancer (PCa) is as high as 21%, ranking first among tumor diseases and second among expected causes of cancer-related death in American male patients.1 In the early stage of PCa, androgen is the main factor that regulates tumor growth.2,3 In the clinic, androgen deprivation therapy is used to treat PCa and has achieved ideal therapeutic effects.2 However, after a period of treatment, tumor cells are transformed from androgen-dependent into androgen-independent cells and become insensitive to chemotherapy.4 Recrudescent tumor growth is often faster and associated with an increased metastasizing tendency.5,6 Bone is the most common metastasizing site of PCa.7 Patients often suffer from bone pain and pathological fractures, which not only seriously affect the quality of life but also directly lead to patient death.8 Therefore, studying the mechanism of PCa metastasis is important to prevent PCa bone metastasis and prolong patient survival.
Metastasis is a process that involves interactions between the cell and extracellular matrix (ECM) as well as cell and target organ.9–11 At the beginning of metastasis, tumor cells must destroy the ECM as this structure prevents cancer cells from leaving their primary site. Cells must survive in the vasculature and finally be deposited in the corresponding tissue.12,13 Currently, a large number of in vivo and in vitro studies have found that the integrin protein family is involved in the entire process of tumor cell invasion and metastasis.5,7,10,14 In PCa, αVβ3 is the most studied integrin during metastasis. Integrin αVβ3 regulates tumor cell adhesion and migration by combining with extracellular matrix ligands and cell-surface ligands.10,15 Dissociative ligand, which contains the RGD structure (Arg-Gly-Asp), is recognized by integrin and triggers signaling to modulate the biological behavior of cells. The structure of αVβ3 contains transmembrane αβ heterodimers, and analyses of the crystal structures of integrin αVβ3 revealed a ligand-binding site on the headpiece of the αVβ3 region. This region is composed of the α and β subunits, ie, the β-propeller domain and the plexin-semaphorin-integrin (PSI) domain of the αV subunit and the βI-like domain and the hybrid domain of the β subunit. In particular, the ligand that contains an RGD sequence is directly recognized by amino acid residues at the interface between the β-propeller domain and the βI-like domain.15
It has been reported that αVβ3 is necessary for tumor cell adhesion to the ECM by targeting the RGD in fibronectin. Once cells adhere on to the ECM, the signal is transmitted intercellularly through the β-subunit. Inside the cell membrane, focal adhesion kinase (FAK), Paxillin and Zyxin gather at the tail of the β-subunit to form a trimer, which connects with the cytoskeleton (such as actin) to promote cytoskeletal protein rearrangement (to eventually yield protein fiber contraction). This series of changes eventually led to cell shape changes via transmission to the next αVβ3 molecule.16,17
Integrin αVβ3 not only directly interacts with ECM but also regulates MMP (especially MMP-2 and MMP-9) expression through the PI3K signaling pathway to hydrolyze collagen in the ECM.18–20 When tumor cells permeate into microvascular or lymphatic vessels, αVβ3 recruits some proteins with affinity to protect cells from complement or immune cell attack.21,22 When tumor cells successfully reach the target organ, αVβ3 acts as a receptor for VEGF to promote neovascularization, providing blood supply to the tumor cell mass and aiding in the establishment of metastases.23
These above functions of αVβ3 are not ascribed to its own activities but rather a series of extracellular ligands and intracellular signaling pathways that participate in the overall effect. Therefore, in this article, we have summarized the ligands and the downstream proteins related to αVβ3 regulating PCa metastasis to help researchers better understand the mechanism by which αVβ3 regulates tumor metastasis.
Extracellular or Membrane Ligands Combine with/Activate αVβ3
Although αVβ3 plays a dominate role in the progress of malignant metastasis, some extracellular or membranous ligands are indispensable for actuate invasion through the regulation of αVβ3 activation. As previously mentioned, the RGD sequence is specifically recognized by the αVβ3 binding site; thus, the majority of the ligand contains an RDG structure. SIBLING family proteins, such as bone sialoprotein (BSP) and osteopontin (OPN) are the classical ligand of αVβ3 (Figure 1).24 These proteins bind to αVβ3 with the help of RGD to hydrolyze ECM. However, the RGD sequence is not an absolute factor for ligand recognition by αVβ3. Several recent studies have demonstrated that cell chemokines and membrane ion channels, such as CXCL16 and BKCa, respectively, are also involved in αVβ3-mediated metastasis. We will summarize the activation of αVβ3 by different ligands below.
The SIBLING Family Regulates PCa Metastasis in an αVβ3-Dependent Manner
Small integrin-binding ligand N-linked glycoproteins (SIBLINGS) are soluble glycophosphoproteins, and members of the SIBLING family include osteopontin (OPN), bone sialoprotein (BSP), dentin matrix protein 1 (DMP1), dentin sialo-phosphoprotein (DSPP) and matrix extracellular phosphoprotein (MEPE). BSP and OPN have been more extensively studied than other SIBLING proteins.24–29 These two proteins have been identified as primary inducers in the progression of PCa metastasis, especially bone metastasis. As mentioned previously, BSP and OPN contain an integrin-binding RGD sequence, which can bind to αVβ3 on the cell surface. After these proteins are immobilized on the tumor cell membrane with the help of αVβ3, BSP or OPN recruit matrix metalloproteinases (for example: MMP-2 and MMP-9) to hydrolyze ECM and promote tumor cell dissemination and seeding in other organs.15,24,30,31 When tumor cells invaded into the vasculature, αVβ3 bound to BSP binds to complement factor H to protect tumor cells from complement attack. After prostate cancer cells pass the blood vessel endothelium and reach the target organ, particularly bone, blood supply becomes the most important factor for tumor survival. Aditi Gupta et al demonstrated that OPN and VEGF act as important regulators of angiogenesis via the integrin αvβ3 signaling pathway involved in cancer cell growth.32–34 These researchers found that OPN induces angiogenesis by enhancing the expression of VEGF in PC3 cells. In this process, integrin is the main intermediary because the mutated RGD sequence of OPN leads to decreased VEGF expression. After treating PC3 with an αVβ3 antagonist, VEGF cannot be restored to its original level even if OPN is overexpressed in PC3.
