Back to Journals » International Journal of Nanomedicine » Volume 21
Research Progress on Mesenchymal Stem Cells-Derived Small Size Vesicles for the Treatment of Liver Diseases
Authors Meng C, Liu M, Qi Q, Cao L, Lei Q, Liu X, Wang M
Received 5 February 2026
Accepted for publication 6 May 2026
Published 20 May 2026 Volume 2026:21 601322
DOI https://doi.org/10.2147/IJN.S601322
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sachin Mali
Chuyang Meng,1,* Min Liu,2,* Qianqian Qi,1 Lixue Cao,1 Qi Lei,1 Xifu Liu,1 Meng Wang1
1Ministry of Education Key Laboratory of Molecular and Cellular Biology; Hebei Research Center of the Basic Discipline of Cell Biology; Hebei Provincial Technology Innovation Center for Anti-Tumor Molecular Targeting New Drugs Development; College of Life Science, Hebei Normal University, Shijiazhuang, 050024, People’s Republic of China; 2Department of Physiology, College of Life Science, Hebei Normal University, Shijiazhuang, 050024, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Meng Wang, Ministry of Education Key Laboratory of Molecular and Cellular Biology; Hebei Research Center of the Basic Discipline of Cell Biology; Hebei Provincial Technology Innovation Center for Anti-tumor Molecular Targeting New Drugs Development; College of Life Science, Hebei Normal University, Shijiazhuang, 050024, People’s Republic of China, Tel/Fax +86 311 80786051, Email [email protected]
Abstract: Liver disease is a major public health challenge, accounting for a substantial burden of mortality and morbidity worldwide. Currently, liver transplantation is the gold standard for treating end-stage liver diseases. However, it is affected by the shortage of organ sources and immune rejection. Mesenchymal stem cells (MSCs) can be used as an alternative therapy for liver diseases, especially in cases of liver cirrhosis, liver failure and complications of liver transplantation. However, MSCs may have potential effects on tumor occurrence. MSCs derived small size vesicles are nanoscale extracellular vesicles (EVs) which secreted by MSCs. MSCs derived small size vesicles are characterized by a lipid bilayer membrane, which encapsulate and transport various biomolecules such as proteins and miRNAs. Compared to MSCs, MSCs derived small size vesicles showed more advantages including lower tumor occurrence and lower toxicity. It has been found that MSCs derived small size vesicles ameliorate liver diseases via regulating lipid metabolism, inhibiting apoptosis, promoting regeneration, suppressing inflammation, drug delivery and so on. Based on the important role of MSCs derived small size vesicles in liver diseases treatment, this review aims to summarize the biological processes of small size vesicles with focusing on the effect of MSCs derived small size vesicles on different liver diseases. Additionally, we also discussed the potential future directions for small size vesicles research, including the challenges to be overcome and the prospects for clinical applications. This review provides theoretical support of MSCs derived small size vesicles as a potential therapeutic agent for liver disease in humans. The diagram illustrates mesenchymal stem cells-derived small size vesicles and their sources, including bone marrow, adipose tissue, Wharton’s jelly, umbilical cord, embryo and liver. Inside the vesicle, components like tetraspanins (CD9, CD63, CD81), DNA, mRNA, microRNA, enzymes and EGFR are shown. The right side depicts the treatment effects on various liver diseases: liver injury, ALD/NAFLD, liver fibrosis, liver cirrhosis, liver failure and HCC. Each liver disease is associated with specific therapeutic mechanism, such as promoting hepatocyte proliferation, anti-inflammatory effects, inhibiting fibrosis and promoting liver tissue repair.Diagram of mesenchymal stem cells-derived small size vesicles and their treatment effects on liver diseases.
Keywords: liver disease, mesenchymal stem cells, MSCs, small size vesicles
Introduction
Various cells can release extracellular vesicles (EVs), which is bounded by a lipid bilayer and cannot replicate by themselves.1 According to the characteristics including the size, density, composition and so on, EVs can be divided into different subtypes, such as microvesicles and apoptotic bodies.2 As a subtype of EVs, small size vesicles usually origin from the endosomal system and are released via the multivesicular bodies (MVBs).3 So far now, small size vesicles have been found to be widely present in the vast majority of body fluids and tissues.4 Small size vesicles are approximately smaller than 200 nm in size and generally considered to be vesicles containing proteins, nucleic acids, and lipids.5 The proteins in small size vesicles exhibit excellent biological activities in physiological processes.6 There are many types of proteins on small size vesicles, such as metabolic enzymes phosphatolycerate kinase 1, NADP dependent male enzymes, heat shock proteins, cytoskeleton, secret proteins, etc.7 Several proteins in small size vesicles serve as biomarkers, such as CD9, CD63, lactadherin, glycosyl-phosphatidyl-inositol and so on, which may explain the targeting and selective characteristics of small size vesicles that entry into the target cells.6 In 2007, Valadi et al demonstrated that there were mRNA and miRNA in small size vesicles originated from the mast cells.8 In addition, it was also found that there were non-coding RNAs and mtDNAs presented in small size vesicles.9 Though the definition of small size vesicles is being updating in real-time, it is been widely recognized that small size vesicles and the contents of small size vesicles participate in several physiological and pathological biological processes.10 Additionally, the miRNA of small size vesicles can be absorbed by neighboring cells, which then may affect the target cells bioactivities.11 The miRNA of small size vesicles can perform negative regulation and cause changes in the expression levels of target genes.12 For example, the exosomes miR-92a-derived from K562 cells can reduce the expression of integrin α5 in human umbilical vein endothelial cells (HUVEC) and enhance endothelial cell migration.13 It has been reported that non-coding RNA from mesenchymal stem cells-derived exosomes (MSCs-exos) has great potential in the treatment of wound healing.14,15 For example, miR-135a in the human amniotic mesenchymal stem cell-derived exosomes can promote fibroblast proliferation and migration.16 The lncRNA KLF3-AS1 on bone marrow mesenchymal stem cell-derived exosomes (BMSC-exos) also can accelerate wound healing by upregulating vascular endothelial growth factor A.17
As a digestive organ, liver serves as a hub for several chemical and physiological reactions that occur in the body, such as the nutrients metabolism, blood volume and endocrine regulation, the synthesis and decomposition of lipids and cholesterol.18 As is well known, liver is composed of hepatocytes, cholangiocytes, hepatic stellate cells (HSCs), Kupffer cells, and liver sinusoidal endothelial cells (LSECs).18 These cells work together to regulate liver function at multiple levels and dimensions.19 Hepatocytes are the most common cells in the liver, and the regeneration of the liver is tightly associated with the proliferation of hepatocytes.20 Regulating the hepatocytes cell cycle can effectively inhibit the senescence of hepatocytes and stimulate their proliferation.19 Cholangiocytes are highly specialized cells and can collaborate with hepatocytes to achieve liver regeneration.21 When one of the two kind cells fail to regenerate, hepatocytes or cholangiocytes can function as facultative stem cells and switch to each other for liver regeneration.22 Different from hepatocytes and cholangiocytes, HSCs have four phenotypes: quiescent, activated, inactivated, and senescent.23 Activated HSCs cause liver damage and induce liver fibrosis.24 Kupffer cells are liver-derived macrophages that are commonly found in the hepatic sinusoids.25 Kupffer cells can resist bacteria and endotoxins, playing an important role in immunity.26 LSECs are well-known for their ability to form the hepatic sinusoidal wall. LSECs are well-known for their ability to form the hepatic sinusoidal wall.27 LSECs usually form an open fenestrae and lack of an organized basement membrane, which play a pivotal role in maintaining hepatic homeostasis.28
Liver disease is a relatively broad concept, specifically including infections of hepatitis B virus (HBV) and hepatitis C virus (HCV), alcoholic liver disease (ALD), non-alcoholic fatty liver disease (NAFLD), and end-stage liver diseases such as liver cirrhosis, liver failure, and hepatocellular carcinoma (HCC).29 Generally speaking, if early-stage liver disease patients have not received systematic treatment, all types of chronic hepatitis may develop into cirrhosis, chronic liver failure, and HCC.30 It has been reported that approximately 2 million people die each year due to liver disease, with the vast majority of deaths caused by cirrhosis and liver cancer.31 Many risk factors can trigger liver disease, including alcohol, high fat diet and so on.29 Meanwhile, some other organs and diseases are also tightly associated with the occurrence of liver disease.32,33 For examples, diabetes has been demonstrated to be closely related with liver disease34 and heart disease may cause acute liver damage, congestive liver disease, and chronic liver disease.32 Several chemical drugs have been demonstrated the positive effect on the treatment of liver disease, for example, lamivudine improves the prognosis of patients with chronic HBV.35 Resveratrol and silymarin can attenuate the development of NAFLD.36,37
This article will detailly introduce the biological processes of exosomes in body and summarizes the research progress of MSCs derived small size vesicles for the treatment of liver diseases.
Biological Processes of Small Size Vesicles
The Biogenesis of Small Size Vesicles
The biogenesis of small size vesicles is related to cell endocytosis. The generation of small size vesicles usually concludes budding, invagination, MVBs formation, and secretion.38 When the plasma membrane buds and enters into the cell, an initial vesicle, which is also called early endosome, is formed. The early endosome is responsible for the initial sorting of internalized cargo.39 The fate of early endosomes is different. Some early endosomes will be recycled by being fused with the Golgi apparatus network or plasma membrane, while some early endosomes are destined to mature into late endosomes.40 During the maturation process of the endosomes, the endosomes membrane is also undergoing changes. A significant feature is the alteration of the membrane composition. For example, sphingolipids in membrane are replaced by ceramides, and Rab5 is substituted by Rab7.41 These changes are necessary for late endosome sorting and play an important role in the transportation of late endosomes. Following with the endosome mature, regions of the endosomal membrane invaginate into the intraluminal space and form multiple intraluminal vesicles (ILVs), which are the precursors of small size vesicles. ILVs transform the compartment into MVBs, however, some MVBs fuse with lysosomes, where the ILVs are degraded. Others MVBs fuse with the plasma membrane and release the ILVs into the extracellular space, which are known as small size vesicles (Figure 1).42,43
 |
Figure 1 The biogenesis of small size vesicles. The biogenesis of small size vesicles begins with the plasma membrane invaginating to form early endosomes. Part of early endosomes recycle their cargo by fusing with the Golgi apparatus network or the plasma membrane. At the same time, others early endosomes mature into late endosomes. Ultimately, some late endosomes are degraded by fusion with lysosomes, while others release small size vesicles fuse with the plasma membrane and then release small size vesicles. After the release of small size vesicles, small size vesicles directly fuse with the target cell membrane and release the contents. Small size vesicles also enter target cells through endocytosis and direct interaction with target cells surface receptors. |
Due for the important role of ILVs on the formation of exosomes, the mechanisms for regulating the formation of ILVs attracts more and more attention. So far now, they are mainly summarized into two main categories: those that rely on endosomal sorting complex required for transport (ESCRT) and those that do not rely on ESCRT.40 ESCRT can be divided into ESCRT-0, I, II and III, which is essentially composed of class E vacuolar protein sorting (Vps). The main function of ESCRT-0, ESCRT-I, and ESCRT-II is to sort the cargos that need to be wrapped into functional microstructural domains on the inner membrane to promote signal transduction. Different from them, ESCRT-III induces budding and scission of the microstructural domains, leading to the production of ILVs.44 Notably, some auxiliary proteins participate in the formation process of ESCRT, such as multi-vesicular body of 12 kDa, tumor susceptibility gene 101 (TSG101) and so on.44,45 Tony et al found that multi-vesicular body of 12 kDa is a component of ESCRT-I complex, which binds to the coiled helical domain of Vps23 to ensure the ordered assembly of ESCRT-I and II.46 Strickland et al reported that TSG101 is a component of ESCRT-I and mainly responsible for the recognition of monoubiquitin.47
Exclude the classical ESCRTs take part in the ILVs generation, there are other mechanisms involved in the process by an ESCRT-independent manner.39 In mammalian cells, ILVs form within the MVBs lumen when ESCRT is depleted, indicating the existence of an ESCRT-independent pathway to form ILVs.48 As is reported, the generation of ceramides, phosphatidic acids, or lipid rafts can facilitate the non-dependent ESCRT pathways.49 For example, ceramides, which have structural and physical properties that promote vesicle biogenesis, works together with membrane sphingolipids to promote the lipid raft microdomains formation and then trigger the spontaneous negative curvature of the lipid raft microdomains membrane to initiate the ILVs generation.41,42
The Release of Small Size Vesicles
It is well known that MVBs can be transported to lysosomes or the plasma membrane. Factors involved in MVBs transport, such as cortactin, Rab proteins, ISGylation, and soluble N-ethylmaleimide-sensitive factor attachment protein receptors, are attracting increasing attention.
The interaction between MVBs and actin with microtubule cytoskeleton has been found to be crucial for the MVBs transport to the plasma membrane.50 Cortactin, a protein that presents in all actin-based structures, participates in the processes including endocytosis, vesicle transport, adhesion, migration, and invasion.51 Known cortactin has been shown to reduce the release of small size vesicles. In addition, Sinha et al reported that cortactin facilitates the transport and docking of MVBs with the plasma membrane, playing a pivotal role in the release of EVs.52
Rab proteins are a key family of small GTPases. As the key regulators of eukaryotic membrane trafficking, Rab proteins play critical roles in vesicle fission and fusion. Parray reported that Rab proteins take part in nearly every stage phase of membrane trafficking, including the formation of transport vesicles, the movement of vesicles and the docking of vesicles with their target compartments.53 It has been reported that silencing Rab27a increases the size of MVBs, while silencing Rab27b leads to regional redistribution of MVBs.54 Hsu et al confirmed that inhibiting Rab35 leads to the accumulation of endosomal vesicles and reduces the secretion of exosomes.55 Additionally, other members of the Rab family such as Rab11and Rab35 are also demonstrated the critical role in regulating the release of small size vesicles.50,56
Meanwhile, ISGylation, a post-translational ubiquitin like modification, was found to be associated with the small size vesicles release. It has been found that the induction of ISGylation promotes the fusion of MVBs with lysosomes, leading to the degradation of MVBs and preventing their transport to the plasma membrane.50,57
After MVBs are transported to the plasma membrane, some molecules, such as soluble N-ethylmaleimide-sensitive factor attachment protein receptor, promote the fusion of vesicles with plasma membrane or organelle membrane.44 With the help of microtubules, Rabs, and tethering factors, one vesicle soluble N-ethylmaleimide-sensitive factor attachment protein receptor and two target soluble N-ethylmaleimide-sensitive factor attachment protein receptors form four-helix bundles to drive the fusion between MVBs and plasma membranes and lead to the small size vesicles secretion.39
The Uptake of Small Size Vesicles
When small size vesicles directly fuse with the target cell membrane, their contents are released into the cytoplasm, completing their uptake process. Although this direct fusion pathway is thought to be one of the most effective means of cargo delivery into target cells, other mechanisms also exist for small size vesicles uptake, such as endocytosis and direct interaction with target cells surface receptors (Figure 1).6,40 Endocytosis is considered to be the main way for cells to take in small size vesicles.1 Small size vesicles undergo uncoating and subsequently fuse with plasma membrane to enter the cells.58 The contents in exosomes need to be released from the vesicles and enter the cytoplasm to exert their functions. Otherwise, they are degraded by lysosomes, recycled within the cell or secreted into the extracellular space.59 Due to the poor predictability of this process among different cell types, there are still many unknown aspects regarding the mechanism by which small size vesicles release their contents into the cytoplasm.1 A proposed mechanism is that small size vesicles fuse with endosomes in a pH-dependent manner, making the plasma membrane permeable and allowing the contents in small size vesicles to reach the cytoplasm.60
Physiological Roles of Exosomes
Garbage Carrier
The formation of small size vesicles originates from endocytosis within cells and carries biological information from the parent cells.1 Due to the characteristics of small size vesicles formation, small size vesicles are regarded as garbage disposers which eliminate unnecessary cellular components, including redundant or misfolded proteins, molecules and metabolic wastes.61 Surya Shrivastava and Kevin V Morris found that overexpressed RNA and DNA fragments were also collected in small size vesicles and expelled outside the cells, which maintained cellular homeostasis.62 These studies indicates that small size vesicles can also be regarded as the garbage carrier in the cells.
