Back to Journals » Infection and Drug Resistance » Volume 13
Protective Efficacy of the OprF/OprI/PcrV Recombinant Chimeric Protein Against Pseudomonas aeruginosa in the Burned BALB/c Mouse Model
Authors Fakoor MH, Mousavi Gargari SL, Owlia P, Sabokbar A
Received 29 December 2019
Accepted for publication 11 May 2020
Published 9 June 2020 Volume 2020:13 Pages 1651—1661
DOI https://doi.org/10.2147/IDR.S244081
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Joachim Wink
Mohammad Hadi Fakoor,1 Seyed Latif Mousavi Gargari,2 Parviz Owlia,3 Azar Sabokbar1
1Department of Microbiology, Karaj Branch, Islamic Azad University, Karaj, Iran; 2Department of Biology, Faculty of Basic Sciences, Shahed University, Tehran, Iran; 3Molecular Microbiology Research Center, Shahed University, Tehran, Iran
Correspondence: Seyed Latif Mousavi Gargari Tel +989123118510
Email [email protected]
Background: Pseudomonas aeruginosa infection is the major cause of death in burn patients. Thus, in this study, a chimeric vaccine harboring the OprF185– 350–OprI22– 83–PcrV was designed and expressed in Escherichia coli. The immunogenicity of the recombinant chimer, OprI, OprF, and PcrV was studied in a burned mouse model.
Methodology: Recombinant proteins including the proposed chimer, OprF, OprI, and PcrV were expressed in the E.coli. Mice were immunized with the purified recombinant proteins, and the antibody titre was estimated in the sera obtained from immunized mice. Immunized and control mice were challenged with 2, 5, and 10xLD50 of the P. aeruginosa strains (PAO1, PAK, and R5), and microbial counts were measured in the skin, liver, spleen, and kidney of the studied mice.
Results: Results showed that the antibody titre (total IgG) was significantly increased by injection of 10 μg of chimeric protein in the experimental groups compared to the control groups. The antibody survival titre was high until 235 days after administration of the second booster. The survival rate of the mice infected with 10xLD50 was significantly increased and the number of bacteria was reduced, especially in the internal organs (kidney, spleen, and liver) compared to the mice immunized with any of the OprF, OprI, and PcrV proteins alone.
Conclusion: The findings of our study revealed that the chimeric protein is a promising vaccine candidate for control of the P. aeruginosa infection.
Keywords: burned, chimeric protein, Pseudomonas aeruginosa, vaccine
Introduction
Pseudomonas aeruginosa (PA) is the most common germ-negative pathogen causing opportunistic infection in the human, particularly in the immunosuppressed patients.1 The Intensive Care Units (ICUs), the respiratory tract infection and burn wards in the hospital are the main places where PA infection can spread.2 Burns and respiratory infections can quickly lead to systemic infection with a mortality rate between 38% and 70%.3 However, the treatment of PA infection is difficult due to high prevalence of drug resistance and limited therapeutic options.4 Therefore, rapid and timely control of the infection is the most effective approach to prevent the PA infection in burn wounds.5 Although, treatment with the antibiotics is an important strategy, medication resistance by Pseudomonas aeruginosa has made it difficalt. Carbapenem antibiotics are used to treat the Multidrug-Resistant Pseudomonas (MDRP). With the prevalence of the carbapenem-resistant pseudomonas (CRP), the scientists have attempted to produce an appropriate vaccine or develop other safety treatments.6,7 So far, several vaccine candidates have been introduced against the PA.8–11 However, no license has been issued yet.12 The presence of several pathways used by the bacteria to cause infection is a major challenge for the development of the vaccine against the PA. Thus, the vaccines that block one or two routes are not able to have the appropriate protective effect. PA has the Type III Secretion System (T3SS) through which directly affects the host cells’ function.13 The PA V-antigen (PcrV) is an extracellular molecule of T3SS that kills the epithelial cells and immune cells by injecting a toxic protein. Promising results have been reported regarding the antibody (Fab) raised against the recombinant PcrV such that it decreased the inflammation and injury in the patients with cystic fibrosis and chronic infections caused by PA.14 Outer membrane protein I (OprI) is another important surface lipoprotein playing a vital role in the PA, particularly making it resistant to antimicrobial peptides.15,16 There is a highly promising vaccine for PA (NCT01563263), which is currently in the Phase III of clinical trial and consists of the OprI and OprF proteins. As an adjuvant, OprI launches the Toll-Like Receptor 2(TLR2)/Toll-Like Receptor 4(TLR4) pathway to improve the function of the immune system.17–19 According to the analysis of the immunogenicity, distribution, and critical role of the OprF, PcrV, and OprI in the pathogenesis of PA, and based on bioinformatics calculations, it was hypothesized that a combination of these proteins as a chimer may provide significant immunity and could provide the protection against the infection. Therefore, in this study, a chimeric vaccine harboring the OprF 185–350 - OprI 22–83 - PcrV was designed and expressed in the E.coli. The immunogenicity of the recombinant chimer was studied in the mouse model of burn.
Materials and Methods
Ethics Statement
All the animal studies were performed in accordance with the protocols provided by the Animal Care Committee of the Shahed University (Tehran, Iran). All the decisions of this committee were made in compliance with the UK Animals Scientific Procedures Act 1986 and related guidelines including the EU Directive 2010/63/EU and National Institutes of Health Guidelines for the Care and Use of Laboratory Animals (NIH Publication No. 8023, modified in 1978). We tried our best to minimize the animal suffering.
Animals, Microorganisms, and Cultures
Six to eight-weeks old male BALB/c mice, weighing 22–25 grams were purchased from the Razi Vaccine and Serum Research Institute (Alborz province, Karaj, Iran). There were eleven mice in each group in all the performed tests. The microorganisms, namely Pseudomonas aeruginosa strains of PAO1 (ATCC15692) and PAK (ATCC25102) used in this study were obtained from the microbial collection of the Faculty of Basic Sciences, Shahed University (Tehran, Iran). Pseudomonas aeruginosa R5, an aggressive and antibiotic-resistant hospital strain was collected from the Faculty of Pharmacy, Tehran University of Medical Sciences (Tehran, Iran). Luria-Bertani (LB) medium, Nutrient Agar (NA), and nutrient broth (all purchased from Merck, Germany) were used for routine culture of all the bacterial strains. All the chemical materials and solvents used in this study were purchased from the Merck Company (Germany).