Connective Tissue Growth Factor Utilizes αVβ3 as a Receptor to Mediate PCa Bone Metastasis
Connective tissue growth factor (CTGF), which is also referred to as CCN2, is a small secreted matrix protein that regulates various cellular functions, including proliferation, adhesion, migration, and invasion.35–38 Under physiological conditions, CCN2 induces skeletogenesis, angiogenesis, and injury repair.39 However, pathologically, CCN2 is closely related to cancer bone metastasis.40,41 It has been reported that αVβ3 act as a major receptor for CCN2 and mediates the migration and adhesion of endothelial and hepatic stellate cells (Figure 1). Based on this theory, Bongjun Kim et al assessed the relationship between CCN2 and αVβ3 in PCa. They found that CCN2 increases matrix invasion activity of the bone metastatic cell line PC3 by upregulation of RUNX2 levels by modulating the promoter activity of RUNX2, which induces the expression of MM-2 and MM-9 proteins. These effects were abolished when PC3 cells were treated with an anti-αvβ3 antibody. To promote growth outside of the primary organ, cancer cells must transform the tissue microenvironment into a suitable site for survival.42 For PCa bone metastasis, tissue reconstruction is inevitable after cancer cells arrive at bone. The osteoclast differentiation factor RANKL is an important regulator involved in bone metabolism. RANKL induces osteoblasts to differentiate into osteoclasts and initiates osteolysis.43 Hydroxyapatite (HA) is exposed on the surface of the dissolved bone, and this structure promotes tumor cell immobilization on the bone surface via αVβ3.44 Bongjun Kim et al confirmed that acetylated RUNX2 induces RANKL expression, and protein acetylation was attributed to the fact that integrin αVβ3 mediates CCN2-stimulated acetylation and stabilization of RUNX2 in highly bone-metastatic cancer cells (Figure 1).40
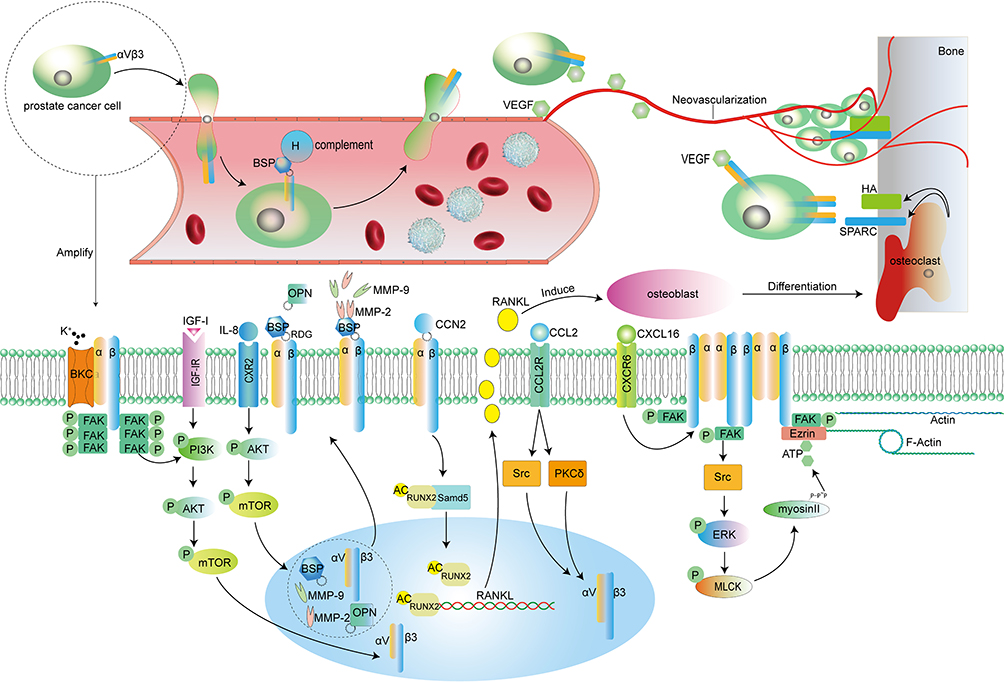 |
Figure 1 The figure describes the process by which αVβ3 mediates prostate cancer cell metastasis. Macroscopically, after cancer cells destroy the ECM and leave their primary site, cancer cells depend on αVβ3, which destroys vascular endothelial cells, to enter blood vessels. In blood vessels, tumor cells under the protection of BSP escape the attack of the immune system and then pass through the blood vessels to reach the metastatic site. Here, bone tissue is used as an example. Under the induction of VEGF, cancer cells recognize SPARC and HA and interact with them on the surface of bone with the help of αVβ3; at the same time, VEGF induces neovascularization to supply blood to the cell mass system. At the microscopic level, IL-8, IGF-I, CCN2 and other ligands activate different signaling pathways to promote the expression of αVβ3 in cancer cells. In addition, CCN2 transmits signals into the cell through αVβ3 to increase the acetylation of the transcription factor RUNX2, and RUNX2 promotes RANKL expression in tumor cells; these tumor cells then secrete RANKL to induce osteoblast differentiation into osteoclasts, and osteoclast-mediated bone tissue remodeling can expose HA and SPARC on the bone surface. BKCa and CXCL16 promote the aggregation and activation of αVβ3 and trigger the activation of downstream FAK. In addition to enhancing the activity of the ERK or PI3K pathway, activated FAK triggers cytoskeletal protein remodeling with the help of adaptor proteins, further enhancing tumor cell invasion ability. Cell information: The PC-3 cell line was initiated from the bone metastasis of a grade IV prostatic adenocarcinoma from a 62-year-old Caucasian male; DU145 was initiated from a 69-year-old Caucasian male with brain metastasis of prostatic adenocarcinoma; LNCAP was isolated from the left supraclavicular lymph node of a 50-year-old prostate cancer patient. LNCAP is sensitive to hormones; C4-2B is a subtype derived from the LNCAP cell line. Abbreviations: BSP, bone sialoprotein; VEGF, vascular endothelial growth factor; HA, hydroxyapatite; SPARC, osteonectin; FAK, focal adhesion kinase; IGF-I, insulin-like growth factor; IL-8, interleukin8; OPN, osteopontin; MMP (2 or 9), matrix metalloproteinase (2 or 9); CCN2, connective tissue growth factor; CCL2, chemokines2; CXCL16, chemokines16; P, phosphorylated; AC, acetylated. |
Cellular Chemokines Can Enhance PCa Invasion by Regulating αVβ3 Expression
Numerous cellular chemokines are related to malignant cell proliferation, survival, spread, and angiogenetic response.45–50 In resistant-prostate cancer, recrudescent cancer cell survival depends on autocrine chemokines that stimulate angiogenetic and drug resistance processes.51–54 Our previous study demonstrates that interleukin-8 (IL-8 or CXCL8) influences αVβ3 level in PC3 cells, and SB225002 (IL-8 receptor antagonist) potently attenuates cancer cell invasion and αVβ3 expression levels. These effects are regulated by the AKT/mTOR signaling pathway (Figure 1). In the SB225002-treated group, phosphorylated AKT is inhibited, and the expression levels of downstream proteins mTOR and P‑mTOR were significantly reduced.
Chemokine 2 (CCL2) is another cytokine that is associated with tumor progression.55 Functional CCL2 is a chemoattractant for monocytes, memory T-cells, and natural killer cells in adaptive immunity responses;56–59 however, CCL2 has also been reported to be implicated in the regulation of cancer cell growth, angiogenesis, and metastasis in prostate and breast cancers.60,61 Tien-Huang Lin et al used shRNA to silence CCL2 expression in PC3 cells and found that inhibition of CCL2 expression inhibited the migratory ability of PC3 cells by approximately 80%. Regarding the mechanism by which CCL2 regulates the PCa invasion, they verified the interrelation between CCL2 and αVβ3. Pretreatment of cells with an anti-αvβ3 integrin monoclonal antibody markedly inhibited CCL2-induced PCa cell migration, and knockdown of CCL2 also reduced αvβ3 integrin expression in PC3 cells. In addition, they also demonstrated how CCL2 affects αvβ3 expression using the PKCδ inhibitor rottlerin and c-Src inhibitor PP2. They confirmed that CCL2 enhances cell migration and αvβ3 expression through CCR2 receptors/PKCδ and/or the c-Src pathway in human PCa cells (Figure 1).62
In addition to the fact that αvβ3 directly enhances PCa invasion via chemokine-mediated processes, membrane dynamics are also involved in the migration of malignant tumors induced by chemokines.63–65 Integrin αvβ3 directly interacts with the ECM. In addition, actin rearrangement also plays a key role in regulating cellular interactions with the ECM, such as motility and vesicular trafficking. No study has confirmed that αvβ3 directly contacts actin, but some studies have noted that ezrin serves as a connector to communicate αvβ3 signals to actin. Ezrin is a scaffold protein that promotes protein interactions with the actin cytoskeleton to regulate cell migration.66,67 Especially in PCa, ezrin can connect the tail of the β-subunit in αvβ3 to actin.68–70 In experiments assessing how CXCL16 affects PCa invasion, researchers found that treatment of PC3 cells with CXCL16 cause αvβ3 molecules to cluster together on the cell surface, and p-Ezrin and F-Actin levels were increased significantly. The phosphorylation of these proteins is most likely transmitted by αvβ3, especially β-subunits because FAK is recruited to sites of integrin clustering following integrin engagement and is activated through undefined mechanisms. Activated FAK stimulates its downstream target ERK, and the ERK signaling pathway is a common pathway for Ezrin phosphorylation.71 All these findings suggest CXCL16 not only by directly enhances αvβ3 expression but also transmit signals into cells via β3-subunits to trigger cytoskeletal protein rearrangement to promote enhanced tumor cell migration (Figure 1).17
VEGF Promotes PCa Bone Metastasis by Controlling αvβ3 and SPARC Interactions
SPARC, which is also called osteonectin, is a component of the bone matrix that modulates cellular interactions with the extracellular matrix, exerting an adhesive, promigratory effect on cells.72–74 Studies confirmed that high expression of SPARC in most of the tumors with a tendency for bone metastasis.75,76 In PCa, Sarmishtha De et al treated tumor cells with different antagonists and conclude that tumor cells on the bone surface linked with SPARC exhibit αvβ3 expression. In addition, αvβ blockade resulted in inhibition of LNCaP cell migration to SPARC. Meanwhile, VEGF produced by tumor cells through autocrine mechanisms enhances the binding of αvβ3 and SPARC. Antibody neutralization of VEGF inhibits αvβ3 activation and decreased SPARC conjugation. Thus, VEGF secreted by tumor cells not only enhances neovascularization but also participates in αvβ3 activation and conjunction with SPARC (Figure 1).77
