Back to Journals » Infection and Drug Resistance » Volume 13
Rapid Identification of KL49 Acinetobacter baumannii Associated with Clinical Mortality
Authors Deng Q, Zhang J ![]() , Zhang M, Liu Z, Zhong Y, Liu S
, Zhang M, Liu Z, Zhong Y, Liu S ![]() , Cui R, Shi Y, Zeng H, Yang X, Lin C, Luo Y
, Cui R, Shi Y, Zeng H, Yang X, Lin C, Luo Y ![]() , Chen H, Wu W, Wu J, Zhang T, Lu Y, Liu X, Zou Q, Huang W
, Chen H, Wu W, Wu J, Zhang T, Lu Y, Liu X, Zou Q, Huang W ![]()
Received 26 August 2020
Accepted for publication 21 October 2020
Published 12 November 2020 Volume 2020:13 Pages 4125—4132
DOI https://doi.org/10.2147/IDR.S278891
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Qiuyang Deng,1,* Jinyong Zhang,2,* Min Zhang,1 Zhou Liu,3 Yuxin Zhong,1 Shiyi Liu,1 Ruiqin Cui,1 Yun Shi,2 Hao Zeng,2 Xiyao Yang,3 Chuchu Lin,4 Yutian Luo,5 Huaisheng Chen,5 Weiyuan Wu,6 Jinsong Wu,6 Tianle Zhang,7 Yuemei Lu,6 Xueyan Liu,5 Quanming Zou,2 Wei Huang1
1Bacteriology & Antibacterial Resistance Surveillance Laboratory, Shenzhen Institute of Respiratory Diseases, Shenzhen People’s Hospital, The Second Clinical Medical College, Jinan University, The First Affiliated Hospital, Southern University of Science and Technology, Shenzhen, Guangdong 518020, People’s Republic of China; 2National Engineering Research Center of Immunological Products, Department of Microbiology and Biochemical Pharmacy, College of Pharmacy, Army Medical University, Chongqing, People’s Republic of China; 3Second Affiliated Hospital of Anhui Medical University, Hefei, People’s Republic of China; 4Department of Nosocomial Infection Control, Shenzhen People’s Hospital, The Second Clinical Medical College, Jinan University, The First Affiliated Hospital, Southern University of Science and Technology, Shenzhen, Guangdong 518020, People’s Republic of China; 5Intensive Care Unit, Shenzhen People’s Hospital, The Second Clinical Medical College, Jinan University, The First Affiliated Hospital, Southern University of Science and Technology, Shenzhen, Guangdong 518020, People’s Republic of China; 6Department of Clinical Microbiology, Shenzhen People’s Hospital, The Second Clinical Medical College, Jinan University, The First Affiliated Hospital, Southern University of Science and Technology, Shenzhen, Guangdong 518020, People’s Republic of China; 7Guangdong Medical University, Dongguan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Wei Huang
Bacteriology & Antibacterial Resistance Surveillance Laboratory, Shenzhen Institute of Respiratory Diseases, Shenzhen People’s Hospital, The Second Clinical Medical College, Jinan University, The First Affiliated Hospital, Southern University of Science and Technology, Shenzhen, Guangdong 518020, People’s Republic of China
Tel +86 7522942513
Fax +86 7522942514
Email [email protected]
Quanming Zou National Engineering Research Center of Immunological Products, Department of Microbiology and Biochemical Pharmacy
College of Pharmacy, Army Medical University, Chongqing 400038, People’s Republic of China
Tel/Fax +86 2368752316
Email [email protected]
Objective: We aimed to establish a tool for rapid identification of KL49 Acinetobacter baumannii.
Methods: Based on the capsular polysaccharide (CPS) synthesis genes database, we investigated the distribution of K locus type 49 (KL49) genes in other KL types and established a rapid identification method for KL49. We collected 61 clinical carbapenem-resistant A. baumannii (CRAB) strains, identified KL49 by gtr100 detection, and used whole genome sequencing (WGS) for verification. A mouse pneumonia model was used to confirm the hypervirulence phenotype. We tested the presence of gtr100 gene in 165 CRAB strains from three provinces in China and evaluated the correlation of gtr100 carrying CRAB infection with mortality.
Results: The gtr100 gene is the CPS synthesis gene found only in KL49. We screened out nine WGS-validated KL49 strains from 61 CRAB clinical strains using polymerase chain reaction (PCR) to detect the gtr100 gene. The survival rates of KL49 strains were significantly lower than nonKL49 strains in a mouse pneumonia model. The survival rates of LAC-4 gtr100 knockout strain decreased significantly. Analysis of phylogenetics showed the worldwide spread of KL49 A. baumannii. Infection of gtr100 carrying CRAB is an independent risk for mortality (OR, 10.76; 95%CI: 3.08– 37.55; p< 0.001).
