")
Back to Journals » Journal of Pain Research » Volume 15
Repeated Sevoflurane Exposure in Neonatal Rats Enhances the Sensitivity to Pain and Traumatic Stress Later in Juvenile Life
Authors Chen BZ, Jiang LH, Zhou W, Shang YC, Li F, Liu B
Received 6 March 2022
Accepted for publication 5 October 2022
Published 12 October 2022 Volume 2022:15 Pages 3171—3178
DOI https://doi.org/10.2147/JPR.S365253
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Qi Fang
Ben-Zhen Chen,1,2,* Li-Hua Jiang,3,* Wenqin Zhou,4 Yu-Chao Shang,4 Fang Li,1 Bin Liu1
1Department of Anesthesiology, West China Hospital, Sichuan University, Chengdu, People’s Republic of China; 2Department of Anesthesiology, Sichuan Provincial Women’s and Children’s Hospital, Chengdu, People’s Republic of China; 3Department of Operating Room Nursing, Sichuan Provincial People’s Hospital, University of Electronic Science and Technology of China, Chengdu, People’s Republic of China; 4Department of Anesthesiology, West China Second University Hospital, Sichuan University, Chengdu, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Bin Liu, Department of Anesthesiology, West China Hospital, Sichuan University, No. 37, Guo Xue Xiang, Chengdu, Sichuan, 610041, People’s Republic of China, Tel +86-13408669172, Email [email protected]
Purposeː: Sevoflurane exposure in the neonatal period of rodent animals was reported to be associated with neuroendocrine dysregulations later in life. We tested the hypothesis that repeated sevoflurane exposure in neonatal rats enhances the sensitivity to pain and acute traumatic stress response later in juvenile life and investigated whether the neonatal brain depolarizing γ-aminobutyric acid type A receptor (GABAAR) activity is involved in mediating these abnormalities.
Methodsː: The postnatal 6 days (P6) Sprague-Dawley male rat pups pretreated with vehicle or the NKCC1 inhibitor, bumetanide, received sequential exposures to 2.1% sevoflurane exposure for 2 hours daily in 3 consecutive days.
Resultsː: The results showed that repeated exposures to sevoflurane in neonatal rats significantly reduced the paw withdrawal thermal latency (PWTL) at P9, P45. Repeated exposures to sevoflurane in neonatal rats did not significantly affect the basal secretion of serum corticosterone at juvenile period P45, whereas the level of corticosterone for neonatal sevoflurane-exposed rats at P45 was significantly higher than the CON group after subject to conditioned fear traumatic stress (CFTS). The resulting NKCC1/KCC2 mRNA ratio was significantly increased immediately after the neonatal rats received the last sevoflurane exposure, which was alleviated by pretreated with the NKCC1 inhibitor bumetanide.
Conclusionː: Repeated exposures to sevoflurane in neonatal rats enhanced the sensitivity to pain and acute traumatic stress response in juvenile life. The neonatal brain depolarizing GABAAR activity is involved in mediating these abnormalities.
Keywords: sevoflurane, neonatal, exposure, paw withdrawal thermal latency, conditioned fear traumatic stress
Introduction
Traumatic stress is an inevitable mental reaction during hospitalization in children. A large number of uncontrollable or necessary medical procedures such as venipuncture, lumbar puncture, insertion of urinary tube or gastric tube, endotracheal intubation, surgical operations often result in nociceptive stimulus. Numerous clinical studies have demonstrated that failure to treat pain in children leads to short-term complications and long-term physiological, behavioral, cognitive sequelae including altered pain processing, attention deficit disorder, impaired visual-perceptual ability or visual-motor integration,1–3 and poor executive functions.4,5 Over 1 million children under the age of 4 years received general anesthesia in the United States every year.6 However, it is only in the last decade that we started to recognize that general anesthesia has deleterious effects on the developing brain.6 With arising concerns regarding the safe use of general anaesthesia in young children, there is an urgent demand to clearly understand the resulting functional changes in the brain in a comprehensive and clinically relevant manner.