Cell Communication
Small size vesicles trigger signal transduction mainly through two primary mechanisms, one is directly interacting with extracellular receptors, the other is internalized by receptor cells and then released cargo.40 For directly interaction, the transmembrane ligands on the surface of small size vesicles can directly bind to surface receptors on target cells for signal transduction. For example, human DEX activate T cells through MHC peptide complexes.63 Meanwhile, small size vesicles can also be internalized via various pathways, including clathrin-mediated endocytosis, lipid raft mediated endocytosis, caveolin mediated endocytosis, phagocytosis and macropinocytosis. These processes are not mutually exclusive and can operate concurrently.40 It has been reported that rat bone marrow MSCs internalize PC12-derived exosomes through clathrin-mediated endocytosis and micropinocytosis, which transfers miR-21 from exosomes to bone marrow MSCs and reduces the expression of TGFβRII and TPM1 in bone marrow MSCs.64
Cell communication is tightly associated with various disease. For instance, small size vesicles have been found to alleviate the symptoms of metabolic diseases.40 Metabolic disorders are characterized by pathogenesis driven by lipotoxicity, chronic inflammation, and oxidative stress, underpinned by dysregulated interorgan communication involving the liver, pancreas, adipose tissue, and immune system.65 Metabolic organs, such as pancreas, adipose tissue, skeletal muscle, and liver, secrete small size vesicles by autocrine or paracrine secretion, which transfer to other metabolic organs. These small size vesicles facilitate the intercellular communication and mediate the cross-talk among multiple metabolic organs, finally alleviating the progression of metabolic diseases.66
Cell Proliferation and Maturation
Small size vesicles contribute to the reproduction and development of organisms, especially in the formation of germ cells. Studies have shown that supporting cells-derived exosomes in the testes facilitate the entrance of seminiferous tubules to mediate sperm differentiation.67–69 Human seminal plasma-derived exosomes can participate in the occurrence, modification, and fertilization of sperm, meanwhile, they are also related to sperm motility, fertilization, and acrosome reactions.67 Human follicular fluid-derived exosomes in female ovaries play an important role in physiological processes including cumulus dilation, fertilization, and embryonic development.67,70 Human endometrial cells-derived exosomes promote implantation by regulating the proliferation and adhesion of trophoblast cells.70 Human endometrial exosomes play an important role in embryonic dormancy. During early embryonic development, endometrial epithelium-derived exosomes induce embryonic diapause by inhibiting the myelocytomatosis viral oncogene homolog / mechanistic target of rapamycin1 and 2 signaling pathways.71 Human placenta contains-derived exosomes various growth factors, miRNAs and mRNAs, which can participate in regulating the physiological functions of the maternal uterus and fetal development, and are important indicators for detecting placental activity.72
Inflammatory Response
Small size vesicles can also effectively suppress inflammatory response. Human BMSC-exos has been demonstrated to downregulate interleukin-1β (IL-1β), interleukin-6 (IL-6), and TNF-α to alleviate the symptoms of Alzheimer’s disease in mice.73 Meanwhile, miRNAs in hUCMSC-exos regulates the canonical inflammatory NF-κB signaling pathway to alleviate chronic skeletal muscle inflammation.74 However, not all small size vesicles can alleviate inflammation. Deng et al reported that after peripheral blood monocytes take up exosomes released from adipose tissue of obese mice, they differentiate into activated macrophages, with increased the secretion of TNF–α and IL-6.75
Immunoregulation
As a type of nano-vesicle, small size vesicles can activate and regulate immune responses.76 DEX contain MHC-I, MHC-II, CD86 and CD86, which contribute to activate CD4+and CD8+T cells, finally leading to anti-tumor immunity.77 Small size vesicles secreted by B lymphoma cells subjected to heat shock contained high level of HSP60 and HSP90, along with enhanced expression of immunogenicity-related molecules including MHC-I, MHC-II, CD40 and CD86. These small size vesicles effectively active CD8+ T cells, leading to a potent anti-tumor immune response.78 NK cells-derived small size vesicles can directly diffuse into tumor tissues to exert their cell lytic activity and participate in immune responses.79 Mast cell-derived small size vesicles upregulate the expression of MHC-II, CD40, CD80 and CD86 in immature DCs, promote antigen-presenting ability of T cells, ultimately activate antigen-specific immune responses.80
Small size vesicles are also associated with the migration of tumor cells. Zhao et al reported that miR-934 in human colorectal cancer cells-derived exosomes active macrophages and promote liver metastasis of colorectal cancer.81 Meanwhile, small size vesicles can also serve as a diagnosis biomarker to detect the type and progression of cancer. A promising diagnostic approach for early cancer detection and metastasis monitoring is non-invasive identification of the location and expression levels of biomarkers in cancer cells. Looking for highly specific and sensitive biomarkers remains a central focus in non-invasive cancer diagnosis.82 Advances in small size vesicles research discovered that some proteins on exosomes can serve as promising biomarkers for cancer. For instance, Melo et al found that Glypican-1 on exosomes in patients’ blood had high sensitivity and specificity in the early stage of pancreatic cancer.83 ECM and protease related proteins were highly expressed in non-small cell lung cancer cells-derived exosomes.84 All these studies indicated the potential role of small size vesicles in cancers.
Miniature Nanoreactors
It is well known that miniature nanoreactors, which can be prepared by combining enzymes with natural or artificially synthesized materials, displays a significant advantage in treating diseases such as cancer and rare genetic disorders.85 Due for the natural nanostructures, small size vesicles may be applied in nanoreactors, carrying a variety of bioactive molecules and possessing strong biocatalytic potential.40 Wan et al found that miniature exosomes nanoreactors enhanced the catalytic efficiency of enzymes and improved therapeutic efficacy They combined M1 phenotype macrophages derived exosomes and zeolitic imidazolate framework-8 nanostructure to construct miniature nanoreactors, which inhibited the activity of P. gingivalis by increasing the biosynthesis of nitric oxide and effectively increased the degree of alveolar bone resorption in periodontal disease rats.86 Though applied in miniature nanoreactors, small size vesicles fully exerted their biochemical characteristics and strengthened their therapeutic effects. Although studies on miniature small size vesicles nanoreactors remain limited, this application offers a powerful solution for future nano-therapeutic engineering.
Interact with Extracellular Matrix
Extracellular matrix is a proteinaceous network that provides a dynamic and heterogeneous microenvironment.87 It has been demonstrated that ECM could mediate the small size vesicles secretion and uptake, meanwhile small size vesicles cargo affects ECM remodeling.88 The physical properties of ECM, such as elasticity, also affects the secretion of small size vesicles.88 For instance, the number of small size vesicles released by MSCs cultured on soft hydrogel is higher than that released cultured on hard hydrogel or on tissue culture plastic.89 Some substances in small size vesicles, such as MMPs and insulin-degrading enzyme, are released into the ECM to participate in the remodeling of the matrix.88 Hoshino et al discovered cancer-associated fibroblasts derived exosomes released MT1-MMP, leading to ECM degradation and promoting the maturation and stability of the invadopodia.90 Insulin-degrading enzyme in microglia derived exosomes enhanced the degradation of Aβ in mice treated with lovastatin.91 All these studies suggest the interaction between small size vesicles and extracellular matrix.
The Application of Mesenchymal Stem Cells-Derived Exosomes in Liver Diseases
Liver is a digestive and metabolic organ and involved in several physiological processes. As a public health problem, liver disease accounts for high morbidity and mortality worldwide.92 Liver disease occurs throughout the human lifespan. For example, cholestatic liver disease commonly caused by structural and genetic factors in newborns.93 In pregnant women, hepatitis can be caused by hepatitis virus or herpes simplex virus.94 There are many methods used to diagnose liver diseases nowadays, including hematological indicators (alanine aminotransferase, aspartate aminotransferase), liver biopsy, ultrasound, magnetic resonance spectroscopy and transient elastography.95
Nowadays, there are many emerging biological therapies for liver diseases treatment, such as miRNA and antibody-drug conjugates.96,97 Among these, growing research is exploring small size vesicles therapy as a novel treatment approach for liver diseases. It has been reported the therapeutic effects of small size vesicles on liver diseases, including hemp sprout-derived exosome, tangerine peel-derived exosome and milk-derived exosomes.98–100 Additionally, the components of small size vesicles, such as some specific miRNAs and protein, were also found to directly stimulate or inhibit certain genes to alleviate liver disease.101 In Table 1, we summarized the therapeutic effects of contents from some small size vesicles on liver diseases to illustrate their functions.
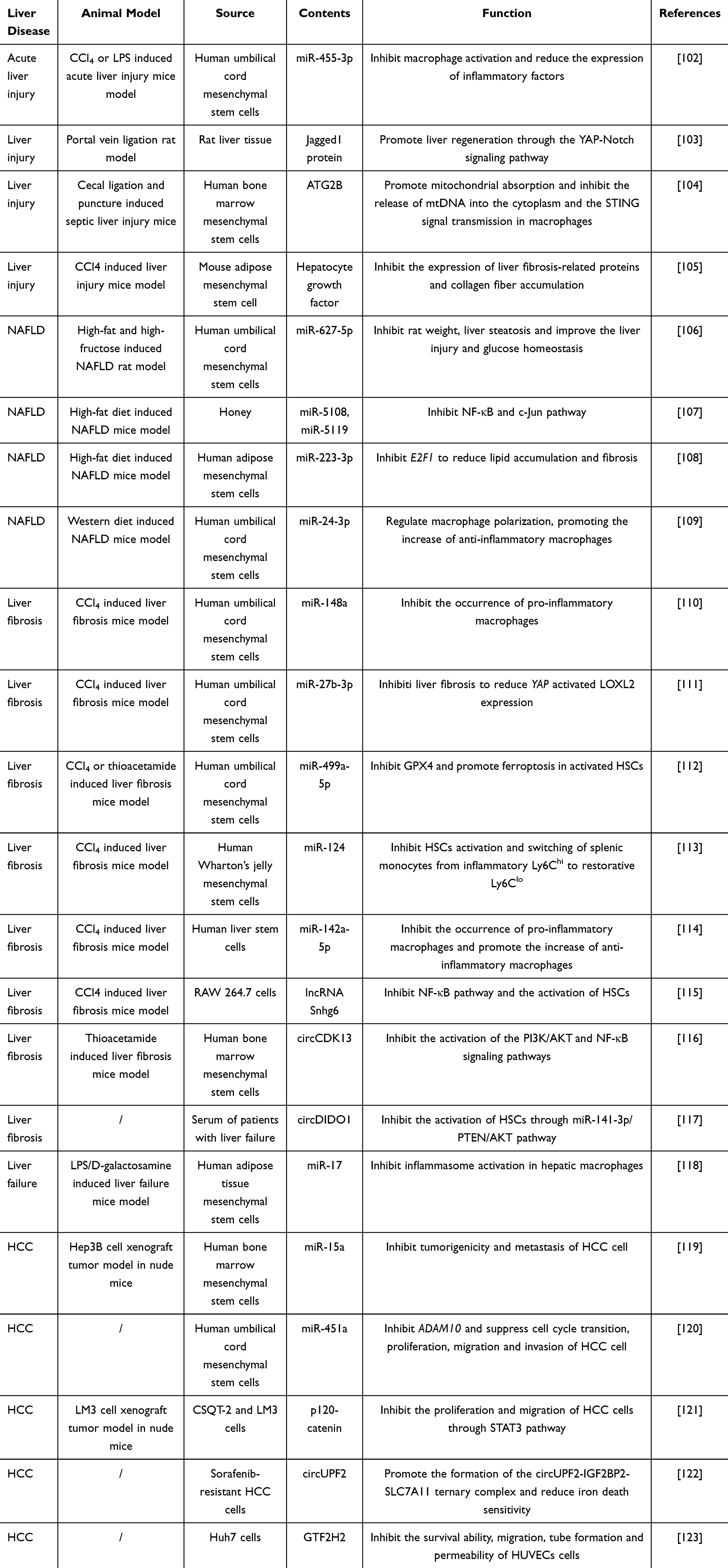 |
Table 1 The Role of Contents in Small Size Vesicles in the Treatment of Liver Diseases |
Small size vesicles also displayed potential capability for the diagnose in liver diseases. In the development of novel, reliable and non-invasive diagnostic strategies for liver diseases, human blood-derived exosomes and human urine-derived exosomes have emerged as promising tools for early detection and prognosis.101,124 The specific protein and nucleic acid in human blood-derived exosomes and human urine-derived exosomes traverse the blood-brain barrier, effectively mirroring the underlying physiological and pathological status of the liver.101 In Table 2, we summarized the small size vesicles as possible biomarkers in different liver diseases.
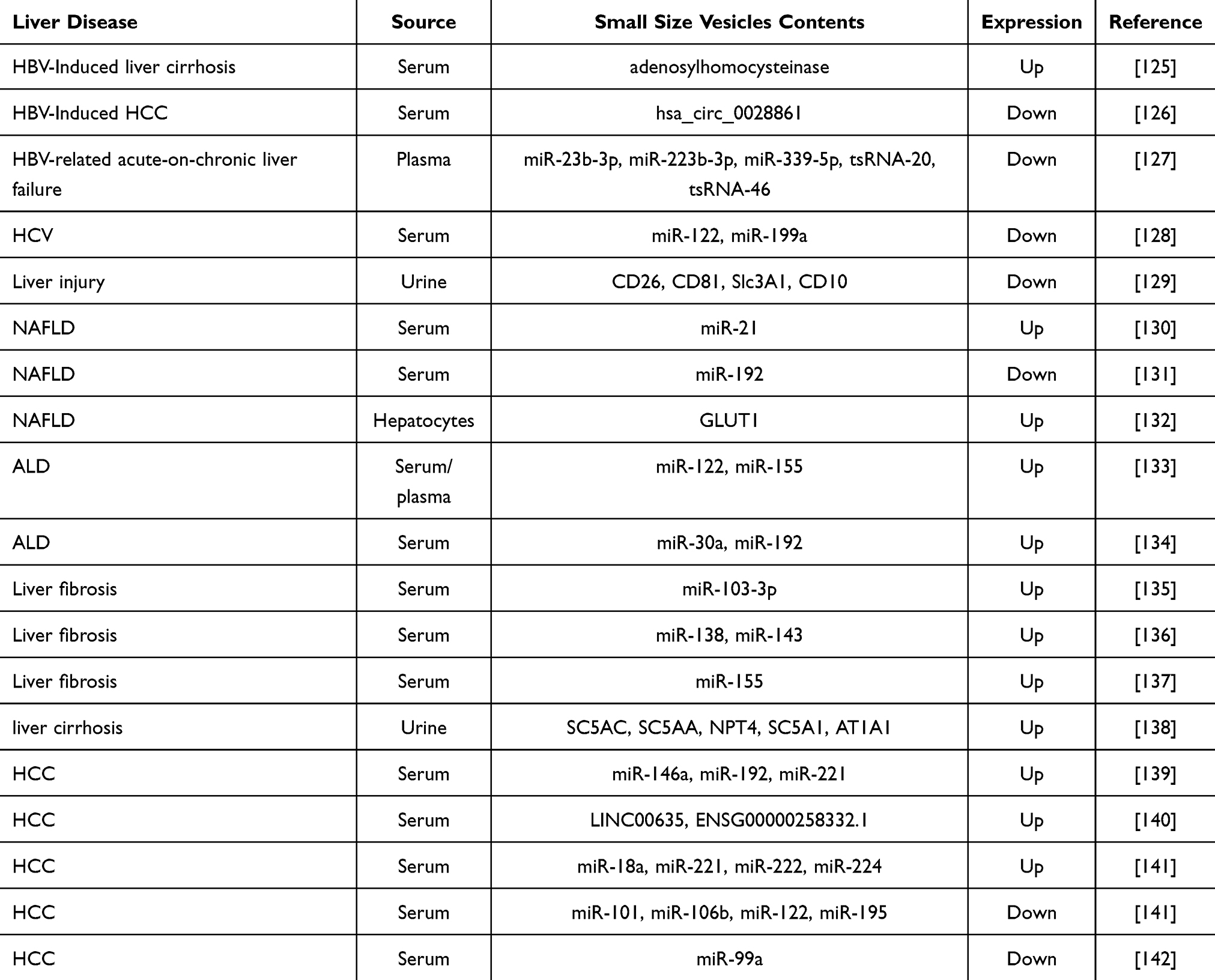 |
Table 2 Potential of Small Size Vesicles as Biomarkers for Liver Disease Diagnosis |
From the aspect of liver disease treatment, utilizing stem cells derived small size vesicles from the liver as a treatment method for liver diseases seems to be the best option. Indeed, liver-derived small size vesicles exhibits biological activity and there are several published articles about the function of EVs from liver cells on liver disease.143,144 We believe that liver-derived small size vesicles will play a significant role in the diagnosis and treatment of liver diseases in the further. Notably, as one of the most available primary stem cells in the human body, MSCs also exhibits excellent biological activity including anti-inflammatory, anti-fibrotic, immunosuppressive effects and so on. MSCs are an ideal source of small size vesicles due to their less adverse reactions.145 Interestingly, exosomes secreted by MSCs was also been found excellent therapeutic effects in liver disease.146,147 Here, we attempt to summary recent advances in the therapeutic applications of MSCs derived small size vesicles for liver diseases.