Cloning, Expression, and Purification of the Recombinant OprF, OprI, PcrV, and OprF185–350–OprI22–83–PcrV Chimer
The reading framework of OprI and PcrV genes for protein production was isolated and amplified by the primers listed in Table 1 and was cloned on the pET28a expression vector, in the frame with a T7 promoter, kanamycin-resistant gene, and the N-terminal six-His-tagged sequence. HindIII and EcoRI (Fermentas, Lithonia) restriction sites were located at the 5′ end PcrV and OprI genes, respectively. The OprF clone was gifted by the research Laboratory, in the Department of Cell Biology, Faculty of Basic Sciences, Shahed University (Tehran, Iran). The OprF-OprI-PcrV chimeric gene was designed according to the genetic code of pseudomonas aeruginosa PAO1 ATCC15692 strain in the National Center for Biotechnology Information (NCBI). The gene was analyzed through the prospective bioinformatics analysis and was synthesized on the pET28a vector by the Biomatik Company (Canada). All the clones were cultured on the LB medium with kanamycin 70 ug/mL (Sigma-Aldrich, Germany) and were expressed in the E.coli BL-21 (DE3) strain (Novagen, Darmstadt, Germany) using 1 mM Isopropyl β-D-1-Thiogalactoside (IPTG) at OD600 of 0.6 as an inducer. After 6 h of incubation at 37°C and 200 rpm, centrifugation was done at 4000 g, 4°C for 15 min. The cell pellets were suspended in the Tris-EDTA buffer (Tris 1 M, EDTA 0.5 M) and were sonicated three times for 45 s at 200 W, cycle 0.5 s with 1 min resting on ice in each interval. The protein solutions were obtained at 17,500 g, 4°C for 20 min, and the supernatant containing recombinant proteins were purified with Ni-NTA agarose column (Qiagen, USA) in a natural condition using the imidazole gradient. Purified recombinant proteins were analyzed using the Sodium Dodecyl Sulfate Polyacrylamide Gel electrophoresis (SDS-PAGE) and were approved by the Western blotting. For Western blot analysis, the antigens were separated by the SDS-PAGE electrophoresis and were transferred onto the nitrocellulose membrane under semi-dry condition. Horseradish Peroxidase (HRP)-conjugated anti-His antibody (diluted 1:10,000) (Abcam, Cambridge, MA, USA) was used to detect the recombinant proteins.
 |
Table 1 Primers Used for Amplification of Open Reading Frame Sequences of the OprI and PcrV |
Immunization of the Mice and Determination of the Antibody Titre
Different mice groups were immunized with 10 μg of each of the recombinant proteins subcutaneously on days 0, 14, and 28. The first injection was performed with complete and two boosters with incomplete Freund’s adjuvant (Sigma-Aldrich, Germany). Burned mice were contaminated with the bacteria and burned mice without any contaminations were kept as two non-immune control groups. Blood samples were collected from the eyes of the mice before the second and third injections and the antibody titre of the sera was measured by the Enzyme-Linked Immunosorbent Assay (ELISA). For this purpose, 96-well flat-bottom plates were coated with 3 μg/well of each antigen in the carbonate–bicarbonate buffer (pH 9.6) at 4°C overnight. Then, the plates were washed three times with Phosphate-Buffered Saline (PBS) containing 0.05% of Tween 20 (PBST) and were blocked using the 5% skim milk (w/v) in PBST at 37°C for 45 min. After washing, serial dilutions of sera were added in duplicates and were subsequently incubated at 37°C for 1–2 h. The plates were washed and then, goat anti-mouse IgG-HRP (diluted 1:10,000) (DNAbiotech Co., Tehran, Iran) was added (100 uL/well). Following 1 h incubation of the plates at 37 °C and washing with the Phosphate-Buffered Saline with Tween® detergent (PBST), 3, 3′, 5, 5′- Tetramethylbenzidine (TMB) (Pasteur Institute, Tehran, Iran) solution was added and incubated in the dark for 20 min at room temperature. The 3M sulfuric acid (H2SO4) was used to stop the reaction followed by absorbance measurement at 450 nm on the ELISA plate reader.
Burn Infection
All the animals (weighing 20–25 g) were anesthetized with 200 μL of anesthetic drug (10 μL of xylazine, 20 μL of ketamine, and 170 μL of Double-Distilled Water (DDW)) (all purchased from Sigma-Aldrich Company, Germany) prior to burning.20 The right side of the waist and lower part of the body of all the mice were shaved using the shaving cream (Depi, Iran). The cylindrical probe made of iron alloy with a diameter of 22 mm, a length of 100 mm, and a weight of 165 g was heated to the temperature of 104°C and was applied on the shaved part of the animals for 8 s to create the 3rd-grade burns. Mice immediately received the peritoneal injection with 500 μL of 0.9% saline and 100 μL of acetaminophen (3.25 gr/mL) to prevent them from being shocked and feeling the pain.
Determination of LD50, 2xLD50, 5xLD50, and 10xLD50 Doses
Bacterial count was measured by estimating the OD600 using two-beam spectrophotometer (UV-Vis model T80Plus - PG, United Kingdom) and the graph was plotted using the Excel 2016 software. Different concentrations (10, 102, 5×102, 103, 2×103, and 104) of P. aeruginosa strains (PAO1, PAK, and R5) were injected subcutaneously at the burn center of all the mice to obtain the LD50 dose.