Conclusion: The hypervirulence phenotype of KL49 CRAB and the association with mortality highlight the urgent need for implementing control measures. The rapid identification assay has the potential to facilitate early medical intervention and worldwide surveillance.
Keywords: Acinetobacter baumannii, hypervirulent, K locus, Kaptive, mortality
Introduction
Acinetobacter baumannii is a gram-negative coccobacillus that can cause serious infections among critically ill patients, particularly in the intensive care unit (ICU) setting.1 Worldwide, A. baumannii accounted for up to 20% of infections in ICUs.2
In general, A. baumannii has been regarded as a low-grade pathogen. Most laboratory strains and clinical strains do not cause severe infections in immunocompetent mice, inducing only a self-limiting pneumonia with very limited local bacterial replication and systemic dissemination, even when a large inoculum is used.3,4 However, two studies have shown that some clinical carbapenem-resistant A. baumannii (CRAB) strains are lethal to immunocompetent mice and thus revealed the range of virulence in different strains of this pathogen5,6 LAC-4 is a hypervirulent strain that killed 100% of mice within 48 h after being inoculated with 108 colony-forming units (CFU).6 Subsequently, there have been some reports about the outbreak and epidemic of A. baumannii with enhanced virulence7,8 However, based on multilocus sequence typing (MLST) genotyping, these hypervirulent strains are of different genotypes.7–9 Genomic analysis showed that these strains all shared a glycosylation gene cluster and type VI secretion system (T6SS), which was thought to be the potential virulence marker.7–9 Notably, the glycosylation gene cluster shared by these hypervirulent strains was part of K locus 49 (KL49) CPS gene cluster, suggesting that the KL49 strain needs to be taken seriously.
In this study, we established a method for rapid identification of the KL49 strain and successfully screened out the hypervirulent strains from clinical strains. The reliability of the method was verified by whole genome sequencing (WGS) and animal experiments. Finally, clinical data analysis showed that KL49 A. baumannii infection is an independent risk for mortality.
Methods
K Loci Analysis
Capsular polysaccharide (CPS) synthesis gene cluster analysis was performed using A. baumannii K locus database.10 All available A. baumannii genomes from NCBI (as of 2019-08-04) were downloaded. Kaptive (version 0.7.0) was used for KL type identification.10
Clinical Strains Collection and Phenotypic Characterization
Consecutive nonreplicate clinical strains of CRAB in the ICU of Shenzhen People’s Hospital in 2017 (n=61) were used for WGS and in vivo assay in this study. Shenzhen People’s Hospital is a medical center with 2500 beds in the Luohu district of Shenzhen, China. A VITEK-2 compact system (bioMérieux, Marcy-l’Étoile, France) was used to establish the strain identity and antimicrobial susceptibilities of the strains. All antimicrobial susceptibilities results were verified by broth microdilution method. The results were interpreted in accordance with the guidelines published by the Clinical and Laboratory Standards Institute (CLSI; document M100-S26).11 The species identity of all strains was confirmed via matrix-assisted laser desorption/ionization mass spectrometry (bioMérieux, Marcy-l’Étoile, France).
WGS, Genomic Characterization and Phylogenetic Analysis
Bacteria from frozen stocks were cultured in Luria–Bertani (LB) broth overnight at 37°C with shaking (220 rpm). Overnight cultures were diluted 1/100 and re-cultured until the optical density at 600 nm (OD600) was 0.6–0.8. Genomic DNA was extracted from each isolate using the SDS method.12 The bacterial genomes were sequenced using Illumina HiSeq 2500 platform (Illumina, San Diego, CA, USA), and fastp was used to remove low-quality and low-complexity reads, and polyG/polyX tails.13 The genomes were assembled with de novo SPAdes Genome Assembler (version 3.12.0).14 Virulence factors in the strains were identified by scanning the genome contigs against VFDB databases using ABRicate (version 0.8.7). Parsnp (version 1.5.2) was used to align the core genome and call the variants (SNP).15 Maximum likelihood tree was generated with RAxML v 7.0.4 using a GTRGAMMAI model.16 Online tool iTOL was used to display, manipulate, and annotate phylogenetic tree.17
gtr100 Deletion and Complement Mutant Construction
The LAC-4 knockout strain was constructed using the suicide vector pCVD442. Briefly, the apramycin resistance (Apr) gene and the flanking sequences of gtr100 was synthesized. The fragments were joined and cloned into the suicide plasmid pCVD442 to yield the plasmid pCVD442-Δgtr100:Apr, which was then transformed into Escherichia coli β2155, and then positive clones were selected on LB agar containing ampicillin (50 μg/mL) and apramycin (50 μg/mL). Next, conjugation between the recipient LAC-4 and the donor β2155/pCVD442-Δgtr100:Apr was performed to transfer the recombinant suicide plasmid pCVD442-Δgtr100:Apr from β2155 to LAC-4. Apramycin- and sucrose-resistant colonies were selected and screened by PCR.