Sevoflurane is currently one of the most commonly used inhalation anaesthetics in pediatric practice. Previous research have indicated that the developing brain is vulnerable to sevoflurane anesthesia. Fetal exposure to high concentrations or long durations of sevoflurane can induce widespread neurodegeneration and long-term learning or memory deficits.7–11 The first 2 postnatal weeks in rodents are characterized by high vulnerability to general anesthetics. A number of previous studies have shown that the corticosteroid-based stress response system and GABAergic signaling during development may be involved in mediating the adverse effects of neonatal exposure to anaesthetics.12,13 GABAAR (γ-aminobutyric acid type A receptor) is the main inhibitory receptor in the mature central nervous system, but it acts as an excitatory receptor in the early stage of neural development. GABAAR is a ligand-gated ion channel receptor, which is coupled with the chloride ion channel. CCC (Cation Chloride Cotransporter) is the key controlling factor in controlling the switch of the GABAAR function.14 Cl−(chloride ion) is the main charge carrier through GABAAR channels,15 which is mainly regulated by the inwardly directed NKCC1 (Sodium Potassium Chloride Cotransporter 1) and the outwardly directed KCC2 (Potassium Chloride Cotransporter 2). In immature neurons, NKCC1 increases the chloride reversal potential; thus, it accumulates Cl− inside the cell. KCC2, on the other hand, reduces the chloride reversal potential; thus, it extrudes Cl− out of the cell and shifts the actions of the GABA from excitation to inhibition. Interfering with the balance of NKCC1/KCC2 may cause excessive excitement of the circuit and lead to neurodevelopmental disorders. NKCC1 and KCC2 are sensitive to neuronal damage, and the imbalance of their expression is thought to cause a variety of neuropathic diseases.16 Sevoflurane exposure enhances GABAAR activity in immature neurons, which induces increased GABAAR-mediated depolarization and corticosteroid levels.17 The excitatory effects of GABAAR play an important role in synaptic formation and neural plasticity. The neuroendocrine system may be involved in sevoflurane-induced neurotoxicity through GABAAR. As an important part of the neuroendocrine system, the limbic-hypothalamic-pituitary-adrenal (LHPA) axis plays an important role in the development of the nervous system.18 According to these studies, the safe use of sevoflurane in pediatric anesthesia and sedation still holds the attention of the public and anaesthesiologists. Thus, in this study, we tested the hypothesis that repeated sevoflurane exposure in neonatal rats enhances the sensitivity to pain and stress response later in juvenile life and investigated whether the neonatal brain depolarizing receptor (GABAAR) activity is involved in mediating these abnormalities.
Materials and Methods
Animals
The current study had been approved by the Ethics Committee of Sichuan Provincial Women’s and Children’s Hospital, Chengdu, China and had been performed in accordance with the ARRIVE guidelines and the Guide for the Care and Use of Laboratory Animals from the National Institutes of Health (Bethesda, MD, USA). Sprague-Dawley (SD) rats were obtained from the Animal Center of Chengdu Medical College (Chengdu, China), and were housed individually in standard conditions with a 12-h light/dark cycle (light from 08:00 to 20:00) at constant incubator (24±1°C, 30% humidity) and ad libitum access to water and rodent chow. To control for the litter variability, several pups from different litters were used for each treatment condition. Multiple sets of animals were used in the experiments. After the experiment, rats were anaesthetized with pentobarbital (70 mg/kg, i.p.) and then euthanized by decapitation.
Experimental Groups and Sevoflurane Exposure
In the current experiment, three groups were divided. The postnatal 6 days (P6) male rat pups were divided randomly to the SEV (sevoflurane) or SEV+BUM (sevoflurane plus bumetanide) or CON (control, not exposed to sevoflurane) groups. The SEV group was exposed to 2.1% sevoflurane in O2/N2 (FiO2 50%) for 2 hours daily in three successive days, in a thermostated box set to 37±1°C. The total gas flow was set at 2 L·min−1. The concentrations of sevoflurane and oxygen were measured continuously using a calibrated Datex side stream analyser that sampled from the interior of the chamber. The rats kept breathing spontaneously. After receiving sevoflurane exposure, the rat pups recovered consciousness and were returned to the mothers on gaining the righting reflex. In the current study, arterial blood for gas analysis demonstrated that 2.1% sevoflurane exposure for 2 h does not significantly change blood gas values and physiological parameters (data not shown). To study the role of GABAAR-mediated depolarization/activation at the time of repeated neonatal sevoflurane exposures, rat pups in the SEV+BUM group received an injection of the NKCC1 inhibitor, bumetanide 1.82 mg·kg−1, intraperitoneally), 15 min prior to receiving the sevoflurane exposure per day. Bumetanide in this concentration/dose range is widely used as the most selective of currently available inhibitors of NKCC1.19,20 The CON group were separated from the dams for identical conditions (37 ± 1°C, FiO2 50%), except for exposure to sevoflurane. In order to control for injections of bumetanide prior to exposure, rat pups in the CON group and SEV group received equal volumes of vehicle (VEH, i.p.) saline at P6, P7 and P8.
