")
Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 13
Progressive Loss of Corneal Nerve Fibers and Sensitivity in Rats Modeling Obesity and Type 2 Diabetes Is Reversible with Omega-3 Fatty Acid Intervention: Supporting Cornea Analyses as a Marker for Peripheral Neuropathy and Treatment
Authors Coppey L, Davidson E, Shevalye H , Obrosov A, Torres M, Yorek MA
Received 29 January 2020
Accepted for publication 31 March 2020
Published 24 April 2020 Volume 2020:13 Pages 1367—1384
DOI https://doi.org/10.2147/DMSO.S247571
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Professor Ming-Hui Zou
Lawrence Coppey,1 Eric Davidson,1 Hanna Shevalye,1 Alexander Obrosov,1 Michael Torres,1 Mark A Yorek1– 4
1Department of Internal Medicine, University of Iowa, Iowa City, IA, USA; 2Department of Veterans Affairs, Iowa City Health Care System, Iowa City, IA, USA; 3Veterans Affairs Center for the Prevention and Treatment of Visual Loss, Iowa City, IA, USA; 4Fraternal Order of Eagles Diabetes Research Center, University of Iowa, Iowa City, IA, USA
Correspondence: Mark A Yorek
Department of Veterans Affairs, Iowa City Health Care System, Iowa City, IA 52246 Tel +1-319-338-0581 ext. 7696
Fax +1-319-339-7162
Email [email protected]
Purpose: To determine whether cornea nerve fiber density and/or corneal function are valid markers for early detection and treatment of peripheral neuropathy in rats modeling prediabetes and type 2 diabetes.
Methods: High-fat feeding combined without or with low-dose streptozotocin was used to create rat models for prediabetes and type 2 diabetes that were longitudinally studied for loss of structure and function of sensory nerves in the cornea and skin as well as nerve conduction velocity and vascular reactivity of epineurial arterioles. There were three time points examined in each of the three conditions with 12 rats per group. The latest time point (24 weeks of high-fat diet with or without 16 weeks of hyperglycemia) was used to examine reversibility of neuro and vascular pathology following 16 weeks of treatment with menhaden oil, a natural source of long-chain omega-3 polyunsaturated fatty acids. The number of rats in the intervention study ranged from 6 to 17.
Results: Our longitudinal study demonstrated that vascular and neural dysfunction associated with obesity or type 2 diabetes occur early and are progressive. Decrease in cornea nerve fiber length and function were valid markers of disease in both the pre-diabetic and diabetic rat models and were more sensitive than decrease in intraepidermal nerve fiber density of the skin and thermal nociception of the hindpaw. Late intervention with menhaden oil significantly reversed both vascular and peripheral nerve damage induced by chronic obesity or type 2 diabetes.
Conclusion: These studies provide support for examination of corneal structure and function as an early marker of peripheral neuropathy in prediabetes and type 2 diabetes. Furthermore, we demonstrate that omega-3 polyunsaturated fatty acids derived from fish oil are an effective treatment for peripheral neuropathy that occurs with chronic obesity or type 2 diabetes.
Keywords: corneal nerves, peripheral neuropathy, omega-3 polyunsaturated fatty acids, diabetes, obesity
Introduction
Diagnosis of diabetic sensorimotor polyneuropathy is challenging and often made during later stages of disease progression in part due to the lack of methodology that can objectively detect its early stages.1 The inability to accurately detect and stage the loss of peripheral nerves and function early has likely contributed to the failure of many clinical trials seeking a treatment for diabetic peripheral neuropathy. Recently, the quantification of intraepidermal nerve fiber density in skin biopsies or using corneal confocal microscopy to non-invasively examine the corneal sub-epithelial nerves have emerged as promising techniques for the early detection of nerve fiber loss.2 It has been proposed that analysis of the function and structure of small nerve fibers in the cornea or skin provide an approach for early detection of diabetic peripheral neuropathy and a means to assess success or failure of treatments.
The cornea derives its innervation from the ophthalmic division of the trigeminal nerve and is the most densely innervated part of the human body containing Aδ and unmyelinated C fibers.3 When stimulated these nerves induce tear production and stimulate the blinking reflex.4 The combination of these events helps to maintain a well-lubricated and smooth ocular surface and helps to minimize visual distortion and dry eye symptoms, while protecting the ocular surface.4 Many disease states are associated with loss of corneal nerves and decreased sensitivity, which leads to the question whether examination of corneal nerve morphometry or cornea sensitivity can be used as markers for early diagnosis, progression of disease or recovery following treatment. In vivo corneal confocal microscopy provides non-invasive imaging of small nerve fibers in the cornea while various adaptations of esthesiometry using Cochet–Bonnet (contact esthesiometry) or Belmonte (non-contact esthesiometry) have been used to measure corneal sensitivity.4–9 In preclinical studies, we have examined the response to a hyperosmotic solution applied to the cornea as another approach that may be adaptable to humans for determining cornea sensitivity.10,11
Corneal confocal microscopy has been used not only to detect changes in the corneal sub-epithelial nerve plexus in cornea-related dystrophies but also in a wide array of systemic and neurodegenerative diseases including diabetic peripheral neuropathy, rheumatoid arthritis, idiopathic small fiber neuropathy, Fabry disease, human immunodeficiency virus-associated peripheral neuropathy, and chemotherapy-induced peripheral neuropathy.5,6,12-17 Morphological changes of these nerves in both type 1 and type 2 diabetes has been shown to be a useful tool to diagnose and stratify the severity of peripheral neuropathy.18 A recent meta-analysis assessing the diagnostic performance of corneal confocal microscopy in patients with diabetic peripheral neuropathy determined that corneal confocal microscopy to be valuable for detecting and assessing early nerve damage.19 Studies have also demonstrated that corneal confocal microscopy can detect neuropathy in human subjects with impaired glucose tolerance.20–22 In rats fed a high-fat diet, a model of prediabetes, we have reported a decrease in sensory nerve conduction velocity as well as a decrease in corneal sub-epithelial nerves and cornea sensitivity in the absence of hyperglycemia, thereby supporting reports at the preclinical level of a decrease in corneal nerves in human subjects deemed to be pre-diabetic.23
Collection and analysis of small nerve C fibers in the skin have also been promoted to be an early marker for diabetic peripheral neuropathy and is a minimally invasive means of diagnosing and staging diabetic neuropathy.24 A study by Divisova et al comparing loss of intraepidermal nerve fibers between healthy controls and patients with type 2 diabetes with or without neuropathy found that the annual rate of nerve fiber loss in the skin was several times higher in diabetic subjects compared to controls irrespective of the presence of signs or symptoms of diabetic polyneuropathy.25 Comparing skin biopsies to corneal confocal microscopy in patients with recently diagnosed type 2 diabetes Ziegler et al found that corneal confocal microscopy detected early nerve fiber loss slightly more frequently than skin biopsy.2
Overall, it is encouraging that changes we found in small nerve densities in cornea and skin in rodent models of prediabetes and diabetes are consistent to those reported to occur in humans. To gain a better understanding of the progressive loss of small nerve fibers in the skin and cornea we have performed a longitudinal study in rats fed a high-fat diet, a model of prediabetes in parallel with a rat modeling late-stage type 2 diabetes, created by feeding rats a high-fat diet followed by a low dose of streptozotocin. Over three different time periods, density and function of small nerve fibers in the skin and cornea were examined along with motor and sensory nerve conduction velocity and vascular relaxation of epineurial arterioles that supply the sciatic nerve. We then examined the effect of menhaden (fish) oil treatment with intervention at the latest time point of the longitudinal study to determine the ability of these methods to detect nerve regeneration and functional recovery.