Effect the Mesenchymal Stem Cells Derived Small Size Vesicles on Liver Injury
Drugs and exogenous toxins may cause liver damage and lesions.148 Acute liver injury usually refers to damage to liver cells caused by excessive use of drugs, toxic foods and so on. Due to the availability of over 1000 drugs on the market and the development of many new drugs and the synthesis of new chemical substances in recent years, drug-induced liver injury attracts more and more attention. According to statistics, the most common drugs involved in liver injury are anti infection drugs, psychotropic drugs and lipid-lowering drugs.149 Exclude the drug abuse induced liver injury, other risk factors such as chemical poison also may induce liver injury. Liver injury is usually accompanied by oxidative stress. In the process of liver drug or chemical poison metabolism, there is a decrease in ATP synthesis and an increase in reactive oxygen species (ROS) levels.150 These drugs or toxic substances interact with cytochrome P450, generating toxic products such as free radicals, which will further bind to proteins and nucleic acids, or cause lipid peroxidation, leading to cell necrosis.151,152 When the risk factors persist for a long time, the acute liver injury may develop into liver fibrosis and HCC.153
Nowadays, MSCs derived small size vesicles have become a potential therapeutic strategy for alleviating liver injury. MSCs derived small size vesicles promote the hepatocyte proliferation by regulating the cell cycle. Human embryonic stem cell-derived exosomes upregulated the expression of cell proliferation markers in the G1 and S phases to promote cell proliferation in acetaminophen (APAP) or hydrogen peroxide (H2O2) stimulated hepatocytes.154 MSCs derived small size vesicles reduce inflammation by targeting inflammatory-related genes through the miRNAs or suppress the protein levels of inflammatory factor-related signaling pathways to mitigate inflammation response. For example, in the carbon tetrachloride (CCl4) or lipopolysaccharide (LPS) induced acute liver injury mice, miR-455-3p in hUCMSC-exos inhibited macrophage activation and the production of inflammatory factors to alleviate liver injury.102 Wu et al found that hUCMSC-exos downregulated the expression of inflammatory factors with upregulation of the ERK1/2 and PI3K/AKT signaling pathway to inhibit oxidative stress-induced cell apoptosis in APAP induced liver injury mice.155 Recent report suggests that the combined cross-effect of ferroptosis and lactate accumulation stimulated the inflammatory response in acute liver injury mice.156,157 Tian et al found that BMSC-exo can also be used as a delivery carrier By loading MnO2 and dexamethasone, MSCs-exo/MnO2@dexamethasone decreased ROS production, which promoted the redox homeostasis and inhibited ferroptosis in the liver. MSC-exo/MnO2@dexamethasone also reduced lactate levels by generating oxygen and alleviate inflammation.158 Most engineered small size vesicles can be used for the treatment of liver disease by acting on multiple targets. Just like the small size vesicles nanoparticles designed by Tian et al, they not only regulated the lactate levels to alleviate inflammation, but also reduced oxidative stress and ferroptosis to achieve multi-trageted treatment.
Effect the Mesenchymal Stem Cells Derived Small Size Vesicles on ALD
ALD is a liver disease caused by long-term alcoholism. According to incomplete statistics, men consume 60–80 g of alcohol per day or women consume 20 g of alcohol per day for 10 years or longer, which usually develop to ALD.159 Individual variation in ALD is primarily attributed to a complex interplay of genetic and environmental factors. ALD involves multiple mechanisms, the most prominent of which is the toxic effect mediated by acetaldehyde, a metabolite of alcohol. In addition, oxidative stress and cytokine mediated inflammatory responses also involved in the progression of ALD.160 The ethanol metabolism in the liver is mediated by alcohol dehydrogenase and generates acetaldehyde, which is then converted to acetate.161 This progression generates a large amount of nicotinamide adenine dinucleotide (NADH). Excessive NADH lead to metabolic disorders, inhibit the oxidation of fatty acids, and cause alcoholic fatty liver.162
Alcohol abuse affects the transcription factors related to lipid metabolism in the liver, including the hepatic fatty acids production and oxidation. It has been found that alcohol increased the synthesis of fatty acids in hepatocytes by upregulating sterol regulatory element-binding protein 1c (SREBP-1c). Knocking off SREBP-1c reduced ethanol-induced fatty liver in mice.163 Alcohol can also inhibit ECM, leading to a decrease in the activity of p-ACC and carnitine palmitoyltransferase1.164
Additionally, gut microbiota is also tightly associated with the alcohol metabolism. Kong et al found that when mice suffer from alcoholic fatty liver, the proportion of Muribaculaceae significantly decreases, while the number of Lachnospiraceae, Runminococcaceae and Erysipelotrichaceae increases significantly.165 They also found that alcohol induced gut microbiota imbalance and reduced the expression of enterohepatic protective factors with activating the NF-κB signaling pathway to produce inflammation in the liver.165
The therapeutic potential of MSCs derived small size vesicles for ALD has not been well explored. Up to now, there are only some small size vesicles from other sources exhibited great potential for ALD treatment. Kim et al found that pomegranate-derived exosomes effectively prevent liver damage by reducing hepatic oxidative stress and apoptosis in mice with alcohol abuse.166 Pueraria lobata root-derived exosomes inhibited alcohol induced ferroptosis and decreased liver lipid accumulation.167 These results indicate the use of MSCs derived small size vesicles as a potential treatment for ALD.
Effect the Mesenchymal Stem Cells Derived Small Size Vesicles on NAFLD
NAFLD is a metabolic disease with high prevalence worldwide, which has become a parallel epidemic to obesity.168 NAFLD ranges from the non-alcoholic fatty liver (NAFL) to the more severe non-alcoholic steatohepatitis (NASH). NAFL typically manifests as steatosis and mild lobular inflammation. NASH includes hepatocyte ballooning degeneration, diffuse lobular inflammation and almost always fibrosis.169 Without intervention, NAFLD can advance to liver fibrosis, which may further progress to liver cirrhosis and HCC.170 In addition, NAFLD is closely associated with other diseases. There is evidence to suggest that NAFLD can affect extrahepatic organs and increase the risk of type 2 diabetes mellitus (T2DM), cardiovascular disease (CVD) and chronic kidney disease.171 In NAFLD, the accumulation of lipid in liver is driven by ceramides and diacylglycerols, which have been demonstrated to inhibit insulin signaling and cause hepatic insulin resistance.172 In addition, lipid overload in pancreatic beta-cells promotes insulin secretion and alters the expression of PPARα, glucokinase, glucose transporter 2 and proinsulin, which enhances insulin resistance in human body.173 Individuals with NAFLD exhibit a specific lipid profile, including elevated triglyceride-rich lipoproteins, low HDL-cholesterol and increased proportion of circulating small-dense particles of LDL-cholesterol. This specific lipid profile causes the occurrence of CVD.171 Exclude T2DM and CVD, there are also studies found the association between NAFLD and colorectal cancer and osteoporosis.174
Many factors, such as lipid accumulation, endoplasmic reticulum stress, mitochondrial dysfunction, oxidative stress and inflammation, are involved in the progression of NAFLD. These complex and intertwined factors lead to significant individual and clinical differences in NAFLD.175 Lipid accumulation in NAFLD is charactered by excessive synthesis of triglycerides (TG) in hepatocytes. Many substrates account for TG synthesis such as white adipose tissue, de-novo lipogenesis and the consumption of a high-fat and/or high-sugar diet. However, a significant increase of hepatic lipid accumulation lead to mitochondrial dysfunction, which results in the inflammation response and oxidative stress in the liver. Also, driven in part by lipid accumulation, endoplasmic reticulum stress results in an accumulation of unfolded proteins, triggering the unfolded protein response and ultimately cause cell apoptosis. Similarly, when the levels of lipophilic substances in hepatocytes continue increasing, liver damage will worsen and progress to more severe disease.176
The therapeutic effect of MSCs derived small size vesicles on NAFLD is manifested through multiple mechanisms, including reducing lipid accumulation, lowering inflammation and reducing oxidative stress. Yang et al found that the calcium/calmodulin-dependent protein kinase 1 in hUCMSC-exos reduced lipid accumulation by inhibiting SREBP-1C mediated fatty acid synthesis and enhancing peroxisome proliferator activated receptor α mediated fatty acid oxidation in NAFLD mice.177 Nie et al discovered that apical papilla stem cells derived exosomes promoted the expression of genes related to fatty acid oxidation and transport in the liver and simultaneously inhibited genes expression related to fatty acid synthesis to regulate fatty acid metabolism in NASH mice model.178
In addition, MSCs derived small size vesicles suppress inflammation response in NAFLD. Zhao et al isolated and obtained ADSC-exos from mice epididymal adipose tissue and administered them to obese mice Results found that STAT3 in ADSC-exos activated arginase-1 and induced the anti-inflammatory M2 phenotype in white adipose tissue.179 Similarly, in mice fed a high-fat high cholesterol (HFHC) diet, hUCMSC-exos treatment decreased the frequency of F4/80+ macrophages in the liver and altered macrophage polarization, reducing the M1 pro-inflammatory phenotype with increasing the M2 anti-inflammatory phenotype.180
In NAFLD, the accumulation of triglycerides leads to excessive production of ROS and oxidative stress. Multiple reports have confirmed that nuclear factor erythroid 2-related factor 2 (Nrf2) participates in cellular antioxidant response.181 Nrf2 translocates from the cytoplasm to the nucleus and activates the expression of antioxidant genes, such as quinone oxidoreductase 1 and superoxide dismutase. Kang et al demonstrated that hUCMSC-exos promote the phosphorylation of Nrf2 to activate quinone oxidoreductase 1 expression in NAFLD mice.180 Du et al found that hUCMSC-exos reduced ROS production and superoxide dismutase content with decreasing kelch like ECH associated protein 1 and NADPH oxidase 2 expression to inhibit oxidative stress in mice primary hepatocytes stimulated with palmitic acid They also found that miR-24-3p derived from hUCMSC-exos inhibited lipid accumulation, ROS production and inflammatory response in palmitic acid stimulated mice primary hepatocyte.182
Exclude these, MSCs derived small size vesicles are found to be involved in some other mechanisms in liver disease. For example, Moayedfard et al discovered that mice ADSC-exos reduce endoplasmic reticulum stress indicators, apoptotic markers and restore autophagy processes in NASH mice.183
Though some studies have only reported the therapeutic effects of small size vesicles themselves, some studies found the specific cargo components of the small size vesicles attributed to regulating the underlying mechanism, such as the miRNAs and proteins. Therefore, subsequent therapeutic studies may be conducted by enriching for these components.
Effect the Mesenchymal Stem Cells Derived Small Size Vesicles on Liver Fibrosis
Liver fibrosis is the advanced progression of hepatitis and fatty liver. Several risk factors induce liver fibrosis such as chronic HCV infection, alcohol abuse and NASH.184 Advanced liver fibrosis results in cirrhosis and liver failure, which often requires liver transplantation.185 The main characteristic of liver fibrosis is the accumulation of ECM. There are significant alterations about the quantity, quality and distribution of ECM in liver fibrosis. Fibrosis alters the composition of the hepatic ECM, replacing the normal collagen IV/VI in the space of Disse with a collagen I/III- and fibronectin-rich matrix, which triggers sinusoidal capillarization.184 HSCs activation accounts for the excessive production of ECM. In a physiological state, HSCs are quiescent and non-proliferative. When the liver is stimulated, HSCs differentiate from a quiescent phenotype to an activated phenotype. During this process, a large amount of ECM components and pro-inflammatory mediators are produced. In addition, activated HSCs express a large amount of alpha smooth muscle actin and tissue inhibitor of metalloproteinases 1, promoting the formation of fibrosis.24 Some cytokines, such as transforming growth factor-β (TGF-β), platelet-derived growth factor, chemokines, leptin and adiponectin, are also involved in liver fibrosis. TGF-β is a key factor in the progression of liver fibrosis. TGF-β activate HSCs, transforming them into myofibroblast like cells. This progression stimulates the synthesis of ECM proteins with blocking their degradation pathways. It has been widely accepted that TGF-β signaling pathway is a promising target for treating liver fibrosis.186 Platelet-derived growth factor is an effective mitogen in HSCs and is upregulated in fibrotic liver. Inhibiting platelet-derived growth factor activity was found to reduce liver fibrosis formation.187 Chemokines drive inflammation response with promoting the migration of fibrotic cells to the injury site, thereby exacerbating the severity of the injury in liver fibrosis. The interaction between chemokine receptors and ligands plays different role in liver fibrotic pathology. For example, the C-C motif chemokine receptor 5, the C-C motif chemokine receptor 1 and the chemokine (C-X-C motif) ligand 4 act synergistically to promote fibrosis, while the interaction between the chemokine (C-X-C motif) ligand 9 and the chemokine (C-X-C motif) receptor 3 prevents fibrosis.188 Some cytokines in adipose tissue also regulate the progression of liver fibrosis. Studies have shown that leptin inhibit the progression of liver fibrosis, while adiponectin possess anti-fibrotic activity.189
Studies indicate that MSCs derived small size vesicles exhibit efficacy in ameliorating liver fibrosis, such as preventing the activation of HSCs and reducing collagen formation. Human ADSC-exos alleviated collagen and liver damage through inhibiting the p38 MAPK/NF-κB pathway in mice.190 Wu et al found that human ADSC-exos treatment inhibited HSCs activation and improved glutamine and ammonia metabolism, thereby improving liver ammonia circulation in CCl4-induced liver fibrosis mice.191 Zhang et al found that human ADSC-exos suppressed LX2 cell activation and inhibited liver fibrosis progression in mice by regulating choline metabolism and inhibiting the PI3K/AKT/mTOR signaling pathway.192 In addition, hUCMSC-exos was found to downregulate the expression of smoothened and inhibited the HSCs activation, exerting anti-fibrotic effects in the liver.193 In line, hUCMSC-exos induced ferroptosis of HSCs by regulating the xCT/GPX4 axis.194 Human BMSC-exos reduced collagen accumulation and inhibited Wnt/β-catenin pathway in HSCs.195
Given the therapeutic role of small size vesicles in liver fibrosis, the small size vesicles miRNA exhibited potential effects on the disease progression.196 MiRNAs in small size vesicles have shown significant efficacy in treating liver fibrosis. Tian et al found that hUCMSC-exos alleviated liver damage and reduced fibrosis-related phenotypes and they also found that miR-148a enriched in hUCMSC-exos targeted Kruppel like factor 6, which further inhibited the STAT3 signaling pathway to suppress the generation of pro-inflammatory macrophages, thereby contributing to protect against liver fibrosis in mice.110 Qu et al reported that human ADSC-exos alleviated liver fibrosis markers such as collagen I and α-SMA in CCl4-induced liver fibrosis mice, while, miR-181-5p in human ADSC-exos downregulated Stat3 and Bcl-2 to activate autophagy in HST-T6 cells.197 Human placenta stem cell-derived exosomes alleviated fibrotic phenotype, and miR-378c in exosomes targeted E3 ubiquitin ligase S phase kinase associated protein 2 to inhibit epithelial mesenchymal transition, inactivating HSCs in mice.198 3D human embryonic stem cells-derived exosomes enriched with miR-6766-3p suppressed HSCs activation through targeting the TGFβRII-SMADS pathway, which ameliorated the liver fibrosis in mice.199
In conclusion, miRNAs in MSCs derived small size vesicles alleviate liver fibrosis through multiple pathways, including regulating the polarization of M1 macrophages to M2 macrophages, inhibiting the activation of HSCs, reducing TGF-β secretion and promoting autophagy.200 Utilizing miRNAs from small size vesicles as therapeutic strategy provides new insights into the prevention of liver fibrosis.
