Back to Journals » Biologics: Targets and Therapy » Volume 20
Research on the Role of Autophagy in Vascular Endothelial Cells in the Healing of Chronic Refractory Wounds
Authors Riling L ![]() , Kunyue F, Liyan G, Yaya W, Baoling N, Jiehui L
, Kunyue F, Liyan G, Yaya W, Baoling N, Jiehui L
Received 12 October 2025
Accepted for publication 23 December 2025
Published 13 January 2026 Volume 2026:20 573844
DOI https://doi.org/10.2147/BTT.S573844
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Shein-Chung Chow
Lao Riling,1 Fan Kunyue,1 Gan Liyan,2 Wang Yaya,1 Ni Baoling,1 Li Jiehui3
1Department of Wound Repair Surgery,Guangxi Province Hospital, Guangxi University of Traditional Chinese Medicine, Nan Ning, Guang Xi, People’s Republic of China; 2Department of Research, Guangxi Zhuang Autonomous Region Institute of Traditional Chinese Medicine, Nan Ning, Guang Xi, People’s Republic of China; 3Department of Wound Repair Surgery,Guangxi Province Hospital, the First Affiliated Hospital of Guangxi University of Traditional Chinese Medicine, Nan Ning, Guang Xi, People’s Republic of China
Correspondence: Li Jiehui, Department of Wound Repair Surgery, Guangxi Province Hospital, the First Affiliated Hospital of Guangxi University of Traditional Chinese Medicine, Nan Ning, Guang Xi, People’s Republic of China, Email [email protected]
Abstract: Chronic refractory wounds exhibit a complex etiology characterized by prolonged healing and pathological chronic inflammation, which pose significant challenges to wound recovery and may increase the risk of cancer over time. In recent years, the autophagy mechanism has emerged as a focal point of research. This process is regulated by nutritional status, growth factors, and cellular stress. Autophagy facilitates the removal of damaged organelles and misfolded proteins, recycles nutrients, contributes to immune defense, and aids in the elimination of pathogens. Endothelial autophagy is particularly crucial in the healing of chronic refractory wounds. It may be activated under conditions of ischemia, hypoxia, inflammation, and infection, thereby inhibiting cell proliferation and tissue repair while assisting cells in maintaining homeostasis and managing various stressors. Consequently, investigating the autophagic mechanisms in vascular endothelial cells during chronic refractory wound healing is essential for the development of novel wound treatment strategies.
Keywords: vascular endothelial cells, autophagy, chronic refractory wound, vascular factors, mechanism
Wound healing is a dynamic and highly coordinated process that encompasses a series of meticulously organized stages, during which various types of tissue cells interact and function to facilitate the orderly remodeling of wound tissue.1 Vascular endothelial cells (VECs) play a crucial role in the formation of new blood vessels during this process. Located on the intima of blood vessels, VECs form the inner wall and serve as a barrier for the transport of substances between blood and tissues. They are involved in immune regulation, the control of vascular permeability, and the secretion of growth factors. Additionally, VECs actively regulate the extravasation of blood cells and platelets, as well as the degree of vasodilation and contraction of blood vessels, thereby maintaining the balance between the internal and external environments of the vascular system. It plays a crucial role in regulating the inflammatory response during the early stages of wound healing by promoting leukocyte adhesion and migration to the injury site through the secretion of various adhesion molecules, while also facilitating the removal of pathogens and necrotic tissue. Vascular endothelial cells are primary contributors to angiogenesis; they can autonomously migrate to the repair site, enhance blood supply to the wound through proliferation and migration, provide essential nutrients, and create a favorable growth environment.2 The formation of new blood vessels is a critical component of the wound healing process. Despite variations in blood pressure and flow dynamics across different body regions, vascular endothelial cells effectively maintain low osmotic pressure within blood vessels and regulate the internal fluid balance, underscoring their vital role in wound healing.3 Many studies have shown that autophagy plays an extremely important role in diseases such as tumors, diabetic wounds, retinopathy, and cardiovascular diseases. By regulating specific autophagy targets and signaling pathways, it provides a theoretical basis for the clinical treatment of related diseases. Common autophagy factors include PI3K, Beclin, P62, etc. By regulating autophagy signaling factors, they activate autophagy in cells, provide nutrients for new blood vessel formation and tissue regeneration, and promote wound healing. For instance, MEBO Moist Burn Ointment regulates the expression of P62, Beclin, and LC3 factors, stimulating autophagy in cells and accelerating the healing of diabetic wounds.4 Autophagy signaling pathways related to wound healing include PI3K/Akt, PINK/parkin, MAPK/ERK1/2, etc. For instance, traditional Chinese medicine regulates the MAPK/ERK1/2 signaling pathway, increases the expression level of LC3-II, stimulates the paracrine secretion of VEGF in mesenchymal stem cells (MSCs), and promotes vascular regeneration in chronic wounds. Shorten the wound healing time.5 This indicates that the autophagy mechanism is closely related to wound healing, and autophagy also affects the level of wound healing to a certain extent.
Overview of Autophagy
Autophagy is a highly conserved self-degradation mechanism in eukaryotic cells. Autophagy also degrades functional proteins to regulate multiple signaling pathways. By doing so, autophagy helps maintain intracellular homeostasis, supplies essential nutrients and energy, and facilitates self-degradation under conditions such as nutrient deprivation, oxidative stress, and pathogen infection, thereby promoting cell survival and renewal. Autophagy can be categorized into microautophagy, macroautophagy, and chaperone-mediated autophagy (CMA).6 Microautophagy involves the direct sequestration and degradation of substrates within lysosomes or vacuoles, playing a crucial role in intracellular protein quality control and organelle turnover.7 Macroautophagy represents the most significant form of autophagy. Autophagosomes are generated from expanding membrane structures derived from the endoplasmic reticulum, which subsequently fuse with lysosomes to degrade their contents. This process is utilized to eliminate intracellular macromolecules, including protein aggregates, damaged organelles, and intracellular pathogens.8 All trabecular autophagy studies cited below are primarily macroautophagy. Chaperone-mediated autophagy (CMA) represents a specialized form of autophagy in which molecular chaperones, such as heat shock proteins, recognize and bind to misfolded or unfolded proteins, directing them to autophagosomes for degradation. Under normal conditions, autophagy effectively removes damaged organelles and denatured proteins, thereby maintaining intracellular homeostasis.9 However, there are certain risks to autophagy in cells, including problems that may be caused by excessive activation or inhibition of autophagy. Excessive activation of autophagy may lead to excessive degradation of cells’ own components, causing cell dysfunction or even death. It may also damage related neurons and induce diseases such as neurodegeneration. Inhibiting autophagy in cells may lead to the accumulation of damaged proteins and organelles within cells that cannot be cleared in a timely manner, increasing the risk of cell canceration, intensifying inflammatory responses, and affecting normal cell metabolism and function, as well as causing adverse reactions such as abnormal immune systems.10 Therefore, it is necessary to precisely control and regulate the autophagy mechanism to prevent the occurrence of adverse reactions.