Materials and Methods
Unless stated otherwise all chemicals used in these studies were obtained from Sigma Aldrich Chemical Co. (St. Louis, MO).
Animals, Diets and Experimental Design
Sprague Dawley male rats were purchased at 11 weeks of age (Harlan Sprague Dawley, Indianapolis, IN). They were housed in the Iowa City Veterans Affairs Medical Center animal care facility, and food (Harlan Teklad, #7001, Madison, WI) and water were provided ad libitum. These studies followed institutional and NIH guidelines and were approved by the Iowa City Veterans Affairs Animal Care and Use Committee (# 1691101). At 12 weeks of age, the longitudinal study was initiated after randomly dividing 108 rats into 3 groups (control, diet-induced obesity and type 2 diabetes; 36 rats per group). The control group remained on the standard diet for the entire study period. This diet contained 4.25 gm% as fat. Rats in the other two groups were placed on a high-fat diet (D12451; Research Diets, New Brunswick, NJ). This diet contained 24 gm% fat (primarily from lard), 24 gm% protein and 41 gm% carbohydrate. After 8 weeks, one group of the high-fat-fed rats was treated with a low dose of streptozotocin (30 mg/kg, i.p.) (MP Biomedicals, Solon, OH) to induce hyperglycemia. These two groups of rats remained on the high-fat diet for the duration of the study (see illustration Supplemental Figure 1). Feeding rodents a high-fat diet is an accepted model for prediabetes since they have been shown to develop a state of impaired glucose tolerance and hyperinsulinemia. Rats fed a high-fat diet and latter treated with a low dose of streptozotocin model late-stage type 2 diabetes since these rats are insulin resistant, hyperglycemic and have low circulating levels of insulin. Studies of these three groups of rats were performed at 22, 28 and 36 weeks of age. For these 3 time points obese rats had been on the high-fat diet for 10, 16, or 24 weeks and the diabetic rats, in addition to being on the high-fat diet, had been hyperglycemic for 2, 8, or 16 weeks. Twelve rats were examined from each of the three groups at each time point.
For the intervention study, we began treatment of the rats with menhaden oil at the latest time point of the longitudinal study. Seventeen rats were initially assigned to five groups (control, diet-induced obesity with or without menhaden oil and diabetic with or without menhaden oil). These many rats were assigned to the groups because it was anticipated that we may lose 2–3 rats during the course of the study but no rats were lost. At the time of treatment 6 of the rats in the control group were diverted into a control with menhaden oil treatment group. For this reason, at the end of the study, the control group had 11 rats and the control with menhaden oil group had 6 rats. For diet-induced obese and diabetic rats treatment with menhaden oil was accomplished by placing them on a diet with 0.5 kcal of fat derived from lard of the high-fat diet replaced with menhaden oil (D16021504 Research Diets). For the control rats that were treated with menhaden oil, they were fed a diet enriched with an equivalent amount of menhaden oil to match the menhaden oil-enriched diets of the obese and diabetic rats. This diet was made by the laboratory. As shown in the forthcoming data, enriching the diet of control rats with menhaden had no effect on any of the outcome measures. The duration of the treatment was 16 weeks. During the 16-week treatment period the amount of diet consumed was determined at 4 different weekly intervals (3, 7, 11 and 15 weeks). The average amount of diet consumed by the control, control + menhaden oil, obese, obese + menhaden oil, diabetic and diabetic + menhaden oil was 43 ± 1, 29 ± 1, 28 ± 1, 30 ± 1, 48 ± 3 and 53 ± 4 g/kg body wt/day, respectively.
Glucose Clearance
To determine the glucose clearance rats were fasted overnight. The next morning rats were briefly anesthetized with isoflurane and injected (i.p.) with a saline solution of glucose in order to deliver a dose of 2 g/kg.26 Prior to this injection and at 15, 30, 45, 60, 120, 180 and 240 min post-injection, blood samples were obtained to measure circulating glucose levels using glucose-oxidase reagent Aviva Accu-Chek strips (Roche Diabetes Care, Inc., Indianapolis, IN).
Endpoints Related to Nerve Function and Vascular Reactivity
To rigorously analyze neural activity multiple endpoints were examined (see below for details). This included thermal sensitivity of the hindpaws and cornea sensitivity in un-anesthetized rats. Motor and Sensory nerve conduction velocity by electrophysiology and corneal nerves density by confocal microscopy were determined in rats anesthetized with sodium pentobarbital (50 mg/kg, i.p., Diamondback Drugs, Scottsdale, AZ). Afterwards, rats were euthanized by exsanguination and tissues were harvested to determine intraepidermal nerve fiber density of the hindpaw by immuno-histochemistry and vascular reactivity of epineurial arterioles of the sciatic nerve to acetylcholine and calcitonin gene-related peptide by videomicroscopy.10,11,23,27 Sample of the liver was also obtained.
