Back to Journals » International Journal of Nanomedicine » Volume 21
RNA Sequencing Explores Potential Mechanisms Underlying the Enhanced Aggressive Phenotype of HepG2 Cells Induced by Aged Zinc Oxide Nanoparticles
Authors Xu J, Yu N, Yang J, Wang S ![]() , Lin T, Wang M
, Lin T, Wang M
Received 19 February 2026
Accepted for publication 6 May 2026
Published 13 May 2026 Volume 2026:21 604287
DOI https://doi.org/10.2147/IJN.S604287
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Kamakhya Prakash Misra
Jin Xu, Na Yu, Jingya Yang, Siyu Wang, Tong Lin, Meimei Wang
Department of Pathophysiology, School of Basic Medical Science, Anhui Medical University, Hefei, Anhui, 230032, People’s Republic of China
Correspondence: Meimei Wang, Email [email protected]
Purpose: While aged zinc oxide nanoparticles (ZnO NPs) show reduced acute cytotoxicity, their chronic effects remain unclear. This study compared the long-term, low-dose impact of environmentally aged versus fresh ZnO NPs on enhanced aggressive phenotype in hepatocellular carcinoma cells.
Material and Methods: HepG2 cells were chronically exposed (approximately 16 weeks) to low doses (1.5 μg/mL) of aged or fresh ZnO NPs. RNA sequencing (n=3 per group) identified transcriptomic changes, while migration and invasion assays validated functional outcomes. A transcription factor (TF) -mRNA network was constructed, and clinical correlation was analyzed using patient survival data.
Results: Aged ZnO NPs were associated with the activation of pro-oncogenic pathways (e.g. JAK-STAT) and induced increased cell migration and invasion compared to fresh NPs. Network analysis suggested CEBPA, CTNNB1, and STAT3 as potential core transcriptional regulators. Consistent with their potential role in promoting an aggressive phenotype, the elevated expression levels of these TFs were associated with reduced overall survival in patients with hepatocellular carcinoma.
Conclusion: Chronic exposure to environmentally aged ZnO NPs, despite lower acute toxicity, may promote a more aggressive phenotype in liver cells compared to fresh NPs. This effect is potentially mediated by the specific modulation of oncogenic pathways and a core transcriptional network linked to poor patient outcomes. Our findings suggest that the environmental aging process could be an important factor influencing the long-term potential of ZnO NPs to promote an aggressive phenotype, which should be considered in nanomaterial risk assessment. The diagram illustrates a process flow starting with aging and cell culture. It shows control, fresh nanoparticles and aged nanoparticles, followed by characterization using TEM, DLS and ICP-OES. Next is RNA sequencing, enrichment and the TF-mRNA network. Functional validation is depicted with a container and particles. Finally, clinical correlation is shown with a graph of survival probability over time, comparing high and low expression levels.Diagram showing aging and cell culture, RNA sequencing, functional validation and clinical correlation processes.
Keywords: zinc oxide nanoparticles, aging, HepG2 cells, transcriptomics, enhanced aggressive phenotype
Introduction
The widespread application of engineered nanomaterials, particularly zinc oxide nanoparticles (ZnO NPs), in consumer products, agriculture, and biomedicine has led to their increasing environmental release,1–3 raising concerns about potential human health risks.4,5 For instance, acute exposure to ZnO NPs has been observed to induce increased airway resistance, hyperinflammatory response, and histological damage in rat lungs, while in vitro studies demonstrate the induction of ROS production, decrease in mitochondrial membrane potential, and activation of the NLRP3 inflammasome in A549 cells.6 Furthermore, Qin et al reported that ZnO NPs can induce ferroptosis in HUVECs and EA.hy926 cells, with elevated intracellular iron levels and lipid peroxidation in a dose- and time-dependent manner.7 Similarly, Valdiglesias et al discovered that ZnO NPs induce considerable cytotoxicity in glial cells, including decreased viability and cytoskeletal alterations, also following dose- and time-dependent kinetics.8
The hepatotoxicity induced by ZnO NPs, as evidenced by the accumulation of experimental data, underscores the liver’s role as a primary target organ for NPs accumulation.9,10 Given this specific susceptibility and the substantial global burden of hepatocellular carcinoma (HCC) – which ranks as the sixth most common malignancy worldwide and the third leading cause of cancer-related mortality11,12 – it is critically important to evaluate the potential risks of ZnO NPs specifically within the context of liver cancer. This concern is particularly acute in Asia, as evidenced by China constituting an estimated 42.4% of global HCC cases.13 Consequently, the human hepatocellular carcinoma cell line HepG2 is frequently selected by nanotoxicologists to directly simulate the potential effects of chronic ZnO NPs exposure on the primary target organ within a disease-relevant context. HepG2 cells are a well-established in vitro model for liver toxicity and cancer biology studies. They not only retain many functions of normal hepatocytes, making them highly relevant for assessing hepatotoxic effects, but also provide a malignant cellular background ideal for investigating the potential of environmental stressors to exacerbate cancer hallmarks, such as increased invasion and migration.14–18 Typically, after internalization, ZnO NPs are primarily absorbed as Zn2+ ions, with a fraction internalized as particulate matter; regardless of the exposure route, this leads to elevated hepatic zinc concentrations.19 Supporting this, a study on crucian carp revealed that ZnO NPs exposure increased serum levels of liver injury biomarkers, including aspartate aminotransferase (AST) and alanine aminotransferase (ALT). Histopathological observations further showed inflammatory cell infiltration within the central vein of the liver following ZnO NPs stimulation.20 Moreover, in a rat model subjected to a 21-day exposure (2 mg/kg/day), liver biopsies exhibited significant tissue damage, including sinusoidal dilatation, Kupffer cell hyperplasia, inflammatory cell infiltration, and hepatocyte apoptosis.21 Here, by employing the HepG2 model, we aim to directly assess whether ZnO NPs enhance the aggressive behaviors of hepatocellular carcinoma cells.
Following their release into the environment, ZnO NPs undergo significant physicochemical transformations influenced by surrounding conditions.22–24 These alterations primarily include dissolution, aggregation, and chemical speciation changes,25 which are influenced by key environmental factors such as pH, organic matter, ionic strength, and sunlight.26,27 For instance, in phosphate-rich waters,28 ZnO NPs can be sequentially transformed into crystalline and subsequently amorphous zinc phosphate phases, a process that significantly attenuates their cytotoxicity.29 Conversely, research by Saji George et al confirmed that exposure to light can significantly increase the toxic potential of ZnO NPs to environmental organisms.30 A critical transformation process that warrants particular attention is environmental aging.31,32 This process can potentially change the NPs’ surface properties, reactivity, and environmental persistence.33,34 Our earlier work has demonstrated that during this aging process, ZnO NPs can transform into secondary species, including zinc ions (Zn2+) and hydrozincite (Zn5(CO3)2(OH)6).35 This transformation may further trigger a series of interrelated physicochemical alterations of NPs. For instance, changes in dissolution kinetics could influence the pattern of Zn2+ release, while the formation of new phases or the adsorption of environmental molecules may change the particles’ surface reactivity, stability, and their mode of interaction with cells. Therefore, to accurately assess the real-world health risks of ZnO NPs, it is essential to investigate how these aging-induced physicochemical alterations translate into biological outcomes, particularly under exposure conditions that mirror environmental relevance. However, the predominant toxicological paradigm for ZnO NPs has relied on acute or sub-acute, high-dose exposures to pristine NPs. While invaluable for elucidating immediate cytotoxic mechanisms, this paradigm does not reflect the chronic, low-level exposure scenarios relevant to environmental or occupational settings.36,37 Consequently, a critical yet underexplored question arises: how does the environmental aging of ZnO NPs – a nanomaterial of high production volume and human exposure potential – modulate their biological effects under chronic, low-dose exposure conditions, and can such exposure drive more insidious pathological processes, such as the progression of a pre-existing malignant phenotype?38–41
Our previous investigations have systematically compared the biological effects of fresh and aged ZnO NPs, providing a foundation for this study. While acute cytotoxicity might be attenuated, the potential to induce DNA damage and gene mutations could be significantly enhanced after aging.42 Building on these findings, this study employs RNA sequencing to delve into the underlying molecular mechanisms. We hypothesized that chronic, low-dose exposure to aged ZnO NPs would induce distinct transcriptomic profiles in HepG2 compared to fresh NPs, potentially activating pathways implicated in cancer progression, such as cell proliferation and invasion. Our specific objectives were to: (1) identify differentially expressed genes (DEGs); (2) perform functional enrichment analysis and Gene Set Enrichment Analysis (GSEA) to delineate perturbed biological processes; and (3) construct regulatory networks to identify key transcription factors and assess the clinical relevance of identified genes. This work aims to provide insights into the mechanisms underlying the health risks associated with chronic exposure to environmentally transformed ZnO NPs.