Paw Withdrawal Thermal Latency (PWTL)
The measurement of PWTL was based on a previous study.21 The experimental rats in each subgroup (n = 10) at P9, P45 were placed on the surface of a 2-mm-thick glass plate in a chamber, then the sensitivity of rat to heat stimuli with a RTY-3 radiant heat stimulator was tested (Xi’an Fenglan Instrumental Factory, Xi’an, P.R. China; 10 V). The experimental rats have been allowed to acclimate for 15 minutes. Testing started once the rat became largely inactive. The heat stimuli was given to a hind paw of the experimental rats. The latency was recorded as the duration starting from the heat stimuli to the occurrence of a hind paw withdrawal reflex. Three heat stimuli were repeatedly given to each rat and the mean PWTL was obtained. The inter-stimulus interval for each heat stimuli was 5 min. Rats after the measurement of PWTL were not used in any other experiments and were euthanized.
Conditioned Fear Traumatic Stress (CFTS)
Fear conditioning system (TSE, Bad Homburg, Germany) was employed to establish the model of CFTS at P45. Contextual fear conditioning took place in a box, which contains a loud speaker which can generate 4.5k Hz, 60 dB sound signal. The floor of the box consists of 28 stainless steel bars (6 millimeter in diameter and 12 millimeter apart), through which foot shock could happen. All P45 rats were put into the conditioned fear chamber, acclimated to the environment for 5 min, and then the sound signal (4.5k Hz, 60 Db, 30 s) occurred intermittently for 6 times with an interval of 2 min. At the end of every sound signal, the experimental rats were paired with a inescapable foot shock (1.2 mA, 5 s). After the last inescapable foot shock, the experimental rats were taken out for measurement of serum corticosterone.
Measurement of Serum Corticosterone
Serum corticosterone of rats was measured using commercial ELISA kits (Cayman Chemical Company, Ann Arbor, MI). In order to explore long-term effects of repeated neonatal sevoflurane exposures on endocrine responses to stress during juvenile period, a subgroup rats of each experiment group (n = 6) were used for measurement of basal serum levels of corticosterone 30 min before the process of CFTS at P45. Another subgroup rats of each experiment group (n = 6) were used for measurement of serum levels of corticosterone 5 min after establishment of model of CFTS. Blood sampling was collected using the “tail clip” method. Specifically, the distal 0.5 mm of the tail was removed using a sterile scalpel blade, and blood was allowed to drain directly into a microcentrifuge tube. These rats used for for measurement of serum levels of corticosterone were not used in any other experiments since the blood sampling was a terminal procedure.
Analyses of mRNA Levels for NKCC1 and K+-2Cl− (KCC2) in the Hypothalamus
Immediately after receiving the last sevoflurane exposure for neonatal rats, a subgroup rats of each experiment group (n = 6) were used for assay of the mRNA levels (NKCC1, KCC2) in the hypothalamus by qRT-PCR (real-time reverse transcription-PCR) in a StepOnePlus™ Real-Time PCR System (Applied Biosystems, CA, USA). RNA was obtained from the samples employing an RNeasy Plus Kit (Qiagen, CA, USA), and using reverse-transcribed with a high-capacity cDNA reverse transcription kit (Bio-Rad Laboratories, Hercules, CA, USA), and then assayed via qRT-PCR. TaqMan probes and oligonucleotide primers specific for the study genes were gained from Applied Biosystems (Carlsbad, CA, USA):NKCC1 (Rn00582505_m1) and KCC2 (Rn00592624_m1). Data were normalized to GAPDH (glyceraldehyde-3-phosphate dehydrogenase) mRNA (Rn01775763_g1). Gene expression was analyzed using the ΔΔCT method, and the data were expressed as relative fold change from that of the control group.