Thermal nociceptive response in the hindpaw was measured using the Hargreaves method.26 The rat was placed in the observation chamber on top of the thermal testing apparatus and allowed to acclimate to the warmed glass surface (30°C) and surroundings for a period of 15 min. The mobile heat source was maneuvered so that it was under the heel of the hindpaw and then activated, a process that initiates a timer and locally warms the glass surface upon which the paw rests, when the rat withdrew its paw, the timer, and the heat source were turned off and the time was recorded. The heat source was set to go off by default after 25 sec to avoid injury to the rat. Following an initial recording, which was discarded, three measurements were made for each hindpaw, with a rest period of 5 min between each measurement. The mean of the measurements reported in seconds were used as the thermal nociceptive response.
Cornea sensitivity was determined by two separate procedures. A Cochet–Bonnet filament esthesiometer was used in un-anaesthetized rats (Luneau Ophtalmogie, France).27 The testing began with the nylon filament extended to the maximal length (6 cm). The end of the nylon filament was touched to the cornea. If the rat blinked (positive response) the length of the filament was recorded. If the rat did not blink then the nylon filament was shortened by 0.5 cm and the test repeated until a positive response was recorded. This process was repeated three times for each eye. Corneal sensation was also measured by applying buffered hyperosmotic eye drops Muro-128 solution 2% sodium chloride (684mOsm/liter) to the right eye of un-anaesthetized rats.10 The study was performed by placing conditioned animals in a custom-made restraining apparatus and allowing 5 minutes for the animal to acclimate to the restraint device and lighting. Six CMOS cameras (Imaging Development Systems GmbH, Obersulm, Germany), one positioned in front of the animal and two positioned laterally, were used to observe both eyes simultaneously. Custom software was used to synchronize video streams and obtain images (MATLAB R2012a, The MathWorks Inc., Natick, MA). Video was taken continuously at 30 fps of each animal as an isotonic sterile saline solution or Muro-128 solution containing 2% sodium chloride was delivered to the right eye. Temperature of the solutions was maintained at a 25 ± 1°C. During each recording epoch, 20 µL of solution was instilled and the response video-recorded for 300 sec after a 10 sec delay. An image collector was used offline to retrieve video frames every 6 sec from each eye drop concentration epoch; therefore 50 images were analyzed per animal. Fiji image analysis software was used by a masked technician to measure the visible surface area of both eyes between the upper and lower eyelids. In each recording epoch, the values obtained were presented as a percentage of the animals’ maximum visible eye surface area between the eyelids during that epoch and final data presented as area under the curve. Corneal sensitivity was examined at least 1 week prior to examination of corneal nerve fiber density. These procedures did not damage the rat’s cornea as no scratching of the cornea upon examination or behavioral abnormalities of the rats were observed.
On the day of terminal studies, rats were weighed and anesthetized with Nembutal i.p. (50 mg/kg, i.p., Abbott Laboratories, North Chicago, IL). Motor nerve conduction velocity was determined using a noninvasive procedure in the sciatic-posterior tibial conducting system.27 The left sciatic nerve was stimulated first at the sciatic notch and then at the Achilles tendon. Stimulation consisted of single 0.2-ms supramaximal (8 V) pulses through a bipolar electrode (Grass S44 Stimulator, Grass Medical Instruments, Quincy, MA). The evoked potentials were recorded from the interosseous muscle with a unipolar platinum electrode and displayed on a digital storage oscilloscope (model 54600A, Hewlett Packard, Rolling Meadows, IL). Motor nerve conduction velocity was calculated by subtracting the distal from the proximal latency measured in milliseconds from the stimulus artifact of the take-off of the evoked potential and the difference was divided into the distance between the two stimulating electrodes measured in millimeters using a Vernier caliper. Sensory nerve conduction velocity was recorded in the digital nerve to the second toe by stimulating with a square-wave pulse of 0.05-ms duration using the smallest intensity current that resulted in a maximal amplitude response. The sensory nerve action potential was recorded behind the medial malleolus. The maximal sensory nerve conduction velocity was calculated by measuring the latency to the onset/peak of the initial negative deflection and the distance between stimulating and recording electrodes. The motor and sensory nerve conduction velocity was reported in meters per second.
Corneal nerve density of the sub-epithelial corneal nerves was imaged using the Rostock cornea module of the Heidelberg Retina Tomograph confocal microscope.23 The anesthetized rat was secured to a platform that allows adjustment and positioning of the rat in three dimensions. A drop of the lubricant eye gel, GenTeal (Alcon Laboratories, Fort Worth, TX) was applied onto the tip of the lens and advanced slowly forward until the gel contacted the cornea of the right eye allowing optical but not physical contact between the objective lens and corneal epithelium. Ten random high-quality images without overlap of the sub-epithelial nerve plexus around the perimeter of the central cornea were acquired by finely focusing the objective lens to maximally resolve the nerve layer just under the corneal epithelium. It has been reported in human studies that acquiring eight random images not overlapping by more than 20% is a sufficient sample size for quantification of corneal sub-epithelial nerve parameters.28 The investigator acquiring these images was masked with respect to identity of the animal condition. For these studies, a single parameter of corneal innervation was quantified. Corneal nerve fiber length defined as the total length of all nerve fibers and branches (in millimeters) present in the acquired images standardized for area of the image (in square millimeters) was determined for each image by tracing the length of each nerve in the image, summing the total length and dividing by the image area. The corneal fiber length for each animal was the mean value obtained from the 10 acquired images and expressed as mm/mm2. Based on receiver operating characteristic (ROC) curve analysis corneal nerve fiber length is the optimal parameter for diagnosing patients with diabetic neuropathy and has the lowest coefficient of variation.18
Immunoreactive nerve fiber profiles innervating the skin from the hindpaw, which are primarily sensory nerves, were visualized using confocal microscopy.23 Samples of skin of the right hindpaw were fixed, dehydrated and embedded in paraffin. Three sections (7 µm each) for each animal were collected and immunostained with anti-PGP9.5 antibody, dilution 1:400 (rabbit anti-human, EMD Millipore Corp. Darmstadt, Germany) overnight followed by treatment with secondary antibody Alexa Fluor 488 goat anti-rabbit, dilution 1:1000 (Invitrogen/Thermo Fischer, Eugene, OR). Profiles were counted by two individual investigators that were masked to the sample identity. All immunoreactive profiles were counted and normalized to length.