Autophagy encompasses initiation, elongation, maturation, and lysosomal binding.11 Following the activation of autophagy, autophagosomes are generated through the involvement of the phosphatidylinositol 3-kinase III (PI3K-III) complex and the ULK complex. These structures extend to encapsulate cellular contents, aided by the autophagy-related protein family and various ubiquitin-like reactions, ultimately resulting in the formation of autophagosomes. Subsequently, autophagosomes fuse with lysosomes, leading to the degradation of the encapsulated substrates into smaller molecules, such as amino acids and fatty acids (Figure 1).12 Autophagy involves the formation of vesicles that encapsulate portions of the cytoplasm, as well as organelles or proteins designated for degradation. This process is regulated by autophagy-related genes (ATG) and is essential for maintaining cellular homeostasis, metabolism, and stress responses. Autophagosomes, which are formed during autophagy, fuse with lysosomes that contain a range of hydrolytic enzymes capable of degrading the materials enclosed within the autophagosomes. This degradation process aids in the removal of damaged or harmful cellular components, thereby contributing to the balance and stability of the intracellular environment.13
 |
Figure 1 The specific process of autophagy in cells. Under the stimulation of oxidative stress and other conditions, cells activate the autophagy mechanism, activating the PI3K signaling pathway to form the RIPK3 complex and the mTOR-related signaling pathway to form the ULK1 complex. Under the combined action of the two, the autophagosome premembrane is formed. During the autophagy elongation, beclin-1 binds to PI3K and recruits ATG proteins to form a double-membrane structure. LC3-1 is cleaved by ATG4 to form LC3-2, and with the help of the ATG family, it is located on the autophagosome membrane, marking autophagosome maturation. Ultimately, the autophagosome membrane enclosing the contents fuses with the lysosomal membrane to form autophagolysosomes, and the substrates they carry are degraded into small molecules such as amino acids and fatty acids (figure was created with Biorender.com). |
Chronic Refractory Wound Healing and Vascular Endothelial Cells
Healing Mechanism of Chronic Refractory Wound
Chronic refractory wounds are classified as “sores” in traditional Chinese medicine. The Ming Dynasty text “Surgical Qixuan” describes “sores” as a general term for lesions characterized by injury, muscle necrosis, and pain, resulting from traumatic wounds. Traditional Chinese medicine posits that deficiency, toxins, blood stasis, and decay are the primary pathogenic factors contributing to the occurrence and progression of chronic refractory ulcers.First, “deficiency is the place where evil remains.” Second, the deficiency of qi and blood, as well as the imbalance of yin and yang caused by chronic illness, leads to impaired blood circulation and stasis; thus, deficiency and blood stasis serve as the root causes of the disease. Third, the accumulation of pathological products, including toxins and decay in the wound and surrounding tissues, disrupts qi and blood circulation, making ulcers difficult to heal repeatedly. Western medicine typically defines chronic refractory wounds as those that have not healed for more than a month despite conventional treatment and show no signs of healing. Factors contributing to the difficulty in wound healing include local blood circulation disorders, infections, and systemic diseases.14 In diabetic patients, elevated blood sugar levels, along with peripheral vascular and neurological lesions, predispose them to foot ulcers that are challenging to heal. Additionally, in the elderly, a decline in physical function and weakened tissue repair capacity often result in delayed wound healing.15 The healing of chronic wounds presents a significant challenge that requires urgent attention in clinical practice. Chronic wound healing is fundamentally a dynamic process that facilitates the regeneration of biological functions in injured tissues, including the inflammatory response, cell proliferation, reepithelialization, collagen formation, angiogenesis, and tissue remodeling16 (Figure 2). During the wound healing process, there are a large number of basal cells around the injury site, which divide into various cells through mitosis, such as monocytes, fibroblasts, endothelial cells, etc. Monocytes migrate from the blood to various tissues to form macrophages, which are crucial components of the immune system and have functions such as phagocytosis of pathogens, immune regulation, tissue repair and remodeling. Fibroblasts are involved in the renewal and remodeling of extracellular matrix, promoting cell proliferation, migration and differentiation by secreting growth factors, and thereby facilitating the re-epithelialization of wounds. Endothelial cells secrete various bioactive substances, such as nitric oxide (NO), endothelin (ET), vascular growth factor (VEGF), etc. Under the guidance of signals like VEGF, they migrate and proliferate to form vascular buds, which form functional vascular networks with adjacent blood vessels. New blood vessels provide nutrients and oxygen for granulation tissue formation, thereby promoting wound tissue remodeling and accelerating wound healing.17–19 However, chronic refractory wounds are frequently characterized by inflammatory cell infiltration, which impedes vascular regeneration and the formation of a vascular network, ultimately prolonging the healing time. Research indicates that the primary cause of delayed healing in chronic wounds is the persistent infiltration of inflammatory cells, which extends the inflammatory reaction period and adversely affects vascular regeneration and network formation.20
 |