PI3K Signaling Pathway Activation is Essential for IGF-I Regulation of αVβ3 Expression
Insulin-like growth factor-I (IGF-I) is implicated in the evolution of various neoplasias.78–82 In PCa, considerable experimental and epidemiological evidence showed that cancer cells produce IGF-I via autocrine mechanisms and express IGF-I receptors (IGF-IR). IGF-I enhances PCa proliferation by increasing caryomitosis, and these effects could maintain LNCAP growth in medium without serum.82 Meanwhile, in the process of tumor conversion, IGF-I transforms androgen-dependent PCa cells to an androgen-independent phase.83–85 In terms of migration, researchers found that IGF-I stimulates PCa invasion by combining with IGF-IR. Blocking αvβ3 with monoclonal antibodies mitigates the effects of IGF-I. In cell proliferation experiments, IGF-I enhances AKT and ERK phosphorylation to promote the proliferation of DU145 PCa cells. On the basis of these findings, researchers used chemical antagonists LY294002 (PI3K antagonist) and UO126 (ERK antagonist) to pretreat DU145 cells. Then, IGF-1 was added to the medium. They found that LY294002 completely counteracted the effect of IGF-1 in DU145 cells, but UO126 did not affect IGF-I induced migration of DU145 cells. They concluded that the mechanism by which IGF-I enhances PCa invasion might depend on the upregulation of αvβ3 expression levels through the PI3K signaling pathway (Figure 1).86
Ion Channel Proteins Take Part in PCa Migration Mediated by αVβ3
BKCa is a large-conductance calcium-activated potassium channel that is mainly expressed in excitable cells. It modulates vasomotor and nerve excitability by regulating membrane potential and calcium signaling.87–89 However, experiments performed by Cheng Du et al demonstrated that both BKCa mRNA and protein levels in PCa cells were increased compared with normal cells. BKCa overexpression strengthens the invasive ability of cancer cell. To investigate mechanisms by which BKCa regulates invasion, immunofluorescence experiments were performed, and BKCa-αvβ3 complex formation was observed. In addition, αvβ3 and BKCa were reciprocally immunoprecipitated by each other. As the important downstream protein of αvβ3, FAK phosphorylation levels were increased after BKCa overexpression. To verify the direct relationship between BKCa and αvβ3/FAK, a BKCa inhibitor (IBTX) and activator (NS1619) were used to block or enhance K+ flow. BKCa inhibition and activation result in enhanced or decreased PCa cell invasion, respectively. These agents did not affect the expression of αvβ3 and FAK. Moreover, FAK phosphorylation is only related to β-subunit regulation; thus, these researchers suggest that BKCa facilitates the coupling between integrin αvβ3 and FAK or recruitment of FAK to the transmembrane BKCa/αvβ3 integrin complex (Figure 1).90
In addition to the above factors that participate in the progression of PCa, many additional proteins or compounds enhance the invasion ability of PCa through the integrin pathway, for example, thyroid hormones (T4) and Gastrin-releasing peptide receptor (GRPR). These proteins regulate invasion generally via the same mechanism, namely enhancing αvβ3 expression and promoting αvβ3 aggregation on the cell surface or increasing αvβ3 activity. Thus, all of these proteins and compounds are not summarized here.
Intercellular Signaling Pathway Stimulated by αVβ3 Related to PCa Metastasis
Ligand combines with its receptor to trigger a series of cascade reactions in the cytoplasm to regulate cell biological behavior.91,92 This theory is adapted for αvβ3, which influences PCa metastasis. Ligand enhances or inhibits PCa behaviors through one or more downstream pathways transmitted by αvβ3. In this section, we will summarize several well-known downstream proteins of αvβ3.
FAK is the First Signaling Molecule of αVβ3 in the Cytoplasm
Focal Adhesion Kinase (FAK) belongs to a kind of tyrosine kinase.93 This protein does not contain a transmembrane structure but harbors two autophosphorylation sites at tyrosine residues 397 and 925.94 Integrin αvβ3 molecules on the cell surface assemble to activate FAK and transmit information to FAK by the β-subunit of its intercellular tail.15,94 After binding to integrin, most ligands transmit the signal to FAK through the β-subunit of integrin. Then, the signals of the FAK signaling pathway are transmitted to the cytoplasm to trigger the corresponding biological effect. The following uses androgens and EMT serves as examples to illustrate how FAK participates in integrin-mediated PCa metastasis.
Androgen is an important factor that regulates PCa growth and mediates metastasis through the integrin-FAK pathway. Gabriella Castoria and coworkers used murine fibroblasts to investigate how androgen-induced PCa migration. They found that NIH3T3 cells (mouse embryo NIH3T3 fibroblasts) treated with RI88I (synthetic androgen) exhibit rapid FAK Tyr397 phosphorylation. Tyr118-phosphorylated paxillin (the focal contact-associated protein) parallels tyrosine phosphorylation of FAK upon androgen stimulation of cells. Meanwhile, cytoskeletal protein remodeling can be detected within a few minutes after androgen treatment. Subsequently, the researchers performed immunoprecipitation and found that the androgen receptor (AR) in androgen-treated NIH3T3 cells rapidly interacted with filamin A (FlnA) at the cytoskeleton. AR-FlnA assembled with integrin β1 to form a tripartite complex, and knockdown of either integrin β1 or FlnA abolished Tyr 397 phosphorylation of FAK induced by androgen. Thus, these researchers conclude that the interaction between AR and FlnA is a key event for FAK activation, and paxillin tyrosine phosphorylation in androgen-stimulated cells further reinforces the role of the AR/FlnA interaction in androgen-mediated motility.95–97
In most cancers, the epithelial-mesenchymal transition (EMT) is a key event in metastasis. At the molecular level, the EMT is characterized by a decrease in the expression of epithelial markers, mainly E-cadherin, which is located on the cell surface of epithelial tissues, and an increase in the expression of mesenchymal markers, such as vimentin, a cytoskeleton protein associated with EMT initiation. It is worth noting that FAK expressed by tumor cells also participates in the EMT process. In A549 lung cancer cells and colorectal cancer cells, FAK can be activated by EGFR, and phosphorylation of FAK at Tyr397 can trigger signal transduction mechanisms, activate downstream signaling molecules, downregulate E-cadherin expression and upregulate vimentin expression. In PCa, EMT does not occur in some PCa cells but occurs in other, such as PC3 and DU145 cells. In in vitro experiments, DU145 and PC3 cells (bone and brain metastasis of prostate cancer cells, respectively) exhibited increased invasiveness and high FAK, FlnA and paxillin expression compared with C4-2B and LNCaP cells, which express lower FAK, do not exhibit the EMT, and move more slowly than DU145 and PC3 cells in migration experiments. Drugs can be used to induce the EMT, for example nerve growth factor (NGF). NGF robustly induces the EMT in DU145 and PC3 cells more effectively compared with C4-2B cells. These phenomena demonstrate that differences in FAK expression in cells with different invasive capabilities highlight the role of FAK in the EMT process of PCa.96,98
FAK is closely related to PCa metastasis. High FAK expression in PCa often indicates the presence of the EMT phenotype or the tendency of cells to metastasize. As mentioned previously, ligand does not have the ability to directly activate FAK, but FAK activity is influenced by integrin.90 Recent studies have indicated that αvβ3 blockade by an antagonist, neutralizing antibody and siRNA inhibit FAK phosphorylation.90,99 In addition, as the direct substrate of FAK, the P85 subunit of PI3K can be phosphorylation by FAK.90,100 Thus, if external stimuli are transmitted to the intracellular pathway through αvβ3, FAK phosphorylation is an important intermediate hinge in this process (Figure 1).