Videomicroscopy was used to investigate the in vitro vasodilatory responsiveness of epineurial arterioles vascularizing the region of the sciatic nerve.29–31 The vessels used for these studies were generally oriented longitudinally in relation to the sciatic nerve; however, radially oriented vessels were also used on occasion. The arterioles used in this study should be regarded as epineurial rather than perineurial vessels. To isolate these vessels, the common iliac was exposed, and the branch points of the internal pudendal and superior gluteal arteries were identified. The internal pudendal and superior gluteal arteries and branches were dissected en bloc. The block of tissue was immediately submerged in a cooled (4°C), oxygenated (20% O2, 5% CO2, and 75% N2) Krebs-Henseleit physiological saline solution (PSS) of the following composition (in millimoles per liter): NaCl (118), KCl (4.7), CaCl2 (2.5), KH2PO4 (1.2), MgSO4 (1.2), NaHCO3 (20), Na2EDTA (0.026), and glucose (5.5). Branches of the superior gluteal and internal pudendal arteries (60- to 100-µm internal diameter and 1–2 mm in length) were carefully dissected and trimmed of fat and connective tissue. Both ends of the isolated vessel segment were cannulated with glass micropipettes filled with PSS (4°C) and secured with 10–0 nylon Ethilon monofilament sutures (Ethicon, Cornelia, GA). The pipettes were attached to a single pressure reservoir (initially set at 0 mmHg) under the condition of no flow. The organ chamber containing the cannulated vessels was then transferred to the stage of an inverted microscope (CK2; Olympus, Lake Success, NY). Attached to the microscope were a closed-circuit television camera (WV-BL200; Panasonic, Secaucus, NJ), a video monitor (Panasonic), and a video caliper (VIA-100K; Boeckeler Instruments, Tucson, AZ). The organ chamber was connected to a rotary pump (Masterflex; Cole Parmer Instrument, Vernon Hills, IL), which continuously circulated 37°C oxygenated PSS at 30 mL/min. The pressure within the vessel was then slowly increased to 40 mmHg. At this pressure, we found that KCl gave the maximal constrictor response. Therefore, all of the studies were conducted at 40 mmHg. Internal vessel diameter (resolution of 2 µm) was measured by manually adjusting the video micrometer. After a 30-min equilibration, 30 mM KCl was added to the bath to test vessel viability. Vessels failing to constrict by at least 30% were discarded. After they were washed with PSS, vessels were incubated for 30 min in PSS and then constricted with phenylephrine (10−7 to 10−6 M) to 30–50% of passive diameter. Afterwards, cumulative concentration–response relationships were evaluated for acetylcholine (10−8–10−4 M) and calcitonin gene-related peptide (10−11–10−8 M). At the end of each dose–response curve for acetylcholine or calcitonin gene-related peptide papaverine (10−5 M) was added to determine maximal vasodilation.
Fatty Acid Composition
To determine the extent menhaden oil treatment changed the fatty acid profile; serum and liver fatty acid composition was measured. Samples were extracted using chloroform/methanol followed by transesterification with 14% boron trifluoride in methanol. The derivatized samples were then extracted using heptane. The fatty acid methyl ester derivatives were resolved by gas-liquid chromatography and individual fatty acids peaks identified by comparison to known fatty acid standards.32 The individual fatty acid data were reported as % of total fatty acids present. These data were used to determine the unsaturation index, ratio of omega-6 to omega-3 polyunsaturated fatty acid and the anti-inflammatory fatty acid index in serum and liver. The anti-inflammatory fatty-acid index is the sum of EPA, DHA, and γ-linolenic acid divided by arachidonic acid.
Physiological Markers
Aviva Accu-Chek strips were used to determine non-fasting blood glucose. After exsanguination, serum was collected by centrifugation and used to determine levels of free fatty acid, triglyceride and free cholesterol. These assays were performed using commercial kits from Roche Diagnostics, Mannheim, Germany; Sigma Chemical Co., St. Louis, MO; Bio Vision, Mountain View, CA, respectively. For a marker of oxidative stress levels of thiobarbituric acid reactive substances were determined from serum according to a method described by Mihara et al.33
Data Analysis
Data are presented as mean ± SEM. Comparisons between the groups were conducted using one-way ANOVA and Bonferroni posttest comparison (Prism software; GraphPad, San Diego, CA). Prism software was also used to verify the normality of each data set. Concentration–response curves for acetylcholine and calcitonin gene-related peptide were compared using a two-way repeated measures analysis of variance with autoregressive covariance structure using proc mixed program of SAS.11,34,35 A p value of less than 0.05 was considered significant.
Results
Data in Supplemental Table 1 provide the weights, blood glucose, glucose clearance and serum-free fatty acid, triglyceride, and cholesterol levels for the rats at the time of termination for the three time points of the longitudinal study. All rats gained weight throughout the study. By the second time point high-fat diet-fed rats weighed significantly more than control rats, and diabetic rats although on a high-fat diet weighed significantly less than the high-fat diet-fed rats. Diabetic rats were significantly hyperglycemic at all three time point periods compared to control and high-fat diet-fed rats. High-fat diet-fed rats and to a greater extent, diabetic rats were insulin resistant as determined by the significant impairment in glucose clearance. Glucose clearance in control rats declined with age and progressively worsened in high-fat diet-fed rats. At all three time points, serum-free fatty acids and triglycerides were significantly increased in the diabetic rats compared to control and high-fat diet-fed rats. Serum cholesterol levels were significantly increased in the high-fat diet-fed and diabetic rats compared to control rats at the final time point.
Figure 1 provides data for motor (top) and sensory (bottom) nerve conduction velocity for rats of the three experimental conditions and three time periods. Motor nerve conduction velocity was significantly decreased in the high-fat diet-fed rats compared to control rats after 24 weeks of the high-fat diet. Motor nerve conduction velocity was significantly decreased in diabetic rats compared to control and high-fat diet-fed rats at all three time points. Sensory nerve conduction velocity was significantly greater in control rats at 22 weeks of age compared to control rats at 28 and 36 weeks of age. Sensory nerve conduction velocity was significantly decreased in high-fat diet-fed and diabetic rats at all three time points studied compared to control rats. Furthermore, sensory nerve conduction velocity was significantly decreased to a greater extent in high-fat diet-fed and diabetic rats at the second and third time points compared to the respective earlier time point of examination.