The results were analyzed using the SPSS software version 20, and LD50, 2xLD50, 5xLD50, and 10xLD50 doses were determined. The test was performed three times individually.21
Burned Mice Challenged with P. aeruginosa
Fourteen days after the third injection, burned immunized mice were challenged with the subcutaneous injection of bacteria at doses of 2xLD50, 5xLD50, and 10xLD50 at the burn center. They were kept for one month after burning until the wound was healed.20
Microbial Counting of the Mouse Tissues
Twelve hours after subcutaneous injection at a dose of 10xLD50 from each bacteria, three challenging mice from each group were sacrificed. Skin, liver, spleen, and kidney samples were homogenized in 4 mL of sterile PBS. Serial dilutions of homogeneous samples were prepared in sterile PBS and 100 μL of the diluted samples was placed on the Nutrient Agar (NA) and was incubated for 24 h at 37°C. The colonies were counted and the number of Colony Forming Units (CFU) from each sample in grams of tissue (CFU/g) was calculated.20,21
Bacterial Resistance
Bacterial resistance of P. aeruginosa strains including PAO1, PAK, and R5 was measured according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2016).22
Statistical Analysis
The Graph Pad Prism software (version 6) and Excel 2016 were used to draw the charts, graphs, and data concepts. Data were analyzed by the Analysis of Variance (ANOVA) and t-test using the SPSS software (version 20). Mean, standard deviation, and significance level were calculated and a p-value of <0.05 was considered as statistically significant.
Results
Characterization of the OprF, OprI, PcrV, and Chimeric Protein
Expression and purification of the OprF, OprI, PcrV, and the chimeric protein consisting of OprF 185–350, OprI 22–83, and PcrV were analyzed on the SDS-PAGE (Figure 1A, C, E, and G), and were verified by the Western blotting (Figure 1B, D, F, and H). The molecular weight of the recombinant proteins of OprF, OprI, PcrV, and the chimer was equal to 38, 8.4, 32, and 66 kDa, respectively.
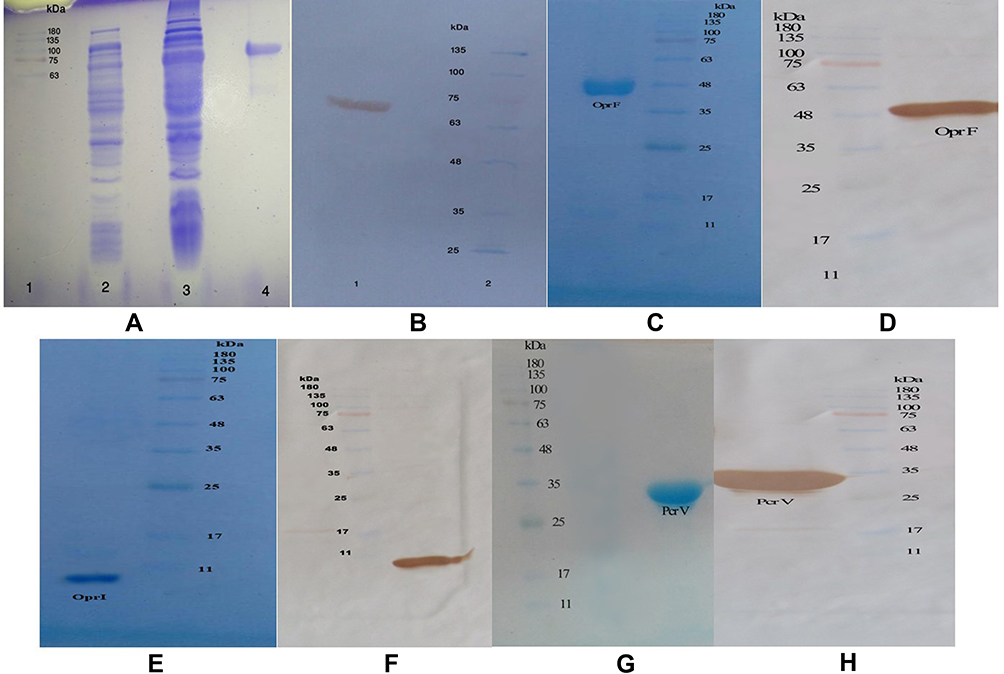 |
Figure 1 (A) Expressed and purified chimeric protein in the SDS-PAGE. The gel (12% w/v) was stained with the Coomassie blue G-250. Lane 2, pellet of the uninduced bacteria with IPTG; lane 1, standard protein size marker (kDa); lane 3, pellet of IPTG-induced bacteria; lane 4, Purification of the chimeric protein on the SDS-PAGE. (B) Confirmation of the chimeric protein through the Western blotting. (C) The OprF protein purified in the SDS-PAGE (12% w/v gel). (D) The OprF protein approved by the Western blotting. (E) The OprI protein purified in the SDS-PAGE by 9% w/v gel. (F) Confirmation of the OprI protein through the Western blotting. (G) The PcrV protein purified in the SDS-PAGE by 12% w/v gel. (H) The PcrV protein approved by the Western blotting. |
Antibody Titer
The ELISA was used to determine the antibody titres raised against the recombinant proteins. Figure 2B, C, and D show the related results. The durability of the antibody titres was also estimated (Figure 2A). The chimeric protein exhibited better immunogenicity and durability compared to other three recombinant proteins. No significant differences were observed in the results of the whole-cell ELISA for P. aeruginosa strains of PAO1 and PAK.
 |
Figure 2 ELISA assay. (A) Total IgG level in the mice immunized with the chimeric and recombinant proteins at different times. (B) The Total IgG bar chart after administration of the second booster in serial dilution antibody for chimeric and recombinant proteins. (C) The binding power of specific antibodies to the Pseudomonas aeruginosa PAO1 whole-cell in the chimeric and recombinant proteins. (D) Comparison of the whole-cell ELISA results between two reference strains of Pseudomonas aeruginosa. *p< 0.05, **p< 0.01 and ***p< 0.001. Abbreviation: Ns, not significant. |
LD50 and Animal Challenges
Table 2 shows the results regarding the animal challenges at a dose of LD50 for each bacteria. The immunized mice were challenged with 2xLD50, 5xLD50, and 10xLD50 doses of P. aeruginosa strains of PAO1, PAK, and R5 to determine the protective potency of our recombinant proteins. A significant (p< 0.05) improvement was observed in the survival rate of the mice vaccinated with the chimeric protein compared to those immunized with any of OprI, PcrV, and OprF proteins (Figure 3). The survival rate of the mice immunized with the OprI protein was much lower than that of other recombinant proteins at the doses of 5xLD50 or 10xLD50. Totally, 75% of the mice immunized with the chimeric protein successfully survived against the 10xLD50 of R5 resistant strain (Figure 3F). The mice immunized with the chimeric protein survived after 3 days of severe infection caused by the 5xLD50 of PAO1, PAK, or R5 resistant strains. However, the mice immunized with each of the proteins tolerated severe infection for 6 days after receiving the 5xLD50 of the bacteria.