Statistical Analysis
Values are reported as mean (SEM). SPSS (version 17.0) was used for statistical analysis. Distribution of data was analysed using the Kolmogorov–Smirnov test. Student’s t-test was used for single comparisons and multiple comparisons among groups were analysed using one-way analysis of variance (ANOVA) followed by post hoc Bonferroni test. A P value less than 0.05 was considered statistically significant.
Results
Neonatal Repeated Sevoflurane Exposure Enhances the Sensitivity to Pain Later in Juvenile Life
In order to evaluate the neonatal repeated sevoflurane exposure in rats on the sensitivity to pain at juvenile period, the PWTL was measured at P9, P45. One-way ANOVA showed repeated exposures to sevoflurane in neonatal rats significantly reduced the PWTL at P9 and P45 compared with the CON group (P<0.01, Figure 1). Pretreated with the NKCC1 inhibitor bumetanide before neonatal sevoflurane exposures increased the paw withdrawal thermal latency compared with the SEV group. These suggested repeated exposures to sevoflurane in neonatal rats enhanced the sensitivity to pain later in life. The neonatal brain depolarizing GABAAR activity might be involved in mediating these abnormalities.
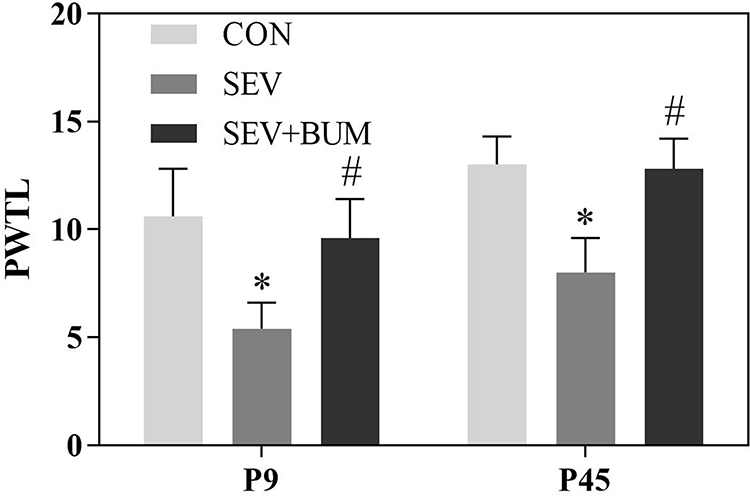 |
Figure 1 Neonatal repeated sevoflurane exposure enhances the sensitivity to pain later in juvenile life, Pretreatment with bumetanide prior to neonatal sevoflurane exposure alleviated this response. PWTL: The paw withdrawal thermal latency. (mean ± SEM; n = 10 rats/group; *p < 0.05 vs CON group; #p < 0.05 vs SEV group, one-way ANOVA). |
Repeated Exposures to Sevoflurane in Neonatal Rats Did Not Significantly Affect the Basal Secretion of Serum Corticosterone at Juvenile Period, but the Serum Corticosterone Levels Under Stress Conditions Were Significantly Enhanced
In order to evaluate the neonatal repeated sevoflurane exposures in rats on the stress response at juvenile period, the basal secretion of serum corticosterone was tested at P45. One-way ANOVA showed repeated exposures to sevoflurane in neonatal rats did not significantly affect the basal secretion of serum corticosterone at juvenile period (Figure 2).