The gtr100 complemented strain was constructed using the vector pWH1266. Briefly, the synthesized gtr100 sequence was joined and cloned into the plasmid pWH1266 to yield the plasmid pWH1266:gtr100 which was then transformed into E. coli DH5a. Positive clones were selected on LB agar containing ampicillin (50 μg/mL) and tetracycline (10 μg/mL). The isolated plasmid pWH1266:gtr100 was electroporated into LAC-4:Δgtr electrocompetent cells. The complemented LAC-4:Δgtr100Ωgtr100 strains were selected on LB agar with tetracycline (10 mg/mL) and confirmed by PCR.
Virulence Test in vivo
We used a pneumonia model of A. baumannii in mice to test the virulence of strains. Inbred C57BL/6 six to eight weeks old female mice under specificpathogen-free grade were purchased from Hunan SJA Laboratory Animal Co., Ltd (Hunan, China). The mice were intraperitoneally anesthetized with pentobarbital sodium (75 mg/kg) and inoculated with 20 μL (2.5×108, 5×107, 1×107, 0 CFU) of A. baumannii by noninvasive intratracheal instillation under direct vision. The survival of the mice was observed for seven days post infection.5 All animal care and use protocols in this study were performed in accordance with the Regulations for the Administration of Affairs Concerning Experimental Animals approved by the State Council of People’s Republic of China. All animal experiments in this study were approved by the Animal Ethical and Experimental Committee of the Army Military Medical University (Chongqing, Permit No. 2011–04) in accordance with their rules and regulations.
Clinical Data Collection and Analysis
CRAB clinical isolates (n=165) were collected from three provinces and used for univariable logistic regression analysis. We retrospectively used rapid identification tool of KL49 to test and collected demographic data, underlying diseases, sites of infection, comorbidities, invasive procedures, antibiotics usage and clinical characteristics of the corresponding patients. The PCR protocol for identifying the presence of gtr100 was as follows: 20 μL reaction containing 1.0 μL of 2×SanTaq PCR Mix (with blue dye); 1.0 μL of bacteria culture (OD600=0.6–0.8); 1.0 μL (10 μM) of forward primer, 1.0 μL (10 μM) of reverse primer; 7.0 μL of sterilized ddH2O. Amplification conditions were: denaturation at 95°C for four minutes; 95°C for 15 seconds, 60°C for 30 seconds, 72°C for 30 seconds (35 cycles); final extension at 72°C for five minutes. Taq DNA polymerase but not high fidelity and hotstart DNA polymerase was recommended. The primary outcome was all-cause in-hospital mortality following the onset of A. baumannii infection. Logistic regression was used for outcome analysis. Variables with a P-value ≤0.05 in the univariable regression were selected for multivariable analysis.
Results
By comparing the gene clusters of different KL in the Kaptive database, we found that only gtr100 gene was specific to KL49, suggesting that the detection of gtr100 has the potential to be used to identify KL49 (Table 1). We performed KL type analysis of all 3341 A. baumannii genomes retrieved from NCBI. Of the 80 genomes that contained the gtr100 gene, 79 were identified as KL49 and one as KL52. Sixteen genomes without the gtr100 gene were identified as KL49. However, 14 of these genomes showed “low” and “none” confidence levels called by Kaptive, probably due to the poor-quality sequence assembly (Table S1).