Figure 2 Healing process of chronic wounds.The healing process of chronic difficult-to-heal wounds begins with the production of inflammatory cells around the injury site, the secretion of inflammatory factors and then the inflammatory response, and the migration of the existing basal cells to the wound, which through division and value-added formation of monocytes, fibroblasts and endothelial cells. Monocytes migrate and differentiate into macrophages, which are capable of phagocytosis and digestion of pathogens, dead cells and foreign bodies. Angiogenic factors secreted by endothelial cells and extracellular matrix secreted by fibroblasts promote the formation of new blood vessels, which in turn promotes the remodeling of the wound and accelerates wound healing (figure was created with Biorender.com). |
Endothelial Cells Participate in Wound Healing
Skin wound healing results from the interplay of various cell types and active factors. Endothelial cells play a crucial role during angiogenesis, as they branch from existing vessels to create immature new vessels through a process known as “budding.” In chronic refractory wounds, a significant number of endothelial cells migrate to the wound periphery, where they develop into vascular buds. These buds penetrate the basement membrane, invade the extracellular matrix, and extend to form branches, ultimately creating a vascular network within the wound. The reduced blood supply to the injured area promotes hypoxia in the wound tissue, which stimulates the formation of new blood vessels to enhance wound healing and accelerate cellular metabolism.21 When the wound healing is in the inflammatory development stage, the space between vascular endothelial cells widens under the action of bradykinin and histamine, the permeability of the vessel wall increases, and plasma components exude. The platelet-derived growth factor (PDGF)secreted by them promotes the proliferation and migration of endothelial cells. Consequently, the newly formed granulation tissue is rich in capillaries, which proliferate, migrate, and extend toward the healing area under the influence of various angiogenic factors. The migration of endothelial cells is facilitated by plasminogen activator and collagenase, which they produce. As a result, bud-like capillaries develop within the granulation tissue, extending and fusing to create a network that ultimately forms new blood vessels, allowing for blood inflow.22,23 Intracellular reactive oxygen species (ROS) exhibit dual effects on angiogenesis, promoting neovascularization at low levels while inhibiting the proliferation, migration, and neovascularization of vascular endothelial cells at elevated levels.24 During the initial phase of wound healing, intracellular ROS levels significantly increase, prompting cells to adopt a healing state. However, inflammatory cells also release substantial amounts of ROS through respiratory bursts to combat infection. Excessive ROS levels can adversely affect cell proliferation and migration, induce apoptosis, impede wound healing, and potentially lead to scar hyperplasia.25 In diabetic wounds, hyperglycemia stimulates ROS production and reduces nitric oxide levels in vascular endothelial cells, severely impairing their normal function, delaying wound healing, and potentially resulting in non-healing wounds.26 Coordinating oxidation and antioxidation may represent an optimal strategy for wound treatment to regulate the levels of reactive oxygen species (ROS) essential for the healing process, preventing excessive ROS expression that could impede vascular endothelial cell proliferation, migration, and angiogenesis, thereby potentially delaying or halting wound healing progress.27
The formation and functional maturation of new blood vessels are initiated by endothelial cells through two primary processes: proliferation and migration. Following injury, the proliferation of vascular endothelial cells increases, and the secretion of angiogenic factors promotes rapid vascular growth.28 Angiogenic factors are crucial for the formation of new blood vessels and can be categorized into two groups. The first group includes basic fibroblast growth factor (bFGF) and PDGF, which induce endothelial cell chemotaxis and stimulate cell division and proliferation. The second group comprises transforming growth factor-β (TGF-β), transforming growth factor-ɑ(TGF-ɑ), and angiopoietin, which inhibit endothelial cell proliferation, promote differentiation, and facilitate the formation of interconnected open capillaries.29 Vascular endothelial growth factor (VEGF) is a crucial mediator in promoting vascular growth, essential for effective wound healing. VEGF specifically stimulates mitosis in vascular endothelial cells, triggering the production of proteases, migration towards signaling cues, cell proliferation, and ultimately tube formation and differentiation in endothelial cells. It enhances the proliferation of vascular endothelial cells in vitro and induces angiogenesis in vivo.30 During wound healing, VEGF prompts coagulation factors to support endothelial cell function by increasing vascular permeability, facilitating new blood vessel formation. Additionally, it modulates the balance between enzyme activators and inhibitors in the local vascular environment, creating a proteolytic milieu that encourages endothelial cell migration beyond customary boundaries.31 As blood reaches tissues post-neovascularization, oxygen levels rise, leading to decreased hypoxia-inducible factor-1 (HIF-1) activity, reduced VEGF secretion, and gradual cessation of angiogenesis.32 The roles and implications of VEGF family factors in various diseases are detailed in Table 1. Therefore, vascular endothelial cells not only play an important role in angiogenesis, but also accelerate proliferation and migration through autocrine and paracrine mechanisms, thus promoting wound healing.