The αVβ3/RUNX2/RANKL Pathway is Closely Related to Osteolytic Metastasis
We mentioned earlier that IGF promotes PCa invasion via the αvβ3/RUNX2/RANKL pathway. Thus, RUNX2/RANKL is another downstream protein of αvβ3. Pathologically, PCa bone metastasis is classified as an osteolytic disease.101 Runx2 is a transcription factor that plays a key regulatory role in osteoblast differentiation.102,103 DNA sequencing found that the receptor activator of nuclear factor kB ligand (RANKL) contains a Runx2 binding site in the promoter region.40,104 Thus, αvβ3-induced bone matrix dissolution likely depends on the delivery of Runx2 to the nucleus to promote RANKL transcription. In terms of mechanism, Aditi Gupta et al did not provide evidence that αvβ3 could directly interact with Runx2, but they determined that αvβ3 activates Samd5. Activated Samd5 subsequently transports Runx2 to the nucleus. In the nucleus, Runx2 guides the transcription of RANKL.34 RANKL is subsequently secreted by tumor and induces multinucleated giant cells to differentiate into osteoclasts and remodel bone matrix.
The Classical Signal Pathway Receives Information Transmitted by αVβ3
In addition to specific proteins that act downstream of αvβ3, numerous well-known signal pathway participate in signal transport initiated by αvβ3. The ERK pathway is one example. Two primary mechanisms are involved in αvβ3-mediated activation of ERK. In one mechanism, FAK recruits a series of adaptor proteins, such as growth-factor-receptor bound protein2 and CRK-associated substrate, and activates protein kinases, such as SRC-family kinases, to activate ERK.99 In another mechanism, αvβ3 colocalizes with growth factor receptors on the surface of cells, and growth factors activate the ERK pathway.105 The alternating process of contact and separation between the cell membrane and the ECM maintains tumor cells movement on the ECM. ERK activation reduces cell adhesion to the ECM and guarantees that the cell membrane is prepared for the next contact. Activated ERK phosphorylates myosin-light-chain kinase (MLCK), and MLCK is a key regulator of contractile force within the cell. Phosphorylated MLCK activates the ATPase of myosin II. Myosin II provides energy for cells to move in the direction of actin contraction, and activated myosin generates contractile forces that pull the cell forward towards newly formed integrin contacts and breaks adhesive contacts at the trailing edge of the cell.106
PI3K is another transmitter of αvβ3 signals. At the beginning of invasion, ligand combines with αvβ3, and the biological signal arrives at PI3K through β-subunit/FAK. Then, the PI3K/AKT/mTOR pathway induces transcription of MMPs and SIBLINGs.55,107 These two families of proteins translocate to the cell surface and combine with αvβ3. MMPs to hydrolyze collagen in the ECM, and SIBLINGs aid in tumor cell adhesion and organization.
The factors mentioned above are reported in the literature as mechanisms by which αvβ3 guides PCa metastasis. However, in other malignant diseases, some small molecules are also important for the intracellular transport of the αvβ3 signal, such as the adaptor protein CAS, SHC and small G proteins.108–110 Some of these proteins are directly used as downstream messengers of αvβ3. Some require the help of a middle messenger. Further research on the role of these factors in PCa will provide more comprehensive mechanistic information on PCa invasion and metastasis.
αVβ3-RGD Specific Recognition Acts as a Tool for PCa Diagnosis and Therapy
Based on the highly specific expression of αvβ3, researchers have exploited various methods based on αvβ3-specific recognition of RGD sequences in the diagnosis and imaging of PCa. Kongzhen Hu et al linked 4-nitrophenyl2-18F-fluoropropionate (18F-NFP) and β-glutamate-RGD with PEG to construct an αvβ3 radiotracer. PET imaging systems are used to detect 18F-FP-PEG2-β-Glu-RGD2 uptake to assess the efficiency of implanted tumors in mice. The results showed 18F-FP-PEG2-β-Glu-RGD2 was rapidly cleared from blood predominately by renal excretion. MicroPET-CT imaging with 18F-FP-PEG2-β-Glu-RGD2 revealed high tumor contrast and low background in PC3 prostate cancer-bearing mouse models. 18F-FP-PEG2-β-Glu-RGD2 exhibited good stability in vivo. These experiments use αvβ3 as the detection object and demonstrate that specific αvβ3-ligand binding can effectively detect the spread of cancer cells in the body.111 Ambros J. Beer and their team further compared the sharpness of [18F] Galacto-RGD and bone scintigraphy in the diagnosis of PCa bone metastases. The diagnosis rate of patients with bone metastasis using [18F] Galacto-RGD is 78% (58 of 74 patients were diagnosed with bone metastases), but the image clarity of [18F] Galacto-RGD is significantly lower than that of bone scintigraphy. Researchers used a method based on [18F] Galacto-RGD tracking αVβ3 imaging to demonstrate that image clarity is closely related to the expression level of αVβ3 in the tumor. Compared with the bone scintigraphy method, the use of [18F] Galacto-RGD to diagnose bone metastases in patients with PCa has the advantages of a good visualization and low background of surrounding tissues.112
In addition to galactose-labeled RGD, researchers also use 99mTc, bombesin, gastrin-releasing peptide receptor and RGD labeled with Afterglow materials to track PCa progression.113–115 According to their results, after chemical- or biomarker-labeled RGD combines with αvβ3, the signal can be visualized in vitro using equipment to clearly distinguish the lesion from the surrounding tissue. This conclusion indicates that clinicians can judge the progression of PCa by detecting the expression of αvβ3 and its location in patients and can predict whether the tumor is at risk of bone metastasis if necessary.
Inspired by RGD’s specific binding to αvβ3, researchers have also developed new PCa treatment options. Traditional chemotherapeutic drugs have a short half-life and are often present at insufficient concentrations when they reach the treatment site, which seriously affects the therapeutic effect of the drug. Docetaxel (an αvβ3 antagonist) is a widely used anticancer drug that is used alone or in combination with other drugs to treat tumors.116 However, this drug suffers from the limitations of nonspecific cytotoxicity in normal tissues. To improve the tissue specificity of this drug, Hitesh Kulhari M Pharm et al used nanomicelles to couple docetaxel and RGD sequences and successfully achieved RGD-αvβ3 targeted drug delivery.117 Yajie Gao et al successfully coupled paclitaxel to RDG to achieve a drug delivery model for mouse xenografts.118 Both of these targeted drug delivery methods show increased therapeutic efficiency and reduced cytotoxicity than traditional drug delivery methods, while chemically modified drugs exhibit increased stability in vivo. In addition to the direct coupling of RGD and drugs, researchers still use liposome-encapsulated drugs to couple RGD for targeted drug delivery experiments to reduce the degradation of chemotherapeutics. In experiments using liposomes to encapsulate cisplatin, liposomes showed significantly increased cellular uptake and increased cytotoxicity of loaded cisplatin. Clinical observation confirmed that the RGD-conjugated liposomal drug system could inhibit osteoclastic and osteoblastic bone lesions, relieve pain, and improve overall survival.119,120
Another novel treatment strategy is RGD-mediated oncolytic virus targeting PCa. Oncolytic adenovirus was able to selectively replicate and kill cancer cells while sparing normal cells.121,122 The anti-tumor efficiency of this virus depends on the level of Coxsackie virus adenovirus receptor (CAR) expression by tumor cells.123 Experimental results by Yue-Hong Shen et al revealed that the E1A/E1B double mutant oncolytic adenovirus AxdAdB-3 exhibited antitumor activity in an orthotopic PCa SCID (severe combined immunodeficiency) mouse model. However, AxdAdB-3 showed insufficient cytopathic effects in some PCa cell lines that expressed low levels of CAR. To overcome the low treatment efficiency caused by low expression of CAR, they insert the RGD peptide into AxdAdB-3 to form a new oncolytic virus: AxdAdB3-F/RGD. They used the novel oncolytic virus to infect PCa cells in vitro, and a PCa xenograft model was constructed using nude mice. AxdAdB-3 was more cytopathic in CAR-positive PCa cells compared with CAR-negative cells, whereas AxdAdB3-F/RGD caused potent oncolysis in both CAR-positive and CAR-negative cancer cells. In contrast, AxdAdB3-F/RGD was not cytopathic against normal prostate epithelial RWPE-1 cells. Intratumoral injection of AxdAdB3-F/RGD into CAR-negative prostate cancer cell xenografts in nude mice can significantly inhibit tumor growth.124
Conclusion
Prostate cancer recurrence and mortality are increasing rapidly. The identification of an effective anti-tumor strategy is crucial to improve patient prognosis and prolong patient survival. The relationship between integrin αvβ3 and prostate cancer is gaining increasing attention. As a transmembrane protein, αvβ3 is involved in almost all prostate cancer progression processes. Different ligands combined with αvβ3 mediate various biological behaviors of tumors, such as proliferation, invasion, migration, angiogenesis and immune escape. Based on this characteristic of αvβ3, multiple diagnostic and therapeutic strategies have been developed to combat prostate cancer. Here, we summarized the current role of αvβ3 in the progression of prostate cancer and new treatments developed for αvβ3 as a therapeutic target. Although these methods show high detection rates and therapeutic effects, the safety of the corresponding reagent or drug needs to be further confirmed. The identification of a safe, effective, and inexpensive antitumor drug is important for prostate cancer patients.