Material and Methods
Characterization of ZnO NPs
Commercially available ZnO NPs with an average particle size of 20 nm, a purity of 99.5%, and a near-spherical morphology were obtained from Nanostructured & Amorphous Materials (Houston, TX, USA). A fresh ZnO NPs suspension (1 mg/mL) was prepared in Milli-Q water and sterilized at 120°C for 30 minutes. Prior to each experiment, the suspension was vortexed for 5 minutes and ultrasonicated for 30 minutes to prevent aggregation. Aged ZnO NPs were obtained by natural aging of the suspension in Milli-Q water at 25°C for 60 days. The synthesis and characterization methodology for the ZnO NPs followed the protocol described by Wang et al35 In detail, fresh and aged ZnO NPs were characterized by transmission electron microscopy (TEM, Carl Zeiss 120, Zeiss International, Germany) to confirm morphological changes. To comprehensively evaluate properties relevant to biological interactions, the dissolution behavior, hydrodynamic size, and surface charge of the NPs were characterized. For dissolution analysis, NPs were suspended at 50 µg/mL in either Milli-Q water or complete cell culture medium (DMEM supplemented with 10% FBS), incubated at 37 °C for 72 hours, and then centrifuged at 20,000 rpm for 1 hour. Zinc concentration in the supernatants was measured by inductively coupled plasma optical emission spectroscopy (ICP-OES, Agilent 5110) against a standard curve. Culture medium samples were subjected to microwave digestion prior to ICP-OES analysis. Hydrodynamic diameter and zeta potential were determined by dynamic light scattering (DLS) using a Zetasizer Nano ZSE (Malvern Instruments, UK). For DLS measurements, NPs were dispersed in ultrapure water at 100 µg/mL, followed by vortex mixing and ultrasonication. All measurements were performed in at least triplicate, and data are presented as mean ± standard deviation (SD).
Cell Culture
The human hepatoma cell line HepG2 was acquired from the China Center for Type Culture Collection (CCTCC, Shanghai, China). Cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM; Gibco, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, USA) and 1% penicillin-streptomycin (Hyclone, USA). All cell cultures were maintained at 37°C in a humidified incubator with 5% CO2.
Cell Viability Assay
To assess acute cytotoxicity, HepG2 cells were seeded in 6-well plates at a density of 1×105 cells per well. After 24 hours of adhesion, cells were exposed to fresh or aged ZnO NPs at concentrations of 1, 5, 10, 15, 20, 30, and 50 µg/mL for 72 hours. Cell viability was then determined by cell counting. Cell viability for each treatment group was expressed as a percentage relative to the untreated control group. Each concentration was tested in duplicate wells, and the entire experiment was independently repeated three times.
In vitro Chronic Exposure to ZnO NPs
Based on the acute cytotoxicity results, a sub-cytotoxic concentration of 1.5 µg/mL was selected for chronic exposure studies. HepG2 cells were continuously exposed to this concentration of fresh or aged ZnO NPs for approximately 16 weeks (equivalent to 40 cell passages). To maintain consistent exposure, the culture medium containing NPs was replenished every two to three days. Control cultures were maintained in parallel without NPs.
RNA-Seq Data Processing, Differential Expression, and Visualization
Total RNA was extracted from chronically exposed HepG2 cells from three independent biological replicates per group (n=3) using TRIzol reagent. Sequencing libraries were prepared and sequenced on an Illumina NovaSeq 6000 platform. After quality control, clean reads were aligned to the reference genome using HISAT2, and gene- level counts were quantified with HTSeq (0.9.1), and the raw expression levels were subsequently normalized to FPKM (Fragments Per Kilobase of transcript per Million mapped reads) to account for gene length and sequencing depth. Differentially expressed genes (DEGs) were identified using the DESeq2 R package (version 1.48.2). Low-expressed genes (counts < 10 in at least 3 samples) were filtered out prior to analysis. Pairwise comparisons (Fresh NPs vs. Control, Aged NPs vs. Control, Aged NPs vs. Fresh NPs) were performed. DEGs were defined as genes with an adjusted p-value (padj; Benjamini-Hochberg correction) < 0.05 and an absolute log2FoldChange (|log2FC|) ≥ 1. Principal component analysis (PCA) was conducted on variance-stabilized counts. The top 500 most variable genes were used for dimensionality reduction, and the results were visualized in a two-dimensional scatter plot of the first two principal components, with 95% confidence ellipses added to illustrate group clustering. Volcano plots for each comparison were generated using ggplot2 (version 4.0.2) and ggrepel (version 0.9.6).
Functional and Pathway Enrichment Analysis
Functional and pathway enrichment analyses were conducted to interpret the biological significance of the differentially expressed genes (DEGs). Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed for each comparison using the clusterProfiler R package (version 4.16.0), with the org.Hs.egdb database (version 3.21.0) used for annotation. The complete set of DEGs (both up- and down-regulated) was analyzed against the background of all expressed genes, with significance defined as p < 0.05. In addition, Gene Set Enrichment Analysis (GSEA) was performed on genome-wide gene lists ranked by the Wald statistic from DESeq2, using candidate GO terms and KEGG pathways identified in the enrichment analysis as input gene sets. The GSEA, implemented with the fgsea method in clusterProfiler, assessed significance with p < 0.05. Results were visualized using dot plots (ggplot2, version 4.0.2) and single-gene-set enrichment plots (GseaVis, version 0.0.5).
Regulatory Network Construction and Clinical Correlation
A transcriptional regulatory network was inferred to identify key transcription factors (TFs) modulating the observed gene expression changes. Potential TF- mRNA interactions were predicted using the collecTRI method within the decoupleR package. The network was visualized using Cytoscape software. To assess the clinical relevance of the identified key TFs, survival analysis was performed using the R package easyTCGA. This analysis utilized the clinical and gene expression data from the Liver Hepatocellular Carcinoma (LIHC) cohort of The Cancer Genome Atlas (TCGA). Kaplan-Meier survival curves were generated based on the optimal cut-point expression level of the key TFs to evaluate the association between their expression and the overall survival of LIHC patients.
Cell Migration and Invasion Assays
Cell migration and invasion were assessed using Transwell inserts (8 μm pore membrane; BD Biosciences, USA). For the migration assay, a suspension of 5×104 cells in 200 µL of serum-free medium was seeded into the upper chamber. The lower chamber contained 600 µL of complete medium with 10% FBS as a chemoattractant. For the invasion assay, the membrane was pre-coated with a thin layer of Matrigel (diluted 1:8 in serum-free medium). Following a 48-hour incubation, non-migratory/non-invasive cells on the upper surface were removed. Cells on the lower surface were fixed with 4% paraformaldehyde, stained with 0.1% crystal violet, and counted under an inverted microscope. Cells from five randomly selected fields per insert were counted using ImageJ software.The experiments were conducted with three independent biological replicates (n=3), each including triplicate wells.
Quantitative Real-Time PCR (RT-qPCR) Analysis
Total RNA was extracted from HepG2 cells using TRIzol reagent (Invitrogen, USA). cDNA was synthesized from total RNA using the ToioScript All-in-one RT EasyMix for qPCR (TOLOBIO, China) under conditions recommended by the manufacturer. Quantitative PCR was performed on a StepOnePlus Real-Time PCR System (Applied Biosystems, USA) using 2 × Q3 SYBR qPCR Master Mix (Universal) (TOLOBIO, China) with gene-specific primers (Sangon Biotech, China). The thermal cycling conditions were: 95°C for 1 minute; 40 cycles of 95°C for 10 seconds, 60°C for 10 seconds, and 72°C for 30 seconds. The relative expression of target genes was normalized to GAPDH and calculated using the 2^(–ΔΔCt) method with triplicate replicates. Each sample was run in technical triplicate, and the experiment was conducted with three independent biological replicates (n=3). The sequences of all primers used are listed in Table S1.
Statistical Analysis
The normality of data distribution was assessed using the Shapiro–Wilk test, and the homogeneity of variances was verified using Levene’s test. Statistical analysis was performed on data from three independent experiments. All data are presented as the mean ± standard deviation (SD). The specific choice of inferential statistical test was determined by the experimental design. For direct comparisons between two independent groups, such as evaluating the difference in migratory or invasive capacity between cells treated with aged versus fresh ZnO NPs, an unpaired two-tailed Student’s t-test was applied. For experiments involving comparisons among three groups, including the cell viability assay and the multi-group gene expression analysis in RT-qPCR, a one-way analysis of variance (ANOVA) was first conducted. Where the ANOVA indicated a statistically significant overall effect, Tukey’s honestly significant difference (HSD) post hoc test was employed for all pairwise comparisons to identify which specific groups differed from one another. In all graphical representations of the data, statistical significance is denoted by asterisks (*) for comparisons against the control group and hash symbols (#) for the direct comparison between aged and fresh NPs treatments, with *# indicating p < 0.05 and **## indicating p < 0.01. A p-value (or the adjusted p-value from post hoc tests) of less than 0.05 was considered statistically significant.