Effect the Mesenchymal Stem Cells Derived Small Size Vesicles on Liver Cirrhosis
According to statistics, approximately 2 million people worldwide die from liver disease each year, with an estimated 1 million dying from cirrhosis and the other 1 million from other liver diseases such as HCC. Liver cirrhosis is one of the common causes of death worldwide and the third leading cause of death among people aged 45–64.201 Liver cirrhosis causes the metabolic disorders in the body leading to an increase in bilirubin and portal vein pressure, along with a decrease in coagulation factors and thrombopoietin.202 Many factors are associated with the progression of liver cirrhosis, such as alcoholism, metabolic liver disease, viral hepatitis, autoimmune hepatitis, primary biliary cholangitis, etc. In some patients, there are more than one factors that accelerates the progression of the disease.203
There are some studies reported the preventive and therapeutic effects of MSCs derived small size vesicles on liver cirrhosis. MSCs derived small size vesicles reduce the expression of fibrosis-related factors to alleviate liver cirrhosis. Abd Allah et al found that hUCMSC-exos reduced the expression of TGF-β, MMP 9 and α-SMA, which contributed to the amelioration of liver cirrhosis in rat.204 Mardpour et al demonstrated that human embryonic stem cells-derived exosomes alleviated liver cirrhosis in rats by upregulating the expression of collagenase, BCL-2 and anti-inflammatory cytokines, while concurrently downregulating α-SMA and TIMP1.205
It is widely known that MSCs are able to exert their effects via exosomes. There are not enough examples of MSCs derived small size vesicles used for treating liver cirrhosis, however, there are many successful cases of MSCs clinical therapy for liver cirrhosis. Shi et al recruited 219 patients with HBV related decompensated liver cirrhosis for their study Their research results indicate that human umbilical cord MSCs treatment can significantly improve the long-term survival rate and liver cirrhosis in patients.206 Lu et al analyzed 11 clinical trials and found that human MSCs therapy reduced MELD scores compared to the liver cirrhosis group.207 Jang et al demonstrated that transplantation of autologous human BMSCs can treat alcoholic liver cirrhosis They showed that 54.5% patients observed histological improvement according to the Laennec fibrosis system, demonstrating the therapeutic potential of MSCs in liver cirrhosis.208 Though there are several examples about MSCs in liver cirrhosis. MSCs derived small size vesicles exhibits more advantages over MSCs in terms of liver cirrhosis treatment. For instance, MSCs derived small size vesicles eliminate the potential tumor formation, unnecessary differentiation and infection transmission.209 All these indicats the therapeutic potential about potential MSCs derived small size vesicles in liver cirrhosis.
Effect the Mesenchymal Stem Cells Derived Small Size Vesicles on Liver Failure
Liver failure is an acute decompensation of liver function with multiple organ dysfunctions in body and its clinical features are manifested as coagulation disorders, jaundice and ascites.210,211 Liver failure divided into four categories: acute liver failure, subacute liver failure, acute non chronic liver failure and chronic liver failure.212 The common causes of liver failure include HBV, HAV, hepatitis E virus (HEV) infection, alcohol abuse and bacterial infection.213 Although liver has regenerative ability, liver failure is a serious and irreversible disease.214 At present, the main methods for treating liver failure are comprehensive medical care, artificial liver support systems and liver transplantation, but the mortality rate of liver failure is still high.210
Although the treatment of liver failure is difficult, there are some examples that demonstrated the great potential of MSCs derived small size vesicles in treating liver failure. MSCs derived small size vesicles alleviate inflammation response in liver failure. For example, mice ADSC-exos reduced the secretion of inflammatory factors by inhibiting the activation of inflammasomes in LPS/GalN induced acute liver failure mice model.118 Zhang et al found that pretreatment of hUCMSC-exos attenuated inflammatory damage in liver and promoted liver tissue repair by inhibiting the activation of the PYD domains-containing protein 3 pathway.215 MSCs derived small size vesicles also induced liver regeneration by upregulating angiogenesis. Human placenta stem cell-derived exosomes effectively increased C-reactive protein levels and upregulated Wnt signaling pathway, which promoted liver regeneration and alleviated liver failure in rat.214 Pyroptosis is a form of programmed cell death in hepatocytes and is regarded as an important pathological feature of liver failure.216 Xie et al found that miR-423-5p in hUCMSC-exos targeted ZBP1 to inhibit ATP and LPS-induced hepatic pyroptosis in LO2 cells.217 MSCs derived small size vesicles restored autophagic flux to protect hepatocyte against death. Lin et al found that let-7a-5p in human BMSC-exos accelerated the nuclear translocation of transcription factor EB (TFEB) and induced the expression of lysosomal genes to promote autophagy, which contributed to the prevention of hepatocyte death.218
It is worth noting that liver transplantation remains the only cure for end-stage liver disease.219 However, liver transplantation often leads to immune rejection and inflammatory responses. Small size vesicles possess immunoregulatory and anti-inflammatory functions, which alleviate the immune rejection and inflammation after the liver transplantation.220 For instance, donor DEX, enriched with immunosuppressive factors such as TGF-β1 and CTLA-4, inhibited the activation of T cells and regulated other immune cells to maintain the immune tolerance.221 DEX also promoted the polarization of macrophages from the pro-inflammatory M1 phenotype to the immunosuppressive M2 phenotype, thereby enhancing the local immunosuppressive microenvironment and alleviating the immune exclusion response.222 Tian et al discovered that mouse MSC-exos inhibited the expression of pro-inflammatory cytokines and chemokines in the non-heart-beating liver transplantation mouse model and suppressed the pyroptosis of Kupffer cells by regulating the TXNIP-NLRP3-Caspase1 signaling pathway.222 Liver transplantation is also susceptible to the risk of ischemia-reperfusion injury. Severe ischemia-reperfusion injury requires retransplantation.223 Deng et al found that human bone marrow MSCs derived exosomes delivered miR-16-5p to liver and inhibited the expression of SLC39A14 to reduce cell ferroptosis in ischemia-reperfusion injury mice.224 All these studies suggest that small size vesicles is a potential therapeutic approach to increase the survival rate by alleviating the immune rejection reaction, inflammation and liver damage after liver transplantation.
Effect the Mesenchymal Stem Cells Derived Small Size Vesicles on HCC
HCC is a primary malignancy of the liver and constitutes a major global health problem.225 Due for the poor prognosis and high mortality rate, HCC is listed as the sixth most common tumor and the third largest cancer. According to statistics, the number of HCC cases continues increasing every year.226 The risk factors associated with HCC include infection with HBV or HCV, exposure to aflatoxin B1, alcohol abuse, metabolic dysfunction associated steatohepatitis and so on.227 HCC involves sustained inflammatory damage, hepatocyte necrosis and fibrotic deposition. The development of liver cirrhosis markedly increases the risk of HCC.228 Nowadays, the treatment methods for HCC include liver resection, liver transplantation, radiation therapy and chemotherapy.227
Recently, MSCs derived small size vesicles therapy is a promising treatment strategy for HCC. Gao et al found that human ADSC-exos reduced tumor volume, promoting natural killer T-cell anti-tumor response and inhibiting HCC in rat.229 Xu et al found that miR-451a in hUCMSC-exos suppressed paclitaxel resistance, proliferation, epithelial-mesenchymal transition and cell cycle progression in Hep3B and SMMC-7721 cell through inhibiting ADAM10.120 lncRNA FAM99B in hUCMSC-exos inhibited malignant cellular phenotypes and tumorigenesis in HCC.230 Ma et al found that miR-15a in human BMSC-exos impeded HCC progression via downregulation of SALL4.119 Ding et al found that human BMSC-exos inhibited the migration, invasion and proliferation in HCCLM3 and SK-Hep-1 cells They also reported that miR-374c-5p in human BMSC-exos suppressed epithelial mesenchymal transition in hepatocellular carcinoma by inhibiting the Wnt/β-catenin pathway.231
Notably, MSCs derived small size vesicles can also be used as a delivery nano-material for the treatment of HCC. Prior study showed that loading miR-122 into ADSC-exos promoted HepG2 and Huh-7 cell sensitivity to sorafenib and 5-fluorouracil.232 Lou et al found that ADSC-exos effectively delivered miR-199a to Huh-7, SMMC-7721 and PLC/PRF/5 cells to inhibit the development of HCC through mTOR signaling pathway.233 MSCs derived small size vesicles can also deliver drugs and others through engineered modification. For example, Liang et al loaded the anti-cancer drug NCTD into human BMSC-exos through electroporation They found that the BMSC-Exos-NCTD delivery system arrested the cell cycle of HCC cells and increased cell apoptosis in rats.234 Lou et al loaded miR-122 into human ADSC-exos, which enhanced the growth inhibitory effect of sorafenib on HCC cells, providing a new therapeutic strategy for HCC.232 In summary, MSCs derived small size vesicles achieve the therapeutic effect on HCC, at least partly, by delivering various types of drugs and others.
Challenges and Future Perspectives
Although the research on EVs has developed rapidly, recent studies have found that not all substances secreted into the extracellular space are carried by EVs.235 Non-vesicular extracellular nanoparticles (NVEPs) are a type of diverse membraneic particles released from cells, and they lack a lipid bilayer membrane.236 NVEPs can carry and release proteins, RNA and DNA, which have been proven to be the main carriers for the release of bioactive substances.235 NVEPs include a variety of types, such as albumin, exomeres, lipoproteins, supermers, vault and viral particles. Albumin can bind and transport endogenous molecules, including long-chain fatty acids, bilirubin, heme and vitamin D, which has antioxidant, antithrombotic and immune-regulating functions.237 Schleicher’s research indicates that some patients with liver cirrhosis and acute kidney injury respond positively to albumin treatment, such as a decrease in serum creatinine levels, which provides a basis for subsequent clinical predictions.238 Lipoproteins are widely known to transport lipids, but they are associated with nucleic acids transportation. High-density lipoprotein and low-density lipoprotein may carry small RNAs such as miRNAs and sRNAs to the target cells.239 Very low-density lipoproteins transport endogenous lipids to adipose tissue and muscle tissue, while chylomicrons transport exogenous lipids from the intestine to the circulation system.240 Vaults are 41 × 73 nm particles that contain multiple copies of three proteins and small untranslated RNA.241 Infected cells release virus particles.235 The virus particles are nanostructures composed of one or more structural proteins.242 Biller et al confirmed that virus particles, like EVs, mediated horizontal gene transfer.243 Exomeres are membrane-like nanoparticles ranging from 28 to 50 nanometers, containing RNA and DNA and metabolic enzymes.244 Exomeres are released under both physiological and pathological conditions in most cells and tissues.245 The size of supermeres is smaller than exomeres (22–32 nm), and supermeres can efficiently cross the blood-brain barrier.235 Meanwhile, the metabolic enzyme enriched in the supermeres is higher than in the exomeres.246 Many clinically relevant proteins, such as amyloid precursor protein, intercellular mesenchymal epithelial transfer factor and TGFβ-induced protein are also highly expressed in supermers.245
Recently, the discovery of supermeres promotes the researchers to pay more attention on re-evaluating the function of EVs in eukaryotic cells.247 The enhanced capacity of NVEPs in transporting biological information has raised doubts about the therapeutic efficacy of EVs. However, during the secretion process, cells simultaneously release both EVs and NVEPs, making it difficult to distinguish between them.235 Therefore, it is necessary to re-examine whether the efficacy of the EVs reported previously comes from the EVs themselves or from NVEP. Nowadays, how EVs can be effectively distinguished from NVEPs remains a challenge. Some scientists have developed various methods in an attempt to solve this problem. A microfluidic platform technology was employed by Mehraji et al for the separation and purification of EVs This technology effectively separated the EVs according to their size and markedly enhanced the EVs recovery rate.248 At the same time, the nanofluidic cytometry can also identify the size of EVs and sort EVs with specific characteristics.249 Super-resolution microscopy provides higher sensitivity, enabling nanoscale analysis of the size and morphological heterogeneity of EVs.250 3D hollow fiber bioreactors combined with fast-protein liquid chromatography-based size-exclusion chromatography achieve the separation of EVs and NVEPs.251 Although these methods effectively separated and purified EVs and NVEPs, a gold standard method still needs to be established urgently, which then can be used to verify the efficacy of EVs.
Due for the ability of penetrating natural barriers and low toxicity, small size vesicles attract more and more attention both as drugs and drug deliveries.252 However, natural small size vesicles also have significant drawbacks. For instance, their clearance rate in the blood is high, which limits the therapeutic efficacy and application of small size vesicles.253 Research have shown that modification of small size vesicles or combining small size vesicles with biocompatible and highly adhesive biomaterials maximized the retention of small size vesicles without their rapid clearance.254 Xing et al used heat-sensitive cell-free extracellular matrix hydrogel to sustained release of rat ADSC-exos, which regulated the matrix synthesis and alleviated intervertebral disc degeneration in rats.255 Wan et al utilized light-crosslinked spherical hydrogels to encapsulate bone marrow MSCs derived exosomes from rat, which prevented the exosomes rapid clearance and destruction in blood.253 Modifying EVs with albumin can also prolong the circulation time of EVs. Liang et al decorated albumin on the surface of EVs and injected them to mice They found these engineered EVs bound to mouse serum albumin, significantly prolonging the circulation time of EVs.256 Based on these studies, the rapid clearance of EVs in the blood may be solved by engineered modification. In conclusion, combining EVs with biomaterials enables their continuous release, meanwhile modifying the surface can extend their circulation time. These methods for modifying small size vesicles should be given full attentions to enhance the application of small size vesicles.
As an emerging therapeutic approach for liver diseases, clinical applications based on MSCs derived small size vesicles require more preclinical studies for comprehensive evaluation. The safety and potential side effects of MSCs derived small size vesicles in liver disease treatment should also be thoroughly considered.257 At present, there is no unified standard for the preparation of small size vesicles.200 Therefore, efficient separation and purification methods are needed to distinguish exosomes from NVEPs and others. In conclusion, MSCs derived small size vesicles exhibits promising potential for clinical translation and application. In the future, MSCs derived small size vesicles may be applied as drugs or delivery vectors for multi-targeted therapy, which could not only deepen existing treatment strategies for liver diseases but also emerge as a novel therapeutic option.
Conclusion
Emerging as a promising diagnostic and therapeutic candidate in clinic, small size vesicles have attracted extensive attention in recent years. Liver diseases constitute a significant global health burden and are a leading cause of mortality worldwide. In-depth investigations into cell-free therapy have demonstrated that MSCs derived small size vesicles possess broad application potential in liver diseases, including drug delivery and liver transplantation. MSCs derived small size vesicles ameliorate liver diseases through multiple mechanisms. For instance, MSCs derived small size vesicles promote liver regeneration, regulate lipid metabolism, inhibit inflammation and oxidative stress, suppress the activation of HSCs and reduce the growth of HCC cells (Figure 2). MSCs derived small size vesicles in the treatment of liver diseases involves multiple types of stem cells, such as human umbilical cord MSCs, human bone marrow MSCs, human adipose MSC, mouse or rat bone marrow MSCs and adipose MSCs. MSCs derived small size vesicles are also be recognized as a kind of biological carrier that can enter target cells, such as HSCs and HCC cells. By transporting cargo, MSCs derived small size vesicles exhibited more potential as an alternative treatment option for various liver diseases. Though there are lots of studies demonstrated the positive effects of MSCs derived small size vesicles on liver disease, the usage of MSCs derived small size vesicles in liver disease is still at the stage of in vitro research and animal levels. Clinical research on MSCs derived small size vesicles in the treatment of liver diseases is still relatively scarce. Exclude the therapeutic effect, small size vesicles can serve as markers for diagnosing the pathological progression of liver diseases. All these underscores the importance of MSCs derived small size vesicles as a key player in the treatment of liver diseases.