Disclosure
Meng Xu is an employee of Seekgene Technology Co., Ltd. The authors report no other possible conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020. CA Cancer J Clin. 2020;70(1):7–30. doi:10.3322/caac.21590
2. Giguere V. DNA-PK, Nuclear mTOR, and the Androgen Pathway in Prostate Cancer. Trends Cancer. 2020;6(4):337–347. doi:10.1016/j.trecan.2020.01.015
3. Zou Y, Tang F, Talbert JC, Ng CM. Using medical claims database to develop a population disease progression model for leuprorelin-treated subjects with hormone-sensitive prostate cancer. PLoS One. 2020;15(3):e230571. doi:10.1371/journal.pone.0230571
4. Liao Y, Liu Y, Xia X, et al. Targeting GRP78-dependent AR-V7 protein degradation overcomes castration-resistance in prostate cancer therapy. Theranostics. 2020;10(8):3366–3381. doi:10.7150/thno.41849
5. Juan-Rivera MC, Martinez-Ferrer M. Integrin Inhibitors in Prostate Cancer. Cancers. 2018;10(2):44. doi:10.3390/cancers10020044
6. Wang Y, Singhal U, Qiao Y, et al. Wnt Signaling Drives Prostate Cancer Bone Metastatic Tropism and Invasion. Transl Oncol. 2020;13(4):100747. doi:10.1016/j.tranon.2020.100747
7. Rizzoli R, Body JJ, Brandi ML, et al. Cancer-associated bone disease. Osteoporos Int. 2013;24(12):2929–2953. doi:10.1007/s00198-013-2530-3
8. Wang N, Xu L, Wang JS, et al. Traditional Chinese medicine on treating pain caused by prostate cancer: A systematic review and meta-analysis. Medicine. 2019;98(44):e17624. doi:10.1097/MD.0000000000017624
9. Selvaggio G, Canato S, Pawar A, et al. Hybrid epithelial-mesenchymal phenotypes are controlled by microenvironmental factors. Cancer Res. 2020;80(11):2407–2420. doi:10.1158/0008-5472.CAN-19-3147
10. Takada Y, Ye X, Simon S. The integrins. Genome Biol. 2007;8(5):215. doi:10.1186/gb-2007-8-5-215
11. Mas-Moruno C, Rechenmacher F, Kessler H. Cilengitide: the first anti-angiogenic small molecule drug candidate design, synthesis and clinical evaluation. Anticancer Agents Med Chem. 2010;10(10):753–768. doi:10.2174/187152010794728639
12. Mishra P, Kiebish MA, Cullen J, et al. Genomic alterations of Tenascin C in highly aggressive prostate cancer: a meta-analysis. Genes Cancer. 2019;10(5–6):150–159. doi:10.18632/genesandcancer.196
13. Quach ND, Kaur SP, Eggert MW, et al. Paradoxical Role of Glypican-1 in Prostate Cancer Cell and Tumor Growth. Sci Rep. 2019;9(1):11478. doi:10.1038/s41598-019-47874-2
14. Zhuang X, Lv M, Zhong Z, Zhang L, Jiang R, Chen J. Interplay between integrin-linked kinase and ribonuclease inhibitor affects growth and metastasis of bladder cancer through signaling ILK pathways. J Exp Clin Cancer Res. 2016;35(1):130. doi:10.1186/s13046-016-0408-x
15. Hood JD, Cheresh DA. Role of integrins in cell invasion and migration. Nat Rev Cancer. 2002;2(2):91–100. doi:10.1038/nrc727
16. Lu J, Doyle AD, Shinsato Y, et al. Basement membrane regulates fibronectin organization using sliding focal adhesions driven by a contractile winch. Dev Cell. 2020;52(5):631–646. doi:10.1016/j.devcel.2020.01.007
17. Singh R, Kapur N, Mir H, Singh N, Lillard JJ, Singh S. CXCR6-CXCL16 axis promotes prostate cancer by mediating cytoskeleton rearrangement via Ezrin activation and alphavbeta3 integrin clustering. Oncotarget. 2016;7(6):7343–7353. doi:10.18632/oncotarget.6944
18. Sun LC, Luo J, Mackey LV, Fuselier JA, Coy DH. A conjugate of camptothecin and a somatostatin analog against prostate cancer cell invasion via a possible signaling pathway involving PI3K/Akt, alphaVbeta3/alphaVbeta5 and MMP-2/-9. Cancer Lett. 2007;246(1–2):157–166. doi:10.1016/j.canlet.2006.02.016
19. Zhao ZS, Wang YY, Chu YQ, Ye ZY, Tao HQ. SPARC is associated with gastric cancer progression and poor survival of patients. Clin Cancer Res. 2010;16(1):260–268. doi:10.1158/1078-0432.CCR-09-1247
20. He Y, Liu XD, Chen ZY, et al. Interaction between cancer cells and stromal fibroblasts is required for activation of the uPAR-uPA-MMP-2 cascade in pancreatic cancer metastasis. Clin Cancer Res. 2007;13(11):3115–3124. doi:10.1158/1078-0432.CCR-06-2088
21. Brown NF, Marshall JF. Integrin-Mediated TGFbeta Activation Modulates the Tumour Microenvironment. Cancers. 2019;11(9):1221. doi:10.3390/cancers11091221
22. Avraamides CJ, Garmy-Susini B, Varner JA. Integrins in angiogenesis and lymphangiogenesis. Nat Rev Cancer. 2008;8(8):604–617. doi:10.1038/nrc2353
23. Dong Y, Xie X, Wang Z, et al. Increasing matrix stiffness upregulates vascular endothelial growth factor expression in hepatocellular carcinoma cells mediated by integrin beta1. Biochem Biophys Res Commun. 2014;444(3):427–432. doi:10.1016/j.bbrc.2014.01.079
24. Bellahcene A, Castronovo V, Ogbureke KU, Fisher LW, Fedarko NS. Small integrin-binding ligand N-linked glycoproteins (SIBLINGs): multifunctional proteins in cancer. Nat Rev Cancer. 2008;8(3):212–226. doi:10.1038/nrc2345
25. Liu B, Xu M, Guo Z, Liu J, Chu X, Jiang H. Interleukin-8 promotes prostate cancer bone metastasis through upregulation of bone sialoprotein. Oncol Lett. 2019;17(5):4607–4613. doi:10.3892/ol.2019.10138
26. Rustamov V, Keller F, Klicks J, Hafner M, Rudolf R. Bone Sialoprotein Shows Enhanced Expression in Early, High-Proliferation Stages of Three-Dimensional Spheroid Cell Cultures of Breast Cancer Cell Line MDA-MB-231. Front Oncol. 2019;9:36. doi:10.3389/fonc.2019.00036
27. Zhu BP, Guo ZQ, Lin L, Liu Q. Serum BSP, PSADT, and Spondin-2 levels in prostate cancer and the diagnostic significance of their ROC curves in bone metastasis. Eur Rev Med Pharmacol Sci. 2017;21(1):61–67.