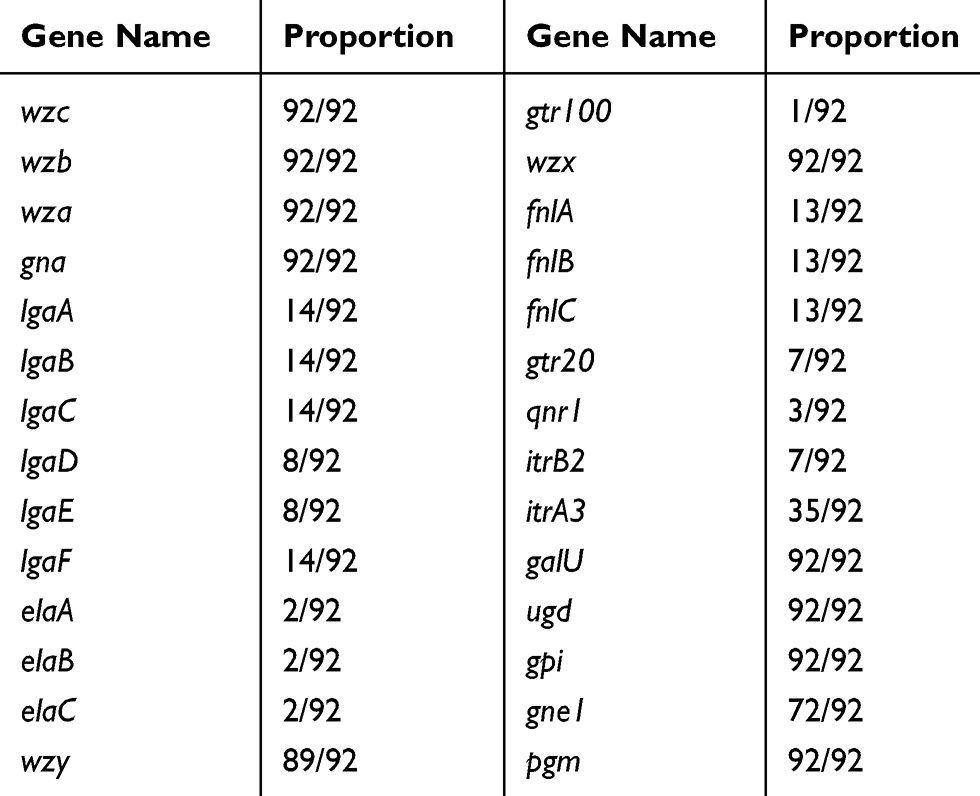 |
Table 1 Distribution of KL49 Capsular Polysaccharide (CPS) Synthesis Genes in Other KL Types |
Nonrepetitive CRAB (n=61) strains collected from patients with confirmed A. baumannii infections were frozen and stored by the clinical microbiology laboratory at the Shenzhen People’s Hospital. We then used forward primer (5ʹ-TTGAGAAGCTAAATTATGGCTCG-3ʹ) and reverse primer (5ʹ-GATAGCACAGAAATCCATAAAGGAA-3ʹ) to screen for the presence of gtr100, as a marker for KL49. Among them, the gtr100 gene was identified in nine of the strains by PCR assay. WGS results confirmed that the nine strains were KL49 (Table 2). Virulence gene analysis showed that most strains shared the same profile except: abaR was missing in isolates 5121, 5122, 5123, 5127, 2096, 2092 and 2093; bap was missing in isolate 2096; and hemO was missing in isolates 20881 and 20883 (Figure S1).
 |
Table 2 KL Types of 61 Clinical Carbapenem Resistant Acinetobacter baumannii Strains in This Study |
To verify the virulence differences between KL49 and nonKL49 strains, two KL49 strains (5122 and 2092) and three nonKL49 strains (7152, 71517 and 20859) were randomly selected to test their virulence in the mouse model (Figure 1). ATCC17978 and LAC-4 was used as control. With an inoculum of 2.5×108 CFU, the survival with strains 5122, 2092 and LAC-4 was 0% at 48 h, and at seven days was 60% with ATCC17978, 50% with 7152, 71517 and 20859. With an inoculum of 5×107 CFU, survival was 0% with LAC-4 at 48 h; 10% with 5122 and 2092, 80% with ATCC17978 and 7152, 60% with 71517, and 90% with 20859 at seven days. With an inoculum of 1×107 CFU, seven-day survival was 10% with LAC-4, 40% with 5122, 30% with 2092, 90% with 71517, 100% with ATCC17978 and 7152. The survival rates of KL49 strains (LAC-4, 5122 and 2092) were significantly lower than nonKL49 strains (7152, 71517, 20859 and ATCC17978) (Figure 1; P<0.0001 by log rank test). With the aim of identifying the role of gtr100 in the virulence of KL49 A. baumannii, we constructed the gtr100 knockout and complemented strain of LAC-4. As shown in Figure 1D, with an inoculum of 1×108 CFU, seven-day survival was 0% with LAC-4, 40% with gtr100 knockout strain and 10% with complemented strain (P=0.0121 by log rank test). With an inoculum of 1×107 CFU, seven-day survival was 20% with LAC-4, 100% with gtr100 knockout strain, and 40% with complemented strain (P=0.0005 by log rank test). The survival rates of gtr100 knockout strain were significantly higher than LAC-4 (1×107 CFU, P=0.0003; 1×108 CFU, P=0.0118) and the complemented strain (1×107 CFU, P=0.0040 by log rank test).