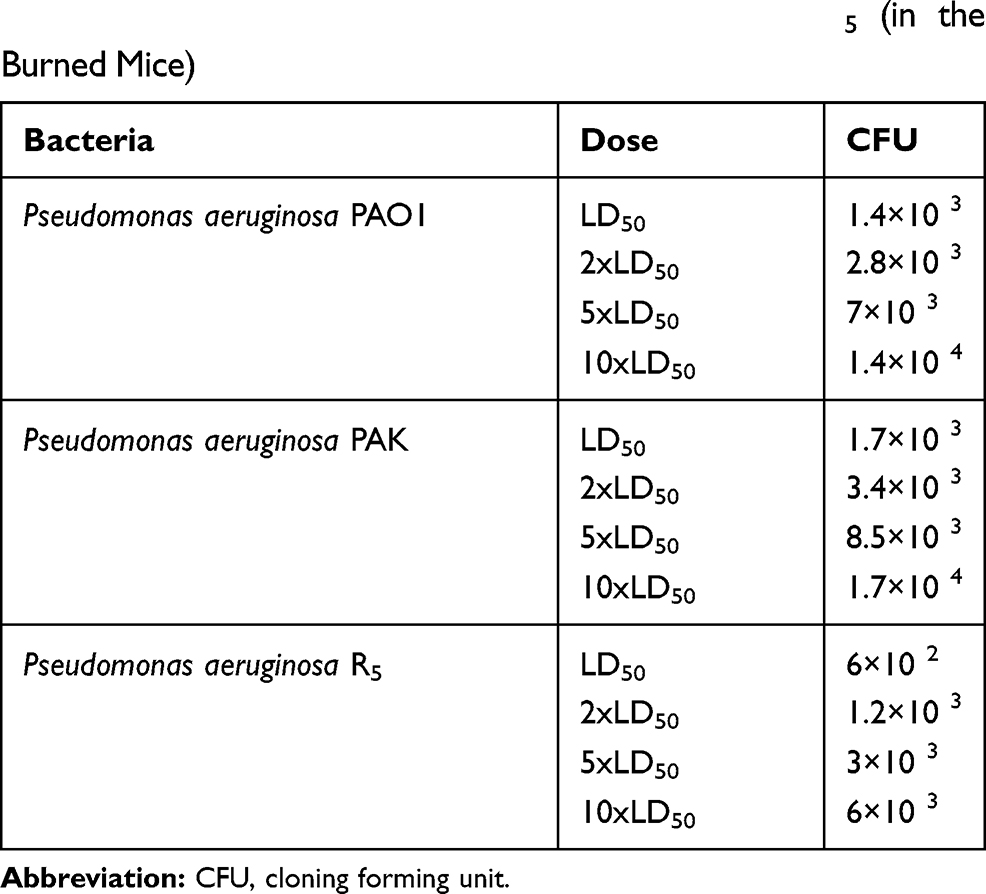 |
Table 2 The Amounts of Inoculation Doses of Death by the Pseudomonas aeruginosa Strains of PAO1, PAK, and R5 (in the Burned Mice) |
 |
Figure 3 Comparison of survival rate in the two control groups (including non-immune mice that were only burned and those received adjuvant with PBS) and the mice immunized with four vaccine candidates (chimeric protein, OprI, OprF, and PcrV) that were challenged with burn wound infections by the P. aeruginosa strains of PAO1, PAK, and R5. *p< 0.05, **p< 0.01 and ***p< 0.001. Abbreviation: ns, not significant. |
Bacterial Burden
As shown in Figure 4A and B, the mice immunized with the recombinant proteins showed significantly lower bacterial loads in the skin, liver, spleen, and kidney compared to the control groups 12 h after infection. However, the bacterial count in the mice immunized with the chimeric protein was much lower in different organs than that of the three individual proteins. Our results exhibited that immunization with the chimeric protein could reduce the bacterial local colonization and systematic spreading simultaneously.