Results
Characterization of NPs
TEM images revealed significant differences in the surface morphology of NPs dispersed in Mill-Q water at a concentration of 100 μg/mL. Although fresh ZnO NPs lack a particularly regular spherical structure, the surface of their regular crystal structure remains relatively clear; in contrast, aged nanoparticles display amorphous and blurred morphological features (Figure 1). Table 1 summarized the hydrodynamic diameter, polydispersity index (PdI), and soluble Zn levels of both fresh and aged ZnO NPs, as measured in Milli-Q water and DMEM medium. Upon dilution in the medium, the agglomerate size of aged ZnO NPs dropped from 692 nm to 434 nm. This may be attributed to fetal bovine serum (FBS) and other components in the cell culture medium, which both stabilize the NPs and aid in their dispersion. Concurrently, dissolved Zn concentrations rose substantially—from 4.61 μg/mL to 13.79 μg/mL for fresh NPs, and from 9.41 μg/mL to 19.85 μg/mL for aged NPs. Such enhanced ion release implies that the observed biological effects of ZnO NPs may be partially driven by Zn ions leached into the culture environment. Notably, dilution in cell culture medium significantly boosted the dissolution of both NP types. The particulars of the physicochemical transformation process have been subjected to exhaustive analysis and detailed exposition in our previous researches.35,
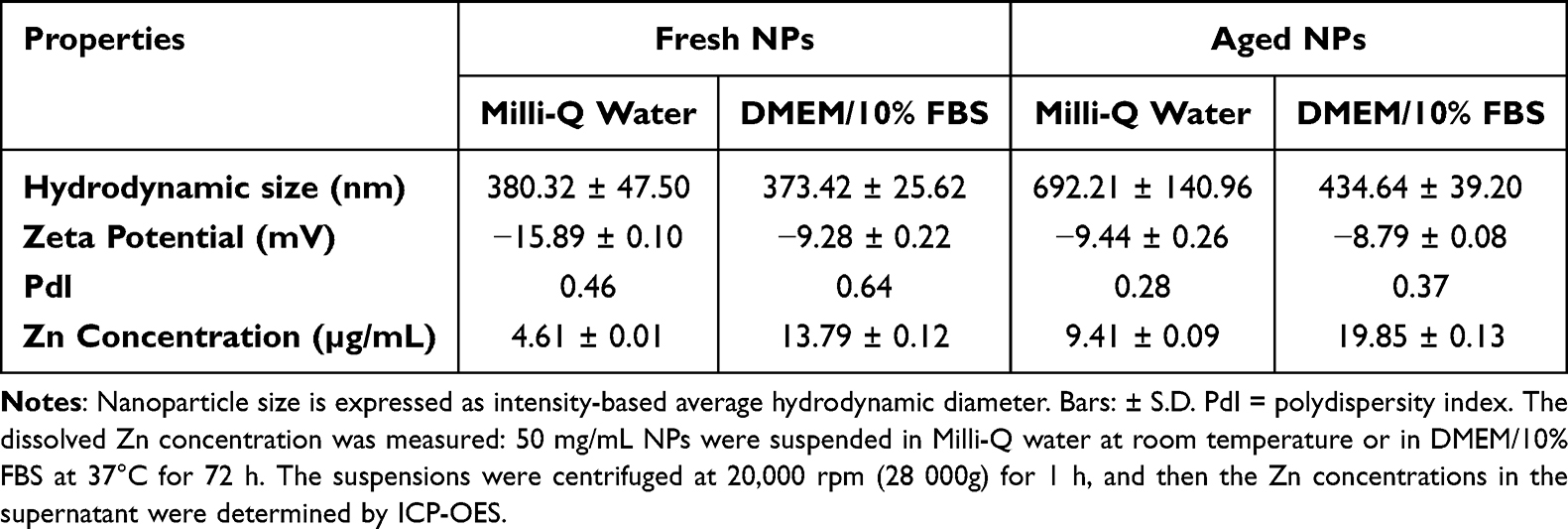 |
Table 1 Particle Size and Zinc Concentration of Fresh and Aged NP Suspensions |
 |
Figure 1 Representative micrographs of fresh and aged NPs in Milli-Q water using low resolution TEM. |
Acute Cytotoxicity of NPs and Dose Selection for Chronic Exposure
HepG2 cells were acutely treated with fresh or aged NPs at varying concentrations for 72 hours. As shown in Figure 2, cell viability progressively decreased with increasing NPs concentration, though the rate of decline differed between aged and fresh groups. Specifically, fresh NPs exhibited significant toxicity from 5 μg/mL onwards, with a marked decline in cell viability; whereas aged NPs exhibited no apparent toxicity below 10 μg/mL, with a comparatively slower decline in cell viability. Notably, when the nanomaterial concentration reached 20 μg/mL, the survival rate in the fresh group fell below 30%, whereas cells in the aged group retained approximately 65% viability. To model a chronic, low-level exposure scenario, a concentration of 1.5 µg/mL, which was demonstrated to exert negligible effects on cell viability in our acute exposure assays, was selected as the possible sub-toxic dose for the subsequent 4-month chronic exposure study.
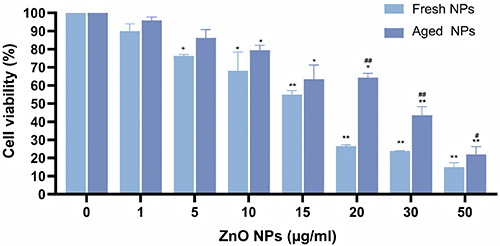 |
Figure 2 Acute cytotoxicity of fresh and aged NPs in HepG2 cells. Cells were treated with the indicated concentrations (0–50 µg/mL) of NPs for 72 h. Cell viability is presented as the mean percentage of the untreated control ± SD (n = 3). *p < 0.05, **p < 0.01 vs control group. ##p < 0.01 for aged NPs versus fresh NPs groups at the same concentration. |
Transcriptome Sequencing Indicates Specific Molecular Alterations Chronically Induced by ZnO NPs
Sequencing of nine samples generated 410.39 million raw reads, averaging 45.60 M reads per sample. Rigorous filtering retained 95.01% of raw reads (389.89 M clean reads), confirming efficient removal of low-quality sequences. The data exhibited high accuracy, with mean Q30 scores of 93.94% across all groups (Control: 94.07%, Fresh NPs: 94.14%, Aged NPs: 93.60%).
Differential gene expression analysis across the three groups was carried out, with thresholds set as |log2FoldChange| ≥ 1 and padj < 0.05. PCA plot (Figure 3a) visually demonstrates the intra -group consistency and inter -group differences, effectively distinguishing the gene expression profiles of the three groups. Venn diagram analysis (Figure 3b) of the DEGs showed that the comparison of fresh NPs versus control groups yielded 33 unique DEGs, the comparison of aged NPs versus control groups yielded 342 unique DEGs, and the two comparisons shared a common set of 66 overlapping DEGs, underscoring both conserved and treatment-specific transcriptional alterations. The volcano plots in Figure 3c–e respectively depict the distribution of DEGs for the comparisons of fresh NPs versus control groups, aged NPs versus control groups, and fresh NPs versus aged NPs groups. These plots clearly illustrate genes with significant expression changes, showcasing the up -regulated and down -regulated genes in each comparison group.
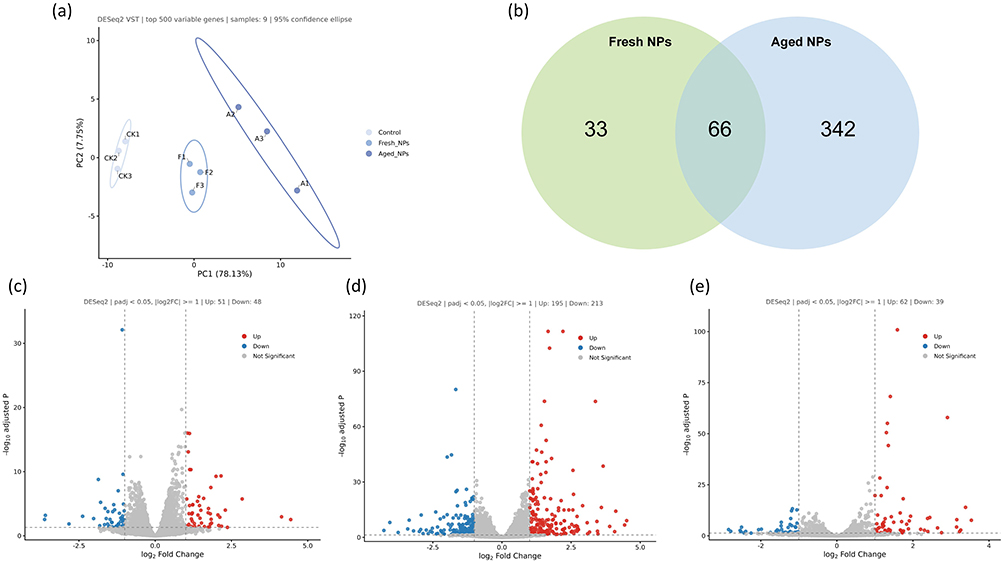 |
Figure 3 Transcriptomic profiling of HepG2 cells after chronic exposure to fresh or aged NPs. (a) PCA plot of transcriptomes from nine samples across control, fresh NPs, and aged NPs groups. (b) Venn diagram illustrating the overlap of DEGs identified in fresh NPs vs control groups comparison and aged NPs vs control groups comparison. (c–e) Volcano plots displaying the distribution of DEGs for the comparisons of (c) fresh NPs vs control groups, (d) aged NPs vs control groups, and (e) fresh NPs vs aged NPs groups. padj < 0.05 was considered statistically significant. |
GO Analysis of Differential Gene Expression Chronically Induced by ZnO NPs
GO enrichment analysis identified a total of 127, 614, and 378 significantly enriched GO terms in the comparison of fresh NPs group versus control group, aged NPs group versus control group, and aged NPs group versus fresh NPs group, respectively. Within the Biological Process (BP) category, 91, 460, and 313 GO terms were significantly enriched across these groups. To further elucidate the most biologically relevant processes, the top 10 most representative GO terms in the BP category for each group were selected and are presented in Figure 4. Positive regulation of cell migration, G protein-coupled receptor signaling pathway, signal release and regulation of epithelial cell proliferation are closely related to fresh NPs exposure (Figure 4a). After treatment with aged NPs, significant changes occur in epithelial cell proliferation, extracellular matrix organization, cell-cell adhesion via plasma-membrane adhesion molecules and extracellular matrix disassembly (Figure 4b). In comparison to fresh NPs group, aged NPs group exhibit alterations in cell-matrix adhesion, cytokine-mediated signaling pathway, fatty acid transport and very long-chain fatty acid metabolic process (Figure 4c).