 |
Figure 1 Virulence potential of Acinetobacter baumannii strains in a mouse infection model. (A–D) The effect of 2.5×108, 1×108, 5×107, 1×107 colony-forming units of each A. baumannii isolate (n=10; *P<0.05, **P<0.001, log-rank test) on survival was assessed in mouse. Strains 5122 and 2092 are two KL49 A. baumannii strains; 7152, 71517 and 20859 are KL3 A. baumannii strains. LAC-4 is a KL49 hypervirulent A. baumannii strain reported in a previous study. LAC-4:Δgtr100 is a gtr100 knockout strain. LAC-4:Δgtr100Ωgtr100 is complemented strain. ATCC17978 is a KL3 A. baumannii strain. |
Eighty-seven KL49 assembly genomes were used for phylogenetic tree construction, including 78 retrieved from NCBI with a “perfect” or “very high” confidence level called by Kaptive and nine genomes in this study. In the phylogenetic tree, KL49 strains are divided into two distinct clades that are prevalent in Americas and Asia, respectively. However, two strains from China and Thailand were found in the Americas clade and one strain from the Czech Republic was found in the Asia clade, representing the worldwide spread of KL49, probably via human travel (Figure 2).
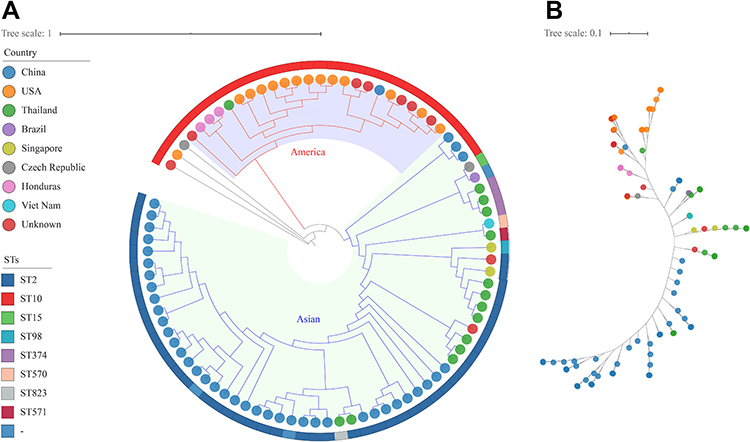 |
Figure 2 Worldwide spread of KL49 Acinetobacter baumannii strains. (A) A phylogenetic tree of the 78 KL49 strains retrieved from NCBI and nine KL49 strains in this study. LAC-4 was used as reference. The colors of the isolate tips represent the country of isolation. STs are shown in the outer ring. (B) An unrooted version of the tree shown in (A). |
A total of 165 clinical CRAB strains were collected from three provinces of China. All-cause in-hospital mortality rate of 63.6% (21/33) in patients with gtr100+ CRAB infection and 24.2% (32/132) in patients with gtr100− CRAB infection. In the univariable logistic regression analysis, the significant variables (P<0.05) associated with all-cause in-hospital mortality were coma, solid malignancy, mechanical ventilation, high level of urea nitrogen (>7.5 mmol/L), C-reactive protein (>10 mg/L), procalcitionin (>2 μg/L), sepsis, shock and gtr100+ CRAB infection (Table 3). Multivariable logistic regression analysis showed that the significant independent predictors for mortality were solid malignancy (OR, 13.66; 95%CI: 3.03–61.47; P=0.001), urea nitrogen (>7.5 mmol/L) (OR, 4.03; 95%CI: 1.31–12.39; P=0.015), sepsis (OR, 6.19; 95%CI: 1.61–23.84; P=0.008), Shock (OR, 5.17; 95%CI: 1.79–14.88; P=0.002) and gtr100+ CRAB infection (OR, 10.76; 95%CI: 3.08–37.55; P<0.001) (Table 4).