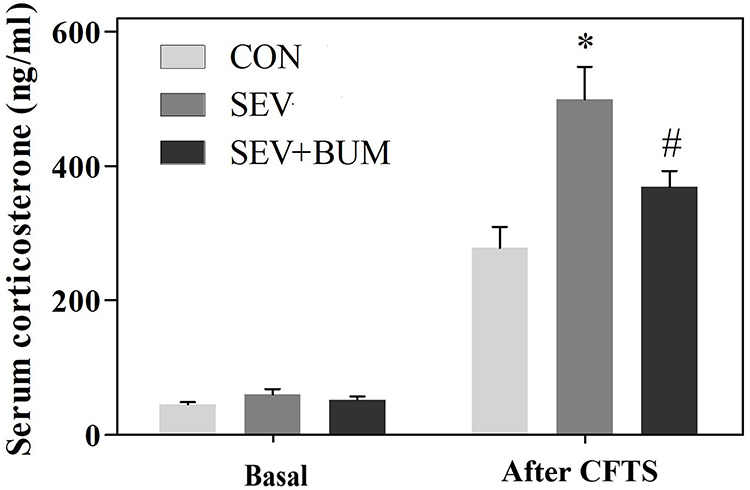 |
Figure 2 Neonatal repeated exposure to sevoflurane in rats enhanced the secretion of corticosterone in the juvenile CFTS model, while the basal secretion of serum corticosterone was not significantly affected at juvenile period (mean ± SEM; n = 6 rats/group; *p < 0.05 vs CON group; #p < 0.05 vs SEV group, one-way ANOVA). |
In order to assess whether neonatal exposure to sevoflurane alters endocrine responses to stress in the CFTS model during juvenile period, serum corticosterone levels under basal and stress condition were measured in blood samples collected from P45 rats 30 min before the process of CFTS and 5 min after subject to CFTS. The results showed that neonatal sevoflurane-exposed rats had unaltered secretion of corticosterone under the basal condition before the process of CFTS at P45; however, the level of corticosterone 5 min after subject to CFTS for neonatal sevoflurane-exposed rats was significantly higher than the CON group ((P < 0.01; Figure 2). Pretreatment of neonatal rats prior to each sevoflurane exposure with bumetanide significantly attenuated the heightened secretion of corticosterone after subject to CFTS ((P < 0.01; Figure 2).
Repeated Sevoflurane Exposures in Neonatal Rats at P6, P7, P8 Increases the NKCC1/KCC2 mRNA Ratio in the Hypothalamus of Rat Pups, Which Was Alleviated by Pretreated with the NKCC1 Inhibitor Bumetanide
The authors investigated whether the neonatal brain depolarizing GABAAR was altered immediately after the neonatal rats receiving the last sevoflurane exposure by examining the expression of NKCC1/KCC2 mRNA ratio in the hypothalamus of rat pups. Repeated exposures to sevoflurane at P6, P7, P8 increased the expression of NKCC1 mRNA in the hypothalamus (P < 0.01; Figure 3A), which was alleviated by pretreating with the NKCC1 inhibitor bumetanide (P < 0.01; Figure 3A). Repeated exposures to sevoflurane at P6, P7, and P8 had no significant effect on the expression of KCC2 mRNA levels in the hypothalamus (Figure 3B). The resulting NKCC1/KCC2 mRNA ratio was significantly increased immediately after the neonatal rats received the last sevoflurane exposure (P < 0.01; Figure 3C), which was alleviated by pretreating with the NKCC1 inhibitor bumetanide (Figure 3C). These results indicated the brain depolarizing GABAAR activity might be increased after neonatal rat pubs were exposed to repeated sevoflurane exposure, which could be attenuated by pretreating with the NKCC1 inhibitor bumetanide.
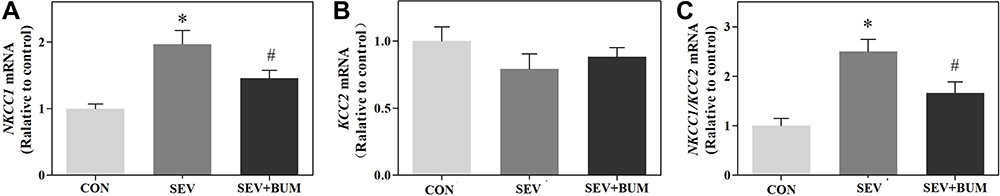 |
Figure 3 Repeated sevoflurane exposures in neonatal rats at P6, P7, P8 increases the NKCC1/KCC2 mRNA ratio in the hypothalamus of rat pups, which was alleviated by pretreated with the NKCC1 inhibitor bumetanide. (A) Repeated sevoflurane exposures in neonatal rats at P6, P7, P8 increased the expression of NKCC1 mRNA in the hypothalamus. (B) Repeated sevoflurane exposures in neonatal rats at P6, P7, P8 had no significant effects on expression of KCC2 mRNA in the hypothalamus. (C) Repeated sevoflurane exposures in neonatal rats at P6, P7, P8 increases the NKCC1/KCC2 mRNA ratio in the hypothalamus of rat pups. (mean ± SEM; n = 6 rats/group; *p < 0.05 vs CON group; #p < 0.05 vs SEV group, one-way ANOVA). |
Discussion
In this study, we tested the hypothesis that neonatal repeated exposures to sevoflurane increased the sensitivity to pain and stress response later in juvenile life, and investigated whether the neonatal brain depolarizing GABAAR was involved in mediating these abnormalities. The results showed repeated exposures to sevoflurane in neonatal rats significantly reduced the PWTL at P9, P45. The brain depolarizing GABAAR activity was enhanced by elevated NKCC1/KCC2 mRNA ratio in the hypothalamus. Repeated exposures to sevoflurane in neonatal rats did not significantly affect the basal secretion of serum corticosterone at juvenile period, whereas, by using the rat model of CFTS induced by electric foot shocks during the juvenile period, we showed that the secretion level of corticosterone 5 min after the establishment of CFTS in neonatal sevoflurane-exposed rats was significantly higher than the CON group. Pretreatment of neonatal rats prior to sevoflurane exposures with NKCC1 inhibitor, bumetanide, ameliorated most of these effects. Our study indicated that repeated exposures to sevoflurane in neonatal rats increased the sensitivity to pain and acute stress response later in juvenile life.