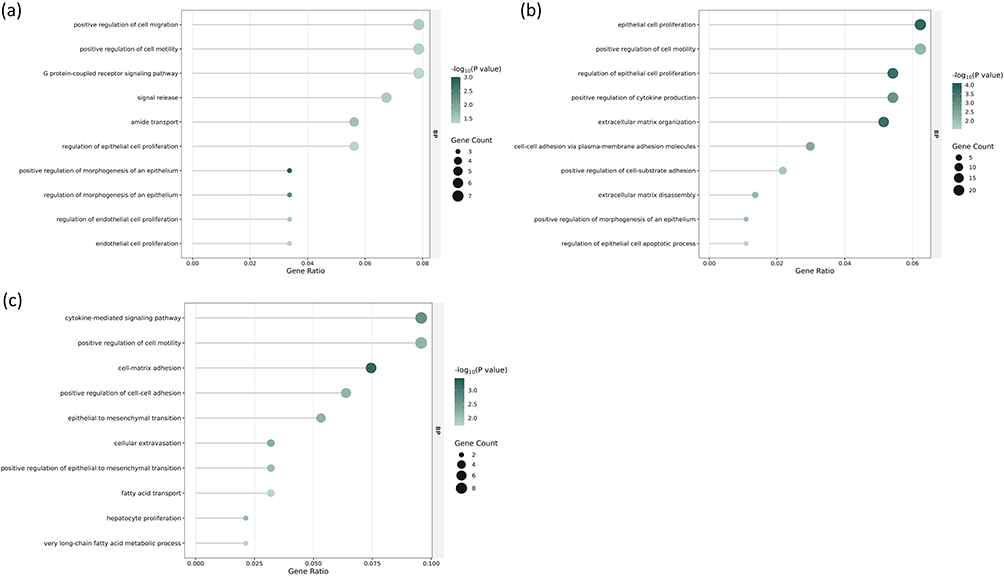 |
Figure 4 GO terms in the BP category for comparisons between fresh ZnO NPs, aged ZnO NPs, and control groups. (a) Top 10 most representative enriched BP terms for fresh ZnO NPs vs control comparison. (b) Top 10 most representative BP terms for aged ZnO NPs vs control comparison. (c) Top 10 most representative BP terms for aged ZnO NPs vs fresh ZnO NPs comparison. −log10(p value), negative logarithm of the p-value. p < 0.05 was considered statistically significant. |
KEGG and GSEA Suggest Specific Pathway Enrichment Profiles Chronically Induced by ZnO NPs
To further elucidate the functional changes of DEGs, KEGG pathway analysis was performed. In the comparison of fresh NPs versus control, the analysis revealed significant enrichment in pathways including Tyrosine metabolism, Phenylalanine metabolism, ECM-receptor interaction, and Calcium signaling pathway (Figure 5a), with these pathways showing notable statistical support among the top-ranked candidates. In aged NP- treated groups, pathways that demonstrated statistically significant enrichment and are mechanistically relevant to cancer biology included Cytokine-cytokine receptor interaction and Tyrosine metabolism and Gap junction (Figure 5b). Furthermore, the comparison between aged and fresh NPs identified several pathways with statistically supported enrichment and clear relevance to hepatocellular carcinoma progression, including Cytokine-cytokine receptor interaction, ECM-receptor interaction, Cytosolic DNA-sensing pathway and the Jak-STAT signaling pathway (Figure 5c). GSEA analysis (Figure 5d–f) revealed that aged NPs group, compared to fresh NPs group, exhibited marked up-regulation of cellular extravasation, positive regulation of cell motility and cytokine-mediated signaling pathway, consistent with prior findings. At the pathway level, the significant up-regulation of the cytokine-cytokine receptor interaction pathway (Figure 5g) and the Jak-STAT signaling pathway (Figure 5h), coupled with the significant down-regulation of the ECM-receptor interaction pathway (Figure 5i) in aged versus fresh NPs, was also confirmed.
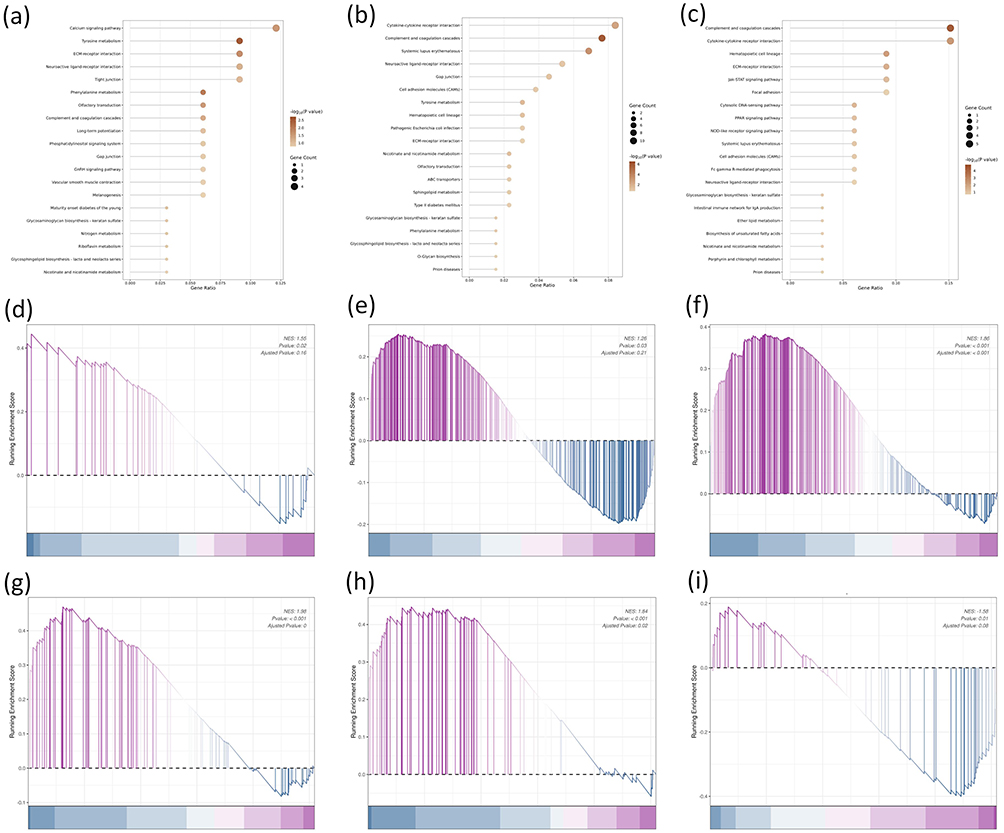 |
Figure 5 Integrated visualization of KEGG pathway enrichment and GSEA analysis. (a) Bubble plot showing the top 20 most enriched KEGG pathways in fresh NPs vs. control comparison. (b) Bubble plot showing the top 20 most enriched KEGG pathways in aged NPs vs. control comparison. (c) Bubble plot showing the top 20 most enriched KEGG pathways in aged NPs vs. fresh NPs comparison. All pathways are ranked by p-value. Dot size represents the number of genes mapped to the pathway, and color intensity reflects the enrichment significance (-log10(p-value)). (d–f) GSEA enrichment plots of aged vs fresh NPs groups comparison. (d–f) Validation of up-regulated biological processes: (d) cellular extravasation, (e) positive regulation of cell motility, (f) cytokine-mediated signaling pathway. (g–i) Validation of dysregulated KEGG pathways: (g) up-regulation of cytokine-cytokine receptor interaction, (h) up-regulation of Jak-STAT signaling pathway, (i) down-regulation of ECM-receptor interaction. In the ranked list, genes up-regulated in the aged group are positioned to the left. Statistical significance was defined as p < 0.05. |
Identifying TFs Associated with Chronic Induction of an Aggressive Phenotype by Aged ZnO NPs Through Integrated Network Analysis
Transcription factors act as master regulators of gene expression, orchestrating diverse cellular processes. Therefore, identifying the key TFs that drive transcriptional programs associated with enhanced aggressiveness is critical for elucidating the mechanisms behind ZnO NP-induced cancer progression. Using the CollecTRI network, we predicted 815 TFs potentially regulating the identified genes. To further investigate the mechanism by which ZnO NPs promote malignancy at the molecular level, we retrieved a set of 16119 increased malignancy-associated genes (encompassing proliferation, migration, and invasion) from the GeneCards database. Subsequently, we intersected these 815 TFs (predicted to regulate the transcriptomic profile) with the 16119 malignancy-associated genes. This analysis revealed a significant overlap of 779 TFs (Figure 6a). From this overlap, the top 100 nodes were selected to construct an increased malignancy associated TF-mRNA regulatory network (Figure 6b). To prioritize the most interconnected regulators, we then screened for genes with more than 50 edges, identifying 89 candidates. Finally, based on log2FoldChange ranking and a review of prior literature, we selected 9 pivotal genes for RT-qPCR validation, including 3 up-regulated genes (CEBPA, CTNNB1, STAT3) and 6 down-regulated genes (CEBPG, EGR1, NR3C1, SP1, STAT5A, TP53). Fortunately, expression of the 9 genes showed a consistent trend with that in RNA-seq in both fresh groups and aged groups (Figure 6c). Survival analysis of hepatocellular carcinoma patients revealed that high expression of CEBPA, CTNNB1, and STAT3 was statistically significantly associated with poor prognosis (Figure 6d–f), indicating that the elevated expression of CEBPA, CTNNB1, and STAT3 identified in our in vitro model is also associated with unfavorable clinical outcomes in HCC patients.