 |
Figure 2 Summary of underlying mechanisms about MSCs derived small size vesicles in the treatment of liver diseases. MSCs derived small size vesicles protect against liver diseases through various mechanisms, such as promoting cell proliferation, inhibiting cell apoptosis and pyroptosis, regulating lipid metabolism and glutamate metabolism, inhibiting endoplasmic reticulum stress, reducing inflammation and oxidative stress, promoting autophagy, inhibiting epithelial-mesenchymal transition, inhibiting the activation of HSCs, promoting ferroptosis of HSCs, inhibiting the proliferation, migration and invasion of HCC cells and promoting the anti-tumor response of nature killer T cells. The black arrows indicate increased levels; the red arrows indicate decreased levels. Abbreviations: HCC, hepatocellular carcinoma; HSCs, hepatic stellate cells; TGF-β, transforming growth factor-β; α-SMA, alpha-smooth muscle actin. |
Abbreviation
ADSC-exos, adipose mesenchymal stem cell-derived exosomes; ALD, alcoholic liver disease; APAP, acetaminophen; BMSC-exos, bone marrow mesenchymal stem cell-derived exosomes; CCl4, carbon tetrachloride; CVD, cardiovascular disease; DCs, dendritic cells; ECM, extracellular matrix; EMT, epithelial-mesenchymal transition; ESCRT, endosomal sorting complex required for transport; EVs, extracellular vesicles; H2O2, hydrogen peroxide; HBV, hepatitis B virus; HCC, hepatocellular carcinoma; HCV, hepatitis C virus; HEV, hepatitis E virus; HFHC, high-fat high cholesterol; HSCs, hepatic stellate cells; hUCMSC-exos, human umbilical cord mesenchymal stem cell-derived exosomes; HUVEC, human umbilical vein endothelial cells; IL-1β, interleukin-1β; IL-6, interleukin-6; ILVs, intraluminal vesicles; LPS, lipopolysaccharide; LSECs, liver sinusoidal endothelial cells; MHC-I, major histocompatibility complex-I; MHC-II, major histocompatibility complex-II; MMP 9, matrix metalloproteinase 9; MSCs, mesenchymal stem cells; MSCs-exos, mesenchymal stem cells-derived exosomes; MVBs, multivesicular bodies; NADH, nicotinamide adenine dinucleotide; NAFL, non-alcoholic fatty liver; NAFLD, non-alcoholic fatty liver disease; NASH, non-alcoholic steatohepatitis; NVEPs, non-vesicular extracellular nanoparticles; NK, natural killer; Nrf2, nuclear factor erythroid 2-related factor 2; ROS, reactive oxygen species; SREBP-1c, sterol regulatory element-binding protein 1c; T2DM, type 2 diabetes mellitus; TEXs, tumor-derived exosomes; TGF-β, transforming growth factor-β; TNF-α, tumor necrosis factor α; TSG101, tumor susceptibility gene 101; Vps, class E vacuolar protein sorting; α-SMA, alpha-smooth muscle actin.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by Hebei Natural Science Foundation (grant number H2025205003) and Science Foundation of Hebei Normal University (grant numbers L2024K03).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Krylova SV, Feng D. The machinery of exosomes: biogenesis, release, and uptake. Int J Mol Sci. 2023;24(2):1337. doi:10.3390/ijms24021337
2. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2019;19(4):213–26. doi:10.1038/nrm.2017.125
3. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the international society for extracellular vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7:1535750. doi:10.1080/20013078.2018.1535750
4. Yáñez-Mó M, Siljander PR, Andreu Z, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4:27066. doi:10.3402/jev.v4.27066
5. Batrakova EV, Kim MS. Using exosomes, naturally-equipped nanocarriers, for drug delivery. J Control Release. 2015;219:396–405. doi:10.1016/j.jconrel.2015.07.030
6. Salunkhe S, Basak M, Chitkara D, Mittal A, Mittal A. Surface functionalization of exosomes for target-specific delivery and in vivo imaging & tracking: strategies and significance. J Control Release. 2020;326:599–614. doi:10.1016/j.jconrel.2020.07.042
7. Hade MD, Suire CN, Suo Z. Mesenchymal stem cell-derived exosomes: applications in regenerative medicine. Cells. 2021;10(8). doi:10.3390/cells10081959
8. Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9:654–659. doi:10.1038/ncb1596
9. Mathivanan S, Fahner CJ, Reid GE, Simpson RJ. ExoCarta 2012: database of exosomal proteins, RNA and lipids. Nucleic Acids Res. 2012;40:D1241–1244. doi:10.1093/nar/gkr828
10. Kimiz-Gebologlu I, Oncel SS. Exosomes: large-scale production, isolation, drug loading efficiency, and biodistribution and uptake. J Control Release. 2022;347:533–543. doi:10.1016/j.jjconrel.2022.05.027
11. Yu X, Odenthal M, Fries JW. Exosomes as miRNA carriers: formation-function-future. Int J Mol Sci. 2016;17(12):2028. doi:10.3390/ijms17122028
12. Lu TX, Rothenberg ME. MicroRNA. J Allergy Clin Immunol. 2018;141(4):1202–1207. doi:10.1016/j.jaci.2017.08.034
13. Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: trafficking, sorting, and function. Genomics Proteomics Bioinf. 2015;13:17–24. doi:10.1016/j.gpb.2015.02.001
14. Rong J, Li YY, Wang X, Wang JN, Song M. Non-coding RNAs in adipose-derived stem cell exosomes: mechanisms, therapeutic potential, and challenges in wound healing. World J Stem Cells. 2025;17(4):102917. doi:10.4252/wjsc.v17.i4.102917
15. Bian D, Wu Y, Song G, Azizi R, Zamani A. The application of mesenchymal stromal cells (MSCs) and their derivative exosome in skin wound healing: a comprehensive review. Stem Cell Res Ther. 2022;13(1):24. doi:10.1186/s13287-021-02697-9
16. Gao S, Chen T, Hao Y, et al. Exosomal miR-135a derived from human amnion mesenchymal stem cells promotes cutaneous wound healing in rats and fibroblast migration by directly inhibiting LATS2 expression. Stem Cell Res Ther. 2020;11(1):56. doi:10.1186/s13287-020-1570-9
17. Xu YX, Pu SD, Li X, et al. Exosomal ncRNAs: novel therapeutic target and biomarker for diabetic complications. Pharmacol Res. 2022;178:106135. doi:10.1016/j.phrs.2022.106135
18. Trefts E, Gannon M, Wasserman DH. The liver. Curr Biol. 2017;27:R1147–r1151. doi:10.1016/j.cub.2017.09.019
19. Wu Q, Yang Y, Lin S, Geller DA, Yan Y. The microenvironment in the development of MASLD-MASH-HCC and associated therapeutic in MASH-HCC. Front Immunol. 2025;16:1569915. doi:10.3389/fimmu.2025.1569915
20. Mangnall D, Bird NC, Majeed AW. The molecular physiology of liver regeneration following partial hepatectomy. Liver Int. 2003;23:124–138. doi:10.1034/j.1600-0676.2003.00812.x
21. Eski SE, Mi J, Pozo-Morales M, et al. Cholangiocytes contribute to hepatocyte regeneration after partial liver injury during growth spurt in zebrafish. Nat Commun. 2025;16(1):5260. doi:10.1038/s41467-025-60334-y
22. Michalopoulos GK, Bhushan B. Liver regeneration: biological and pathological mechanisms and implications. Nat Rev Gastroenterol Hepatol. 2021;18:40–55. doi:10.1038/s41575-020-0342-4
23. Higashi T, Friedman SL, Hoshida Y. Hepatic stellate cells as key target in liver fibrosis. Adv Drug Deliv Rev. 2017;121:27–42. doi:10.1016/j.addr.2017.05.007
24. Kisseleva T, Brenner D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat Rev Gastroenterol Hepatol. 2021;18:151–166. doi:10.1038/s41575-020-00372-7
25. Sato K, Hall C, Glaser S, Francis H, Meng F, Alpini G. Pathogenesis of kupffer cells in cholestatic liver injury. Am J Pathol. 2016;186(9):2238–2247. doi:10.1016/j.ajpath.2016.06.003
26. Li P, He K, Li J, Liu Z, Gong J. The role of kupffer cells in hepatic diseases. Mol Immunol. 2017;85:222–229. doi:10.1016/j.molimm.2017.02.018
27. Poisson J, Lemoinne S, Boulanger C, et al. Liver sinusoidal endothelial cells: physiology and role in liver diseases. J Hepatol. 2017;66(1):212–227. doi:10.1016/j.jhep.2016.07.009
28. Gracia-Sancho J, Caparrós E, Fernández-Iglesias A, Francés R. Role of liver sinusoidal endothelial cells in liver diseases. Nat Rev Gastroenterol Hepatol. 2021;18:411–431. doi:10.1038/s41575-020-00411-3
29. Neshat SY, Quiroz VM, Wang Y, Tamayo S, Doloff JC. Liver disease: induction, progression, immunological mechanisms, and therapeutic interventions. Int J Mol Sci. 2021;22:6777. doi:10.3390/ijms22136777
30. Taru V, Szabo G, Mehal W, Reiberger T. Inflammasomes in chronic liver disease: hepatic injury, fibrosis progression and systemic inflammation. J Hepatol. 2024;81(5):895–910. doi:10.1016/j.jhep.2024.06.016
31. Wang FS, Fan JG, Zhang Z, Gao B, Wang HY. The global burden of liver disease: the major impact of China. Hepatology. 2014;60:2099–2108. doi:10.1002/hep.27406
32. El Hadi H, Di Vincenzo A, Vettor R, Rossato M. Relationship between heart disease and liver disease: a two-way street. Cells. 2020;9:567. doi:10.3390/cells9030567
33. Yang K, Song M. New insights into the pathogenesis of metabolic-associated fatty liver disease (MAFLD): gut-liver-heart crosstalk. Nutrients. 2023;15(18):3970. doi:10.3390/nu15183970
34. Ahmadieh H, Azar ST. Liver disease and diabetes: association, pathophysiology, and management. Diabet Res Clin Pract. 2014;104:53–62. doi:10.1016/j.diabres.2014.01.003
35. Schiff ER. Lamivudine for hepatitis B in clinical practice. J Med Virol. 2000;61:386–391. doi:10.1002/1096-9071(200007)61:3<386::aid-jmv18>3.0.co;2-x
36. Ma Z, Zhang Y, Li Q, Xu M, Bai J, Wu S. Resveratrol improves alcoholic fatty liver disease by downregulating HIF-1α expression and mitochondrial ROS production. PLoS One. 2017;12:e0183426. doi:10.1371/journal.pone.0183426
37. Gillessen A, Schmidt HH. Silymarin as supportive treatment in liver diseases: a narrative review. Adv Ther. 2020;37:1279–1301. doi:10.1007/s12325-020-01251-y
38. Liang Y, Duan L, Lu J, Xia J. Engineering exosomes for targeted drug delivery. Theranostics. 2021;11:3183–3195. doi:10.7150/thno.52570
39. Woodman PG, Futter CE. Multivesicular bodies: co-ordinated progression to maturity. Curr Opin Cell Biol. 2008;20:408–414. doi:10.1016/j.ceb.2008.04.001
40. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367. doi:10.1126/science.aau6977
41. Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319:1244–1247. doi:10.1126/science.1153124
42. Gurunathan S, Kang MH, Kim JH. A comprehensive review on factors influences biogenesis, functions, therapeutic and clinical implications of exosomes. Int J Nanomed. 2021;16:1281–1312. doi:10.2147/ijn.S291956
43. Kowal J, Tkach M, Théry C. Biogenesis and secretion of exosomes. Curr Opin Cell Biol. 2014;29:116–125. doi:10.1016/j.ceb.2014.05.004
44. Oestreich AJ, Davies BA, Payne JA, Katzmann DJ. Mvb12 is a novel member of ESCRT-I involved in cargo selection by the multivesicular body pathway. Mol Biol Cell. 2007;18:646–657. doi:10.1091/mbc.e06-07-0601
45. Liu CC, Liu YY, Cheng Y, et al. The ESCRT-I subunit Tsg101 plays novel dual roles in entry and replication of classical swine fever virus. J Virol. 2021:95. doi:10.1128/jvi.01928-20
46. Chu T, Sun J, Saksena S, Emr SD. New component of ESCRT-I regulates endosomal sorting complex assembly. J Cell Biol. 2006;175:815–823. doi:10.1083/jcb.200608053
47. Strickland M, Watanabe S, Bonn SM, et al. Tsg101/ESCRT-I recruitment regulated by the dual binding modes of K63-linked diubiquitin. Structure. 2022;30:289–299.e286. doi:10.1016/j.str.2021.09.006
48. Stuffers S, Sem Wegner C, Stenmark H, Brech A. Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic. 2009;10:925–937. doi:10.1111/j.1600-0854.2009.00920.x
49. Arya SB, Collie SP, Parent CA. The ins-and-outs of exosome biogenesis, secretion, and internalization. Trends Cell Biol. 2024;34:90–108. doi:10.1016/j.tcb.2023.06.006
50. Hessvik NP, Llorente A. Current knowledge on exosome biogenesis and release. Cell Mol Life Sci. 2018;75:193–208. doi:10.1007/s00018-017-2595-9
51. Jeannot P, Besson A. Cortactin function in invadopodia. Small GTPases. 2020;11:256–270. doi:10.1080/21541248.2017.1405773
52. Sinha S, Hoshino D, Hong NH, et al. Cortactin promotes exosome secretion by controlling branched actin dynamics. J Cell Biol. 2016;214:197–213. doi:10.1083/jcb.201601025
53. Parray ZA. A review on evolution, structural characteristics, interactions, and regulation of the membrane transport protein: the family of Rab proteins. Int J Biol Macromol. 2025;296:139828. doi:10.1016/j.ijbiomac.2025.139828
54. Ostrowski M, Carmo NB, Krumeich S, et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol. 2010;12:19–30. doi:10.1038/ncb2000
55. Hsu C, Morohashi Y, Yoshimura S, et al. Regulation of exosome secretion by Rab35 and its GTPase-activating proteins TBC1D10A-C. J Cell Biol. 2010;189:223–232. doi:10.1083/jcb.200911018
56. Savina A, Vidal M, Colombo MI. The exosome pathway in K562 cells is regulated by Rab11. J Cell Sci. 2002;115:2505–2515. doi:10.1242/jcs.115.12.2505
57. Villarroya-Beltri C, Baixauli F, Mittelbrunn M, et al. ISGylation controls exosome secretion by promoting lysosomal degradation of MVB proteins. Nat Commun. 2016;7:13588. doi:10.1038/ncomms13588
58. Mulcahy LA, Pink RC, Carter DR. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles. 2014;3. doi:10.3402/jev.v3.24641
59. Somiya M. Where does the cargo go?: solutions to provide experimental support for the “extracellular vesicle cargo transfer hypothesis”. J Cell Commun Signal. 2020;14(2):135–146. doi:10.1007/s12079-020-00552-9
60. Polanco JC, Hand GR, Briner A, Li C, Götz J. Exosomes induce endolysosomal permeabilization as a gateway by which exosomal tau seeds escape into the cytosol. Acta Neuropathol. 2021;141(2):235–256. doi:10.1007/s00401-020-02254-3
61. Goh CY, Wyse C, Ho M, et al. Advances in extracellular vesicle nanotechnology for precision theranostics. Cancer Lett. 2020;473:90–97. doi:10.1016/j.canlet.2019.12.046
62. Shrivastava S, Morris KV. The multifunctionality of exosomes; from the garbage bin of the cell to a next generation gene and cellular therapy. Genes. 2021;12(2):173. doi:10.3390/genes12020173
63. Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med. 1996;183:1161–1172. doi:10.1084/jem.183.3.1161
64. Tian T, Zhu YL, Zhou YY, et al. Exosome uptake through clathrin-mediated endocytosis and macropinocytosis and mediating miR-21 delivery. J Biol Chem. 2014;289:22258–22267. doi:10.1074/jbc.M114.588046
65. Dini L, Tacconi S, Carata E, Tata AM, Vergallo C, Panzarini E. Microvesicles and exosomes in metabolic diseases and inflammation. Cytokine Growth Factor Rev. 2020;51:27–39. doi:10.1016/j.cytogfr.2019.12.008
66. Guay C, Regazzi R. Exosomes as new players in metabolic organ cross-talk. Diabetes Obes Metab. 2017;19(Suppl 1):137–146. doi:10.1111/dom.13027
67. Machtinger R, Laurent LC, Baccarelli AA. Extracellular vesicles: roles in gamete maturation, fertilization and embryo implantation. Hum Reprod Update. 2016;22:182–193. doi:10.1093/humupd/dmv055
68. Sullivan R, Saez F, Girouard J, Frenette G. Role of exosomes in sperm maturation during the transit along the male reproductive tract. Blood Cells Mol Dis. 2005;35:1–10. doi:10.1016/j.bcmd.2005.03.005
69. Ali W, Deng K, Bian Y, Liu Z, Zou H. Spectacular role of epididymis and bio-active cargo of nano-scale exosome in sperm maturation: a review. Biomed Pharmacother. 2023;164:114889. doi:10.1016/j.biopha.2023.114889
70. Chen C, Zhang Z, Gu X, Sheng X, Xiao L, Wang X. Exosomes: new regulators of reproductive development. Mater Today Bio. 2023;19:100608. doi:10.1016/j.mtbio.2023.100608