With the progress of medical technology, greater numbers of neonates and infants receive surgery, interventions, or examination under general anesthesia or sedation at an early stage after birth. Anesthesia or sedation is required for a large number of medical examinations and procedures in pediatric outpatients and inpatients which are non-invasive and do not cause obvious surgical trauma, but require immobilization. Sevoflurane has become one of the most commonly used general anesthetics in pediatrics due to its characteristics of small respiratory tract irritation, rapid induction and recovery, and stable hemodynamics. Sevoflurane, in particular multiple or prolonged exposures, has been reported to be neurotoxic in the developing brain in animal studies and lead to abnormal neurological development including attention-deficit/hyperactivity disorder (ADHD).8–10,22,23 The causative links between neonatal sevoflurane exposure and later life neurocognitive dysfunctions were always the subject of heat dispute. So, we established “repeated sevoflurane exposure” model in neonatal rats without surgical procedures in our study. In the present study, repeated sevoflurane exposures in neonatal rats did not significantly affect the basal secretion of serum corticosterone level later in juvenile life. However, the authors showed that repeated exposures to sevoflurane in neonatal rats enhanced the sensitivity to pain and secretion of corticosterone in the CFTS model. These results suggested a possibility the later life neurocognitive dysfunctions were not solely driven by the effect of early life sevoflurane exposure, the adverse effects of neonatal anaesthesia in rodents might be associated with post-anaesthesia life experiences. Thus, we proposed the neurocognitive dysfunctions might be the result of a cumulative impact initially programmed by neonatal sevoflurane exposure and later exacerbated by adverse stressful factors such as trauma and various types of psychosocial stress.
During brain development, GABAergic synapses are formed prior to the formation of glutamatergic synapses and the activation of the GABAAR depolarizes immature neurons.24,25 NKCC1 was reported to play a critical role in regulating the function of brain GABAergic transmission and modulating the vulnerability to maternal separation-induced susceptibility to later life stress.16 Disturbing NKCC1/KCC2 balance was associated with brain circuit hyperexcitability and contributed to a wide spectrum of stress‐related neurocognitive dysfunctions.26–29 In the present study, the NKCC1/KCC2 mRNA ratio was increased in the hypothalamus immediately after neonatal rats receiving the last sevoflurane exposure at P8, the stress susceptibility exhibited in the CFTS model during juvenile period was significantly increased for neonatal sevoflurane-exposed rats, which was alleviated by administration of NKCC1 inhibitor bumetanide before neonatal sevoflurane exposures. We proposed the mechanism that the enhanced depolarizing GABAAR activity after neonatal repeated sevoflurane exposures at P6-P8 contributed to the dysregulation of the neuroendocrine response and increased the risk of stress exposure later in life. The alleviating effect of bumetanide on neuroendocrine dysregulation in the CFTS model for neonatal sevoflurane-exposed rats is that bumetanide, by reversing GABA-initiated signaling from depolarizing to inhibitory, facilitates adaptation of the neonatal brain to environmental stressful factors later in life.