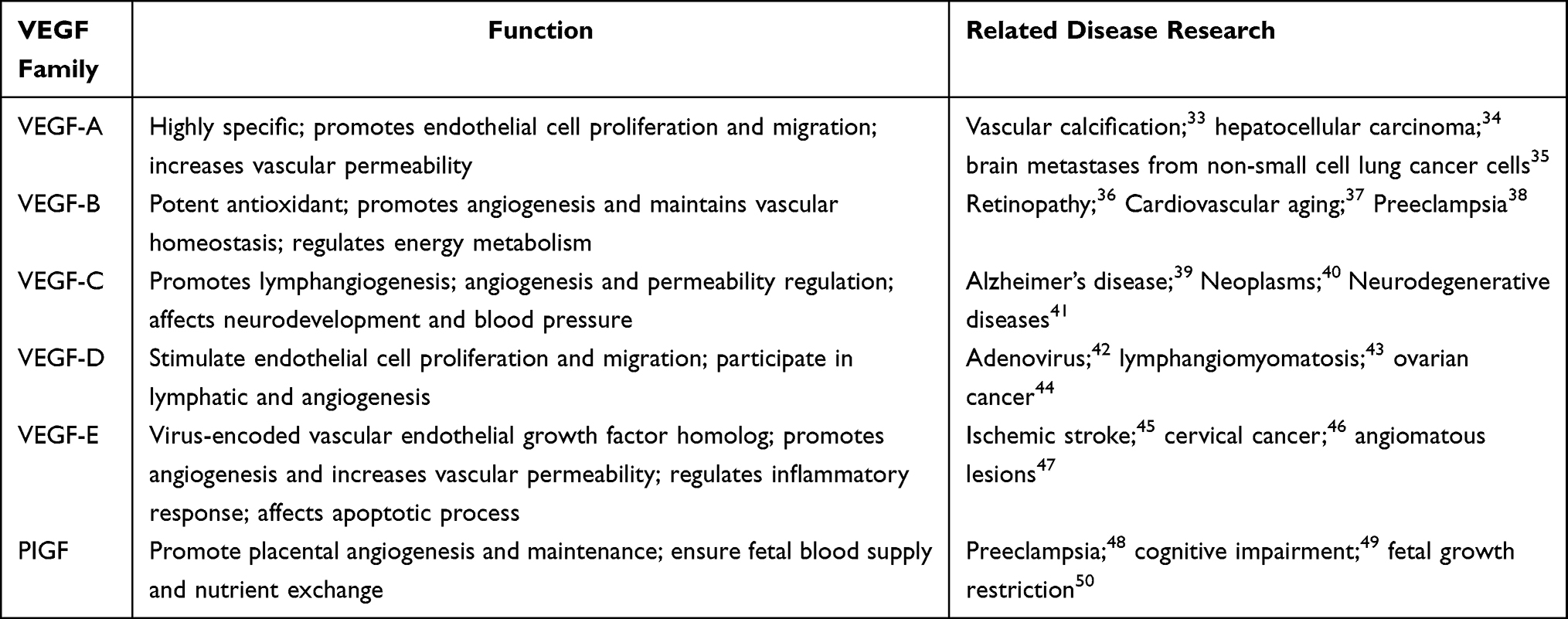 |
Table 1 Various Factor Functions of the VEGF Family |
Wound Healing and Autophagy
During chronic refractory wound healing, factors such as mitochondrial stress, inflammatory mediators, hypoxia, and infection can trigger autophagy in vascular endothelial cells. This process subsequently impacts both the structure and function of these cells, thereby modulating apoptosis and disrupting normal wound healing. It has been pointed out that proper autophagy contributes to angiogenesis and tissue repair. Because proper autophagy not only needs to maintain a certain basic autophagic activity of the cell to remove aged or damaged organelles, protein aggregates, etc., but also needs to be induced by stress to activate autophagy, when the cell is faced with nutrient deficiencies, oxidative stress, infections and other stresses, autophagy will be activated to enhance the ability to remove harmful substances, helping the cell to survive.51 According to the findings,52,53 vascular endothelial autophagy plays a role in the wound healing process in the following ways:
Autophagy and the Inflammatory Response
During wound inflammation, vascular endothelial cell autophagy plays a crucial role in influencing the survival of inflammatory cells, the release of inflammatory factors, and the elimination of invading pathogens. It exerts anti-infective effects, negatively regulates the inflammatory response, partially modulates the wound’s inflammatory reaction, and promotes wound cleansing and healing.54 Gao Haixia55 demonstrated that the knockout of the endothelial cell autophagy-related gene Atg7 reduces the secretion of inflammatory factors in diabetic wounds and modulates the wound healing process via a paracrine pathway. Atg7 is a critical protein involved in cell autophagy, mitophagy, and the induction of the mitochondrial stress response, thereby regulating cell cycle. In the study conducted by Pan Dongmei et al56 it was found that the novel MoS2 nanoenzyme protects against inflammatory endothelial cell injury by regulating mitophagy and vascular endothelial cells, suggesting its potential for preventing and treating diseases related to chronic inflammatory vascular endothelial damage.
Gene knockout technology serves as a pivotal method in molecular biology, fundamentally based on the targeted removal or inactivation of specific genes within an organism. This approach allows for the observation of resultant phenotypic changes, thereby elucidating the direct causal relationship between genes and phenotypes. The technology offers highly precise gene-targeting tools that facilitate the investigation of wound repair mechanisms and enable a systematic analysis of the roles of specific genes in critical biological processes, including tissue regeneration, inflammation regulation, and cell proliferation. Nonetheless, gene knockout technology is confronted with significant safety concerns in practical applications. The procedure may inadvertently induce unexpected gene mutations or alterations in chromosomal structure, potentially disrupting normal immune regulatory functions and leading to abnormal immune responses or the onset of autoimmune diseases. Furthermore, the long-term biological effects and genetic stability associated with gene editing remain inadequately studied and understood, rendering the prediction and control of its potential risks challenging. Consequently, the clinical application of gene knockout technology has been mired in extensive ethical scrutiny and safety debates. Such controversies have hindered the translation and implementation of this technology in clinical contexts, such as wound treatment. Currently, its use in human medicine is primarily confined to basic research and preclinical testing stages.
Autophagy and Neovascularization
During wound angiogenesis, autophagy in vascular endothelial cells eliminates damaged organelles and proteins, decreases the buildup of harmful substances, enhances cell survival, and exerts anti-apoptotic and anti-oxidative effects. Additionally, autophagy breaks down and reuses cellular components, supplying essential nutrients and energy for new tissue formation and vascular regeneration, thereby promoting vascular endothelial cell development.57 Wei Haibin58 demonstrated in animal experiments that the autophagy of vascular endothelial cells regulates the activity of apoptosis factors, including Bcl-2 and Caspase-8. This regulation influences the death receptor pathways associated with apoptosis and endoplasmic reticulum stress, subsequently affecting vascular permeability and leukocyte adhesion. Hao Dan et al59 reported that the expression of LC3-II and Beclin-1 in vascular endothelial cells was upregulated, which activated autophagy, contributed to the oxidative stress response in endothelial cells, facilitated the degradation of oxidative proteins, and induced apoptosis. Li Yuanyuan60 found that H2S may inhibit both autophagy and apoptosis in vascular endothelial cells by activating the PI3K/Akt/mTOR signaling pathway, thereby promoting angiogenesis in wound tissue and enhancing diabetic wound healing.