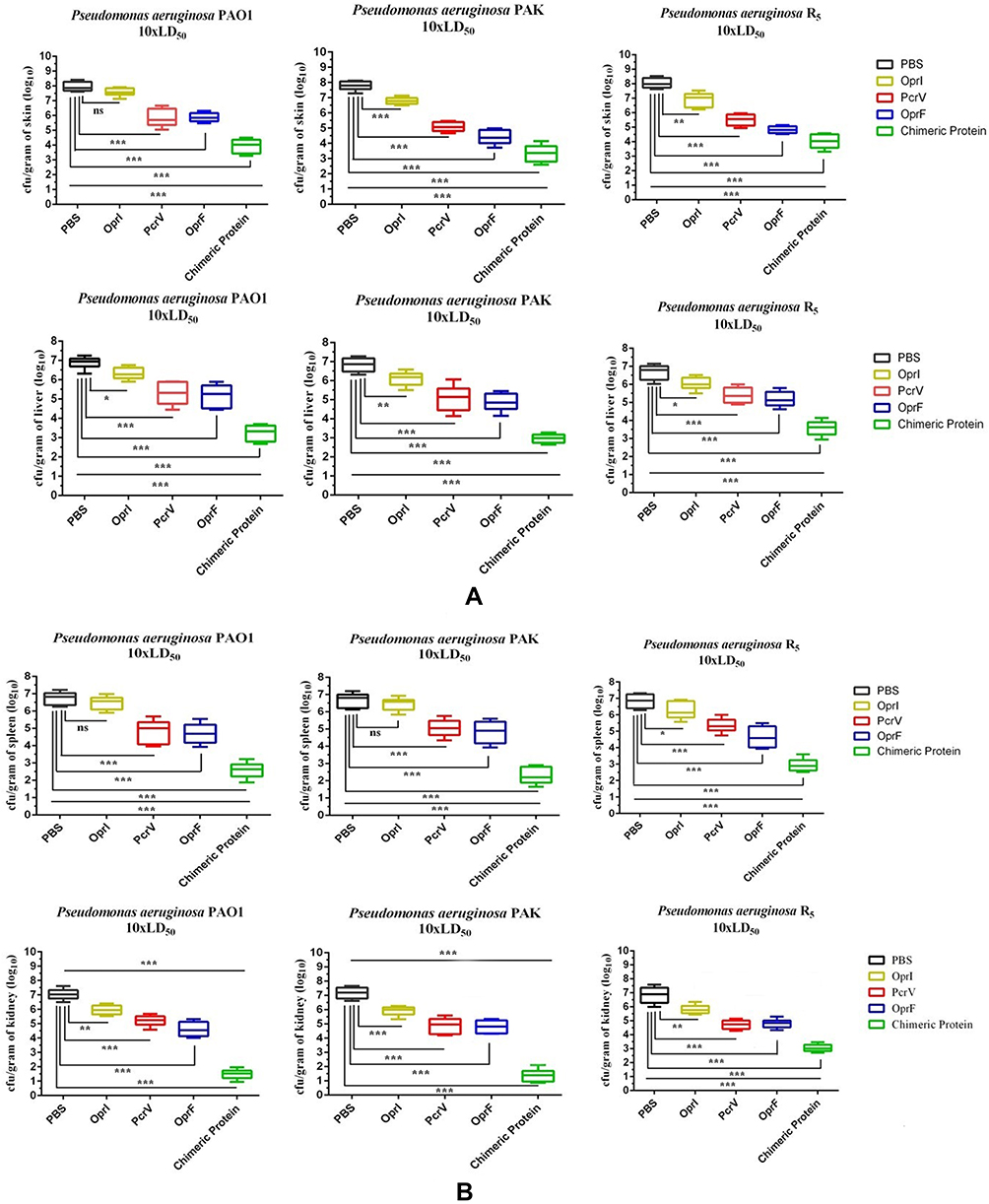 |
Figure 4 Bacterial loads in the skin and liver (A) spleen and kidney (B) of the mice immunized 12 h after infection with the 10xLD50 CFU of PAO1, PAK, and R5. *p< 0.05, **p< 0.01 and ***p< 0.001. Abbreviation: ns, not significant. |
Bacterial Resistance
Disc diffusion method (CLSI-2016) was used to evaluate the resistance to the P. aeruginosa.22 The R5 hospital strain showed resistance to ciprofloxacin 5 μg and imipenem 10 μg. However, the growth inhibition zone was equal to 28 mm in the imipenem disk for the P. aeruginosa PAO1 and it was equal to 30 mm for the P. aeruginosa PAK. For the ciprofloxacin (5 μg), the growth inhibition zone was equal to 34 and 37 mm for the PAO1 and PAK strains, respectively (Figure 5).
 |
Figure 5 Bacterial resistance tested by the disc diffusion method on the Pseudomonas aeruginosa strains of PAO1, PAK, and R5. R5 hospital strain was resistant to ciprofloxacin 5 μg and imipenem 10 μg. |
Discussion
Currently, there are several proteins available as a vaccine candidate for protection against the Pseudomonas bacteria. Some of these proteins have undergone research tests and some are under clinical trials.23,24 Among these, OprI, OprF, and PcrV are of considerable importance, since they are expressed in most of the pathogenic strains of PA and play an essential role in the pathogenesis of PA.24–26 In the present study, these proteins were selected and produced to investigate their potency as new vaccine candidates against PA infection. Several reports have indicated that the antibodies raised against the OprF, OprI, and PcrV could provide protection against the PA infection.14,27–29 Active immunization with the OprI and OprF,30–32 and passive immunization with the PcrV14,27–29 have been shown to protect the animals challenged with the PA.10,32 The three antigens selected in this study are conserved among different PA isolates, and as a result, one can expect a cross-protective effect when challenged with different clinical isolates. Previous studies have revealed that single subunit vaccines are mostly inadequate in protecting against the infections. Therefore, in this study, the immunogenic regions of the three proteins were selected and designed to develop a chimeric protein using the bioinformatics tools. The antigenic and immunogenic MHC class I, MHC class II, and B cell epitopes present in the OprI and OprF proteins were identified by the reverse vaccinology approach. The selected epitopes were listed based on their antigenicity and toxicity potentials in order to design our chimeric vaccine. The immunogenicity of the chimeric protein was compared with each of the OprI, OprF, and PcrV proteins alone (Figure 1C–H). The anti-chimer antibodies could protect against the potential infections by the 10xLD50 of P. aeruginosa strains (PAO1, PAK, and R5) in the mouse model of burn wound. The survival rate of the mice immunized with the chimeric protein was increased compared to other recombinant proteins applied alone against infections by all the strains (Figure 3). Hydrophobicity is considered as an important virulence factor of various pathogens. Factors increasing the hydrophobicity can enhance the adhesion of bacteria to intestinal epithelial cells and thereby facilitate the invasion. Adhesion to the epithelial cells has been found to be associated with the hydrophobicity of the Streptococcus pyogenes strain, and sub-inhibitory concentrations of rifampicin have been shown to lower the hydrophobicity and reduce the cell adhesion.33 The hydrophobicity of the bacterial surface acts as opsonin, creates the phagocytic capacity, and leads to the development of the complement system and IgG opsonization of Salmonella enterica serovar Typhimurium.34 Calvinho et al have reported that fast-growing strains of Staphylococcus aureus exhibited high surface hydrophobicity and consequently high pathogenicity compared to slow-growing hydrophilic strains.35 The engulfment of the bacteria by the phagocytic cells as a result of treatment with the antibiotics occurs due to increased bacterial surface hydrophobicity. In a study, hydrophobicity and phagocytic killing of the PA were increased when treated with sub-inhibitory concentration of aztreonam, whereas low exposures to inhibitory concentrations caused similar effects on other Gram-negative bacteria such as E. coli, Serratia marcescens, Klebsiella pneumoniae, and Salmonella typhimurium.36 On the other hand, the increase in the bacterial surface hydrophobicity seems to fulfill the lack of opsonization and leads to competent phagocytosis. In this regard, non-opsonized hydrophobic strains of Bacteroides buccae, Porphyromonas gingivalis, Staphylococcus saprophyticus, and Fusobacterium nucleatum have been shown to be readily phagocytosed by the Polymorphonuclear leukocytes (PMNs), whereas the phagocytic killing of hydrophilic strains required human serum opsonization.37,38 Thus, it is assumed that the hydrophobicity confirmed by the bioinformatics analysis of the selected recombinant proteins consisting of the OprF 185–350, OprI 22–83, and PcrV increases the phagocytic ability of the PMNs by increasing the bacterial aggregation and may facilitate a rapid and competent host response. It has been reported that the antibodies act against the PcrV, OprI, and OprF virulence factors