71. Liu WM, Cheng RR, Niu ZR, et al. Let-7 derived from endometrial extracellular vesicles is an important inducer of embryonic diapause in mice. Sci Adv. 2020:6. doi:10.1126/sciadv.aaz7070
72. Czernek L, Düchler M. Exosomes as messengers between mother and fetus in pregnancy. Int J Mol Sci. 2020;21:4264. doi:10.3390/ijms21124264
73. Liu S, Fan M, Xu JX, et al. Exosomes derived from bone-marrow mesenchymal stem cells alleviate cognitive decline in AD-like mice by improving BDNF-related neuropathology. J Neuroinflammation. 2022;19:35. doi:10.1186/s12974-022-02393-2
74. Tian Y, Wang TS, Bu H, Shao G, Zhang W, Zhang L. Role of exosomal miR-223 in chronic skeletal muscle inflammation. Orthop Surg. 2022;14:644–651. doi:10.1111/os.13232
75. Deng ZB, Poliakov A, Hardy RW, et al. Adipose tissue exosome-like vesicles mediate activation of macrophage-induced insulin resistance. Diabetes. 2009;58:2498–2505. doi:10.2337/db09-0216
76. Rezaie J, Etemadi T, Feghhi M. The distinct roles of exosomes in innate immune responses and therapeutic applications in cancer. Eur J Pharmacol. 2022;933:175292. doi:10.1016/j.ejphar.2022.175292
77. Xia J, Miao Y, Wang X, Huang X, Dai J. Recent progress of dendritic cell-derived exosomes (Dex) as an anti-cancer nanovaccine. Biomed Pharmacother. 2022;152:113250. doi:10.1016/j.biopha.2022.113250
78. Chen W, Wang J, Shao C, et al. Efficient induction of antitumor T cell immunity by exosomes derived from heat-shocked lymphoma cells. Eur J Immunol. 2006;36:1598–1607. doi:10.1002/eji.200535501
79. Di Pace AL, Tumino N, Besi F, et al. Characterization of human NK cell-derived exosomes: role of DNAM1 receptor in exosome-mediated cytotoxicity against tumor. Cancers. 2020;12:661. doi:10.3390/cancers12030661
80. Skokos D, Botros HG, Demeure C, et al. Mast cell-derived exosomes induce phenotypic and functional maturation of dendritic cells and elicit specific immune responses in vivo. J Immunol. 2003;170:3037–3045. doi:10.4049/jimmunol.170.6.3037
81. Zhao S, Mi Y, Guan B, et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J Hematol Oncol. 2020;13:156. doi:10.1186/s13045-020-00991-2
82. Hanjani NA, Esmaelizad N, Zanganeh S, et al. Emerging role of exosomes as biomarkers in cancer treatment and diagnosis. Crit Rev Oncol Hematol. 2022;169:103565. doi:10.1016/j.critrevonc.2021.103565
83. Melo SA, Luecke LB, Kahlert C, et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature. 2015;523:177–182. doi:10.1038/nature14581
84. Wang X, Huang J, Chen W, Li G, Li Z, Lei J. The updated role of exosomal proteins in the diagnosis, prognosis, and treatment of cancer. Exp Mol Med. 2022;54:1390–1400. doi:10.1038/s12276-022-00855-4
85. Wu Q, Fu S, Xiao H, et al. Advances in extracellular vesicle nanotechnology for precision theranostics. Adv Sci. 2023;10(3):e2204814. doi:10.1002/advs.202204814
86. Wan S, Liu W, Wu Q, et al. Nanobiohybrid extracellular vesicle nanoreactor with improving metabolical activity for biocatalytic therapy. ACS Nano. 2024;18(47):32899–32909. doi:10.1021/acsnano.4c12458
87. Hellewell AL, Adams JC. Insider trading: extracellular matrix proteins and their non-canonical intracellular roles. Bioessays. 2016;38(1):77–88. doi:10.1002/bies.201500103
88. Karampoga A, Tzaferi K, Koutsakis C, Kyriakopoulou K, Karamanos NK. Exosomes and the extracellular matrix: a dynamic interplay in cancer progression. Int J Dev Biol. 2022;66(1–2–3):97–102. doi:10.1387/ijdb.210120nk
89. Lenzini S, Debnath K, Joshi JC, et al. Cell-matrix interactions regulate functional extracellular vesicle secretion from mesenchymal stromal cells. ACS Nano. 2021;15(11):17439–17452. doi:10.1021/acsnano.1c03231
90. Hoshino D, Kirkbride KC, Costello K, et al. Exosome secretion is enhanced by invadopodia and drives invasive behavior. Cell Rep. 2013;5(5):1159–1168. doi:10.1016/j.celrep.2013.10.050
91. Tamboli IY, Barth E, Christian L, et al. Statins promote the degradation of extracellular amyloid {beta}-peptide by microglia via stimulation of exosome-associated insulin-degrading enzyme (IDE) secretion. J Biol Chem. 2010;285(48):37405–37414. doi:10.1074/jbc.M110.149468
92. Asrani SK, Devarbhavi H, Eaton J, Kamath PS. Burden of liver diseases in the world. J Hepatol. 2019;70:151–171. doi:10.1016/j.jhep.2018.09.014
93. Feldman AG, Sokol RJ. Neonatal cholestasis: updates on diagnostics, therapeutics, and prevention. Neoreviews. 2021;22:e819–e836. doi:10.1542/neo.22-12-e819
94. García-Romero CS, Guzman C, Cervantes A, Cerbón M. Liver disease in pregnancy: medical aspects and their implications for mother and child. Ann Hepatol. 2019;18:553–562. doi:10.1016/j.aohep.2019.04.009
95. Nassir F. NAFLD: mechanisms, treatments, and biomarkers. Biomolecules. 2022;12(6). doi:10.3390/biom12060824
96. Hochreuter MY, Dall M, Treebak JT, Barrès R. MicroRNAs in non-alcoholic fatty liver disease: progress and perspectives. Mol Metab. 2022;65(101581):101581. doi:10.1016/j.molmet.2022.101581
97. Fu Y, Urban DJ, Nani RR, et al. Glypican-3-specific antibody drug conjugates targeting hepatocellular carcinoma. Hepatology. 2019;70:563–576. doi:10.1002/hep.30326
98. Kim JS, Eom JY, Kim HW, et al. Hemp sprout-derived exosome-like nanovesicles as hepatoprotective agents attenuate liver fibrosis. Biomater Sci. 2024;12:5361–5371. doi:10.1039/d4bm00812j
99. Zou J, Song Q, Shaw PC, Wu Y, Zuo Z, Yu R. Tangerine peel-derived exosome-like nanovesicles alleviate hepatic steatosis induced by type 2 diabetes: evidenced by regulating lipid metabolism and intestinal microflora. Int J Nanomed. 2024;19:10023–10043. doi:10.2147/ijn.S478589
100. El-Kattawy AM, Algezawy O, Alfaifi MY, et al. Therapeutic potential of camel milk exosomes against HepaRG cells with potent apoptotic, anti-inflammatory, and anti-angiogenesis effects for colostrum exosomes. Biomed Pharmacother. 2021;143:112220. doi:10.1016/j.biopha.2021.112220
101. Wang C, Liu J, Yan Y, Tan Y. Role of exosomes in chronic liver disease development and their potential clinical applications. J Immunol Res. 2022;2022:1695802. doi:10.1155/2022/1695802
102. Shao M, Xu Q, Wu Z, et al. Exosomes derived from human umbilical cord mesenchymal stem cells ameliorate IL-6-induced acute liver injury through miR-455-3p. Stem Cell Res Ther. 2020;11:37. doi:10.1186/s13287-020-1550-0
103. Liu M, Zhu Y, Li Z, et al. Exosomes from liver progenitor cells carrying JAG1 activate notch signaling to promote liver regeneration in PVL rats. Cell Death Dis. 2025;16(1):609. doi:10.1038/s41419-025-07925-1
104. Liu J, Tang M, Li Q, Li Q, Dai Y, Zhou H. ATG2B upregulated in LPS-stimulated BMSCs-derived exosomes attenuates septic liver injury by inhibiting macrophage STING signaling. Int Immunopharmacol. 2023;117:109931. doi:10.1016/j.intimp.2023.109931
105. Yu L, Xue J, Wu Y, Zhou H. Therapeutic effect of exosomes derived from hepatocyte-growth-factor-overexpressing adipose mesenchymal stem cells on liver injury. Folia Histochem Cytobiol. 2023;61(3):160–171. doi:10.5603/fhc.95291
106. Cheng L, Yu P, Li F, et al. Human umbilical cord-derived mesenchymal stem cell-exosomal miR-627-5p ameliorates non-alcoholic fatty liver disease by repressing FTO expression. Hum Cell. 2021;34:1697–1708. doi:10.1007/s13577-021-00593-1
107. Liu B, Nguyen PL, Yu H, et al. Honey vesicle-like nanoparticles protect aged liver from non-alcoholic steatohepatitis. Acta Pharm Sin B. 2024;14:3661–3679. doi:10.1016/j.apsb.2024.05.002
108. Niu Q, Wang T, Wang Z, et al. Adipose-derived mesenchymal stem cell-secreted extracellular vesicles alleviate non-alcoholic fatty liver disease via delivering miR-223-3p. Adipocyte. 2022;11:572–587. doi:10.1080/21623945.2022.2098583
109. Jiang W, Zeng Q, Liu CH, et al. Huc-MSCs-derived exosomes alleviate non-alcoholic steatohepatitis by regulating macrophages polarization through miR-24-3p/STING axis. Stem Cell Res Ther. 2025;16:74. doi:10.1186/s13287-025-04197-6
110. Tian S, Zhou X, Zhang M, et al. Mesenchymal stem cell-derived exosomes protect against liver fibrosis via delivering miR-148a to target KLF6/STAT3 pathway in macrophages. Stem Cell Res Ther. 2022;13:330. doi:10.1186/s13287-022-03010-y
111. Cheng F, Yang F, Wang Y, Zhou J, Qian H, Yan Y. Mesenchymal stem cell-derived exosomal miR-27b-3p alleviates liver fibrosis via downregulating YAP/LOXL2 pathway. J Nanobiotechnology. 2023;21:195. doi:10.1186/s12951-023-01942-y
112. Wang Z, Yuan M, Yao L, et al. Exosomal miR-499a-5p from human umbilical cord mesenchymal stem cells attenuates liver fibrosis via targeting ETS1/GPX4-mediated ferroptosis in hepatic stellate cells. J Nanobiotechnology. 2025;23:222. doi:10.1186/s12951-025-03291-4
113. Niknam B, Baghaei K, Mahmoud Hashemi S, Hatami B, Reza Zali M, Amani D. Human wharton’s jelly mesenchymal stem cells derived-exosomes enriched by miR-124 promote an anti-fibrotic response in an experimental model of liver fibrosis. Int Immunopharmacol. 2023;119:110294. doi:10.1016/j.intimp.2023.110294
114. Hu Z, Zhao Y, Jiang J, et al. Exosome-derived miR-142-5p from liver stem cells improves the progression of liver fibrosis by regulating macrophage polarization through CTSB. Environ Toxicol. 2023;38:1860–1873. doi:10.1002/tox.23813
115. Zhang T, Jing J, Liang Y, et al. Resveratrol-stimulated macrophage exosomes delivering lncRNA Snhg6 inhibit liver fibrosis by modulating the NF-κB pathway. Genomics. 2025;117(3):111043. doi:10.1016/j.ygeno.2025.111043
116. Ma J, Li Y, Chen M, et al. hMSCs-derived exosome circCDK13 inhibits liver fibrosis by regulating the expression of MFGE8 through miR-17-5p/KAT2B. Cell Biol Toxicol. 2023;39(2):1–22. doi:10.1007/s10565-022-09714-4
117. Ma L, Wei J, Zeng Y, et al. Mesenchymal stem cell-originated exosomal circDIDO1 suppresses hepatic stellate cell activation by miR-141-3p/PTEN/AKT pathway in human liver fibrosis. Drug Deliv. 2022;29(1):440–453. doi:10.1080/10717544.2022.2030428
118. Liu Y, Lou G, Li A, et al. AMSC-derived exosomes alleviate lipopolysaccharide/d-galactosamine-induced acute liver failure by miR-17-mediated reduction of TXNIP/NLRP3 inflammasome activation in macrophages. EBioMedicine. 2018;36:140–150. doi:10.1016/j.ebiom.2018.08.054
119. Ma YS, Liu JB, Lin L, et al. Exosomal microRNA-15a from mesenchymal stem cells impedes hepatocellular carcinoma progression via downregulation of SALL4. Cell Death Discov. 2021;7:224. doi:10.1038/s41420-021-00611-z
120. Xu Y, Lai Y, Cao L, et al. Human umbilical cord mesenchymal stem cells-derived exosomal microRNA-451a represses epithelial-mesenchymal transition of hepatocellular carcinoma cells by inhibiting ADAM10. RNA Biol. 2021;18:1408–1423. doi:10.1080/15476286.2020.1851540
121. Cheng Z, Lei Z, Yang P, et al. Exosome-transmitted p120-catenin suppresses hepatocellular carcinoma progression via STAT3 pathways. Mol, Carcinog. 2019;58(8):1389–1399. doi:10.1002/mc.23022
122. Dong FL, Xu ZZ, Wang YQ, Li T, Wang X, Li J. Exosome-derived circUPF2 enhances resistance to targeted therapy by redeploying ferroptosis sensitivity in hepatocellular carcinoma. J Nanobiotechnology. 2024;22(1):298. doi:10.1186/s12951-024-02582-6
123. Li Z, Li Y, Ouyang Q, Li X, Huang J. Exosome-derived GTF2H2 from Huh7 cells can inhibit endothelial cell viability, migration, tube formation, and permeability. Tissue Cell. 2022;79:101922. doi:10.1016/j.tice.2022.101922
124. Basthi Mohan P, Rajpurohit S, Musunuri B, Bhat G, Lochan R, Shetty S. Exosomes in chronic liver disease. Clin Chim Acta. 2023;540:117215. doi:10.1016/j.cca.2022.117215
125. Tong L, Yan C, Wang M, Yang J, Wang H, Wang Y. Prognostic value of serum exosomal AHCY expression in hepatitis B-induced liver cirrhosis. Front Med. 2021;8:777452. doi:10.3389/fmed.2021.777452
126. Wang Y, Pei L, Yue Z, Jia M, Wang H, Cao LL. The potential of serum exosomal hsa_circ_0028861 as the novel diagnostic biomarker of HBV-derived hepatocellular cancer. Front Genet. 2021;12:703205. doi:10.3389/fgene.2021.703205
127. Xu W, Yu M, Wu Y, et al. Plasma-derived exosomal SncRNA as a promising diagnostic biomarker for early detection of HBV-related acute-on-chronic liver failure. Front Cell Infect Microbiol. 2022;12:923300. doi:10.3389/fcimb.2022.923300
128. Jiao X, Fan Z, Chen H, et al. Serum and exosomal miR-122 and miR-199a as a biomarker to predict therapeutic efficacy of hepatitis C patients. J Med Virol. 2017;89:1597–1605. doi:10.1002/jmv.24829
129. Conde-Vancells J, Rodriguez-Suarez E, Gonzalez E, et al. Candidate biomarkers in exosome-like vesicles purified from rat and mouse urine samples. Proteomics Clin Appl. 2010;4:416–425. doi:10.1002/prca.200900103
130. Zhang J, Li D, Zhang R, Gao P, Peng R, Li J. The miR-21 potential of serving as a biomarker for liver diseases in clinical practice. Biochem Soc Trans. 2020;48:2295–2305. doi:10.1042/bst20200653
131. Lin Y, Ding D, Huang Q, et al. Downregulation of miR-192 causes hepatic steatosis and lipid accumulation by inducing SREBF1: novel mechanism for bisphenol a-triggered non-alcoholic fatty liver disease. Biochim Biophys Acta Mol Cell Biol Lipids. 2017;1862:869–882. doi:10.1016/j.bbalip.2017.05.001
132. Zhang W, Zhang J, Shi H, Liu F, Yu H, Shi H. Exosome GLUT1 derived from hepatocyte identifies the risk of non-alcoholic steatohepatitis and fibrosis. Hepatol Int. 2023;17:1170–1181. doi:10.1007/s12072-023-10520-1
133. Bala S, Petrasek J, Mundkur S, et al. Circulating microRNAs in exosomes indicate hepatocyte injury and inflammation in alcoholic, drug-induced, and inflammatory liver diseases. Hepatology. 2012;56:1946–1957. doi:10.1002/hep.25873
134. Momen-Heravi F, Saha B, Kodys K, Catalano D, Satishchandran A, Szabo G. Increased number of circulating exosomes and their microRNA cargos are potential novel biomarkers in alcoholic hepatitis. J Transl Med. 2015;13:261. doi:10.1186/s12967-015-0623-9
135. Chen L, Yao X, Yao H, Ji Q, Ding G, Liu X. Exosomal miR-103-3p from LPS-activated THP-1 macrophage contributes to the activation of hepatic stellate cells. FASEB j. 2020;34:5178–5192. doi:10.1096/fj.201902307RRR
136. El-Ahwany E, Nagy F, Zoheiry M, et al. Circulating miRNAs as predictor markers for activation of hepatic stellate cells and progression of HCV-induced liver fibrosis. Electron Physician. 2016;8:1804–1810. doi:10.19082/1804
137. Niu LJ, Zhang YM, Huang T, Sun XF, Luo SX. Exosomal microRNA-155 as a biomarker for hepatic fibrosis diagnosis and progression. Ann Transl Med. 2021;9:137. doi:10.21037/atm-20-7787
138. Gonzalez E, Azkargorta M, Garcia-Vallicrosa C, et al. Could protein content of urinary extracellular vesicles be useful to detect cirrhosis in alcoholic liver disease? Int J Biol Sci. 2021;17:1864–1877. doi:10.7150/ijbs.59725
139. Fründt T, Krause L, Hussey E, et al. Diagnostic and prognostic value of miR-16, miR-146a, miR-192 and miR-221 in exosomes of hepatocellular carcinoma and liver cirrhosis patients. Cancers. 2021;13. doi:10.3390/cancers13102484
140. Xu H, Chen Y, Dong X, Wang X. Serum exosomal long noncoding RNAs ENSG00000258332.1 and LINC00635 for the diagnosis and prognosis of hepatocellular carcinoma. Cancer Epidemiol Biomarkers Prev. 2018;27:710–716. doi:10.1158/1055-9965.Epi-17-0770
141. Sohn W, Kim J, Kang SH, et al. Serum exosomal microRNAs as novel biomarkers for hepatocellular carcinoma. Exp Mol Med. 2015;47:e184.