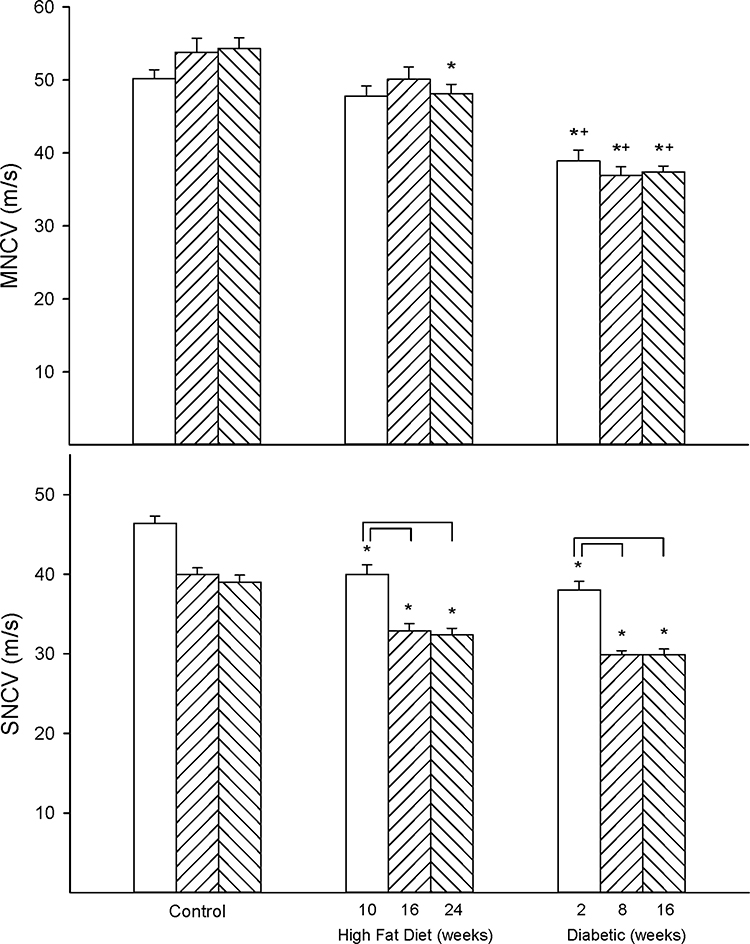 |
Figure 1 Effect of duration of obesity or type 2 diabetes on motor and sensory nerve conduction velocity. Motor (top) and sensory (bottom) nerve conduction velocity was examined as described in the 'Methods' section after 22 (open bars), 28 (left hatch bars) or 36 (right hatch bars) weeks of age in Sprague Dawley rats fed a normal diet or in rats fed a high-fat diet at 12 weeks of age for 10 (open bars), 16 (left hatch bars) or 24 (right hatch bars) weeks or in type 2 diabetic rats that were treated with a low dose of streptozotocin after 8 weeks of a high-fat diet and 2 (open bars), 8 (left hatch bars) or 16 (right hatch bars) weeks of hyperglycemia. Data are presented as the mean ± SEM. in m/s. The number of rats in each group was 12. *p < 0.05 compared to control rats; +p < 0.05 compared to high-fat diet-fed rats; brackets indicate significance (p < 0.05) compared to early time point for that condition. |
Figure 2 provides data for intraepidermal nerve fiber density (top) and corneal nerve fiber length (bottom). Intraepidermal nerve fiber density was significantly decreased early in both high-fat-diet-fed rats (10 weeks) and diabetic rats (10 weeks high-fat diet with 2 weeks hyperglycemia) compared to control rats but do not appear to be progressive. Loss of corneal nerve fiber length also occurs early with a significant decrease after 10 weeks of a high-fat diet and a significantly greater decrease in matched diabetic rats after 10 weeks of high-fat diet and 2 weeks of hyperglycemia compared to control rats. Unlike intraepidermal nerve fibers decrease in corneal nerve fiber length was progressive with increased loss of nerve fibers with duration of high-fat diet and/or hyperglycemia.
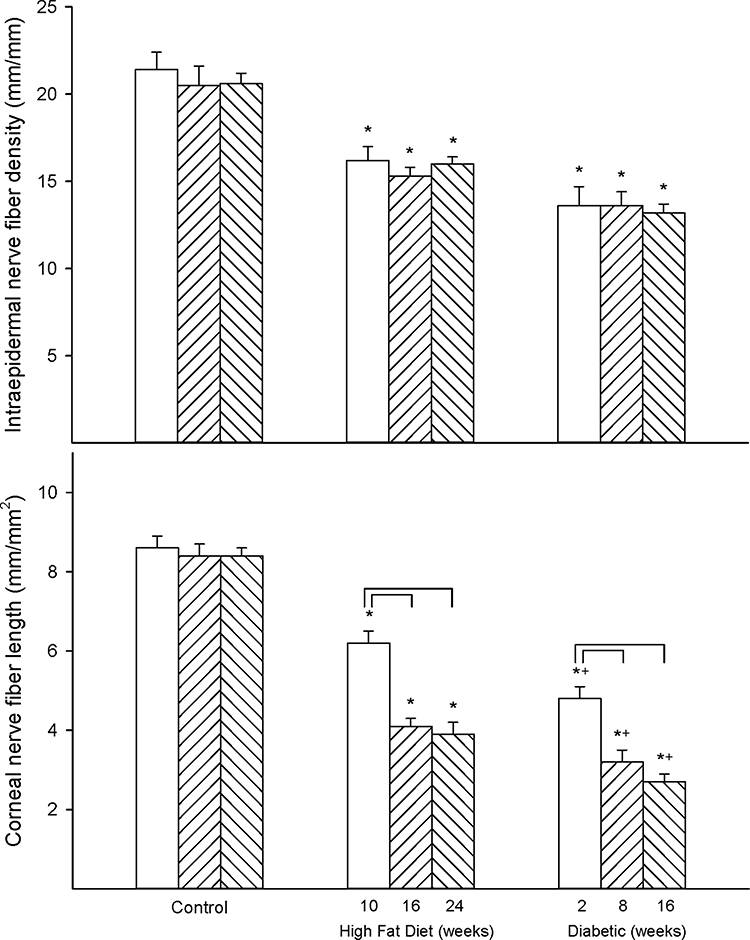 |
Figure 2 Effect of duration of obesity or type 2 diabetes on intraepidermal nerve fiber density and corneal nerve fiber length. Intraepidermal nerve fiber density (top) and corneal nerve fiber length (bottom) were examined as described in the 'Methods' section after 22 (open bars), 28 (left hatch bars) or 36 (right hatch bars) weeks of age in Sprague Dawley rats fed a normal diet or in rats fed a high-fat diet at 12 weeks of age for 10 (open bars), 16 (left hatch bars) or 24 (right hatch bars) weeks or in type 2 diabetic rats that were treated with a low dose of streptozotocin after 8 weeks of a high-fat diet and 2 (open bars), 8 (left hatch bars) or 16 (right hatch bars) weeks of hyperglycemia. Data are presented as the mean ± SEM. in fibers/mm for intraepidermal nerve fibers and mm/mm2 for corneal nerve fiber length. The number of rats in each group was 12. *p < 0.05 compared to control rats; +p < 0.05 compared to high-fat diet-fed rats; brackets indicate significance (p < 0.05) compared to early time point for that condition. |
Figure 3 provides data for thermal nociception in high-fat diet-fed and diabetic rats. Thermal sensitivity was significantly decreased in both high-fat diet-fed and diabetic rats to a similar extent after 16 weeks of high-fat diet with or without 8 weeks of hyperglycemia compared to control rats. Figure 4 provides data for the effect of high-fat diet with or without hyperglycemia on cornea sensitivity measured by two methodologies. Using Cochet–Bonnet filament esthesiometer, a significant decrease in corneal sensitivity was detected early in high-fat diet-fed rats (10 weeks of high-fat diet) and