of PA speculate and interfere with the early stages of its pathogenesis, as well as later stages such as systemic dissemination to the organs.39 Our results showed that resistance against pathogenic bacteria was started from the skin and more bacteria were killed in this area. None of PA strains (PAO1, PAK, and R5) were found in the internal organs at the dose of 2xLD50. Bacteria were able to cross the damaged skin to a low extent at the doses of 5xLD50 and 10xLD50 (Figure 4A and B). The OprI, OprF, and PcrV proteins cause humoral and cellular immunity.23 OprI increases the production of Interleukin 4 (IL-4), Interleukin 6 (IL-6), Interleukin 10 (IL-10), Interferon gamma (IFN-Ƴ), Tumor Necrosis Factor-alpha (TNF-α), and Immunoglobulin G (IgG)3,18 involved in the humoral and cellular immunity function. Rau et al have shown that the OprI can act as an adjuvant.18 In our study, when the OprI was used along with the PcrV and OprF proteins, the antibody rise was significantly increased compared to any of the antigens used alone (Figure 2A–C). The antibody titre (IgG total), the concentration, and shelf-life of the antibody produced against the PA were much higher in the chimeric protein-immunized mice than the single proteins (Figure 1A and B). Weimer et al40 have shown the synergistic effect of the OprI and OprF proteins in the rise of IgG titre, which seems to be a property of the OprI adjuvant.18 The neutralization power of the antibody produced by the chimeric protein was much higher than any of the single proteins (Figure 2C). This means that the chimeric protein could attach to the whole-cell with high affinity. This was approximately the same in the whole-cell ELISA results for both strains P. aeruginosa of PAO1 and PAK (Figure 2D) attributing to the presence of the conserved domains of the selected proteins in all the three strains studied in our research. Zhang et al have reported that the immune induction occurred due to the presence of OprF in their proposed vaccine.21 Besides increasing the IgG antibody titre, OprF also increases the production of the Interleukin 1 beta (IL-1β), Interleukin 2 (IL-2), IL-4, IL-6, Interleukin 12 (IL-12), Interleukin 17 (IL-17), INF-Ƴ, and TNF-α.23,41 Chuang Wan et al have shown that the PcrVNH protein not only increases the levels of IgG1, IgG2a, and IgG2b but also increases the titre of cytokines including IL-4, IL-17, and IL-1β, suggesting the humoral immunity. The PcrVNH protein is composed of the N-terminal (1–127 amino acid) and the helix 12 in C-terminal (251–294 aa.) of PcrV.42 PcrV protein as a vaccine could increase the titres of the TNF-α, INF-Ƴ, Cluster of Differentiation 3 (CD3), Cluster of Differentiation 28 (CD28), and Cluster of Differentiation 4 (CD4).3 Therefore, all the three proteins selected to design the chimeric antigen have both the ability to activate cellular and humoral immunity. In general, outer membrane proteins can increase the humoral immunity by increasing the (Th2) CD4 T Cells (CD44 + IL-4) and can cause cellular immunity by increasing the (Th1) CD4 T Cells (CD44 +IFN-Ƴ).21,43 Many attempts have also been made to develop multiple vaccines using some outer membrane proteins and some success has been achieved.3,23,44 Other researchers have turned to development of a multi-antigen vaccine since PA has many factors involved in its pathogenesis. Yang et al have designed a trivalent vaccine (PcrV28–294-OprI25–83-Hcp11–162) with Al(OH)3 adjuvant and evaluated its protective efficacy in mouse models of pneumonia and burn. Immunization with this trivalent vaccine induced strong immune responses and reduced the bacterial loads, decreased the pathology, inflammatory cytokine expression, and inflammatory cell infiltration.3 Weimer et al have indicated that intramuscular immunization with a combination of OprF epitope 8 (OprF311–341), OprI, and flagellins (types A or B) produced high-affinity IgG antibodies specific to the flagellins, OprI, and OprF that individually promoted extensive deposition of the Complement Component 3 (C3) on PA.30 Hassan et al have conducted a research on the antigens of trivalent vaccine (OprF, OprI, and flagellin B) and confirmed many results reported in the study by Weimer et al. Both studies were performed to select the same antigens and evaluate their efficacy on non-mucoid pseudomonas infection.44 Despite significant efforts in raising awareness in producing a vaccine against the PA, to the best of our knowledge, no effective vaccine has been developed yet, suggesting the potential for developing a vaccine to prevent infection.
Conclusion
In summary, our results demonstrated that the chimeric protein could be an effective vaccine candidate for protection against the P. aeruginosa infection. Combination of these proteins in the chimer increased the survival rate of the mice compared to any of the antigens applied alone.
Ethical Approval
All animal studies were performed based on the protocols reviewed and approved by the Animal Care Committee of the Shahed University in Iran. The Ethics Committee endorsement number is the IR.SHAHED.REC.1398.057 of Shahed University.
Acknowledgment
This article is a part of Mohammad Hadi Fakoor’s PhD thesis in Azad Islamic University (Karaj Branch). The authors extend their gratitude to Shahed University for providing facilities to conduct this study.