Conclusion
Our current findings indicated that repeated exposures to sevoflurane in neonatal rats enhanced the sensitivity to pain and acute traumatic stress response in juvenile life. The neonatal brain depolarizing GABAAR activity in the hypothalamus might be involved in mediating these abnormalities.
Ethics Approval
The present study was approved by the Ethics Committee of Sichuan Provincial Women’s and Children’s Hospital, and was conducted in accordance with the ARRIVE guidelines and the Guide for the Care and Use of Laboratory Animals from the National Institutes of Health (Bethesda, MD, USA).
Author Contributions
All authors read and approved the final version of the manuscript. All authors contributed to data analysis, drafting or revising the article, have agreed on the journal to which the article will be submitted, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
This work was supported by the grants from the National Natural Science Foundation of China (Nos. 81701090) and Key research project of Health Commission of Sichuan Province (Grant Number: 19ZD009).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Ferguson SA, Ward WL, Paule MG, et al. A pilot study of preemptive morphine analgesia in preterm neonates: effects on head circumference, social behavior, and response latencies in early childhood. Neurotoxicol Teratol. 2012;34(1):47–55. doi:10.1016/j.ntt.2011.10.008
2. de Graaf J, van Lingen RA, Simons SH, et al. Long-term effects of routine morphine infusion in mechanically ventilated neonates on children’s functioning: five-year follow-up of a randomized controlled trial. Pain. 2011;152:1391–1397. doi:10.1016/j.pain.2011.02.017
3. Perry M, Tan Z, Chen J, et al. Neonatal pain perceptions and current practice. Crit Care Nurs Clin North Am. 2018;30(4):549–561. doi:10.1016/j.cnc.2018.07.013
4. Anand KJS, Palmer FB, Papanicolaou AC. Repetitive neonatal pain and neurocognitive abilities in ex-preterm children. Pain. 2013;154:1899–1901. doi:10.1016/j.pain.2013.06.027
5. Doesburg SM, Chau CM, Cheung TP, et al. Neonatal pain-related stress, functional cortical activity and visual-perceptual abilities in school-age children born at extremely low gestational age. Pain. 2013;154:1946–1952. doi:10.1016/j.pain.2013.04.009
6. Rabbitts JA, Groenewald CB, Moriarty JP, et al. Epidemiology of ambulatory anesthesia for children in the United States: 2006 and 1996. Anesth Analg. 2010;111:1011–1015. doi:10.1213/ANE.0b013e3181ee8479
7. Chung W, Park S, Hong J, et al. Sevoflurane exposure during the neonatal period induces long-term memory impairment but not autism-like behaviors. Paediatr Anaesth. 2015;25(10):1033–1045. doi:10.1111/pan.12694
8. Liu B, Gu Y, Xiao H, et al. Altered metabolomic profiles may be associated with sevoflurane-induced neurotoxicity in neonatal rats. Neurochem Res. 2015;40(4):788–799. doi:10.1007/s11064-015-1529-x
9. Amrock LG, Starner ML, Murphy KL, et al. Long-term effects of single or multiple neonatal sevoflurane exposures on rat hippocampal ultrastructure. Anesthesiology. 2015;122(1):87–95. doi:10.1097/ALN.0000000000000477
10. Zhang X, Liu S, Newport GD, et al. In vivo monitoring of sevoflurane induced adverse effects in neonatal nonhuman primates using small-animal positron emission tomography. Anesthesiology. 2016;125(1):133–146. doi:10.1097/ALN.0000000000001154
11. Yu Y, Yang Y, Tan H, et al. Tau contributes to sevoflurane-induced neurocognitive impairment in neonatal mice. Anesthesiology. 2020;133:595–610. doi:10.1097/ALN.0000000000003452
12. Cao W, Pavlinec C, Gravenstein N, et al. Roles of aldosterone and oxytocin in abnormalities caused by sevoflurane anesthesia in neonatal rats. Anesthesiology. 2012;117:791–800. doi:10.1097/ALN.0b013e318266c62d