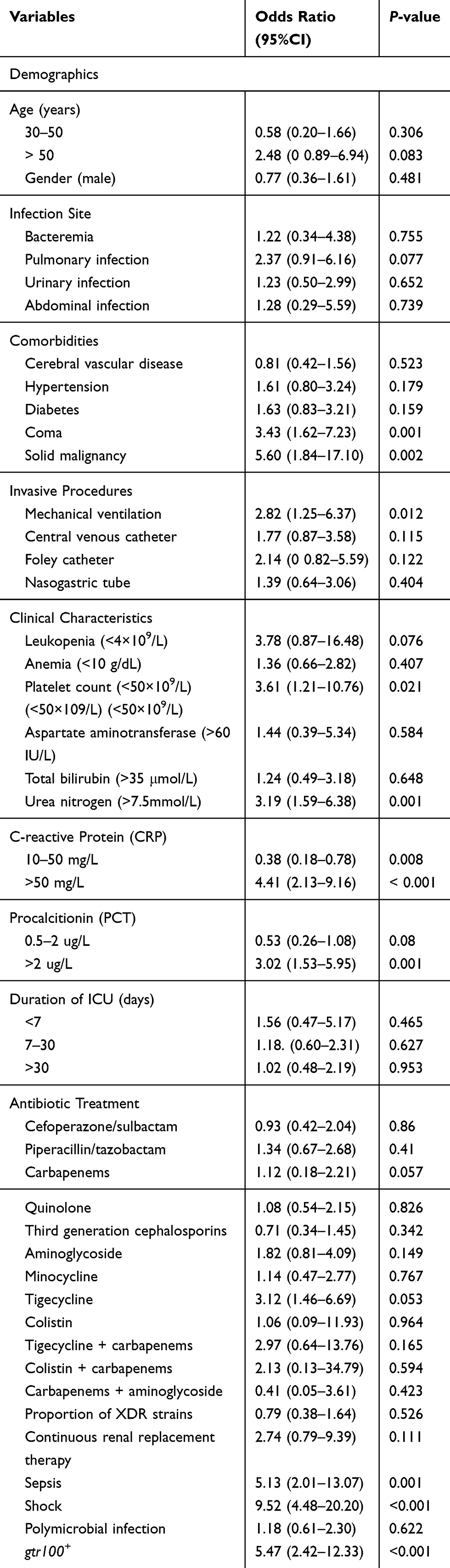 |
Table 3 Univariable Logistic Regression Analysis of the Factors Associated with Mortality |
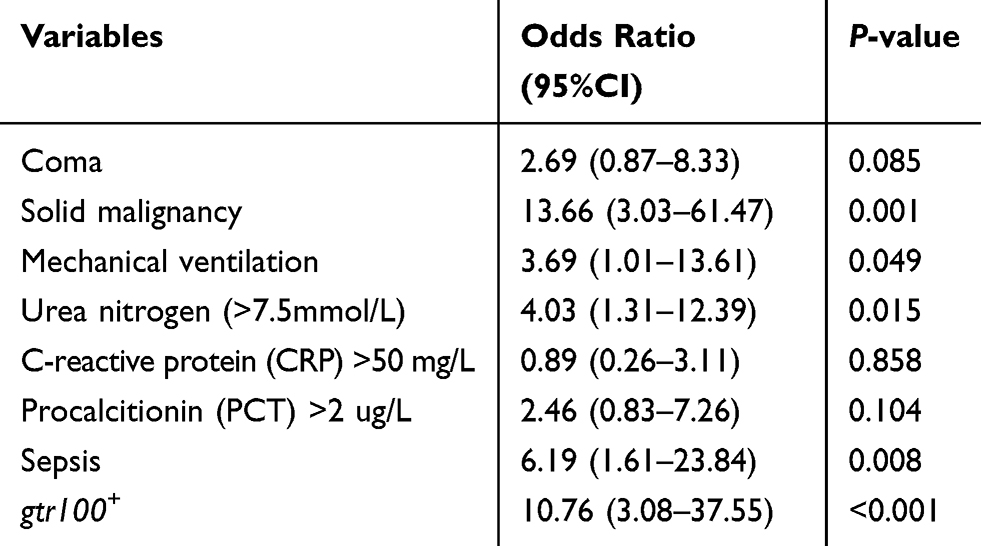 |
Table 4 Multivariable Logistic Regression Analysis of the Factors Associated with Mortality |
Discussion
The relationship between capsular type and bacterial virulence has been demonstrated in different bacteria. For Klebsiella pneumoniae, approximately 70% hypervirulent strains were of the K1 and K2.18–20
For A. baumannii, there have been many reports of KL49 strains being more virulent than others.6–8 High virulence phenotype of KL49 may be explained by the existence of an unusual sugar legionaminic acid, which can be used to mimic the host cell surface and benefit to escape from host immune surveillance.21 In this study, a glycosyltransferase encoding gene gtr100 is highlighted because it is specifically present in the CPS gene cluster of KL49, and is regarded as a good candidate for rapid identification of KL49. Moreover, animal experiment proved that gtr100 is crucial to the virulence of LAC-4, indicating that it has the potential to be a target of antivirulence.