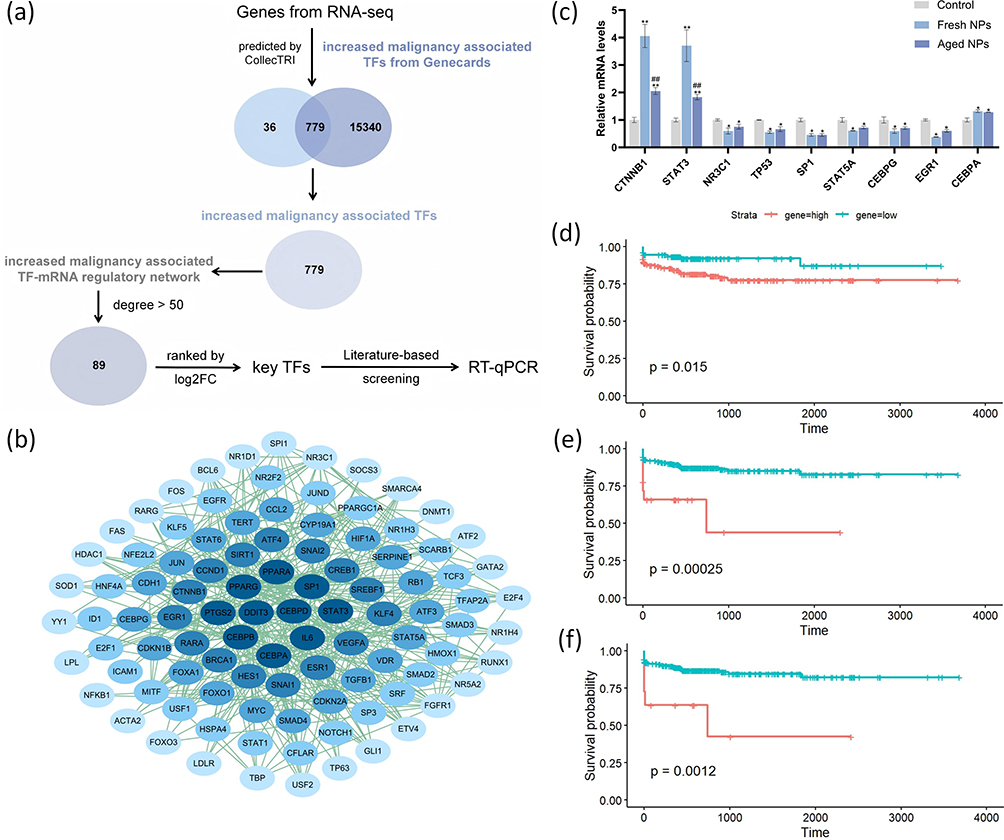 |
Figure 6 Identification of key transcription factors associated with aged ZnO NP-induced malignancy. (a) Schematic flowchart of the bioinformatic screening pipeline from the transcriptomic data derived from the comparison between aged NP-treated and control groups. (b) The TF-mRNA regulatory network visualizing the interactions among the top key TFs and their target genes. (c) RT-qPCR validation of the expression levels of nine pivotal TFs treated with fresh or aged NPs, compared to control. Data were based on 3 independent experiments and expressed as mean ± SD, *p < 0.05, **p < 0.01 vs control group, ##p < 0.01 for aged NPs versus fresh NPs groups. (d–f) Kaplan-Meier survival analysis of hepatocellular carcinoma patients from the TCGA database, showing that high expression of CEBPA (d), CTNNB1 (e) and STAT3 (f) is significantly associated with poor overall survival. |
Chronic Exposure to ZnO NPs Promotes the Migration and Invasion of HepG2 Cells
To further explore the impact of chronic exposure to ZnO NPs on the aggressive behaviors of HepG2 cells, we performed cell migration and invasion assays. The results showed that both fresh and aged NPs significantly increased the number of migrating and invading cells compared to control group. This effect was substantially stronger in aged NP group (Figure 7). In conclusion, these collective findings suggest that chronic exposure to low dose of ZnO NPs can significantly enhance the migratory and invasive capacities of HepG2 cells in vitro, and that aged NPs markedly promote this process.
 |
Figure 7 Chronic exposure to ZnO NPs promoted the migration and invasion capacity of HepG2 cells. (a) Representative images from the cell migration and invasion assays for control, fresh NPs, and aged NPs groups. (b and c) Quantitative analysis of cell migration (b) and invasion (c). Data are presented as the mean from three or more independent experiments (n = 3). *p < 0.05, **p < 0.01 vs control group. #p < 0.05 for aged NPs versus fresh NPs groups at the same concentration. |
Discussion
ZnO NPs have become one of the most widely utilized engineered nanomaterials in consumer products,43,44 leading to their inevitable release into the environment.45 Human exposure to ZnO NPs occurs through various pathways, including inhalation, ingestion, and dermal contact,46 raising significant public health concerns regarding their potential toxicity.47 Notably, upon environmental release, fresh ZnO NPs undergo an aging process, resulting in complex physicochemical transformations,48,49 such as the formation of hydrozincite (Zn5(CO3)2(OH)6) and increased dissolution into Zn2+ ions.50,51 Consequently, aged ZnO NPs, rather than their fresh counterparts, could be the primary form to which humans are exposed.52 The liver, as a major organ for xenobiotic metabolism and NPs accumulation, is a primary target for nanomaterial-induced toxicity.53,54 It is, therefore, critically important to evaluate the potential of these materials to enhance cancer cell aggressiveness in hepatic contexts,55 particularly those related to enhanced aggressive phenotype such as enhanced migration and invasion,56 induced by chronic exposure to low dose of aged ZnO NPs.57
Here, we employed a chronic exposure model combined with transcriptomic profiling in HepG2 cells, aiming to compare the effects of fresh versus aged ZnO NPs while acknowledging the potential confounding role of long-term selection pressures. To control for batch effects and isolate the NP-specific impact, all experimental groups (fresh or aged NPs groups and control groups) were cultured in parallel and passaged synchronously throughout the 16-week exposure period. Our analysis included identifying DEGs, conducting functional enrichment analyses (GO, KEGG, GSEA), and constructing a TF-mRNA regulatory network to identify key regulators. The clinical relevance of these findings was further assessed by correlating key TFs with patient survival data in hepatocellular carcinoma (HCC), and experimental validation was performed using RT-qPCR and Transwell assays. Collectively, our data indicate that chronic exposure to aged ZnO NPs is associated with a more aggressive phenotype in HCC. This phenotypic shift may result from the direct transcriptional changes induced by the NPs or the selective enrichment of a pre-existing aggressive subpopulation over the prolonged culture period—processes that our present study cannot fully disentangle but are important to acknowledge.58
Functional assays provided direct evidence supporting the enhanced aggressive capabilities induced by aged ZnO NPs. Transwell assays showed that chronic exposure to aged ZnO NPs significantly augmented the migratory and invasive capacities of HepG2 cells compared to both control and fresh ZnO NP-treated groups. This suggests that transformed NPs are more potent in promoting behaviors associated with enhanced aggressive phenotype. This finding is consistent with reports that certain nanomaterials can enhance aggressive behaviors. For instance, a study on platinum NPs revealed that they promote breast cancer metastasis by disrupting the vascular endothelial barrier through a mechanism involving ROS-induced damage to intercellular junctions.59 Our results contribute to this body of evidence by specifically highlighting that environmental aging is a crucial factor that can modulate the potential of ZnO NPs to promote an aggressive phenotype in HepG2 cells.
To decipher the molecular mechanisms underlying the differential effects of fresh and aged ZnO NPs, we conducted a comprehensive transcriptomic analysis. KEGG and GSEA revealed differences in pathway enrichment profiles between the two treatments. Fresh ZnO NPs appeared to primarily affect pathways related to basic cellular metabolism (eg., Tyrosine and Phenylalanine metabolism) and signal transduction (eg., Calcium signaling pathway). In comparison, aged ZnO NPs showed significant enrichment in pathways implicated in cancer progression, notably the cytokine-cytokine receptor interaction and the JAK-STAT signaling pathway, which have been reported as regulators of cell proliferation, migration, and invasion.60 GSEA validated the up-regulation of core processes like cellular extravasation, positive regulation of cell motility, and cytokine-mediated signaling pathway in aged NPs compared to fresh NPs. This coordinated activation of pro-migratory signaling networks provides an important mechanistic basis for the enhanced invasive phenotype we observed.61 GO term analysis provided supporting evidence, showing that aged NPs were associated with biological processes such as epithelial cell proliferation, positive regulation of cell motility, and extracellular matrix organization. This observed reshaping of the transcriptome induced by aged ZnO NPs, characterized in part by the activation of pro-oncogenic signals and the alteration of adhesion-related pathways, provides a plausible mechanistic explanation for their enhanced ability to promote cell migration and invasion observed in our phenotypic assays.