28. Pang X, Gong K, Zhang X, Wu S, Cui Y, Qian BZ. Osteopontin as a multifaceted driver of bone metastasis and drug resistance. Pharmacol Res. 2019;144:235–244. doi:10.1016/j.phrs.2019.04.030
29. Wisniewski T, Zyromska A, Makarewicz R, Zekanowska E. Osteopontin and angiogenic factors as new biomarkers of prostate cancer. Urol J. 2019;16(2):134–140. doi:10.22037/uj.v0i0.4282
30. Balera BV, Chaves-Neto AH, Landim DBT, Penha OS. Soluble yerba mate (Ilex Paraguariensis) extract enhances in vitro osteoblastic differentiation of bone marrow-derived mesenchymal stromal cells. J Ethnopharmacol. 2019;244:112131. doi:10.1016/j.jep.2019.112131
31. Hassan B, Baroukh B, Llorens A, et al. NAMPT expression in osteoblasts controls osteoclast recruitment in alveolar bone remodeling. J Cell Physiol. 2018;233(9):7402–7414. doi:10.1002/jcp.26584
32. Gupta A, Zhou CQ, Chellaiah MA. Osteopontin and MMP9: associations with VEGF Expression/Secretion and Angiogenesis in PC3 Prostate Cancer Cells. Cancers. 2013;5(2):617–638. doi:10.3390/cancers5020617
33. Gupta A, Cao W, Sadashivaiah K, Chen W, Schneider A, Chellaiah MA. Promising noninvasive cellular phenotype in prostate cancer cells knockdown of matrix metalloproteinase 9. ScientificWorldJournal. 2013;2013:493689. doi:10.1155/2013/493689
34. Gupta A, Cao W, Chellaiah MA. Integrin alphavbeta3 and CD44 pathways in metastatic prostate cancer cells support osteoclastogenesis via a Runx2/Smad 5/receptor activator of NF-kappaB ligand signaling axis. Mol Cancer. 2012;11(1):66. doi:10.1186/1476-4598-11-66
35. Shimbo A, Kajiyama H, Tamauchi S, et al. Expression of connective tissue growth factor as a prognostic indicator and its possible involvement in the aggressive properties of epithelial ovarian carcinoma. Oncol Rep. 2019;42(6):2323–2332. doi:10.3892/or.2019.7352
36. Yao Y, Wang Y, Li L, et al. Down-regulation of interferon regulatory factor 2 binding protein 2 suppresses gastric cancer progression by negatively regulating connective tissue growth factor. J Cell Mol Med. 2019;23(12):8076–8089. doi:10.1111/jcmm.14677
37. Li X, Liu H, Sun L, et al. MicroRNA-302c modulates peritoneal dialysis-associated fibrosis by targeting connective tissue growth factor. J Cell Mol Med. 2019;23(4):2372–2383. doi:10.1111/jcmm.14029
38. Jun JI, Lau LF. Taking aim at the extracellular matrix: CCN proteins as emerging therapeutic targets. Nat Rev Drug Discov. 2011;10(12):945–963. doi:10.1038/nrd3599
39. Kanyama M, Kuboki T, Akiyama K, et al. Connective tissue growth factor expressed in rat alveolar bone regeneration sites after tooth extraction. Arch Oral Biol. 2003;48(10):723–730. doi:10.1016/S0003-9969(03)00153-5
40. Kim B, Kim H, Jung S, et al. A CTGF-RUNX2-RANKL axis in breast and prostate cancer cells promotes tumor progression in bone. J Bone Miner Res. 2020;35(1):155–166. doi:10.1002/jbmr.3869
41. Nakata E, Nakanishi T, Kawai A, et al. Expression of connective tissue growth factor/hypertrophic chondrocyte-specific gene product 24 (CTGF/Hcs24) during fracture healing. Bone. 2002;31(4):441–447. doi:10.1016/S8756-3282(02)00846-3
42. Zhang X. Interactions between cancer cells and bone microenvironment promote bone metastasis in prostate cancer. Cancer Commun. 2019;39(1):76. doi:10.1186/s40880-019-0425-1
43. Muruganandan S, Ionescu AM, Sinal CJ. At the crossroads of the adipocyte and osteoclast differentiation programs: future therapeutic perspectives. Int J Mol Sci. 2020;21(7):7. doi:10.3390/ijms21072277
44. Wang L, Zheng S, Huang G, et al. Osthole-loaded N-octyl-O-sulfonyl chitosan micelles (N SC-OST) inhibits RANKL-induced osteoclastogenesis and prevents ovariectomy-induced bone loss in rats. J Cell Mol Med. 2020;2:254.
45. Qi S, Perrino S, Miao X, Lamarche-Vane N, Brodt P. The chemokine CCL7 regulates invadopodia maturation and MMP-9 mediated collagen degradation in liver-metastatic carcinoma cells. Cancer Lett. 2020;483:98–113. doi:10.1016/j.canlet.2020.03.018
46. Qeadan F, Bansal P, Hanson JA, Beswick EJ. The MK2 pathway is linked to G-CSF, cytokine production and metastasis in gastric cancer: a novel intercorrelation analysis approach. J Transl Med. 2020;18(1):137. doi:10.1186/s12967-020-02294-z
47. Yang Z, Yue Z, Ma X, Xu Z. Calcium Homeostasis: A Potential Vicious Cycle of Bone Metastasis in Breast Cancers. Front Oncol. 2020;10:293. doi:10.3389/fonc.2020.00293
48. Bagheri H, Pourhanifeh MH, Derakhshan M, et al. CXCL-10: a new candidate for melanoma therapy? Cell Oncol. 2020;43(3):353–365. doi:10.1007/s13402-020-00501-z
49. Mao L, Zhuang R, Qin L, et al. CCL18 overexpression predicts a worse prognosis in oral squamous cell carcinoma (OSCC). Neoplasma. 2020;67(03):700–706. doi:10.4149/neo_2020_190821N802
50. Vieira-Filho D, Amarante MK, Ishibashi CM, et al. CCR5 and CXCL12 allelic variants: possible association with childhood neuroblastoma susceptibility? J Neuroimmunol. 2020;342:577193. doi:10.1016/j.jneuroim.2020.577193
51. Huang Y, Hakamivala A, Li S, et al. Chemokine releasing particle implants for trapping circulating prostate cancer cells. Sci Rep. 2020;10(1):4433. doi:10.1038/s41598-020-60696-x
52. Maynard JP, Ertunc O, Kulac I, Baena-Del VJ, De Marzo AM, Sfanos KS. IL8 Expression Is Associated with Prostate Cancer Aggressiveness and Androgen Receptor Loss in Primary and Metastatic Prostate Cancer. Mol Cancer Res. 2020;18(1):153–165. doi:10.1158/1541-7786.MCR-19-0595
53. Hellsten R, Lilljebjorn L, Johansson M, Leandersson K, Bjartell A. The STAT3 inhibitor galiellalactone inhibits the generation of MDSC-like monocytes by prostate cancer cells and decreases immunosuppressive and tumorigenic factors. Prostate. 2019;79(14):1611–1621. doi:10.1002/pros.23885
54. Dambal S, Alfaqih M, Sanders S, et al. 27-hydroxycholesterol impairs plasma membrane lipid raft signaling as evidenced by inhibition of IL6-JAK-STAT3 signaling in prostate cancer cells. Mol Cancer Res. 2020;18(5):671–684. doi:10.1158/1541-7786.MCR-19-0974
55. Xu M, Jiang H, Wang H, Liu J, Liu B, Guo Z. SB225002 inhibits prostate cancer invasion and attenuates the expression of BSP, OPN and MMP2. Oncol Rep. 2018;40(2):726–736. doi:10.3892/or.2018.6504
56. Chawla AS, Khalsa JK, Dhar A, et al. A role for cell-autocrine interleukin-2 in regulatory T cell homeostasis. Immunology. 2020;160(3):295–309. doi:10.1111/imm.13194
57. Ma QY, Chen J, Zhao J. Follicular cytotoxic CD8 T cells present high cytokine expression, and are more susceptible to Breg-mediated suppression in non-small cell lung cancer. Immunol Res. 2020;68(1):54–62. doi:10.1007/s12026-020-09120-0
58. Dang N, Lin Y, Waer M, Sprangers B. Donor lymphocyte-derived natural killer cells control major histocompatibility complex class I-negative melanoma. Cancer Immunol Res. 2020;8:756.
59. Hosseinzadeh F, Ai J, Ebrahimi-Barough S, et al. Natural killer cell expansion with autologous feeder layer and anti-cd3 antibody for immune cell therapy of hepatocellular carcinoma. Asian Pac J Cancer Prev. 2019;20(12):3797–3803. doi:10.31557/APJCP.2019.20.12.3797
60. Kulbe H, Levinson NR, Balkwill F, Wilson JL. The chemokine network in cancer–much more than directing cell movement. Int J Dev Biol. 2004;48(5–6):489–496. doi:10.1387/ijdb.041814hk
61. Craig MJ, Loberg RD. CCL2 (Monocyte Chemoattractant Protein-1) in cancer bone metastases. Cancer Metastasis Rev. 2006;25(4):611–619. doi:10.1007/s10555-006-9027-x
62. Lin TH, Liu HH, Tsai TH, et al. CCL2 increases alphavbeta3 integrin expression and subsequently promotes prostate cancer migration. Biochim Biophys Acta. 2013;1830(10):4917–4927. doi:10.1016/j.bbagen.2013.06.033
63. Chen Q, Lei JH, Bao J, et al. BRCA1 Deficiency Impairs Mitophagy and Promotes Inflammasome Activation and Mammary Tumor Metastasis. Adv Sci. 2020;7(6):1903616. doi:10.1002/advs.201903616
64. Song X, Wang W, Wang H, et al. Acetylation of ezrin regulates membrane-cytoskeletal interaction underlying CCL18-elicited cell migration. J Mol Cell Biol. 2019;12:424.