Glycosyltransferases encoded by gtr genes are primarily responsible for the linkages between sugars in CPS. There are two gtr genes (gtr20 and gtr100) located in KL49 locus. gtr20 glycosyltransferase, also encoded in KL8, KL9, KL49, KL54, KL63, KL108 and KL109 (Table 1), forms the α-L-FucpNAc-(1→3)-D-GlcpNAc linkage.22
Previous study has confirmed that the structure of the polysaccharide isolated from LAC-4 was built of trisaccharide repeating units α-L-FucNAc-3-α-D-GlcNAc-8-α-Leg5,7Ac,21 it is reasonable to speculate that gtr100 probably forms the α-D-GlcNAc-8-α-Leg5,7Ac linkage, and this linkage may be closely related to the virulence of KL49 because the virulence of LAC-4 decreased significantly when gtr100 was knocked out and gtr20 was retained. These inferences need to be verified by further experiments.
Although both previous studies and this study have demonstrated that KL49 A. baumannii is more virulent than nonKL49, the clinical characteristics of KL49 infection remain unclear. In general, A. baumannii is an opportunistic human pathogen that predominantly infects critically ill patients.1 However, due to the different virulence of different KL types, we have to regard them differently. Fatal outbreak of nosocomial infections caused by KL49 A. baumannii alerts the threat of this particular strain in clinic.7 Clinical characteristics analysis showed that CRAB infections with strains carrying gtr100 were the independent predictors for mortality, this is consistent with the conclusion that KL49 is a hypervirulent strain. Moreover, phylogenetic analysis suggests that KL49 A. baumannii has spread worldwide (Figure 2). Therefore, the development of a rapid identification assay without the need for WGS in this study should be very useful in advising the clinician to implement strict control measures and medical intervention earlier in order to avoid transmission and improve the prognosis of infection. Meanwhile, a rapid identification tool also makes the surveillance of these hypervirulent CRAB strains more convenient and faster.
The question raised by this study is how gtr100 affects the virulence of KL49. Correlation of the genes found in the KL gene cluster and the CPS structure from the same isolate has successfully confirmed the functions of several genes or gene clusters involved in the bonds formed by the encoded glycosyltransferases.23 In view of the important role of gtr100 gene in virulence, further studies on its function are necessary.
Data Sharing Statement
The genome sequences were deposited in GenBank under BioProject PRJNA533558.
Acknowledgments
We thank Yuting Huang for preparation of the figures.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the International Collaborative Research Fund (GJHZ20180413181716797) and Free Inquiry Fund (JCYJ20180305163929948) of Shenzhen Science and Technology Innovation Commission.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Dijkshoorn L, Nemec A, Seifert H. An increasing threat in hospitals: multidrug-resistant Acinetobacter baumannii. Nat Rev Microbiol. 2007;5(12):939–951. doi:10.1038/nrmicro1789
2. Evans BA, Hamouda A, Amyes SG. The rise of carbapenem-resistant Acinetobacter baumannii. Curr Pharm Des. 2013;19(2):223–238. doi:10.2174/138161213804070285
3. van Faassen H, KuoLee R, Harris G, Zhao X, Conlan JW, Chen W. Neutrophils play an important role in host resistance to respiratory infection with Acinetobacter baumannii in mice. Infect Immun. 2007;75(12):5597–5608. doi:10.1128/IAI.00762-07
4. Knapp S, Wieland CW, Florquin S, et al. Differential roles of CD14 and toll-like receptors 4 and 2 in murine Acinetobacter pneumonia. Am J Respir Crit Care Med. 2006;173(1):122–129. doi:10.1164/rccm.200505-730OC
5. Zeng X, Gu H, Cheng Y, et al. A lethal pneumonia model of Acinetobacter baumannii: an investigation in immunocompetent mice. Clin Microbiol Infect. 2019;25(4):516 e511–516 e514. doi:10.1016/j.cmi.2018.12.020
6. Harris G, Kuo Lee R, Lam CK, et al. A mouse model of Acinetobacter baumannii-associated pneumonia using a clinically isolated hypervirulent strain. Antimicrob Agents Chemother. 2013;57(8):3601–3613. doi:10.1128/AAC.00944-13
7. Jones CL, Clancy M, Honnold C, et al. Fatal outbreak of an emerging clone of extensively drug-resistant Acinetobacter baumannii with enhanced virulence. Clin Infect Dis. 2015;61(2):145–154. doi:10.1093/cid/civ225
8. Zhou K, Tang X, Wang L, et al. An emerging clone (ST457) of Acinetobacter baumannii clonal complex 92 with enhanced virulence and increasing endemicity in South China. Clin Infect Dis. 2018;67(suppl_2):S179–S188. doi:10.1093/cid/ciy691
9. Ou HY, Kuang SN, He X, et al. Complete genome sequence of hypervirulent and outbreak-associated Acinetobacter baumannii strain LAC-4: epidemiology, resistance genetic determinants and potential virulence factors. Sci Rep. 2015;5:8643.