Transcription factors are DNA-binding proteins that regulate gene expression by controlling the initiation of transcription.62,63 They function as key molecular switches, and their dysregulation is frequently associated with disease pathogenesis.64 Given this critical role, identifying which specific TFs drive the transcriptional response to a stimulus is a central goal in mechanistic toxicology. To achieve this goal in our model, we constructed a TF-mRNA regulatory network by integrating our sequencing data with known malignancy-associated TFs. This analysis identified 89 highly interconnected TFs, from which we prioritized 9 pivotal TFs-CEBPA, CTNNB1, STAT3, CEBPG, EGR1, NR3C1, SP1, STAT5A, and TP53 -based on their high connectivity and significant differential expression. Experimental validation confirmed consistent expression trends for these TFs between RNA-seq and RT-qPCR data. To explore the potential clinical relevance of our in vitro findings, survival analysis using clinical data from HCC patients was performed. It revealed that elevated expression of CEBPA, CTNNB1, and STAT3 was significantly associated with poor patient prognosis. Although this correlation does not prove a direct causal link between NP exposure and clinical outcomes, it raises the possibility that the transcriptional signature (ie., co-upregulation of these three TFs) identified in our cellular model may reflect a molecular state pertinent to a more aggressive disease course in patients. This observation lends support to the potential pathophysiological relevance of the mechanisms proposed in this study. CTNNB1 (β-catenin), the central effector of the Wnt signaling pathway, drives enhanced aggressive phenotype by enhancing cell proliferation and migration when it accumulates in the nucleus.65 STAT3, a key signaling molecule, promotes cancer cell survival, proliferation, and immune evasion upon persistent activation.66 CEBPA, an important regulator of hepatocyte differentiation and metabolism, is a well-characterized tumor suppressor frequently inactivated in HCC.67 However, its role is highly context-dependent. The observed up-regulation of CEBPA in our model, particularly in concert with CTNNB1 and STAT3, suggests a potential pathological rewiring of its function under chronic stress. This suggests a potential functional alteration or dysregulation under chronic NP stress. This could involve a shift in its transcriptional program towards supporting stress adaptation and anabolic processes, or a change in its interaction with the broader oncogenic signaling network involving Wnt/β-catenin and STAT3 pathways.68 The concerted up-regulation of these factors by aged ZnO NPs suggests a synergistic oncogenic network: CEBPA may facilitate metabolic adaptation, CTNNB1 drives proliferative signaling, and STAT3 enhances cell survival and invasiveness. This network likely contributes to the observed aggressive phenotype. Our study provides the first evidence that under chronic exposure to aged ZnO NPs, the concerted up-regulation of CEBPA, CTNNB1, and STAT3 is critically associated with increased aggressiveness in HCC cells. This finding suggests that these three TFs may form a synergistic oncogenic network, potentially through metabolic reprogramming, enhanced proliferative signaling, and immune evasion, thereby playing a pivotal role in aged ZnO NP-induced aggravation of hepatocellular carcinoma malignancy. Further investigation into the specific roles of these TFs, particularly how aged ZnO NPs modulate their activity, will be crucial for understanding the detailed mechanism of action.
While this study provides novel insights into the chronic effects of environmentally aged ZnO NPs, several limitations should be acknowledged. First, the findings are based on a single in vitro hepatocellular carcinoma model (HepG2). Future studies employing additional cell lines (including non-cancerous hepatocytes) and in vivo models are necessary to assess the generalizability of these observations. Second, although our transcriptomic data strongly implicate pathways such as JAK-STAT and Wnt/β-catenin, the functional activation of these pathways and the specific role of identified transcription factors (eg., STAT3, CTNNB1) were not directly validated at the protein or functional level. Experiments utilizing pathway-specific inhibitors, siRNA knockdown, or luciferase reporter assays are warranted to establish causality. Third, the chronic exposure paradigm itself introduces interpretative complexity. The observed phenotypic shift could result from a combination of the differential gene expression directly induced by the NPs, adaptive cellular reprogramming, the selective pressure leading to clonal evolution, or the biological effects of sustained Zn2+ ion release.69 Our experimental design controlled for batch effects via parallel culture, but cannot definitively disentangle these intertwined possibilities. Future single-cell sequencing or lineage-tracing studies could help resolve the contribution of selection versus induction. Addressing these limitations in subsequent work will be crucial for a definitive mechanistic understanding.70
Conclusion
In summary, this study demonstrated that although aged ZnO NPs exhibit reduced acute cytotoxicity compared to fresh NPs, they are associated with a greater capacity to promote an aggressive phenotype in HepG2 cells following low-dose, long-term exposure. Transcriptomic and functional analyses suggest that this phenotypic shift may be linked to the modulation of pro-oncogenic pathways (eg., JAK-STAT and Wnt/β-catenin) and the differential expression of a key transcriptional network (CEBPA, CTNNB1, STAT3), a proposed mechanism summarized in Figure 8. The clinical correlation of this network with poor patient survival adds potential pathophysiological relevance to our in vitro observations. It is important to acknowledge the study’s limitations. The observed effects may stem from a combination of direct NPs action, sustained Zn2+ release, long-term adaptive cellular reprogramming, and/or clonal selection. Additionally, findings from the single HepG2 cell line model require further investigation for generalizability. Nevertheless, our work underscores that environmental aging is a critical factor modulating the long-term biological impact and potential health risks of ZnO NPs. It highlights the necessity of incorporating nanomaterial transformation into environmental and health risk assessments.
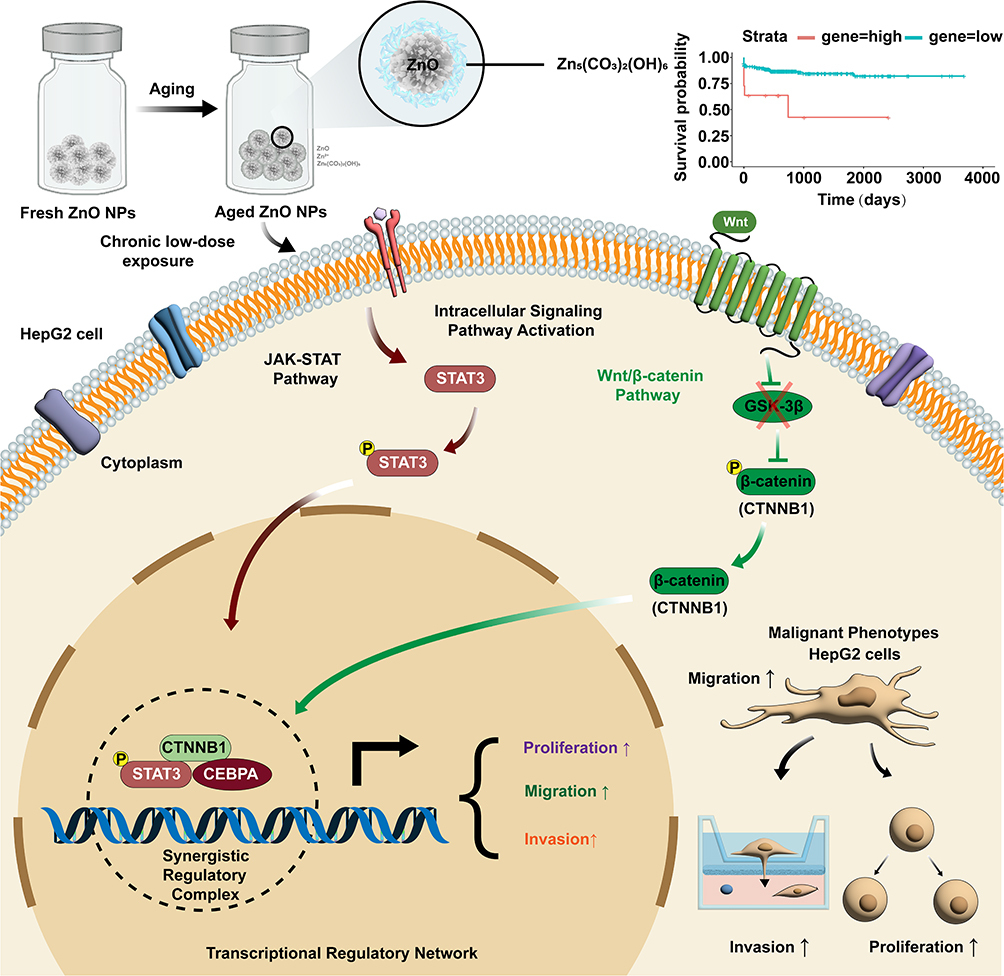 |
Figure 8 Schematic representation of the proposed mechanism for promoting malignant behaviors induced by aged ZnO NPs in HepG2 Cells. Arrows (↑) denote enhancement of the indicated cellular phenotypes and the cross symbol (×) indicates reduced phosphorylation of β-catenin in the pathway. |
Abbreviations
ZnO NPs, zinc oxide nanoparticles; TF, transcription factor; HCC, hepatocellular carcinoma; AST, aspartate aminotransferase; ALT, alanine aminotransferase; DEGs, differentially expressed genes; GSEA, Gene Set Enrichment Analysis; TEM, transmission electron microscopy; CCTCC, China Center for Type Culture Collection; DMEM, Dulbecco’s Modified Eagle’s Medium; FBS, fetal bovine serum; PCA, principal component analysis; GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; LIHC, liver hepatocellular carcinoma; TCGA, The Cancer Genome Atlas; RT-qPCR, Quantitative Real-Time PCR; SD, standard deviation; ANOVA, one-way analysis of variance; BP, biological process.
Data Sharing Statement
Data will be made available from the corresponding author on request.