Animal wound models, with their capacity to closely mimic the human wound healing process, have demonstrated significant value in drug development and the investigation of wound healing mechanisms. They serve as essential tools for the comprehensive exploration of related pathophysiological processes and the evaluation of potential treatment options. Nonetheless, these models encounter several challenges in practical applications. Firstly, inherent species differences can compromise the accuracy and reliability of translating results from animal experiments to human clinical trials. This discrepancy may result in certain drugs or therapies that appear effective in animal models proving ineffective or even posing safety risks in human trials. Secondly, as awareness of animal welfare increases and ethical standards become more stringent, the use of animal models faces growing ethical restrictions, which, to some extent, limits the scope of experimental research.Moreover, the construction and maintenance of animal wound models typically necessitate significant financial investment. This includes expenses related to the procurement and sustenance of experimental animals, the utilisation of specialised equipment, and the training of technical staff, all of which contribute to elevated research costs. Simultaneously, the establishment of these models, along with subsequent observation and detection procedures, is relatively intricate. This complexity demands a high level of technical expertise from experimental personnel, thereby not only increasing the operational challenges but also presenting substantial obstacles to large-scale experimental research. The cumulative impact of these factors hinders animal wound models from adequately addressing the practical requirements for extensive screening and the swift advancement of clinical translation applications. Consequently, there is an urgent necessity to explore and develop more refined alternative research models.
Autophagy and Scar Repair
Autophagy of vascular endothelial cells can regulate the production of transforming growth factor-β (TGF-β) by cells, expel “abnormal” extracellular matrix from cells, and reduce wound scar hyperplasia.ZAMANI61 reported decreased expression levels of TGF-β1and VEGF in treatment groups in an in vitro wound healing model, validating the efficacy of alginate hydrogel with losartan in wound repair through scar size reduction and tissue remodeling. Losartan mitigates angiotensin II–induced damage to endothelial cells by inhibiting the angiotensin II-receptor connection. It modulates endothelial cell autophagy levels and enhances autophagy, thereby ameliorating endothelial cell function via activation of the AMPK signaling pathway.62.
In vitro wound healing models offer several advantages for research applications, including the ability to create wounds of varying types and sizes, effectively simulate multiple stages of inflammatory responses, and observe the migration behaviours of epithelial cells and fibroblasts. Additionally, these models feature a relatively straightforward operational process and low experimental costs, rendering them suitable for large-scale preliminary screening and mechanism exploration. However, they also exhibit notable limitations, the most significant being their inability to replicate the complex physiological environment of the body. This includes the absence of functional vascular networks, the involvement of various immune cells, and in vivo factors such as neural and endocrine regulation. Consequently, these models struggle to accurately represent the dynamic distribution, metabolic processes, and specific target sites of drugs in actual wounds, thereby constraining their value in translational medicine research.
Autophagy therapy, as a disease treatment approach that has emerged in recent years, plays a significant role in the clinical treatment of various diseases. By targeting and regulating the relevant autophagy proteins or signaling pathways in diseases and precisely delivering drugs to therapeutic targets, the therapeutic effect can be improved and the side effects caused by drugs can be reduced. Therefore, autophagy therapy has broad prospects for development in clinical practice and is expected to become a new treatment option for various diseases. Provide a theoretical basis for the research and development of new types of traditional Chinese medicine plasters.
Drug Development and Challenges in Clinical Targeted Therapy
The new type of Chinese herbal ointment refers to the combination of advanced technological means on the basis of traditional ointment or compound formula to precisely locate the therapeutic target point and regulate the relevant signaling pathway, so as to enhance the therapeutic effect, safety and popularity. For example, MEBO, Shengmian Yu Hong Ointment and Comfrey Ointment are new types of Chinese herbal ointment for clinical treatment of wounds. After relevant studies, it was found that MEBO can regulate the expression of TNF-a, IL-6, mTOR and other factors, reduce inflammatory response and promote wound healing.63 Comfrey ointment can increase the expression of VEGF and other growth factors, improve local blood circulation, accelerate the growth of traumatic granulation tissues and epithelialization process and so on.64 Therefore, this paper provides a theoretical basis for the development of a new type of herbal ointment that promotes wound healing by regulating vascular endothelial cell autophagy by exploring the connection between the mechanism of vascular endothelial cell autophagy and the healing of chronic difficult-to-heal wounds.
Clinical targeted autophagy therapy may face multiple challenges. Firstly, autophagy has a dual function. In the early stage of tumors, autophagy can inhibit tumor occurrence by eliminating damaged organelles. However, in the advanced stage, autophagy may not provide nutritional support for the survival of tumor organelles, which requires precise control of the degree of autophagy in cells to avoid excessive autophagy or inhibit autophagy.65 Secondly, the level of autophagy in cells is related to tissue specificity. There are significant differences in the expression levels and functions of autophagy-related proteins (such as LC3 and ATG5) in different tissues, which can lead to inconsistent drug delivery efficiencies.66 Clinical targeted autophagy therapeutic drugs may also cause drug delivery and toxicity issues, as autophagy is a widespread cellular metabolic process. Systemic inhibition of autophagy may lead to serious side effects, such as autophagy deficiency causing myocardial cell damage and neurodegeneration.67 At present, in vivo measurement of autophagic flux in human wounds mainly relies on technological extensions in animal model research, and there is no non-invasive, real-time measurement method that is fully applicable to the human body. Animal experimental wound models obtain wound tissue samples and use experimental techniques such as immunofluorescence, genetic testing, and transmission electron microscopy to detect changes in the expression and distribution of autophagy-related proteins (such as LC3 and p62), which indirectly reflects autophagy flux. However, the accuracy, effectiveness and safety of these technologies for measuring human wounds have yet to be studied. In the future, it is necessary to develop safer, non-invasive and in vivo measurement and quantification technologies for real-time monitoring of autophagic flux in human wounds. In addition, precisely delivering biological agents to specific cells in chronic and refractory wounds requires precise regulation of autophagy targets and signaling pathways. However, there are currently no reliable biomarkers that can predict the response of autophagy inhibitors to wound cells. Moreover, external conditions or changes in the internal environment can also affect the accessibility and localization accuracy of the targets. Since people’s understanding of the autophagy mechanism is mostly still at the theoretical and animal experiment stage, transforming theoretical research into clinical research means conducting a large number of ethical experiments for verification.