diabetic rats (10 weeks of high-fat diet and 2 weeks of hyperglycemia) compared to control rats (Figure 4 top). Cornea sensitivity continued to decrease with the duration of high-fat diet and/or hyperglycemia compared to the early time point in both models. Furthermore, cornea sensitivity was more significantly decreased in diabetic rats at the two later time points compared to high-fat diet-fed rats. Cornea sensitivity was also determined by measuring the squinting response to a hyperosmotic solution (Figure 4 bottom).10 Using this approach, a significant decrease in corneal sensitivity was also detected early in high-fat diet-fed and diabetic rats compared to control rats. This methodology did not detect any progression in loss of sensation with the duration of high-fat diet or diabetes and there was no difference in the decrease in cornea sensitivity between high-fat diet-fed rats and diabetic rats.
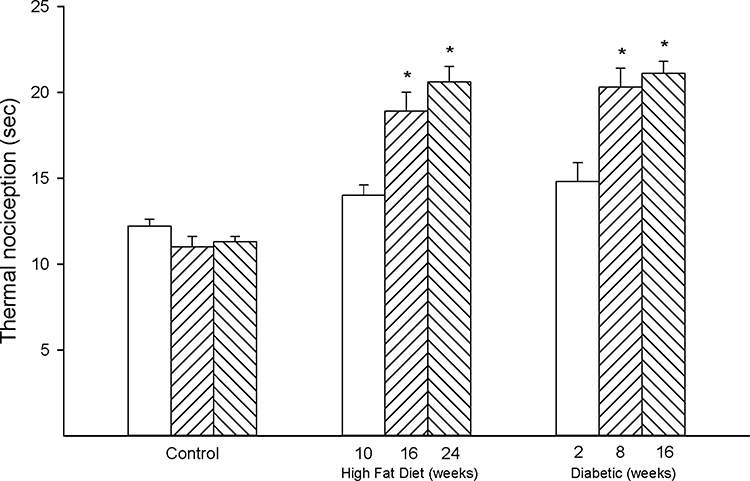 |
Figure 3 Effect of duration of obesity or type 2 diabetes on thermal nociception. Thermal nociception was examined as described in the 'Methods' section after 22 (open bars), 28 (left hatch bars) or 36 (right hatch bars) weeks of age in Sprague Dawley rats fed a normal diet or in rats fed a high-fat diet at 12 weeks of age for 10 (open bars), 16 (left hatch bars) or 24 (right hatch bars) weeks or in type 2 diabetic rats that were treated with a low dose of streptozotocin after 8 weeks of a high-fat diet and 2 (open bars), 8 (left hatch bars) or 16 (right hatch bars) weeks of hyperglycemia. Data are presented as the mean ± SEM. in sec. The number of rats in each group was 12. *p < 0.05 compared to control rats. |
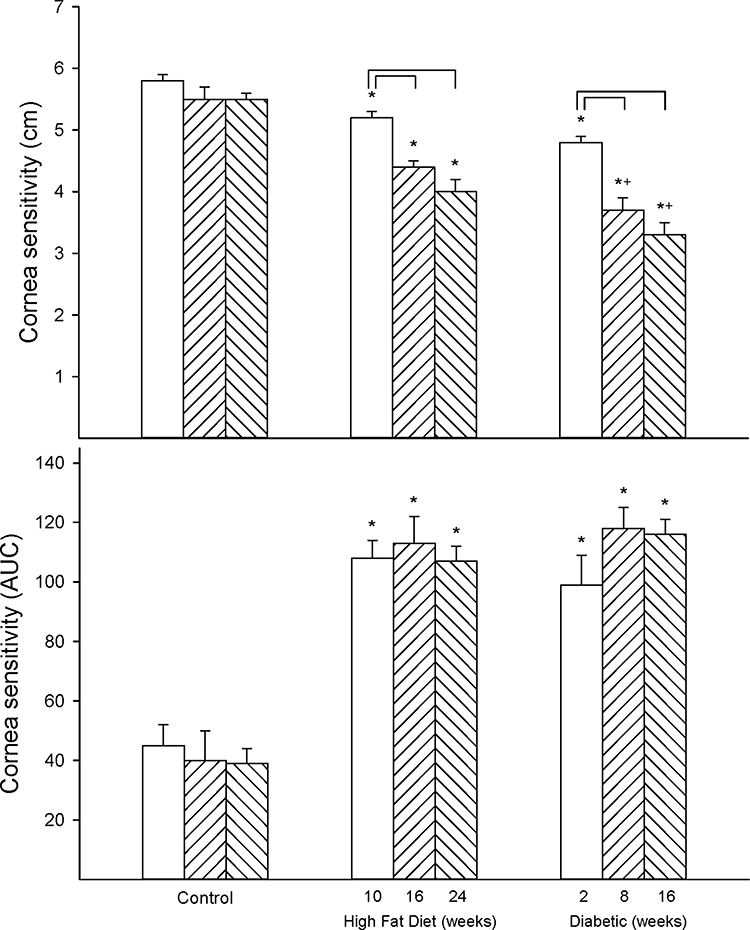 |
Figure 4 Effect of duration of obesity or type 2 diabetes on cornea sensitivity. Cornea sensitivity was determined using a Cochet–Bonnet filament esthesiometer (top) or by response to a hyperosmolar solution (bottom) as described in the 'Methods' section after 22 (open bars), 28 (left hatch bars) or 36 (right hatch bars) weeks of age in Sprague Dawley rats fed a normal diet or in rats fed a high-fat diet at 12 weeks of age for 10 (open bars), 16 (left hatch bars) or 24 (right hatch bars) weeks or in type 2 diabetic rats that were treated with a low dose of streptozotocin after 8 weeks of a high-fat diet and 2 (open bars), 8 (left hatch bars) or 16 (right hatch bars) weeks of hyperglycemia. Data are presented as the mean ± SEM. in cm for determination of cornea sensitivity by the filament esthesiometer and area under the curve (AUC) for cornea sensitivity by the hyperosmolar solution approach. The number of rats in each group was 12. *p < 0.05 compared to control rats; +p < 0.05 compared to high-fat diet-fed rats; brackets indicate significance (p < 0.05) compared to early time point for that condition. |
We have previously reported that decrease in vascular relaxation to acetylcholine by the small blood vessels that supply circulation to the sciatic nerve occurs early in diabetic rats.29 Data in Figure 5 demonstrate that decrease in vasodilation to acetylcholine by epineurial arterioles of the sciatic nerve was progressive in both the high-fat diet-fed and type 2 diabetic rat models compared to age-matched control rats. Moreover, the decrease in vascular relaxation to acetylcholine was impaired to a greater extent in diabetic rats compared to high-fat diet-fed rats. We have also previously reported that calcitonin gene-related peptide is associated with sensory neurons innervating epineurial arterioles and is a potent vasodilator of these blood vessels.36 Data in Figure 6 demonstrate that vascular relaxation to calcitonin gene-related peptide is impaired in a progressive manner in both high-fat diet-fed and diabetic rats at physiological doses compared to control rats. Impairment is also greater in diabetic rats compared to high-fat diet-fed rats at 10−9 M calcitonin gene-related peptide.