Funding
This work was supported by grants from National Natural Science Foundation of China (22176002); Discipline (Professional) Leader Cultivation Project for Middle-aged and Young Teachers in Higher Education Institutions (DTR2025006); Funded Project of Anhui Medical University’s Research Level Improvement Program (2021xkjT004); Anhui Provincial undergraduate Innovation and Entrepreneurship Training Program (S202310366007 & S202310366110); Provincial Quality Project of Anhui Province in the New Era of Nurturing People (Graduate Education, 2024xscx061 & 2023xscx051); Anhui Medical University Postgraduate Research and Practice Innovation Program (YJS20240047); Early Exposure to Scientific Research Projects of Anhui Medical University (2023-ZQKY-119 & 2024-ZQKY-029).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Liu T, Lu Y, Zhan R, Qian W, Luo G. Nanomaterials and nanomaterials-based drug delivery to promote cutaneous wound healing. Adv Drug Deliv Rev. 2023;193:114670. doi:10.1016/j.addr.2022.114670
2. Liu M, Liu T, Chen X, et al. Nano-silver-incorporated biomimetic polydopamine coating on a thermoplastic polyurethane porous nanocomposite as an efficient antibacterial wound dressing. J Nanobiotechnol. 2018;16(1):89. doi:10.1186/s12951-018-0416-4
3. Mishra PK, Mishra H, Ekielski A, Talegaonkar S, Vaidya B. Zinc oxide nanoparticles: a promising nanomaterial for biomedical applications. Drug Discov Today. 2017;22(12):1825–17. doi:10.1016/j.drudis.2017.08.006
4. Efthimiou I, Kalamaras G, Papavasileiou K, et al. ZnO, Ag and ZnO-Ag nanoparticles exhibit differential modes of toxic and oxidative action in hemocytes of mussel Mytilus galloprovincialis. Sci Total Environ. 2021;767:144699. doi:10.1016/j.scitotenv.2020.144699
5. Singh TA, Sharma A, Tejwan N, Ghosh N, Das J, Sil PC. A state of the art review on the synthesis, antibacterial, antioxidant, antidiabetic and tissue regeneration activities of zinc oxide nanoparticles. Adv Colloid Interface Sci. 2021;295:102495. doi:10.1016/j.cis.2021.102495
6. Zhuo LB, Liu YM, Jiang Y, Yan Z. Zinc oxide nanoparticles induce acute lung injury via oxidative stress-mediated mitochondrial damage and NLRP3 inflammasome activation: in vitro and in vivo studies. Environ Pollut. 2024;341:122950. doi:10.1016/j.envpol.2023.122950
7. Qin X, Zhang J, Wang B, et al. Ferritinophagy is involved in the zinc oxide nanoparticles-induced ferroptosis of vascular endothelial cells. Autophagy. 2021;17(12):4266–4285. doi:10.1080/15548627.2021.1911016
8. Valdiglesias V, Alba-González A, Fernández-Bertólez N, et al. Effects of Zinc Oxide Nanoparticle Exposure on Human Glial Cells and Zebrafish Embryos. Int J Mol Sci. 2023;24(15):12297. doi:10.3390/ijms241512297
9. Zhang YN, Poon W, Tavares AJ, McGilvray ID, Chan WCW. Nanoparticle-liver interactions: cellular uptake and hepatobiliary elimination. J Control Release. 2016;240:332–348. doi:10.1016/j.jconrel.2016.01.020
10. Poon W, Zhang YN, Ouyang B, et al. Elimination Pathways of Nanoparticles. ACS Nano. 2019;13(5):5785–5798. doi:10.1021/acsnano.9b01383
11. JM Llovet, RK Kelley, A Villanueva, et al. Hepatocellular carcinoma. Nat Rev Dis Primers. 2021;7(1):7. doi:10.1038/s41572-021-00245-6
12. Zhan Q, Liu B, Situ X, et al. New insights into the correlations between circulating tumor cells and target organ metastasis. Signal Transduct Target Ther. 2023;8(1):465. doi:10.1038/s41392-023-01725-9
13. Chan SL, Sun HC, Xu Y, et al. The Lancet Commission on addressing the global hepatocellular carcinoma burden: comprehensive strategies from prevention to treatment. Lancet. 2025;406(10504):731–778. doi:10.1016/s0140-6736(25)01042-6
14. Stanley LA, Wolf CR. Through a glass, darkly? HepaRG and HepG2 cells as models of human Phase I drug metabolism. Drug Metab Rev. 2022;54(1):46–62. doi:10.1080/03602532.2022.2039688
15. Lin Z, Monteiro-Riviere NA, Riviere JE. Pharmacokinetics of metallic nanoparticles. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2015;7(2):189–217. doi:10.1002/wnan.1304
16. El-Kady AM, Sh A, Mohamed K, et al. Zinc oxide nanoparticles produced by Zingiber officinale ameliorates acute toxoplasmosis-induced pathological and biochemical alterations and reduced parasite burden in mice model. PLoS Negl Trop Dis. 2023;17(7):e0011447. doi:10.1371/journal.pntd.0011447
17. S S, Ss A, Sb S, Hk V, Pv M. Determination of the bioavailability of zinc oxide nanoparticles using ICP-AES and associated toxicity. Colloids Surf B Biointerfaces. 2020;188:110767. doi:10.1016/j.colsurfb.2019.110767
18. He M, Li X, Yu L, et al. Double-Sided Nano-ZnO: superior Antibacterial Properties and Induced Hepatotoxicity in Zebrafish Embryos. Toxics. 2022;10(3):144. doi:10.3390/toxics10030144
19. Fujihara J, Nishimoto N. Review of Zinc Oxide Nanoparticles: toxicokinetics, Tissue Distribution for Various Exposure Routes, Toxicological Effects, Toxicity Mechanism in Mammals, and an Approach for Toxicity Reduction. Biol Trace Elem Res. 2024;202(1):9–23. doi:10.1007/s12011-023-03644-w
20. Hong H, Liu Z, Li S, et al. Zinc oxide nanoparticles (ZnO-NPs) exhibit immune toxicity to crucian carp (Carassius carassius) by neutrophil extracellular traps (NETs) release and oxidative stress. Fish Shellfish Immunol. 2022;129:22–29. doi:10.1016/j.fsi.2022.07.025
21. Almansour MI, Alferah MA, Shraideh ZA, Jarrar BM. Zinc oxide nanoparticles hepatotoxicity: histological and histochemical study. Environ Toxicol Pharmacol. 2017;51:124–130. doi:10.1016/j.etap.2017.02.015
22. Karwadiya J, Lützenkirchen J, Darbha GK. Retention of ZnO nanoparticles onto polypropylene and polystyrene microplastics: aging-associated interactions and the role of aqueous chemistry. Environ Pollut. 2024;352:124097. doi:10.1016/j.envpol.2024.124097
23. Gomez-Gonzalez MA, Da Silva-Ferreira T, Clark N, Clough R, Quinn PD, Parker JE. Toward Understanding the Environmental Risks of Combined Microplastics/Nanomaterials Exposures: unveiling ZnO Transformations after Adsorption onto Polystyrene Microplastics in Environmental Solutions. Glob Chall. 2023;7(8):2300036. doi:10.1002/gch2.202300036
24. Keller AA, Wang H, Zhou D, et al. Stability and aggregation of metal oxide nanoparticles in natural aqueous matrices. Environ Sci Technol. 2010;44(6):1962–1967. doi:10.1021/es902987d
25. Yan Z, Liu C, Liu Y, et al. The interaction of ZnO nanoparticles, Cr(VI), and microorganisms triggers a novel ROS scavenging strategy to inhibit microbial Cr(VI) reduction. J Hazard Mater. 2023;443(Pt B):130375. doi:10.1016/j.jhazmat.2022.130375
26. Lai RWS, Zhou GJ, Yung MMN, Djurišić AB, Leung KMY. Interactive effects of temperature and salinity on toxicity of zinc oxide nanoparticles towards the marine mussel Xenostrobus securis. Sci Total Environ. 2023;889:164254. doi:10.1016/j.scitotenv.2023.164254
27. Eixenberger JE, Anders CB, Hermann RJ, et al. Rapid Dissolution of ZnO Nanoparticles Induced by Biological Buffers Significantly Impacts Cytotoxicity. Chem Res Toxicol. 2017;30(8):1641–1651. doi:10.1021/acs.chemrestox.7b00136
28. Jones P, Sugino S, Yamamura S, Lacy F, Biju V. Impairments of cells and genomic DNA by environmentally transformed engineered nanomaterials. Nanoscale. 2013;5(20):9511–9516. doi:10.1039/c3nr03118g
29. Zhang H, Chen Z, Huang Q. Study of the toxicity of ZnO nanoparticles to Chlorella sorokiniana under the influence of phosphate: spectroscopic quantification, photosynthetic efficiency and gene expression analysis. Environ Sci Nano. 2020;7(5):1431–1443. doi:10.1039/C9EN01464K
30. George S, Yin H, Liu Z, Shen S, Cole I, Khiong CW. Hazard profiling of a combinatorial library of zinc oxide nanoparticles: ameliorating light and dark toxicity through surface passivation. J Hazard Mater. 2022;434:128825. doi:10.1016/j.jhazmat.2022.128825
31. Yang Q, Xu L, Yang X, et al. Fate and enzymatic response of co-exposed photoaged nanoplastic and PFAS: insights from a human gastrointestinal simulation. J Hazard Mater. 2025;496:139274. doi:10.1016/j.jhazmat.2025.139274
32. Amde M, Liu JF, Tan ZQ, Bekana D. Transformation and bioavailability of metal oxide nanoparticles in aquatic and terrestrial environments. A review. Environ Pollut. 2017;230:250–267. doi:10.1016/j.envpol.2017.06.064