Conclusions
The treatment of chronic and refractory wounds remains a major challenge in medicine. Both traditional Chinese medicine and Western medicine have limitations in clinical treatment. Traditional Chinese medicine mainly focuses on the mechanism research of one-way pathways such as herbal medicines, compound prescriptions, and external treatments of traditional Chinese medicine, lacking a systematic network analysis of “multi-component - multi-target - dynamic regulation”. The absence of objectification of syndrome differentiation standards leads to significant fluctuations in therapeutic effects. In Western medicine, techniques such as negative pressure drainage and flap transplantation are mainly used for treatment. For instance, stem cell therapy, as an emerging method for treating wounds, has achieved remarkable therapeutic effects. However, due to the risks of unexpected differentiation and genomic instability, as well as the high cost of treatment, it is difficult to be widely popularized in clinical practice. Therefore, there is an urgent need to seek new solutions for improving the clinical treatment of chronic and refractory wounds. Vascular endothelial cells, as one of the key cells for forming new blood vessels during wound healing, participate in the important stage of wound healing. They promote angiogenesis and tissue remodeling by secreting various growth factors and activating related signaling pathways. The signaling pathways of vascular endothelial cells are mainly related to proteins such as protein kinase C and MAPK. Various inflammatory mediators and cytokines act on MAPK or connexin by activating downstream Rho kinase, causing the tight junctions and adhesion junction structures between vascular endothelial cells to loosen, increasing vascular permeability and reducing blood volume. As an autophagy signaling factor, MAPK can interact with other proteins to induce autophagy in cells and maintain the homeostasis of the intracellular environment. This indicates that the growth factors secreted by vascular endothelial cells may, under certain conditions, be able to combine with MAPK protein to activate the autophagy mechanism of cells. Therefore, in-depth exploration of the sequential regulatory rules of vascular endothelial cytokines and autophagy-related signaling pathways, combined with the exploration of metabolokinetics to explore the dynamic action trajectory of the wound microenvironment, provides a new direction for the clinical treatment of chronic refractory wounds.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
National Natural Science Foundation of China (82360931), Guangxi Natural Science Foundation of China (2024GXNSFAA010288), Guangxi University of Traditional Chinese Medicine 2025 Postgraduate Scientific Research Innovation Project (YCSY2025031, YCSY2025032).
Disclosure
The authors declare no competing interests in this work.
References
1. Seth G, Herron A, Davis J, Phillips M, Chakrabarti M, Evans CE. Control of inflammatory lung injury and repair by metabolic signaling in endothelial cells. Curr Opinion Hematol. 2025;32(3):157–11. doi:10.1097/MOH.0000000000000848
2. Wen SJ, Liu JL. Review of vascular endothelial cell function. J Reprod Med. 2005;14(5):310–313.
3. Guo L. Protective Effects of Shikonin on H2O2-Induced Vascular Endothelial Cell Injury and Skin Microvascular Reconstruction. Xinjiang: Tarim University; 2023.
4. Ma HL, Huang Q, Zheng AT, et al. Regulation of trabecular autophagy signaling factors Beclin1, LC3, and p62 by MEBO in diabetic rats. Chin J Gerontol. 2022;42(2):356–365.
5. Qi YZ, Wang Z, Chen C, et al. Research progress on the regulation of p38 MAPK, ERK1/2, JNK, etc. by traditional chinese medicine in the treatment of osteoporosis. Chin J Exp Traditional Med Formulae. 2023;29(12):222–229.
6. Dikic I, Elazar Z. Mechanism and medical implications of mammalian autophagy. Nat Rev Mol Cell Biol. 2018;19(6):349–364. doi:10.1038/s41580-018-0003-4
7. Takahashi Y, San TT, Manik MIN, et al. The greatwall kinase Rim15 promotes microautophagy and microlipophagy under the control of TORC1. Biochem Biophys Res Commun. 2025;752:151468. doi:10.1016/j.bbrc.2025.151468
8. Shi B, Ma M, Zheng Y, et al. mTOR and Beclin1: two key autophagy-related molecules and their roles in myocardial ischemia/reperfusion injury. J Cell Physiol. 2019;234(8):12562–12568. doi:10.1002/jcp.28125
9. Luo R. Unraveling the complexity of chaperone-mediated autophagy in aging: insights into sex-specific and cell-type-specific regulation. Autophagy. 2025;1–2. doi:10.1080/15548627.2025.2601849
10. Wu JS. A talk on autophagy. Biol Teaching. 2017;42(4):66–67.
11. Chen XL, Wang SL. Autophagy process, pathway, regulation and its multiple correlation with pulmonary hypertension. Chin J Tissue Eng Res. 2021;25(2):311–316.
12. Zhou P, Cui YL, Zhang WT, et al. Role of autophagy in cerebral ischemia injury and regulatory mechanism of traditional Chinese medicine. Chin J Tissue Eng Res. 2025;29(8):1650–1658.
13. Cheng YQ, Peng BW. Effect of lysosomal pH homeostasis on autophagy and autophagy flow. Guangdong Chem Indus. 2024;51(12):47–49.
14. Wang P, Yin B, Su YJ, et al. Research advances in healing mechanism of chronic refractory wounds mediated by long non-coding RNA. Chinese J Burns. 2020;36(8):758–761. doi:10.3760/cma.j.cn501120-20190526-00254
15. Tang J, Guan H, Dong W, et al. Application of compound polymyxin b ointment in the treatment of chronic refractory wounds. Int J Low Extrem Wounds. 2022;21(3):320–324. doi:10.1177/1534734620944512
16. Dong Y, Yang Q, Sun X. Comprehensive analysis of cell therapy on chronic skin wound healing: a meta-analysis. Human Gene Therapy. 2021;32(15–16):787–795. doi:10.1089/hum.2020.275
17. Zhou Z, Sun X, Huang X, et al. Application of SVF microinjection combined with skin tension in ischemic chronic refractory wounds. Regenesis Repair Rehabil. 2025;1(1):58–64. doi:10.1016/j.rerere.2024.11.002
18. Zhao YM, Chen Y, Xie B, et al. Progress in the application of induced membrane technology in chronic refractory wounds. Chinese J Burns Wounds Surface Ulcers. 2025;37(1):9–12.