Author Contributions
Prof. Owlia and Mousavi are the supervisor of the PhD thesis of Mohammad Hadi Fakoor. Mohammad Hadi Fakoor as a PhD student performed the experiments and wrote the draft of the article. Ms. Sabokbar participated as a consultant in this thesis project. All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Moradali MF, Ghods S, Rehm BH. Pseudomonas aeruginosa lifestyle: a paradigm for adaptation, survival, and persistence. Front Cell Infect Microbiol. 2017;7:39. doi:10.3389/fcimb.2017.00039
2. Nathwani D, Raman G, Sulham K, et al. Clinical and economic consequences of hospital-acquired resistant and multidrug-resistant Pseudomonas aeruginosa infections: a systematic review and meta-analysis. Antimicrob Resist Infect Control. 2014;3(1):32. doi:10.1186/2047-2994-3-32
3. Yang F, Gu J, Yang L, et al. Protective efficacy of the trivalent Pseudomonas aeruginosa vaccine candidate PcrV-OprI-Hcp1 in murine pneumonia and burn models. Sci Rep. 2017;7:3957. doi:10.1038/s41598-017-04029-5
4. Ding C, Yang Z, Wang J, et al. Prevalence of Pseudomonas aeruginosa and antimicrobial-resistant Pseudomonas aeruginosa in patients with pneumonia in mainland China: a systematic review and meta-analysis. Int J Infect Dis. 2016;49:119–128. doi:10.1016/j.ijid.2016.06.014
5. Potron A, Poirel L, Nordmann P. Emerging broad-spectrum resistance in Pseudomonas aeruginosa and Acinetobacter baumannii: mechanisms and epidemiology. Int J Antimicrob Agents. 2015;45:568–585. doi:10.1016/j.ijantimicag.2015.03.001
6. Mikucionyte G, Zamorano L, Vitkauskiene A, et al. Nosocomial dissemination of VIM-2-producing ST235 Pseudomonas aeruginosa in Lithuania. Eur J Clin Microbiol Infect Dis. 2016;35:195–200. doi:10.1007/s10096-015-2529-0
7. Leylabadlo HE, Asgharzadeh M, Aghazadeh M. Dissemination of carbapenemases producing gram negative bacteria in the Middle East. Iran J Microbiol. 2015;7:226.
8. Döring G, Meisner C, Stern M, et al. A double-blind randomized placebo-controlled phase III study of a Pseudomonas aeruginosa flagella vaccine in cystic fibrosis patients. Proc Natl Acad Sci. 2007;104:11020–11025. doi:10.1073/pnas.0702403104
9. Westritschnig K, Hochreiter R, Wallner G, et al. A randomized, placebo-controlled Phase I study assessing the safety and immunogenicity of a Pseudomonas aeruginosa hybrid outer membrane protein OprF/I vaccine (IC43) in healthy volunteers. Hum Vaccines Immunother. 2014;10:170–183. doi:10.4161/hv.26565
10. François B, Luyt C-E, Dugard A, et al. Safety and pharmacokinetics of an anti-PcrV PEGylated monoclonal antibody fragment in mechanically ventilated patients colonized with Pseudomonas aeruginosa: a randomized, double-blind, placebo-controlled trial. Crit Care Med. 2012;40:2320–2326. doi:10.1097/CCM.0b013e31825334f6
11. Lu Q, Rouby -J-J, Laterre P-F, et al. Pharmacokinetics and safety of panobacumab: specific adjunctive immunotherapy in critical patients with nosocomial Pseudomonas aeruginosa O11 pneumonia. J Antimicrob Chemother. 2011;66:1110–1116. doi:10.1093/jac/dkr046
12. Vincent J-L. Vaccine development and passive immunization for Pseudomonas aeruginosa in critically ill patients: a clinical update. Future Microbiol. 2014;9:457–463. doi:10.2217/fmb.14.10
13. Gellatly SL, Hancock RE. Pseudomonas aeruginosa: new insights into pathogenesis and host defenses. Pathog Dis. 2013;67:159–173. doi:10.1111/2049-632X.12033
14. Milla CE, Chmiel JF, Accurso FJ, et al. Anti‐PcrV antibody in cystic fibrosis: a novel approach targeting Pseudomonas aeruginosa airway infection. Pediatr Pulmonol. 2014;49:650–658. doi:10.1002/ppul.22890
15. Tseng T-S, Wang S-H, Chang T-W, et al. Sarkosyl-induced helical structure of an antimicrobial peptide gw-q6 plays an essential role in the binding of surface receptor OprI in Pseudomonas aeruginosa. PLoS One. 2016;11:e0164597. doi:10.1371/journal.pone.0164597
16. Chang T-W, Wang C-F, Huang H-J, et al. Key residues of outer membrane protein OprI involved in hexamer formation and bacterial susceptibility to cationic antimicrobial peptides. Antimicrob. Agents Chemother. 2015;59:6210–6222. doi:10.1128/AAC.01406-15
17. Gartner T, Baeten M, Otieno S, et al. Mucosal prime-boost vaccination for tuberculosis based on TLR triggering OprI lipoprotein from Pseudomonas aeruginosa fused to mycolyl-transferase Ag85A. Immunol Lett. 2007;111:26–35. doi:10.1016/j.imlet.2007.04.010
18. Rau H, Revets H, Cornelis P, et al. Efficacy and functionality of lipoprotein OprI from Pseudomonas aeruginosa as adjuvant for a subunit vaccine against classical swine fever. Vaccine. 2006;24(22):4757–4768. doi:10.1016/j.vaccine.2006.03.028
19. Revets H, Pynaert G, Grooten J, et al. Lipoprotein I, a TLR2/4 ligand modulates Th2-driven allergic immune responses. J. Immunol. 2005;174:1097–1103. doi:10.4049/jimmunol.174.2.1097
20. Behrouz B, Mahdavi M, Amirmozafari N, et al. Immunogenicity of Pseudomonas aeruginosa recombinant b-type fagellin as a vaccine candidate: protective efficacy in a murine burn wound sepsis model. Burns. 2016. doi:10.1016/j.burns.2016.03.015
21. Zhang X, Yang F, Zou J, et al. Immunization with Pseudomonas aeruginosa outer membrane vesicles stimulates protective immunity in mice. Vaccine. 2018;36(8):1047–1054. doi:10.1016/j.vaccine.2018.01.034