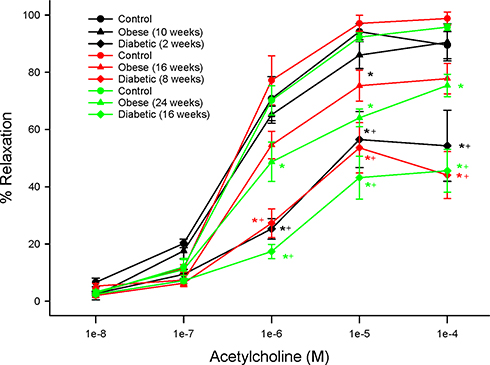 |
Figure 5 Effect of duration of obesity or type 2 diabetes on vascular reactivity to acetylcholine by epineurial arterioles of the sciatic nerve. Vascular reactivity was determined as described in the 'Methods' section after 22 (black circles), 28 (red circles) or 36 (green circles) weeks of age in Sprague Dawley rats fed a normal diet or in rats fed a high-fat diet at 12 weeks of age for 10 (black triangles), 16 (red triangles) or 24 (green triangles) weeks or in type 2 diabetic rats that were treated with a low dose of streptozotocin after 8 weeks of a high-fat diet and 2 (black diamonds), 8 (red diamonds) or 16 (green diamonds) weeks of hyperglycemia. Data are presented as the mean ± SEM. in % relaxation. The number of rats in each group was 12. *p < 0.05 compared to control rats; +p < 0.05 compared to high-fat diet-fed rats. |
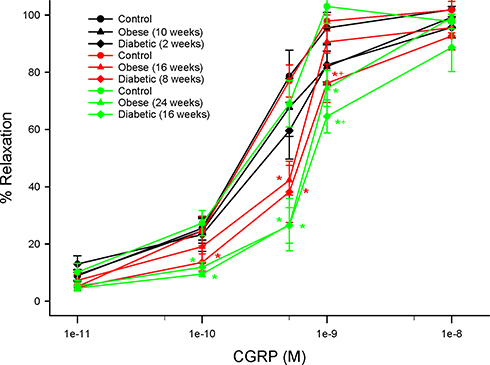 |
Figure 6 Effect of duration of obesity or type 2 diabetes on vascular reactivity to calcitonin gene-related peptide by epineurial arterioles of the sciatic nerve. Vascular reactivity was determined as described in the 'Methods' section after 22 (black circles), 28 (red circles) or 36 (green circles) weeks of age in Sprague Dawley rats fed a normal diet or in rats fed a high-fat diet at 12 weeks of age for 10 (black triangles), 16 (red triangles) or 24 (green triangles) weeks or in type 2 diabetic rats that were treated with a low dose of streptozotocin after 8 weeks of a high-fat diet and 2 (black diamonds), 8 (red diamonds) or 16 (green diamonds) weeks of hyperglycemia. Data are presented as the mean ± SEM. in % relaxation. The number of rats in each group was 12. *p < 0.05 compared to control rats; +p < 0.05 compared to high-fat diet-fed rats. |
In the intervention experiment, we examined whether the neural and vascular damage induced by a chronic high-fat diet with or without hyperglycemia was reversible and whether recovery of nerve fiber loss and function in the skin and cornea could be markers for regeneration. For this study, we treated obese or type 2 diabetic rats with menhaden oil for 16 weeks following 24 weeks of a high-fat diet (obese) or 24 weeks of a high-fat diet and 16 weeks of hyperglycemia (diabetic), respectively. Treatment consisted of replacing 50% of the kcal of the high-fat diet derived from lard with menhaden oil. Six control rats were also treated with a similar amount of menhaden oil incorporated into the normal diet. We have previously demonstrated the success of this approach in treating obese and type 2 diabetic rats with dietary oils.35,37,38 At the beginning of this study rats in all six groups weighed about the same (Table 1). At the end of the study, obese rats treated with or without menhaden oil weighed significantly more than the control or diabetic rats treated with or without menhaden oil, respectively. At the end of the study, the weight of diabetic rats was