19. Yang X, Zhang X, Tian Y, et al. Srsf3-dependent APA drives macrophage maturation and limits atherosclerosis. Circ Res. 2025;136(9):985–1009. doi:10.1161/CIRCRESAHA.124.326111
20. Guo L, Wei B, Pan F, et al. Effects of dual-gene modification on biological characteristics of vascular endothelial cells and their significance as reserving cells for chronic wound repair. Growth Factors. 2022;40(5–6):221–230. doi:10.1080/08977194.2022.2118119
21. Gold Stone. Effect of Platelet on Vascular Endothelial Cells and Wound Healing Induced by Oxidative Stress. Guangdong: Southern Medical University; 2023.
22. Xu DM, Du JJ, Yang Y, et al. Research progress on mechanism of polysaccharides from traditional Chinese medicine promoting chronic refractory wound healing. Chin J Exp Traditional Med Formulae. 2024;30(8):245–252.
23. Li XL. Differential Expression Analysis of circRNAs and miRNAs in Diabetic Wound and Mechanism of hsa_circRNA_000554 Regulating Wound Angiogenesis. Jiangxi: Nanchang University Medical Department; 2023.
24. Schiffmann LM, Werthenbach JP, Heintges-Kleinhofer F, et al. Mitochondrial respiration controls neoangiogenesis during wound healing and tumour growth. Nat Commun. 2020;11(1):3653. doi:10.1038/s41467-020-17472-2
25. Zheng B, Wang Q, Xiao YB, et al. Progress of mitochondrial autophagy in atherosclerosis. Chinese J Pathophysiol. 2025;41(03):585–592.
26. Li MY, Liu S, Chen KY, et al. MXene nanoparticles Ti3C2TX and photothermal effect promote wound healing in diabetic mice. Chin J Tissue Eng Res. 2025;29(28):6052–6060.
27. Dunnill C, Patton T, Brennan J, et al. Reactive oxygen species (ROS) and wound healing: the functional role of ROS and emerging ROS-modulating technologies for augmentation of the healing process. Int Wound J. 2017;14(1):89–96. doi:10.1111/iwj.12557
28. Fang Y, Qian ZY, Hada HY, et al. Potential mechanism of Mongolian medicine Echinops lanatus on proliferation and angiogenesis of vascular endothelial cells. Chin J Tissue Eng Res. 2025;29:35):7519–7528.
29. Zhang K, Zhu CL, Zhang Y. Study on the protective effect of growth differentiation factor 15 in heart transplantation. Chinese J Extracorporeal Circul. 2025;23(1):76–80.
30. Revilla G, Mulyani H, Yerizel E, et al. Human mesenchymal stem cell promotes burn wound healing by inducing FGF and VEGF in diabetic rat. Pakistan J Biologic Sci. 2024;27(10):506–511. doi:10.3923/pjbs.2024.506.511
31. Zhang Y, Zhang YY, Pan ZW, et al. GDF11 promotes wound healing in diabetic mice via stimulating HIF-1ɑ-VEGF/SDF-1ɑ-mediated endothelial progenitor cell mobilization and neovascularization. Acta Pharmacol Sin. 2023;44(5):999–1013. doi:10.1038/s41401-022-01013-2
32. Robin H, Hannig L, Bernhard JS. Tissue origin of endothelial cells determines immune system modulation and regulation of HIF-1α-, TGF-β-, and VEGF signaling. iScience. 2025;28(2):111740. doi:10.1016/j.isci.2024.111740
33. Fu C, Liang Q, Ma L, et al. miR-29a-3p/Vegfa axis modulates high phosphate-induced vascular smooth muscle cell calcification. Ren Fail. 2025;47(1):2489712. doi:10.1080/0886022X.2025.2489712
34. Xu X, Zhang Y, Wu S, et al. Hepatitis B virus promotes angiogenesis in hepatocellular carcinoma by increasing m6A modification of VEGFA mRNA via IGF2BP3. J Med Virol. 2025;97(5):e70356. doi:10.1002/jmv.70356
35. Xu X, Yang G, Shi N. Ceritinib reduces transendothelial invasion of non-small cell lung cancer cells by restoring claudin-10 and suppressing VEGF-A signaling. Biochem Genet. 2025. doi:10.1007/s10528-025-11103-5
36. Chen R, Lee C, Lin X, et al. Novel function of VEGF-B as an antioxidant and therapeutic implications. Pharmacol Res. 2019;143:33–39. doi:10.1016/j.phrs.2019.03.002
37. Manickam N, Sultan I, Panthel J, et al. Beneficial effects of vascular endothelial growth factor B gene transfer in the aged heart. Cardiovasc Res. 2025;121(10):1594–1608. doi:10.1093/cvr/cvaf046
38. Eslahi H, Saravani M, Shahraki M, et al. Investigation of the placental levels of vascular endothelial growth factor B and selenoprotein P and their relation with birth weight in patients with pre-eclampsia: a case-control study. Int J Fertil Steril. 2025;19(2):246–250. doi:10.22074/ijfs.2024.2030292.1698
39. Ma YN, Wang Z, Tang W. Deep cervical lymphaticovenous anastomosis in Alzheimer’s disease: a promising frontier or premature enthusiasm? Biosci Trends. 2025;19(2):144–149. doi:10.5582/bst.2025.01108
40. Elieh-Ali-Komi D, Maurer M, Siebenhaar F. The impact of mast cells on the anatomy, cellular communication, and molecular immune network of lymph nodes. Clin Rev Allergy Immunol. 2025;68(1):35. doi:10.1007/s12016-025-09050-5
41. Kim K, Abramishvili D, Du S, et al. Meningeal lymphatics-microglia axis regulates synaptic physiology. Cell. 2025;188(10):2705–2719.e23. doi:10.1016/j.cell.2025.02.022
42. Rissanen TT, Markkanen JE, Gruchala M, et al. VEGF-D is the strongest angiogenic and lymphangiogenic effector among VEGFs delivered into skeletal muscle via adenoviruses. Circ Res. 2003;92(10):1098–1106. doi:10.1161/01.RES.0000073584.46059.E3
43. Lagos Castillo M, Reyes Cartes F. Lymphangioleiomyomatosis: update on pathophysiology, diagnosis, and treatment. Rev Med Chil. 2025;153(1):63–71. doi:10.4067/s0034-98872025000100063
44. Náležinská M, Chovanec J. Vascular endothelial growth factor D potential predictor and screening marker in ovarian carcinoma. Ceska Gynekol. 2025;90(1):22–37. doi:10.48095/cccg202522
45. Wang J, Wu X, Fang J, et al. Intervention of exogenous VEGF protect brain microvascular endothelial cells from hypoxia-induced injury by regulating PLCγ/RAS/ERK and PI3K/AKT pathways. Exp Gerontol. 2024;192:112452. doi:10.1016/j.exger.2024.112452
46. Nayarisseri A, Abdalla M, Joshi I, et al. Potential inhibitors of VEGFR1, VEGFR2, and VEGFR3 developed through deep learning for the treatment of cervical cancer. Sci Rep. 2024;14(1):13251. doi:10.1038/s41598-024-63762-w
47. Fagiani E, Lorentz P, Bill R, Pavotbawan K, Kopfstein L, Christofori G. VEGF receptor-2-specific signaling mediated by VEGF-E induces hemangioma-like lesions in normal and in malignant tissue. Angiogenesis. 2016;19(3):339–358. doi:10.1007/s10456-016-9508-7
48. Luo W, Tang P, LV Y, et al. Maternal PlGF and sFlt-1 are associated with low birth weight and/or small-for-gestational age neonates in pregnancy with or without preeclampsia. AJP Rep. 2025;15(1):e36–e42. doi:10.1055/a-2555-1742
49. Lam JC, Louras P, Savettiere A, et al. Worry moderates plasma placental growth factor (PIGF) and cognition in older adults with amnestic mild cognitive impairment (aMCI). Exp Aging Res. 2025;1–16.