142. Ning S, Liu H, Gao B, et al. miR-155, miR-96 and miR-99a as potential diagnostic and prognostic tools for the clinical management of hepatocellular carcinoma. Oncol Lett. 2019;18:3381–3387. doi:10.3892/ol.2019.10606
143. Ying SQ, Cao Y, Zhou ZK, et al. Hepatocyte-derived tissue extracellular vesicles safeguard liver regeneration and support regenerative therapy. J Nanobiotechnology. 2024;22(1):521. doi:10.1186/s12951-024-02790-0
144. Liu X, Tan S, Liu H, et al. Hepatocyte-derived MASP1-enriched small extracellular vesicles activate HSCs to promote liver fibrosis. Hepatology. 2023;77(4):1181–1197. doi:10.1002/hep.32662
145. Lotfy A, AboQuella NM, Wang H. Mesenchymal stromal/stem cell (MSC)-derived exosomes in clinical trials. Stem Cell Res Ther. 2023;14:66. doi:10.1186/s13287-023-03287-7
146. Wang Z, Zhong D, Yan T, et al. Stem cells from human exfoliated deciduous teeth-derived exosomes for the treatment of acute liver injury and liver fibrosis. ACS Appl Mater Interfaces. 2025;17(12):17948–17964. doi:10.1021/acsami.4c19748
147. Wu Y, Wang L, Deng Z, et al. Human umbilical cord-derived mesenchymal stem cell exosomes ameliorate hepatocyte ferroptosis and sepsis-associated liver injury by restoring iron homeostasis in hepatocytes. J Nanobiotechnology. 2026. doi:10.1186/s12951-026-04270-z
148. Li L, Dong Y, Liu X, Wang M. Mangiferin for the management of liver diseases: a review. Foods. 2023;12. doi:10.3390/foods12132469
149. Li M, Luo Q, Tao Y, Sun X, Liu C. Pharmacotherapies for drug-induced liver injury: a current literature review. Front Pharmacol. 2021;12:806249. doi:10.3389/fphar.2021.806249
150. Fisher K, Vuppalanchi R, Saxena R. Drug-induced liver injury. Arch Pathol Lab Med. 2015;139:876–887. doi:10.5858/arpa.2014-0214-RA
151. Yuan R, Tao X, Liang S, et al. Protective effect of acidic polysaccharide from schisandra chinensis on acute ethanol-induced liver injury through reducing CYP2E1-dependent oxidative stress. Biomed Pharmacother. 2018;99:537–542. doi:10.1016/j.biopha.2018.01.079
152. Iorga A, Dara L. Cell death in drug-induced liver injury. Adv Pharmacol. 2019;85:31–74. doi:10.1016/bs.apha.2019.01.006
153. Wei H, Zhao T, Liu X, et al. Mechanism of action of dihydroquercetin in the prevention and therapy of experimental liver injury. Molecules. 2024;29(15):3537. doi:10.3390/molecules29153537
154. Tan CY, Lai RC, Wong W, Dan YY, Lim SK, Ho HK. Mesenchymal stem cell-derived exosomes promote hepatic regeneration in drug-induced liver injury models. Stem Cell Res Ther. 2014;5:76. doi:10.1186/scrt465
155. Wu HY, Zhang XC, Jia BB, et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate Acetaminophen-induced acute liver failure through activating ERK and IGF-1R/PI3K/AKT signaling pathway. J Pharmacol Sci. 2021;147:143–155. doi:10.1016/j.jphs.2021.06.008
156. Wang Y, Li H, Jiang S, et al. The glycolytic enzyme PFKFB3 drives kidney fibrosis through promoting histone lactylation-mediated NF-κB family activation. Kidney Int. 2024;106(2):226–240. doi:10.1016/j.kint.2024.04.016
157. Wang M, Shen G, Xu L, et al. IL-1 receptor like 1 protects against alcoholic liver injury by limiting NF-κB activation in hepatic macrophages. J Hepatol. 2017;21:S0168–8278(17)32263–8. doi:10.1016/j.jhep.2017.08.023
158. Tian Y, Zhang J, Jia Z, et al. Biomimetic mineralized mesenchymal stem cell-derived exosomes for dual modulation of ferroptosis and lactic acid-driven inflammation in acute liver injury therapy. J Colloid Interface Sci. 2025;687:489–506. doi:10.1016/j.jcis.2025.02.078
159. Rocco A, Compare D, Angrisani D, Sanduzzi Zamparelli M, Nardone G. Alcoholic disease: liver and beyond. World J Gastroenterol. 2014;20:14652–14659. doi:10.3748/wjg.v20.i40.14652
160. Yan C, Hu W, Tu J, Li J, Liang Q, Han S. Pathogenic mechanisms and regulatory factors involved in alcoholic liver disease. J Transl Med. 2023;21:300. doi:10.1186/s12967-023-04166-8
161. Hurley TD, Edenberg HJ. Genes encoding enzymes involved in ethanol metabolism. Alcohol Res. 2012;34:339–344.
162. Chen Z, Tian R, She Z, Cai J, Li H. Role of oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free Radic Biol Med. 2020;152:116–141. doi:10.1016/j.freeradbiomed.2020.02.025
163. Ji C, Chan C, Kaplowitz N. Predominant role of sterol response element binding proteins (SREBP) lipogenic pathways in hepatic steatosis in the murine intragastric ethanol feeding model. J Hepatol. 2006;45:717–724. doi:10.1016/j.jhep.2006.05.009
164. You M, Matsumoto M, Pacold CM, Cho WK, Crabb DW. The role of AMP-activated protein kinase in the action of ethanol in the liver. Gastroenterology. 2004;127:1798–1808. doi:10.1053/j.gastro.2004.09.049
165. Kong L, Chen J, Ji X, et al. Alcoholic fatty liver disease inhibited the co-expression of Fmo5 and PPARα to activate the NF-κB signaling pathway, thereby reducing liver injury via inducing gut microbiota disturbance. J Exp Clin Cancer Res. 2021;40:18. doi:10.1186/s13046-020-01782-w
166. Kim JS, Kim DH, Gil MC, et al. Pomegranate-derived exosome-like nanovesicles alleviate binge alcohol-induced leaky gut and liver injury. J Med Food. 2023;26:739–748. doi:10.1089/jmf.2023.K.0060
167. Zhang W, Song Q, Bi X, et al. Preparation of pueraria lobata root-derived exosome-like nanovesicles and evaluation of their effects on mitigating alcoholic intoxication and promoting alcohol metabolism in mice. Int J Nanomed. 2024;19:4907–4921. doi:10.2147/ijn.S462602
168. Younossi Z, Anstee QM, Marietti M, et al. Global burden of NAFLD and NASH: trends, predictions, risk factors and prevention. Nat Rev Gastroenterol Hepatol. 2018;15:11–20. doi:10.1038/nrgastro.2017.109
169. Cobbina E, Akhlaghi F. Non-alcoholic fatty liver disease (NAFLD) - pathogenesis, classification, and effect on drug metabolizing enzymes and transporters. Drug Metab Rev. 2017;49:197–211. doi:10.1080/03602532.2017.1293683
170. Guo X, Yin X, Liu Z, Wang J. Non-alcoholic fatty liver disease (NAFLD) pathogenesis and natural products for prevention and treatment. Int J Mol Sci. 2022;23:15489. doi:10.3390/ijms232415489
171. Wang Z, Ye M, Zhang XJ, et al. Impact of NAFLD and its pharmacotherapy on lipid profile and CVD. Atherosclerosis. 2022;355:30–44. doi:10.1016/j.atherosclerosis.2022.07.010
172. Petersen MC, Shulman GI. Roles of diacylglycerols and ceramides in hepatic insulin resistance. Trends Pharmacol Sci. 2017;38(7):649–665. doi:10.1016/j.tips.2017.04.004
173. Oberhauser L, Maechler P. Lipid-induced adaptations of the pancreatic beta-cell to glucotoxic conditions sustain insulin secretion. Int J Mol Sci. 2021;23(1):324. doi:10.3390/ijms23010324
174. Cui A, Xiao P, Fan Z, et al. Causal association of NAFLD with osteoporosis, fracture and falling risk: a bidirectional mendelian randomization study. Front Endocrinol. 2023;14:1215790. doi:10.3389/fendo.2023.1215790
175. Friedman SL, Neuschwander-Tetri BA, Rinella M, Sanyal AJ. Mechanisms of NAFLD development and therapeutic strategies. Nat Med. 2018;24:908–922. doi:10.1038/s41591-018-0104-9
176. Wei S, Wang L, Evans PC, Xu S. NAFLD and NASH: etiology, targets and emerging therapies. Drug Discov Today. 2024;29:103910. doi:10.1016/j.drudis.2024.103910
177. Yang F, Wu Y, Chen Y, et al. Human umbilical cord mesenchymal stem cell-derived exosomes ameliorate liver steatosis by promoting fatty acid oxidation and reducing fatty acid synthesis. JHEP Rep. 2023;5:100746. doi:10.1016/j.jhepr.2023.100746
178. Nie YF, Shang JM, Liu DQ, et al. Apical papilla stem cell-derived exosomes regulate lipid metabolism and alleviate inflammation in the MCD-induced mouse NASH model. Biochem Pharmacol. 2024;222:116073. doi:10.1016/j.bcp.2024.116073
179. Zhao H, Shang Q, Pan Z, et al. Exosomes from adipose-derived stem cells attenuate adipose inflammation and obesity through polarizing M2 macrophages and beiging in white adipose tissue. Diabetes. 2018;67:235–247. doi:10.2337/db17-0356
180. Kang Y, Song Y, Luo Y, et al. Exosomes derived from human umbilical cord mesenchymal stem cells ameliorate experimental non-alcoholic steatohepatitis via Nrf2/NQO-1 pathway. Free Radic Biol Med. 2022;192:25–36. doi:10.1016/j.freeradbiomed.2022.08.037
181. Bellezza I, Giambanco I, Minelli A, Donato R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim Biophys Acta Mol Cell Res. 2018;1865:721–733. doi:10.1016/j.bbamcr.2018.02.010
182. Du X, Li H, Han X, Ma W. Mesenchymal stem cells-derived exosomal miR-24-3p ameliorates non-alcohol fatty liver disease by targeting Keap-1. Biochem Biophys Res Commun. 2022;637:331–340. doi:10.1016/j.bbrc.2022.11.012
183. Moayedfard Z, Bagheri Lankarani K, Alizadeh AA, et al. The ameliorative effect of adipose-derived mesenchymal stem cells and their exosomes in non-alcoholic steatohepatitis by simultaneously enhancing autophagic flux and suppressing endoplasmic reticulum stress. Iran J Med Sci. 2025;50(5):334–350. doi:10.30476/ijms.2024.103376.3660
184. Aydın MM, Akçalı KC. Liver fibrosis. Turk J Gastroenterol. 2018;29:14–21. doi:10.5152/tjg.2018.17330
185. Bataller R, Brenner DA. Liver fibrosis. J Clin Invest. 2005;115:209–218. doi:10.1172/jci24282
186. Dewidar B, Meyer C, Dooley S, Meindl-Beinker AN. TGF-β in hepatic stellate cell activation and liver fibrogenesis-updated 2019. Cells. 2019;8. doi:10.3390/cells8111419
187. Roehlen N, Crouchet E, Baumert TF. Liver fibrosis: mechanistic concepts and therapeutic perspectives. Cells. 2020;9. doi:10.3390/cells9040875
188. Parola M, Pinzani M. Liver fibrosis: pathophysiology, pathogenetic targets and clinical issues. Mol Aspects Med. 2019;65:37–55. doi:10.1016/j.mam.2018.09.002
189. Marra F, Bertolani C. Adipokines in liver diseases. Hepatology. 2009;50:957–969. doi:10.1002/hep.23046
190. Gan L, Zheng L, Yao L, et al. Exosomes from adipose-derived mesenchymal stem cells improve liver fibrosis by regulating the miR-20a-5p/TGFBR2 axis to affect the p38 MAPK/NF-κB pathway. Cytokine. 2023;172:156386. doi:10.1016/j.cyto.2023.156386
191. Wu B, Feng J, Guo J, et al. ADSCs-derived exosomes ameliorate hepatic fibrosis by suppressing stellate cell activation and remodeling hepatocellular glutamine synthetase-mediated glutamine and ammonia homeostasis. Stem Cell Res Ther. 2022;13:494. doi:10.1186/s13287-022-03049-x
192. Zhang Z, Shang J, Yang Q, et al. Exosomes derived from human adipose mesenchymal stem cells ameliorate hepatic fibrosis by inhibiting PI3K/Akt/mTOR pathway and remodeling choline metabolism. J Nanobiotechnology. 2023;21:29. doi:10.1186/s12951-023-01788-4
193. Zong R, Zheng Y, Yan Y, et al. Mesenchymal stem cells-derived exosomes alleviate liver fibrosis by targeting Hedgehog/SMO signaling. Hepatol Int. 2024;18:1781–1791. doi:10.1007/s12072-024-10717-y
194. Tan Y, Huang Y, Mei R, et al. HucMSC-derived exosomes delivered BECN1 induces ferroptosis of hepatic stellate cells via regulating the xCT/GPX4 axis. Cell Death Dis. 2022;13:319. doi:10.1038/s41419-022-04764-2
195. Rong X, Liu J, Yao X, Jiang T, Wang Y, Xie F. Human bone marrow mesenchymal stem cells-derived exosomes alleviate liver fibrosis through the Wnt/β-catenin pathway. Stem Cell Res Ther. 2019;10:98.