65. Hetmanski J, de Belly H, Busnelli I, et al. Membrane Tension Orchestrates Rear Retraction in Matrix-Directed Cell Migration. Dev Cell. 2019;51(4):460–475. doi:10.1016/j.devcel.2019.09.006
66. Moodley S, Lian EY, Crupi M, Hyndman BD, Mulligan LM. RET isoform-specific interaction with scaffold protein Ezrin promotes cell migration and chemotaxis in lung adenocarcinoma. Lung Cancer. 2020;142:123–131. doi:10.1016/j.lungcan.2020.02.004
67. Garcia-Ortiz A, Serrador JM. ERM Proteins at the Crossroad of Leukocyte Polarization, Migration and Intercellular Adhesion. Int J Mol Sci. 2020;21(4):1502. doi:10.3390/ijms21041502
68. Bompard G, Sharp SJ, Freiss G, Machesky LM. Involvement of Rac in actin cytoskeleton rearrangements induced by MIM-B. J Cell Sci. 2005;118(Pt 22):5393–5403. doi:10.1242/jcs.02640
69. Khanna C, Wan X, Bose S, et al. The membrane-cytoskeleton linker ezrin is necessary for osteosarcoma metastasis. Nat Med. 2004;10(2):182–186. doi:10.1038/nm982
70. Valdman A, Fang X, Pang ST, Nilsson B, Ekman P, Egevad L. Ezrin expression in prostate cancer and benign prostatic tissue. Eur Urol. 2005;48(5):852–857. doi:10.1016/j.eururo.2005.03.013
71. Mytilinaiou M, Nikitovic D, Berdiaki A, et al. IGF-I regulates HT1080 fibrosarcoma cell migration through a syndecan-2/Erk/ezrin signaling axis. Exp Cell Res. 2017;361(1):9–18. doi:10.1016/j.yexcr.2017.09.035
72. Ren F, Wang D, Wang Y, Chen P, Guo C. SPOCK2 Affects the Biological Behavior of Endometrial Cancer Cells by Regulation of MT1-MMP and MMP2. Reprod Sci. 2019;289123557.
73. Singh N, Rashid S, Rashid S, Dash NR, Gupta S, Saraya A. Clinical significance of promoter methylation status of tumor suppressor genes in circulating DNA of pancreatic cancer patients. J Cancer Res Clin Oncol. 2020;146(4):897–907. doi:10.1007/s00432-020-03169-y
74. Munasinghe A, Malik K, Mohamedi F, et al. Fibronectin acts as a molecular switch to determine SPARC function in pancreatic cancer. Cancer Lett. 2020;477:88–96. doi:10.1016/j.canlet.2020.02.031
75. Armstrong AJ, Gupta S, Healy P, et al. Pharmacodynamic study of radium-223 in men with bone metastatic castration resistant prostate cancer. PLoS One. 2019;14(5):e216934. doi:10.1371/journal.pone.0216934
76. Yang Y, Niu X, Liu W, Xu H. Expression and significance of secreted protein acidic and rich in cysteine in human osteosarcoma. Oncol Lett. 2017;14(5):5491–5496. doi:10.3892/ol.2017.6871
77. De S, Chen J, Narizhneva NV, et al. Molecular pathway for cancer metastasis to bone. J Biol Chem. 2003;278(40):39044–39050. doi:10.1074/jbc.M304494200
78. Kedzierska L, Madej-Michniewicz A, Marczuk N, Dolegowska B, Starzynska T, Blogowski W. Clinical significance of various growth factors in patients with different gastric neoplasms. Am J Transl Res. 2020;12(1):118–129.
79. Suh S, Kim KW. Diabetes and Cancer: cancer Should Be Screened in Routine Diabetes Assessment. Diabetes Metab J. 2019;43(6):733–743. doi:10.4093/dmj.2019.0177
80. Pirouzpanah S, Asemani S, Shayanfar A, Baradaran B, Montazeri V. The effects of Berberis vulgaris consumption on plasma levels of IGF-1, IGFBPs, PPAR-gamma and the expression of angiogenic genes in women with benign breast disease: a randomized controlled clinical trial. BMC Complement Altern Med. 2019;19(1):324. doi:10.1186/s12906-019-2715-1
81. Werner H, Sarfstein R, Bruchim I. Investigational IGF1R inhibitors in early stage clinical trials for cancer therapy. Expert Opin Investig Drugs. 2019;28(12):1101–1112. doi:10.1080/13543784.2019.1694660
82. Connolly JM, Rose DP. Regulation of DU145 human prostate cancer cell proliferation by insulin-like growth factors and its interaction with the epidermal growth factor autocrine loop. Prostate. 1994;24(4):167–175. doi:10.1002/pros.2990240402
83. Assikis VJ, Simons JW. Novel therapeutic strategies for androgen-independent prostate cancer: an update. Semin Oncol. 2004;31(2 Suppl 4):26–32. doi:10.1053/j.seminoncol.2004.02.008
84. Martel CL, Gumerlock PH, Meyers FJ, Lara PN. Current strategies in the management of hormone refractory prostate cancer. Cancer Treat Rev. 2003;29(3):171–187. doi:10.1016/S0305-7372(02)00090-7
85. Iwamura M, Sluss PM, Casamento JB, Cockett AT. Insulin-like growth factor I: action and receptor characterization in human prostate cancer cell lines. Prostate. 1993;22(3):243–252. doi:10.1002/pros.2990220307
86. Montagnani MM, Moretti RM, Procacci P, Motta M, Limonta P. Insulin-like growth factor-I promotes migration in human androgen-independent prostate cancer cells via the alphavbeta3 integrin and PI3-K/Akt signaling. Int J Oncol. 2006;28(3):723–730.
87. Noda S, Chikazawa K, Suzuki Y, Imaizumi Y, Yamamura H. Involvement of the gamma1 subunit of the large-conductance Ca(2+)-activated K(+) channel in the proliferation of human somatostatinoma cells. Biochem Biophys Res Commun. 2020;525(4):1032–1037. doi:10.1016/j.bbrc.2020.02.176
88. Rojas S, Basualto E, Valdivia L, et al. The activity of IKCa and BKCa channels contributes to insulin-mediated NO synthesis and vascular tone regulation in human umbilical vein. Nitric Oxide. 2020;99:7–16. doi:10.1016/j.niox.2020.03.004
89. Parlar A, Arslan SO, Cam SA. Glabridin Alleviates Inflammation and Nociception in Rodents by Activating BKCa Channels and Reducing NO Levels. Biol Pharm Bull. 2020;43(5):884–897. doi:10.1248/bpb.b20-00038
90. Du C, Zheng Z, Li D, et al. BKCa promotes growth and metastasis of prostate cancer through facilitating the coupling between alphavbeta3 integrin and FAK. Oncotarget. 2016;7(26):40174–40188. doi:10.18632/oncotarget.9559
91. Lau YK, Ramaiyer M, Johnson DE, Grandis JR. Targeting STAT3 in Cancer with Nucleotide Therapeutics. Cancers. 2019;11(11):1681.
92. Huang B, Deng W, Wang DX. [Effects of p38 mitogen-activated protein kinase in rats with oleic acid-induced acute lung injury]. Zhongguo Ying Yong Sheng Li Xue Za Zhi. 2017;33(4):334–339. Chinese. doi:10.12047/j.cjap.5552.2017.081
93. Mohanty A, Pharaon RR, Nam A, Salgia S, Kulkarni P, Massarelli E. FAK-targeted and combination therapies for the treatment of cancer: an overview of Phase I and II clinical trials. Expert Opin Investig Drugs. 2020;29:1–11.