10. Wyres KL, Cahill SM, Holt KE, Hall RM, Kenyon JJ. Identification of Acinetobacter baumannii loci for capsular polysaccharide (KL) and lipooligosaccharide outer core (OCL) synthesis in genome assemblies using curated reference databases compatible with Kaptive. Microb Genom. 2020;6(3).
11. Clinical, Institute LS. Performance Standards for Antimicrobial Susceptibility Testing. Wayne, PA: Clinical and Laboratory Standards Institute; 2017.
12. Chen WP, Kuo TT. A simple and rapid method for the preparation of gram-negative bacterial genomic DNA. Nucleic Acids Res. 1993;21(9):2260. doi:10.1093/nar/21.9.2260
13. Chen S, Zhou Y, Chen Y, Gu J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 2018;34(17):i884–i890. doi:10.1093/bioinformatics/bty560
14. Antipov D, Korobeynikov A, McLean JS, Pevzner PA. hybridSPAdes: an algorithm for hybrid assembly of short and long reads. Bioinformatics. 2016;32(7):1009–1015. doi:10.1093/bioinformatics/btv688
15. Treangen TJ, Ondov BD, Koren S, Phillippy AM. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014;15(11):524. doi:10.1186/s13059-014-0524-x
16. Stamatakis A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006;22(21):2688–2690. doi:10.1093/bioinformatics/btl446
17. Letunic I, Bork P. Interactive Tree Of Life (iTOL) v4: recent updates and new developments. Nucleic Acids Res. 2019;47(W1):W256–W259. doi:10.1093/nar/gkz239
18. Russo TA, Luke NR, Beanan JM, et al. The K1 capsular polysaccharide of Acinetobacter baumannii strain 307-0294 is a major virulence factor. Infect Immun. 2010;78(9):3993–4000. doi:10.1128/IAI.00366-10
19. Yu W-L, Ko W-C, Cheng K-C, Lee -C-C, Lai -C-C, Chuang Y-C. Comparison of prevalence of virulence factors for Klebsiella pneumoniae liver abscesses between isolates with capsular K1/K2 and non-K1/K2 serotypes. Diagn Microbiol Infect Dis. 2008;62(1):1–6. doi:10.1016/j.diagmicrobio.2008.04.007
20. Fang C-T, Lai S-Y, Yi W-C, Hsueh P-R, Liu K-L, Chang S-C. Klebsiella pneumoniae genotype K1: an emerging pathogen that causes septic ocular or central nervous system complications from pyogenic liver abscess. Clin Infect Dis. 2007;45(3):284–293. doi:10.1086/519262
21. Vinogradov E, Maclean L, Xu HH, Chen W. The structure of the polysaccharide isolated from Acinetobacter baumannii strain LAC-4. Carbohydr Res. 2014;390:42–45. doi:10.1016/j.carres.2014.03.001
22. Arbatsky NP, Kenyon JJ, Kasimova AA, et al. K units of the K8 and K54 capsular polysaccharides produced by Acinetobacter baumannii BAL 097 and RCH52 have the same structure but contain different di-N-acyl derivatives of legionaminic acid and are linked differently. Carbohydr Res. 2019;483:107745. doi:10.1016/j.carres.2019.107745
23. Kasimova AA, Kenyon JJ, Arbatsky NP, et al. Acinetobacter baumannii K20 and K21 capsular polysaccharide structures establish roles for UDP-glucose dehydrogenase Ugd2, pyruvyl transferase Ptr2 and two glycosyltransferases. Glycobiology. 2018;28(11):876–884.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.