50. Kolawole E, Duggirala A, Gronow O, et al. Differential expression of maternal plasma microRNAs and their respective gene targets can predict early fetal growth restriction. Life. 2025;15(2):167. doi:10.3390/life15020167
51. Zhang JJ. Type 2 diabetes is involved in vascular aging mechanism. J Med Inform. 2022;35(11):37–39.
52. Hu Q, Chi YF. Progress in autophagy of vascular endothelial cells in early severe burn. Chinese J Med. 2024;59(3):261–264.
53. Xiao CG, Li B, Zheng QY. Progress in research on the relationship between autophagy mechanism and inflammasome. Mod J Integr Traditional Chinese Western Med. 2019;28(18):2044–2047.
54. Qiang L, Yang S, Cui YH, et al. Keratinocyte autophagy enables the activation of keratinocytes and fibroblastsand facilitates wound healing. Autophagy. 2021;17(9):2128–2143. doi:10.1080/15548627.2020.1816342
55. Gao HX. Effect of Specific Knockout of Endothelial Cell Autophagy-Related Gene Atg7 on Wound Healing in Diabetic Mice. Guangdong: Jinan University; 2018.
56. Pan DG, Ke SK, Yin QH, et al. MoS2 nanoenzyme alleviates inflammatory endothelial cell injury by regulating mitochondrial dynamics and autophagy. Acta Pharmacol Sin. 2024;59(10):2791–2799.
57. Ren H, Zhao F, Zhang Q, et al. Autophagy and skin wound healing. Burns Trauma. 2022;10:tkac003. doi:10.1093/burnst/tkac003
58. Wei H, Yin Y, Yang W, et al. Nuciferine induces autophagy to relieve vascular cell adhesion molecule 1 activation via repressing the Akt/mTOR/AP1 signal pathway in the vascular endothelium. Front Pharmacol. 2023;14:1264324. doi:10.3389/fphar.2023.1264324
59. Hao D, Zhou DL, Yu XY, et al. The role of p38MAPK in Ox-LDL-induced vascular endothelial cell injury. Chinese J Evidence Based Cardiovasc Med. 2018;10(11):1343–1347.
60. Li YY, Zhao FH, Zhang KX, et al. Effects of hydrogen sulfide on autophagy and angiogenesis in diabetic rats. Chinese J Pathophysiol. 2023;39(12):2223–2233.
61. Zamani S, Salehi M, Abbaszadeh-Goudarzi G, et al. Evaluation effect of alginate hydrogel containing losartan on wound healing and gene expression. J Biomater Appl. 2025;39(7):762–788. doi:10.1177/08853282241292144
62. Shan SZ. Effect of AT1-AA on Platelet Activation and Its Mechanism. Shanxi: Shanxi Medical University; 2023.
63. Zheng A, Ma H, Liu X, et al. Effects of moist exposed burn therapy and ointment (MEBT/MEBO) on the autophagy mTOR signalling pathway in diabetic ulcer wounds. Pharm Biol. 2020;58(1):124–130. doi:10.1080/13880209.2019.1711430
64. Wang L, Liu G, Han Q, et al. The protective effect of erythracol extract on vascular endothelial cell injury induced by high sugar and high fat. Chin J Tissue Eng Res. 2023;27(26):4132–4138.
65. Debnath J, Gammoh N, Ryan KM. Autophagy and autophagy-related pathways in cancer. Nat Rev Mol Cell Biol. 2023;24(8):560–575. doi:10.1038/s41580-023-00585-z
66. Khawaja RR, Martín-Segura A, Santiago-FERNÁNDEZ O, et al. Sex-specific and cell-type-specific changes in chaperone-mediated autophagy across tissues during aging. Nat Aging. 2025;5(4):691–708. doi:10.1038/s43587-024-00799-6
67. Qi YR, Chao R, Fan XZ, Ming YY. Organelle-specific autophagy in inflammatory diseases: a potential therapeutic target underlying the quality control of multiple organelles. Autophagy. 2021;17(2):385–401. doi:10.1080/15548627.2020.1725377
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Research Hotspots and Frontier Trends of Autophagy in Diabetic Cardiomyopathy From 2014 to 2024: A Bibliometric Analysis
Yan M, Zhang S, Liang P, Huang H, Li G, A R, Wu H
Journal of Multidisciplinary Healthcare 2025, 18:837-860
Published Date: 13 February 2025
The Critical Role of Autophagy in the Pathogenesis of Diabetic Osteoporosis: Mechanisms and Therapeutic Measures
Hou K, Shi W, Xu K, Wang T, Zhang Y
Drug Design, Development and Therapy 2025, 19:9913-9942
Published Date: 6 November 2025