94. Tapial MP, Lopez NP, Lietha D. FAK Structure and Regulation by Membrane Interactions and Force in Focal Adhesions. Biomolecules. 2020;10(2):179.
95. Castoria G, D’Amato L, Ciociola A, et al. Androgen-induced cell migration: role of androgen receptor/filamin A association. PLoS One. 2011;6(2):e17218. doi:10.1371/journal.pone.0017218
96. Di Donato M, Cernera G, Auricchio F, Migliaccio A, Castoria G. Cross-talk between androgen receptor and nerve growth factor receptor in prostate cancer cells: implications for a new therapeutic approach. Cell Death Discov. 2018;4(1):5. doi:10.1038/s41420-017-0024-3
97. Di Donato M, Cernera G, Migliaccio A, Castoria G. Nerve Growth Factor Induces Proliferation and Aggressiveness In Prostate Cancer Cells. Cancers. 2019;11(6):784. doi:10.3390/cancers11060784
98. Spans L, Helsen C, Clinckemalie L, et al. Comparative genomic and transcriptomic analyses of LNCaP and C4-2B prostate cancer cell lines. PLoS One. 2014;9(2):e90002. doi:10.1371/journal.pone.0090002
99. Deng Z, Zeng Q, Chai J, et al. Disintegrin tablysin-15 suppresses cancer hallmarks in melanoma cells by blocking FAK/Akt/ERK and NF-kappaB signaling. Curr Cancer Drug Targets. 2019;20:306.
100. Zacharopoulou N, Kallergi G, Alkahtani S, et al. The histone demethylase KDM2B activates FAK and PI3K that control tumor cell motility. Cancer Biol Ther. 2020;21:1–8.
101. Molla MS, Katti DR, Iswara J, Venkatesan R, Paulmurugan R, Katti KS. Prostate cancer phenotype influences bone mineralization at metastasis: a study using an in vitro prostate cancer metastasis testbed. JBMR Plus. 2020;4(2):e10256. doi:10.1002/jbm4.10256
102. Liu FL, Chen CL, Lai CC, Lee CC, Chang DM. Arecoline suppresses RANKL-induced osteoclast differentiation in vitro and attenuates LPS-induced bone loss in vivo. Phytomedicine. 2020;69:153195. doi:10.1016/j.phymed.2020.153195
103. Hong Z, Fang Z, Lei J, et al. The significance of Runx2 mediating alcohol-induced Brf1 expression and RNA Pol III gene transcription. Chem Biol Interact. 2020;323:109057. doi:10.1016/j.cbi.2020.109057
104. Geng YM, Liu CX, Lu WY, et al. LAPTM5 is transactivated by RUNX2 and involved in RANKL trafficking in osteoblastic cells. Mol Med Rep. 2019;20(5):4193–4201. doi:10.3892/mmr.2019.10688
105. Miyamoto S, Teramoto H, Gutkind JS, Yamada KM. Integrins can collaborate with growth factors for phosphorylation of receptor tyrosine kinases and MAP kinase activation: roles of integrin aggregation and occupancy of receptors. J Cell Biol. 1996;135(6 Pt 1):1633–1642. doi:10.1083/jcb.135.6.1633
106. Klemke RL, Cai S, Giannini AL, Gallagher PJ, de Lanerolle P, Cheresh DA. Regulation of cell motility by mitogen-activated protein kinase. J Cell Biol. 1997;137(2):481–492. doi:10.1083/jcb.137.2.481
107. Khwaja A, Rodriguez-Viciana P, Wennstrom S, Warne PH, Downward J. Matrix adhesion and Ras transformation both activate a phosphoinositide 3-OH kinase and protein kinase B/Akt cellular survival pathway. EMBO J. 1997;16(10):2783–2793. doi:10.1093/emboj/16.10.2783
108. Cary LA, Han DC, Polte TR, Hanks SK, Guan JL. Identification of p130Cas as a mediator of focal adhesion kinase-promoted cell migration. J Cell Biol. 1998;140(1):211–221. doi:10.1083/jcb.140.1.211
109. Wary KK, Mainiero F, Isakoff SJ, Marcantonio EE, Giancotti FG. The adaptor protein Shc couples a class of integrins to the control of cell cycle progression. Cell. 1996;87(4):733–743. doi:10.1016/S0092-8674(00)81392-6
110. Horwitz AR, Parsons JT. Cell migration–movin’ on. Science. 1999;286(5442):1102–1103. doi:10.1126/science.286.5442.1102
111. Hu K, Tang X, Tang G, et al. 18F-FP-PEG2-beta-Glu-RGD2: A Symmetric Integrin alphavbeta3-Targeting Radiotracer for Tumor PET Imaging. PLoS One. 2015;10(9):e138675.
112. Beer AJ, Schwarzenbock SM, Zantl N, et al. Non-invasive assessment of inter-and intrapatient variability of integrin expression in metastasized prostate cancer by PET. Oncotarget. 2016;7(19):28151–28159. doi:10.18632/oncotarget.8611
113. Lucente E, Liu H, Liu Y, et al. Novel (64)Cu Labeled RGD2-BBN Heterotrimers for PET Imaging of Prostate Cancer. Bioconjug Chem. 2018;29(5):1595–1604. doi:10.1021/acs.bioconjchem.8b00113
114. Cook G, Azad GK, Taylor BP, et al. Imaging alphavbeta3 integrin expression in skeletal metastases with (99m)Tc-maraciclatide single-photon emission computed tomography: detection and therapy response assessment. Eur J Nucl Med Mol Imaging. 2018;45(6):898–903. doi:10.1007/s00259-017-3926-7
115. Zhang J, Niu G, Lang L, et al. Clinical translation of a dual integrin alphavbeta3- and gastrin-releasing peptide receptor-targeting PET Radiotracer, 68Ga-BBN-RGD. J Nucl Med. 2017;58(2):228–234. doi:10.2967/jnumed.116.177048
116. Nouman M, Haider G, Bukhari N, et al. Response rate of cisplatin plus docetaxel as primary treatment in locally advanced head and neck carcinoma (squamous cell types). Asian Pac J Cancer Prev. 2020;21(3):825–830. doi:10.31557/APJCP.2020.21.3.825
117. Kulhari H, Pooja D, Shrivastava S, et al. Cyclic-RGDfK peptide conjugated succinoyl-TPGS nanomicelles for targeted delivery of docetaxel to integrin receptor over-expressing angiogenic tumours. Nanomedicine-Uk. 2015;11(6):1511–1520. doi:10.1016/j.nano.2015.04.007
118. Gao Y, Zhou Y, Zhao L, et al. Enhanced antitumor efficacy by cyclic RGDyK-conjugated and paclitaxel-loaded pH-responsive polymeric micelles. Acta Biomater. 2015;23:127–135. doi:10.1016/j.actbio.2015.05.021
119. Zhang L, Shan X, Meng X, et al. The first integrins beta3-mediated cellular and nuclear targeting therapeutics for prostate cancer. Biomaterials. 2019;223:119471. doi:10.1016/j.biomaterials.2019.119471
120. Wang F, Chen L, Zhang R, Chen Z, Zhu L. RGD peptide conjugated liposomal drug delivery system for enhance therapeutic efficacy in treating bone metastasis from prostate cancer. J Control Release. 2014;196:222–233. doi:10.1016/j.jconrel.2014.10.012
121. Dong W, van Ginkel JW, Au KY, Alemany R, Meulenberg JJ, van Beusechem VW. ORCA-010, a novel potency-enhanced oncolytic adenovirus, exerts strong antitumor activity in preclinical models. Hum Gene Ther. 2014;25(10):897–904. doi:10.1089/hum.2013.229
122. Yang YF, Xue SY, Lu ZZ, et al. Antitumor effects of oncolytic adenovirus armed with PSA-IZ-CD40L fusion gene against prostate cancer. Gene Ther. 2014;21(8):723–731. doi:10.1038/gt.2014.46
123. Satoh M, Wang H, Ishidoya S, et al. Oncolytic virotherapy for prostate cancer by E1A, E1B mutant adenovirus. Urology. 2007;70(6):1243–1248. doi:10.1016/j.urology.2007.09.031
124. Shen YH, Yang F, Wang H, et al. Arg-Gly-Asp (RGD)-Modified E1A/E1B double mutant adenovirus enhances antitumor activity in prostate cancer cells in vitro and in mice. PLoS One. 2016;11(1):e147173.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.