33. Abbas Q, Yousaf B, Amina, et al. Transformation pathways and fate of engineered nanoparticles (ENPs) in distinct interactive environmental compartments: a review. Environ Int. 2020;138:105646. doi:10.1016/j.envint.2020.105646
34. Irumva O, Twagirayezu G, Xia A, et al. Environmental fate, transport, impacts, and future perspectives of engineered nanoparticles in surface waters. Environ Res. 2025;285(Pt 1):122267. doi:10.1016/j.envres.2025.122267
35. Wang MM, Wang YC, Wang XN, et al. Mutagenicity of ZnO nanoparticles in mammalian cells: role of physicochemical transformations under the aging process. Nanotoxicology. 2015;9(8):972–982. doi:10.3109/17435390.2014.992816
36. Lee MKH, Lim HK, Su C, et al. 3D Airway Epithelial-Fibroblast Biomimetic Microfluidic Platform to Unravel Engineered Nanoparticle-Induced Acute Stress Responses as Exposome Determinants. Environ Sci Technol. 2023;57(48):19223–19235. doi:10.1021/acs.est.3c03678
37. Hernández-Moreno D, Valdehita A, Conde E, Rucandio I, Navas JM, Fernández-Cruz ML. Acute toxic effects caused by the co-exposure of nanoparticles of ZnO and Cu in rainbow trout. Sci Total Environ. 2019;687:24–33. doi:10.1016/j.scitotenv.2019.06.084
38. Raha S, Ahmaruzzaman M. ZnO nanostructured materials and their potential applications: progress, challenges and perspectives. Nanoscale Adv. 2022;4(8):1868–1925. doi:10.1039/d1na00880c
39. Al-Darwesh MY, Manai M, Chebbi H, Klein A. Green Synthesis of Chitosan-Coated Selenium Nanoparticles for Paclitaxel Delivery. Nanomaterials. 2025;15(16):1276. doi:10.3390/nano15161276
40. Al-darwesh MY, Manai M, Boughzala N, Chebbi H. Green synthesis and characterization of platinum nanoparticles loaded with paclitaxel and coated with chitosan: a promising approach for breast cancer treatment. Mater Chem Phys. 2026;354:132192. doi:10.1016/j.matchemphys.2026.132192
41. Al-darwesh MY, Akool A, Khalaf K, Faisal S, Ahmed TBZ. Novel Enhanced Delivery of Prednisolone via Chitosan-Coated Zinc Oxide Nanoparticles: characterization and Evaluation of Its Anti-inflammatory, Antibacterial, and Antioxidant Potential. BioNanoScience. 2025;15(3):452. doi:10.1007/s12668-025-02071-6
42. Wang MM, Cao R, Jiang WG, et al. Long-term exposure to low doses of fresh and aged zinc oxide nanoparticles causes cell malignant progression enhanced by a tyrosine phosphatase SHP2 gain-of-function mutation. Environ Sci Nano. 2019;6(8):2389–2404. doi:10.1039/C8EN01191E
43. El-Saadony MT, Fang G, Yan S, et al. Green Synthesis of Zinc Oxide Nanoparticles: preparation, Characterization, and Biomedical Applications - A Review. Int J Nanomed. 2024;19:12889–12937. doi:10.2147/ijn.S487188
44. Pal P, Pareek A. Zinc oxide nanoparticles: a comprehensive review on its synthesis, characterization, and role in biomedical applications as well as health risks. Inorg Chem Commun. 2025;181:115314. doi:10.1016/j.inoche.2025.115314
45. Sun H, Guo W, Zhou Q, et al. Uptake, transformation, and environmental impact of zinc oxide nanoparticles in a soil-wheat system. Sci Total Environ. 2023;857(Pt 1):159307. doi:10.1016/j.scitotenv.2022.159307
46. Youn SM, Choi SJ. Food Additive Zinc Oxide Nanoparticles: dissolution, Interaction, Fate, Cytotoxicity, and Oral Toxicity. Int J Mol Sci. 2022;23(11):6074. doi:10.3390/ijms23116074
47. Lee HI, Kwon RY, Choi SJ. Food Additive Solvents Increase the Dispersion, Solubility, and Cytotoxicity of ZnO Nanoparticles. Nanomaterials. 2023;13(18):2573. doi:10.3390/nano13182573
48. Feng X, Yan Y, Wan B, et al. Enhanced Dissolution and Transformation of ZnO Nanoparticles: the Role of Inositol Hexakisphosphate. Environ Sci Technol. 2016;50(11):5651–5660. doi:10.1021/acs.est.6b00268
49. Bathi JR, Moazeni F, Upadhyayula VKK, et al. Behavior of engineered nanoparticles in aquatic environmental samples: current status and challenges. Sci Total Environ. 2021;793:148560. doi:10.1016/j.scitotenv.2021.148560
50. Zhang H, Huang Q, Xu A, Wu L. Spectroscopic probe to contribution of physicochemical transformations in the toxicity of aged ZnO NPs to Chlorella vulgaris: new insight into the variation of toxicity of ZnO NPs under aging process. Nanotoxicology. 2016;10(8):1177–1187. doi:10.1080/17435390.2016.1196252
51. Spurgeon DJ, Lahive E, Schultz CL. Nanomaterial Transformations in the Environment: effects of Changing Exposure Forms on Bioaccumulation and Toxicity. Small. 2020;16(36):e2000618. doi:10.1002/smll.202000618
52. Fang X, Jiang L, Gong Y, Li J, Liu L, Cao Y. The presence of oleate stabilized ZnO nanoparticles (NPs) and reduced the toxicity of aged NPs to Caco-2 and HepG2 cells. Chem Biol Interact. 2017;278:40–47. doi:10.1016/j.cbi.2017.10.002
53. Sun T, Kang Y, Liu J, et al. Nanomaterials and hepatic disease: toxicokinetics, disease types, intrinsic mechanisms, liver susceptibility, and influencing factors. J Nanobiotechnol. 2021;19(1):108. doi:10.1186/s12951-021-00843-2
54. Xu J, Liu Y. Nanomaterials for liver cancer targeting: research progress and future prospects. Front Immunol. 2025;16:1496498. doi:10.3389/fimmu.2025.1496498
55. Yan D, Long J, Liu J, Cao Y. The toxicity of ZnO nanomaterials to HepG2 cells: the influence of size and shape of particles. J Appl Toxicol. 2019;39(2):231–240. doi:10.1002/jat.3712
56. Kitchin KT, Richards JA, Robinette BL, et al. Biochemical effects of copper nanomaterials in human hepatocellular carcinoma (HepG2) cells. Cell Biol Toxicol. 2023;39(5):2311–2329. doi:10.1007/s10565-022-09720-6
57. Sutunkova MP, Ryabova YV, Minigalieva IA, et al. Features of the response to subchronic low-dose exposure to copper oxide nanoparticles in rats. Sci Rep. 2023;13(1):11890. doi:10.1038/s41598-023-38976-z
58. Bolton KL, Ptashkin RN, Gao T, et al. Cancer therapy shapes the fitness landscape of clonal hematopoiesis. Nat Genet. 2020;52(11):1219–1226. doi:10.1038/s41588-020-00710-0
59. Wang D-P, Shen J, Qin C-Y, et al. Platinum nanoparticles promote breast cancer cell metastasis by disrupting endothelial barrier and inducing intravasation and extravasation. Nano Res. 2022;15(8):7366–7377. doi:10.1007/s12274-022-4404-5
60. Kroemer G, Pouyssegur J. Tumor cell metabolism: cancer’s Achilles’ heel. Cancer Cell. 2008;13(6):472–482. doi:10.1016/j.ccr.2008.05.005
61. Jiang Y, Zhang H, Wang J, Liu Y, Luo T, Hua H. Targeting extracellular matrix stiffness and mechanotransducers to improve cancer therapy. J Hematol Oncol. 2022;15(1):34. doi:10.1186/s13045-022-01252-0
62. Spitz F, Furlong EE. Transcription factors: from enhancer binding to developmental control. Nat Rev Genet. 2012;13(9):613–626. doi:10.1038/nrg3207
63. Kumar A, Chan J, Taguchi M, Kono H. Interplay among transacting factors around promoter in the initial phases of transcription. Curr Opin Struct Biol. 2021;71:7–15. doi:10.1016/j.sbi.2021.04.008
64. Hasan A, Khan NA, Uddin S, Khan AQ, Steinhoff M. Deregulated transcription factors in the emerging cancer hallmarks. Semin Cancer Biol. 2024;98:31–50. doi:10.1016/j.semcancer.2023.12.001
65. Calderaro J, Ziol M, Paradis V, Zucman-Rossi J. Molecular and histological correlations in liver cancer. J Hepatol. 2019;71(3):616–630. doi:10.1016/j.jhep.2019.06.001
66. He G, Karin M. NF-κB and STAT3 - key players in liver inflammation and cancer. Cell Res. 2011;21(1):159–168. doi:10.1038/cr.2010.183
67. Tan EH, Hooi SC, Laban M, et al. CCAAT/enhancer binding protein alpha knock-in mice exhibit early liver glycogen storage and reduced susceptibility to hepatocellular carcinoma. Cancer Res. 2005;65(22):10330–10337. doi:10.1158/0008-5472.Can-04-4486
68. Lourenço AR, Coffer PJ. A tumor suppressor role for C/EBPα in solid tumors: more than fat and blood. Oncogene. 2017;36(37):5221–5230. doi:10.1038/onc.2017.151
69. Liu J, Feng X, Wei L, Chen L, Song B, Shao L. The toxicology of ion-shedding zinc oxide nanoparticles. Crit Rev Toxicol. 2016;46(4):348–384. doi:10.3109/10408444.2015.1137864
70. Jovic D, Liang X, Zeng H, Lin L, Xu F, Luo Y. Single-cell RNA sequencing technologies and applications: a brief overview. Clin Transl Med. 2022;12(3):e694. doi:10.1002/ctm2.694
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.