22. CLSI C. Performance Standards for Antimicrobial Susceptibility Testing. Clinical Lab Standards Institute; 2016.
23. Aguilera-Herce J, García-Quintanilla M, Romero-Flores R, et al. A live salmonella vaccine delivering PcrV through the type III secretion system protects against Pseudomonas aeruginosa. mSphere. 2019;4(2):e00116–e00119. doi:10.1128/mSphere.00116-19
24. Hoggarth A, Weaver A, Pu Q, et al. Mechanistic research holds promise for bacterial vaccines and phage therapies for Pseudomonas aeruginosa. Drug Des Dev Ther. 2019;13:909. doi:10.2147/DDDT.S189847
25. Nilsson E, Larsson A, Olesen HV, et al. Good effect of IgY against Pseudomonas aeruginosa infections in cystic fibrosis patients. Pediatr Pulmonol. 2008;43:892–899. doi:10.1002/ppul.20875
26. Sato H, Frank D. Multi-functional characteristics of the Pseudomonas aeruginosa type III needle-tip protein, PcrV; comparison to orthologs in other gram-negative bacteria. Front Microbiol. 2011;2:142. doi:10.3389/fmicb.2011.00142
27. Warrener P, Varkey R, Bonnell JC, et al. A novel anti-PcrV antibody providing enhanced protection against Pseudomonas aeruginosa in multiple animal infection models. Antimicrob Agents Chemother. 2014;58:4384–4391. doi:10.1128/AAC.02643-14
28. Wang Q, Li H, Zhou J, et al. PcrV antibody protects multi-drug resistant Pseudomonas aeruginosa induced acute lung injury. Respir Physiol Neurobiol. 2014;193:21–28. doi:10.1016/j.resp.2014.01.001
29. Song Y, Baer M, Srinivasan R, et al. PcrV antibody–antibiotic combination improves survival in Pseudomonas aeruginosa-infected mice. Eur J Clin Microbiol Infect Dis. 2012;31:1837–1845. doi:10.1007/s10096-011-1509-2
30. Weimer ET, Lu H, Kock ND, et al. A fusion protein vaccine containing OprF epitope 8, OprI, and type A and B flagellins promotes enhanced clearance of nonmucoid Pseudomonas aeruginosa. Infect Immun. 2009;77:2356–2366. doi:10.1128/IAI.00054-09
31. Haberstroh J, Gilleland H
32. Baumann U, Mansouri E, Von Specht B-U. Recombinant OprF–OprI as a vaccine against Pseudomonas aeruginosa infections. Vaccine. 2004;22:840–847. doi:10.1016/j.vaccine.2003.11.029
33. Tylewska S, Hjertén S, Wadström T. Effect of subinhibitory concentrations of antibiotics on the adhesion of Streptococcus pyogenes to pharyngeal epithelial cells. Antimicrob Agents Chemother. 1981;20:563–566. doi:10.1128/AAC.20.5.563
34. Stendahl O, Tagesson C, Magnusson K, et al. Physiochemical consequences of opsonization of Salmonella typhimurium with hyperimmune IgG and complement. Immunology. 1977;32:11.
35. Calvinho L, Almeida R, Oliver S. Influence of Streptococcus dysgalactiae surface hydrophobicity on adherence to mammary epithelial cells and phagocytosis by mammary macrophages. J Vet Med Ser B. 1996;43:257–266. doi:10.1111/j.1439-0450.1996.tb00313.x
36. Pruul H, Lewis G, McDonald PJ. Enhanced susceptibility of gram-negative bacteria to phagocytic killing by human polymorphonuclear leucocytes after brief exposure to aztreonam. J Antimicrob Chemother. 1988;22:675–686. doi:10.1093/jac/22.5.675
37. Kerosuo E, Haapasalo M, Alli K, et al. Ingestion of Bacteroides buccae, Bacteroides oris, Porphyromonas gingivalis, and Fusobacterium nucleatum by human polymorphonuclear leukocytes in vitro. Oral Microbiol Immunol. 1990;5:202–207. doi:10.1111/j.1399-302X.1990.tb00646.x
38. Maródi L, BURJÁN P, Rozgonyi F. Opsonic requirements and surface hydrophobicity of novobiocin-resistant, coagulase-negative staphylococci. J Med Microbiol. 1990;32:19–24. doi:10.1099/00222615-32-1-19
39. Dacheux D, Goure J, Chabert J, et al. Pore‐forming activity of type III system‐secreted proteins leads to oncosis of Pseudomonas aeruginosa‐infected macrophages. Mol Microbiol. 2001;40:76–85. doi:10.1046/j.1365-2958.2001.02368.x
40. Weimer ET, Ervin SE, Wozniak DJ, et al. Immunization of young African green monkeys with OprF epitope 8–OprI–type A-and B-flagellin fusion proteins promotes the production of protective antibodies against nonmucoid Pseudomonas aeruginosa. Vaccine. 2009;27:6762–6769. doi:10.1016/j.vaccine.2009.08.080
41. Gong Q, Ruan M, Niu M, et al. Immune efficacy of DNA vaccines based on oprL and oprF genes of Pseudomonas aeruginosa in chickens. Poult Sci. 2018;97:4219–4227. doi:10.3382/ps/pey307
42. Chuang Wan JZ, Zhao L, Cheng X, et al. Rational design of a chimeric derivative of PcrV as a subunit vaccine against Pseudomonas aeruginosa. Front Immunol. 2019;10. doi:10.3389/fimmu.2019.00781
43. Baker SM, Pociask D, Clements JD, et al. Intradermal vaccination with a Pseudomonas aeruginosa vaccine adjuvanted with a mutant bacterial ADP-ribosylating enterotoxin protects against acute pneumonia. Vaccine. 2019;37:808–816. doi:10.1016/j.vaccine.2018.12.053
44. Hassan R, El-Naggar W, El-Aziz AMA, et al. Immunization with outer membrane proteins (OprF and OprI) and flagellin B protects mice from pulmonary infection with mucoid and nonmucoid Pseudomonas aeruginosa. J Microbiol Immunol Infect. 2018;51:312–320. doi:10.1016/j.jmii.2016.08.014
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.