196. Liu Y, Zheng Y, Yang Y, et al. Exosomes in liver fibrosis: the role of modulating hepatic stellate cells and immune cells, and prospects for clinical applications. Front Immunol. 2023;14:1133297. doi:10.3389/fimmu.2023.1133297
197. Qu Y, Zhang Q, Cai X, et al. Exosomes derived from miR-181-5p-modified adipose-derived mesenchymal stem cells prevent liver fibrosis via autophagy activation. J Cell Mol Med. 2017;21:2491–2502. doi:10.1111/jcmm.13170
198. Zheng W, Bian S, Qiu S, et al. Placenta mesenchymal stem cell-derived extracellular vesicles alleviate liver fibrosis by inactivating hepatic stellate cells through a miR-378c/SKP2 axis. Inflamm Regen. 2023;43:47. doi:10.1186/s41232-023-00297-z
199. Wang N, Li X, Zhong Z, et al. 3D hESC exosomes enriched with miR-6766-3p ameliorates liver fibrosis by attenuating activated stellate cells through targeting the TGFβRII-SMADS pathway. J Nanobiotechnology. 2021;19:437. doi:10.1186/s12951-021-01138-2
200. Li L, Liu Y, Wang K, et al. Stem cell exosomes: new hope and future potential for relieving liver fibrosis. Clin Mol Hepatol. 2025;31(2):333–349. doi:10.3350/cmh.2024.0854
201. Barnett R. Liver cirrhosis. Lancet. 2018;392(10144):275. doi:10.1016/s0140-6736(18)31659-3
202. Starr SP, Raines D. Cirrhosis: diagnosis, management, and prevention. Am Fam Physician. 2011;84(12):1353–1359.
203. Juanola A, Pose E, Ginès P. Liver cirrhosis: ancient disease, new challenge. Med Clin. 2025;164:238–246. doi:10.1016/j.medcli.2024.11.002
204. Abd-Allah SH, Khamis T, Samy W, Alsemeh AE, Abdullah DM, Hussein S. Mesenchymal stem cells and their derived exosomes mitigated hepatic cirrhosis in rats by altering the expression of miR-23b and miR-221. Iran J Med Sci. 2024;49:724–740. doi:10.30476/ijms.2023.99524.3159
205. Mardpour S, Hassani SN, Mardpour S, et al. Extracellular vesicles derived from human embryonic stem cell-MSCs ameliorate cirrhosis in thioacetamide-induced chronic liver injury. J Cell Physiol. 2018;233:9330–9344. doi:10.1002/jcp.26413
206. Shi M, Li YY, Xu RN, et al. Mesenchymal stem cell therapy in decompensated liver cirrhosis: a long-term follow-up analysis of the randomized controlled clinical trial. Hepatol Int. 2021;15:1431–1441. doi:10.1007/s12072-021-10199-2
207. Lu W, Qu J, Yan L, et al. Efficacy and safety of mesenchymal stem cell therapy in liver cirrhosis: a systematic review and meta-analysis. Stem Cell Res Ther. 2023;14:301. doi:10.1186/s13287-023-03518-x
208. Jang YO, Kim YJ, Baik SK, et al. Histological improvement following administration of autologous bone marrow-derived mesenchymal stem cells for alcoholic cirrhosis: a pilot study. Liver Int. 2014;34:33–41. doi:10.1111/liv.12218
209. Wang Y, Kong Y, Du J, et al. Injection of human umbilical cord mesenchymal stem cells exosomes for the treatment of knee osteoarthritis: from preclinical to clinical research. J Transl Med. 2025;23(1):641. doi:10.1186/s12967-025-06623-y
210. Ye C, Li W, Li L, Zhang K. Glucocorticoid treatment strategies in liver failure. Front Immunol. 2022;13:846091. doi:10.3389/fimmu.2022.846091
211. Sarin SK, Choudhury A, Sharma MK, et al. Acute-on-chronic liver failure: consensus recommendations of the Asian Pacific association for the study of the liver (APASL): an update. Hepatol Int. 2019;13:353–390. doi:10.1007/s12072-019-09946-3
212. Wang X, Ning Q. Immune mediated liver failure. Excli J. 2014;13:1131–1144.
213. Br VK, Sarin SK. Acute-on-chronic liver failure: terminology, mechanisms and management. Clin Mol Hepatol. 2023;29:670–689. doi:10.3350/cmh.2022.0103
214. Jun JH, Kim JY, Choi JH, Lim JY, Kim K, Kim GJ. Exosomes from placenta-derived mesenchymal stem cells are involved in liver regeneration in hepatic failure induced by bile duct ligation. Stem Cells Int. 2020;2020:5485738. doi:10.1155/2020/5485738
215. Zhang S, Jiang L, Hu H, et al. Pretreatment of exosomes derived from hUCMSCs with TNF-α ameliorates acute liver failure by inhibiting the activation of NLRP3 in macrophage. Life Sci. 2020;246:117401. doi:10.1016/j.lfs.2020.117401
216. Liu J, Du S, Kong Q, et al. HSPA12A attenuates lipopolysaccharide-induced liver injury through inhibiting caspase-11-mediated hepatocyte pyroptosis via PGC-1α-dependent acyloxyacyl hydrolase expression. Cell Death Differ. 2020;27(9):2651–2667. doi:10.1038/s41418-020-0536-x
217. Xie D, Yu L, Wang Z, Qiu G, Ouyang S. Exosomes derived from human umbilical cord mesenchymal stem cells inhibit hepatocyte pyroptosis via miR-423-5p/ZBP1 in acute liver failure. Hum Cell. 2025;38:124. doi:10.1007/s13577-025-01248-1
218. Lin D, Chen H, Xiong J, et al. Mesenchymal stem cells exosomal let-7a-5p improve autophagic flux and alleviate liver injury in acute-on-chronic liver failure by promoting nuclear expression of TFEB. Cell Death Dis. 2022;13:865. doi:10.1038/s41419-022-05303-9
219. Chen X, Yang Z, Li M. Molecular regulatory mechanisms and diagnostic potential of dendritic cell-derived exosomes in liver transplantation: from immune tolerance induction to translational challenges. Front Immunol. 2025;16:1657956. doi:10.3389/fimmu.2025.1657956
220. Yao L, Hu X, Dai K, et al. Mesenchymal stromal cells: promising treatment for liver cirrhosis. Stem Cell Res Ther. 2022;13(1):308. doi:10.1186/s13287-022-03001-z
221. Cai Z, Zhang W, Yang F, et al. Immunosuppressive exosomes from TGF-β1 gene-modified dendritic cells attenuate Th17-mediated inflammatory autoimmune disease by inducing regulatory T cells. Cell Res. 2012;22(3):607–610. doi:10.1038/cr.2011.196
222. Wang J, Zhao X, Wan YY. Intricacies of TGF-β signaling in Treg and Th17 cell biology. Cell Mol Immunol. 2023;20(9):1002–1022. doi:10.1038/s41423-023-01036-7
223. Jung KW, Kang J, Kwon HM, et al. Effect of remote ischemic preconditioning conducted in living liver donors on postoperative liver function in donors and recipients following liver transplantation: a randomized clinical trial. Ann Surg. 2020;271(4):646–653. doi:10.1097/SLA.0000000000003498
224. Deng Z, Zeng W, Gao Y, et al. Mesenchymal stem cells prevent SLC39A14-dependent hepatocyte ferroptosis through exosomal miR-16-5p in liver graft. Adv Sci. 2025;12(6):e2411380. doi:10.1002/advs.202411380
225. Gilles H, Garbutt T, Landrum J. Hepatocellular carcinoma. Crit Care Nurs Clin North Am. 2022;34:289–301. doi:10.1016/j.cnc.2022.04.004
226. Foglia B, Turato C, Cannito S. Hepatocellular carcinoma: latest research in pathogenesis, detection and treatment. Int J Mol Sci. 2023;24(15):12224. doi:10.3390/ijms241512224
227. Park S, Davis AM, Pillai AA. Prevention, diagnosis, and treatment of hepatocellular carcinoma. JAMA. 2024;332(12):1013–1014. doi:10.1001/jama.2024.14101
228. Forner A, Reig M, Bruix J. Hepatocellular carcinoma. Lancet. 2018;391:1301–1314. doi:10.1016/s0140-6736(18)30010-2
229. Ko SF, Yip HK, Zhen YY, et al. Adipose-derived mesenchymal stem cell exosomes suppress hepatocellular carcinoma growth in a rat model: apparent diffusion coefficient, natural killer T-cell responses, and histopathological features. Stem Cells Int. 2015;2015:853506. doi:10.1155/2015/853506
230. Xu G, Ban K, Mu H, Wang B. Human umbilical cord mesenchymal stem cells-derived exosomal lncRNA FAM99B represses hepatocellular carcinoma cell malignancy. Mol Biotechnol. 2024;66:1389–1401. doi:10.1007/s12033-023-00795-y
231. Ding B, Lou W, Fan W, Pan J. Exosomal miR-374c-5p derived from mesenchymal stem cells suppresses epithelial-mesenchymal transition of hepatocellular carcinoma via the LIMK1-Wnt/β-catenin axis. Environ Toxicol. 2023;38:1038–1052. doi:10.1002/tox.23746
232. Lou G, Song X, Yang F, et al. Exosomes derived from miR-122-modified adipose tissue-derived MSCs increase chemosensitivity of hepatocellular carcinoma. J Hematol Oncol. 2015;8:122. doi:10.1186/s13045-015-0220-7
233. Lou G, Chen L, Xia C, et al. MiR-199a-modified exosomes from adipose tissue-derived mesenchymal stem cells improve hepatocellular carcinoma chemosensitivity through mTOR pathway. J Exp Clin Cancer Res. 2020;39:4. doi:10.1186/s13046-019-1512-5235
234. Liang L, Zhao L, Wang Y, Wang Y. Treatment for hepatocellular carcinoma is enhanced when norcantharidin is encapsulated in exosomes derived from bone marrow mesenchymal stem cells. Mol Pharm. 2021;18(3):1003–1013. doi:10.1021/acs.molpharmaceut.0c00976
235. Jeppesen DK, Zhang Q, Coffey RJ. Extracellular vesicles and nanoparticles at a glance. J Cell Sci. 2024;137(23):jcs260201. doi:10.1242/jcs.260201
236. Zhang H, Freitas D, Kim HS, et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat Cell Biol. 2018;20(3):332–343. doi:10.1038/s41556-018-0040-4
237. Spinella R, Sawhney R, Jalan R. Albumin in chronic liver disease: structure, functions and therapeutic implications. Hepatol Int. 2016;10(1):124–132. doi:10.1007/s12072-015-9665-6
238. Schleicher EM, Karbannek H, Weinmann-Menke J, et al. Effect of albumin treatment duration on response rates and outcomes in patients with cirrhosis and acute kidney injury. J Hepatol. 2025;83(3):682–691. doi:10.1016/j.jhep.2025.03.008
239. Allen RM, Michell DL, Cavnar AB, et al. LDL delivery of microbial small RNAs drives atherosclerosis through macrophage TLR8. Nat Cell Biol. 2022;24(12):1701–1713. doi:10.1038/s41556-022-01030-7
240. Mehta A, Shapiro MD. Apolipoproteins in vascular biology and atherosclerotic disease. Nat Rev Cardiol. 2022;19(3):168–179. doi:10.1038/s41569-021-00613-5
241. Steiner E, Holzmann K, Elbling L, Micksche M, Berger W. Cellular functions of vaults and their involvement in multidrug resistance. Curr Drug Targets. 2006;7(8):923–934. doi:10.2174/138945006778019345
242. Sun X, Lian Y, Tian T, Cui Z. Advancements in functional nanomaterials inspired by viral particles. Small. 2024;20(36):e2402980. doi:10.1002/smll.202402980
243. Biller SJ, Ryan MG, Li J, et al. Distinct horizontal gene transfer potential of extracellular vesicles versus viral-like particles in marine habitats. Nat Commun. 2025;16(1):2126. doi:10.1038/s41467-025-57276-w
244. Zhang Q, Jeppesen DK, Higginbotham JN, et al. Angiotensin-converting enzyme 2-containing small extracellular vesicles and exomeres bind the severe acute respiratory syndrome coronavirus 2 spike protein. Gastroenterology. 2021;160(3):958–961.e3. doi:10.1053/j.gastro.2020.09.042
245. Zhang Q, Jeppesen DK, Higginbotham JN, et al. Supermeres are functional extracellular nanoparticles replete with disease biomarkers and therapeutic targets. Nat Cell Biol. 2021;23(12):1240–1254. doi:10.1038/s41556-021-00805-8
246. Heiser CN, Simmons AJ, Revetta F, et al. Molecular cartography uncovers evolutionary and microenvironmental dynamics in sporadic colorectal tumors. Cell. 2023;186(25):5620–5637.e16. doi:10.1016/j.cell.2023.11.006
247. Fu C, Li Z, Sun Y, Lin S, Yang X, Li X. Advances in staphylococcus aureus extracellular vesicles: navigating challenges, embracing prospects. Lett Appl Microbiol. 2026;79(3):ovag009. doi:10.1093/lambio/ovag009
248. Mehraji S, Pirolli NH, Nowak R, Jay SM, DeVoe DL. Therapeutic extracellular vesicle preparation via sequential electrophoretic enrichment and counterflow microdialysis. Lab Chip. 2026;26(2):353–363. doi:10.1039/d5lc00870k
249. Tirelli V, Grasso F, Barreca V, et al. Flow cytometric procedures for deep characterization of nanoparticles. Biol Methods Protoc. 2025;10(1):bpaf019. doi:10.1093/biomethods/bpaf019
250. Alexandre L, D’Arrigo D, Kuszla N, et al. Illuminating extracellular vesicles biology with super-resolution microscopy: insights into morphology and composition. ACS Nano. 2025;19(27):24148–24173. doi:10.1021/acsnano.5c00380
251. Tutanov OS, Massick C, Ramirez M, et al. A comprehensive analysis of supermere, exomere, and extracellular vesicle isolation and cargo in colorectal cancer. Cell Rep. 2025;44(10):116287. doi:10.1016/j.celrep.2025.116287
252. Vader P, Mol EA, Pasterkamp G, Schiffelers RM. Extracellular vesicles for drug delivery. Adv Drug Deliv Rev. 2016;106(Pt A):148–156. doi:10.1016/j.addr.2016.02.006
253. Wan J, He Z, Peng R, et al. Injectable photocrosslinking spherical hydrogel-encapsulated targeting peptide-modified engineered exosomes for osteoarthritis therapy. J Nanobiotechnology. 2023;21(1):284. doi:10.1186/s12951-023-02050-7
254. Fan WJ, Liu D, Pan LY, et al. Exosomes in osteoarthritis: updated insights on pathogenesis, diagnosis, and treatment. Front Cell Dev Biol. 2022;10:949690. doi:10.3389/fcell.2022.949690
255. Xing H, Zhang Z, Mao Q, et al. Injectable exosome-functionalized extracellular matrix hydrogel for metabolism balance and pyroptosis regulation in intervertebral disc degeneration. J Nanobiotechnology. 2021;19(1):264. doi:10.1186/s12951-021-00991-5
256. Liang X, Niu Z, Galli V, et al. Extracellular vesicles engineered to bind albumin demonstrate extended circulation time and lymph node accumulation in mouse models. J Extracell Vesicles. 2022;11(7):e12248. doi:10.1002/jev2.12248
257. Lu X, Guo H, Wei X, et al. Current status and prospect of delivery vehicle based on mesenchymal stem cell-derived exosomes in liver diseases. Int J Nanomed. 2023;18:2873–2890. doi:10.2147/IJN.S404925
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Activation of Dynamin-Related Protein 1 and Induction of Mitochondrial Apoptosis by Exosome-Rifampicin Nanoparticles Exerts Anti-Osteosarcoma Effect

Chen W, Lin W, Yu N, Zhang L, Wu Z, Chen Y, Li Z, Gong F, Li N, Chen X, He X, Wu Y, Zeng X, Yueh Y, Xu R, Ji G
International Journal of Nanomedicine 2022, 17:5431-5446
Published Date: 18 November 2022
Mesenchymal Stem Cell–Derived Exosomes in Various Chronic Liver Diseases: Hype or Hope?
Zhu L, Wang Q, Guo M, Fang H, Li T, Zhu Y, Jiang H, Xiao P, Hu M
Journal of Inflammation Research 2024, 17:171-189
Published Date: 10 January 2024
Exosomes Derived from Antler Mesenchymal Stem Cells Promote Wound Healing by miR-21-5p/STAT3 Axis
Meng D, Li Y, Chen Z, Guo J, Yang M, Peng Y
International Journal of Nanomedicine 2024, 19:11257-11273
Published Date: 4 November 2024
Exosomes Derived from Apelin-Pretreated Mesenchymal Stem Cells Ameliorate Sepsis-Induced Myocardial Dysfunction by Alleviating Cardiomyocyte Pyroptosis via Delivery of miR-34a-5p

Li T, Zhao Y, Cao Z, Shen Y, Chen J, Huang X, Shao Z, Zeng Y, Chen Q, Yan X, Li X, Zhang Y, Hu B
International Journal of Nanomedicine 2025, 20:687-703
Published Date: 17 January 2025
Mesenchymal Stem Cell-Derived Exosomes for Ocular Diseases: Therapeutic Mechanisms and Clinical Perspectives
Cheng S, Ma Y, Huang F, Luo R, Han L, He L, Yuan ZX
International Journal of Nanomedicine 2025, 20:14521-14550
Published Date: 6 December 2025