13. Crowley SK, Girdler SS. Neurosteroid, GABAergic and hypothalamic pituitary adrenal (HPA) axis regulation: what is the current state of knowledge in humans? Psychopharmacology. 2014;231:3619–3634. doi:10.1007/s00213-014-3572-8
14. Kaila K, Price TJ, Payne JA, et al. Cation-chloride cotransporters in neuronal development, plasticity and disease. Nat Rev Neurosci. 2014;15:637–654. doi:10.1038/nrn3819
15. Salmon CK, Pribiag H, Gizowski C, et al. Depolarizing GABA transmission restrains activity-dependent glutamatergic synapse formation in the developing hippocampal circuit. Front Cell Neurosci. 2020;14:36. doi:10.3389/fncel.2020.00036
16. Cabrera OH, Tesic V, Tat QL, et al. Sevoflurane-induced dysregulation of cation-chloride cotransporters NKCC1 and KCC2 in neonatal mouse brain. Mol Neurobiol. 2019;57:1–10. doi:10.1007/s12035-019-01751-1
17. Xu C, Tan S, Zhang J, et al. Anesthesia with sevoflurane in neonatal rats: developmental neuroendocrine abnormalities and alleviating effects of the corticosteroid and Cl−importer antagonists. Psychoneuroendocrinology. 2015;60:173–181. doi:10.1016/j.psyneuen.2015.06.016
18. Hankin BL, Badanes LS, Smolen A, et al. Cortisol reactivity to stress among youth: stability over time and genetic variants for stress sensitivity. J Abnorm Psychol. 2015;124:54–67. doi:10.1037/abn0000030
19. Quirk GJ, Garcia R, González-Lima F, et al. Prefrontal mechanisms in extinction of conditioned fear. Biol Psychiatry. 2006;60:337–343. doi:10.1016/j.biopsych.2006.03.010
20. Liu R, Wang J, Liang S, et al. Role of NKCC1 and KCC2 in epilepsy: from expression to function. Front Neurol. 2020;10:1407. doi:10.3389/fneur.2019.01407
21. Li KC, Chen J. Altered pain-related behaviors and spinal neuronal responses produced by s.c. injection of melittin in rats. Neuroscience. 2004;126:753–762. doi:10.1016/j.neuroscience.2004.03.050
22. Yang J, Lingsha J, Jia M, et al. Subsequent maternal separation exacerbates neurobehavioral abnormalities in rats neonatally exposed to sevoflurane anesthesia. Neurosci Lett. 2017;661:137–142. doi:10.1016/j.neulet.2017.09.063
23. Ju L-S, Yang -J-J, Gravenstein N, et al. Role of environmental stressors in determining the developmental outcome of neonatal anesthesia. Psychoneuroendocrinology. 2017;81:96–104. doi:10.1016/j.psyneuen.2017.04.001
24. Ben-Ari Y, Gaiarsa J-L, Tyzio R, et al. GABA: a pioneer transmitter that excites immature neurons and generates primitive oscillations. Physiol Rev. 2007;87(4):1215–1284. doi:10.1152/physrev.00017.2006
25. Tang X, Jaenisch R, Sur M. The role of GABAergic signalling in neurodevelopmental disorders. Nat Rev Neurosci. 2021;22(5):290–307. doi:10.1038/s41583-021-00443-x
26. Yang J, Ju L, Yang C, et al. Effects of combined brief etomidate anesthesia and postnatal stress on amygdala expression of Cl− cotransporters and corticotropin-releasing hormone and alcohol intake in adult rats. Neurosci Lett. 2018;685:83–89. doi:10.1016/j.neulet.2018.08.019
27. Chew TA, Orlando BJ, Zhang J, et al. Structure and mechanism of the cation-chloride cotransporter NKCC1. Nature. 2019;572(7770):488–492. doi:10.1038/s41586-019-1438-2
28. Delpire E, Gagnon KB. Na+-K+-2Cl− cotransporter (NKCC) physiological function in nonpolarized cells and transporting epithelia. Compr Physiol. 2018;8:871–901. doi:10.1002/cphy.c170018
29. Monette MY, Somasekharan S, Forbush B. Molecular motions involved in Na-K-Cl cotransporter-mediated ion transport and transporter activation revealed by internal cross-linking between transmembrane domains 10 and 11/12. J Biol Chem. 2014;289:7569–7579. doi:10.1074/jbc.M113.542258
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.