not significantly different than control rats. Non-fasting blood glucose levels were significantly increased in diabetic rats compared to control or obese rats. Treating rats with menhaden oil had no effect on blood glucose levels and glucose clearance remained impaired after treatment of obese and diabetic rats with menhaden oil (data not shown). Weight of the left epididymal fat pad was significantly increased in obese rats compared to control rats and untreated diabetic rats. Treating control or obese rats with menhaden oil did not affect the weight of the left epididymal fat pad. However, treating diabetic rats with menhaden significantly decreased the weight of the left epididymal fat pad compared to obese rats treated with or without menhaden oil and untreated diabetic rats. Serum-free fatty acid levels trended to be higher in obese rats compared to control rats and this trend was not altered by treating control or obese rats with menhaden oil. Serum-free fatty acid levels were significantly increased in diabetic rats and these levels were decreased in diabetic rats treated with menhaden oil. Serum triglyceride and cholesterol levels were significantly increased in obese rats compared to control rats and improved when obese rats were treated with menhaden oil. Serum triglyceride and cholesterol levels were increased to a greater extent in diabetic rats. Treating diabetic rats with menhaden oil trended to improve serum triglyceride and cholesterol levels but they remained significantly increased compared to control rats. Serum thiobarbituric acid levels, a marker for oxidative stress, were significantly increased in obese and diabetic rats compared to control rats. Treating obese and diabetic rats with menhaden oil reduced serum thiobarbituric acid levels.
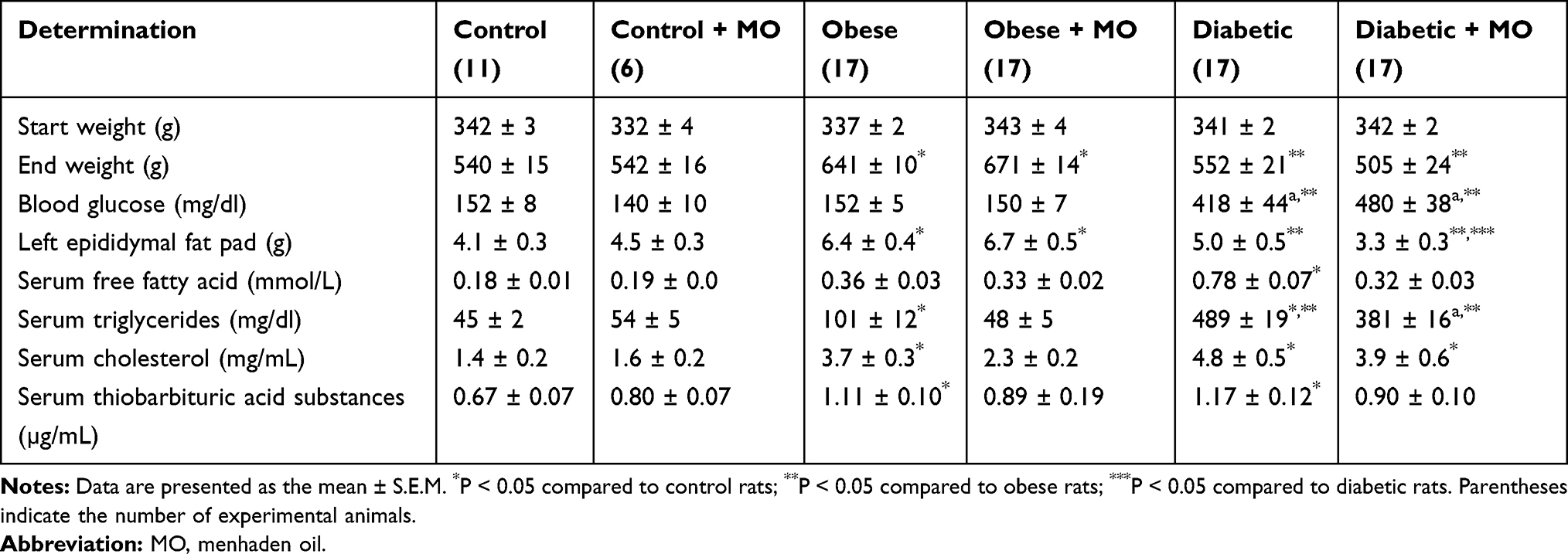 |
Table 1 Effect of Menhaden Oil Treatment on Weight Gain, Serum Lipids and Thiobarbituric Acid Substances in Diet-Induced Obese and Type 2 Diabetic Sprague Dawley Rats |
Data in Supplemental Tables 2 and 3 provide the fatty acid composition of the serum and liver from control, high-fat fed and diabetic rats treated with or without menhaden oil, respectively. As expected treating rats with menhaden oil significantly increased levels of eicosapentaenoic and docosahexaenoic acids in serum and liver of control, obese and diabetic rats. This was accompanied by a significant decrease in linoleic and arachidonic acid levels. Table 2 provides data on the unsaturation index, omega-6 to omega-3 polyunsaturated fatty acid ratio, and anti-inflammatory index for serum and liver of control, obese and diabetic rats treated with or without menhaden oil, respectively. The unsaturation indices of serum and liver are significantly increased in control, obese and diabetic rats treated with menhaden oil compared to their respective untreated rats. The omega-6 to omega-3 polyunsaturated fatty acid ratio was significantly decreased and the anti-inflammatory fatty acid index was significantly increased in control, obese and diabetic rats treated with menhaden oil compared to respective untreated rats. Data in Figure 7 provide the serum resolvin D1 levels for control rats and obese and diabetic rats treated with or without menhaden oil. At the end of the study, resolvin D1 levels were significantly decreased in obese and diabetic rats. Treating obese rats with menhaden oil significantly improved serum resolvin D1 levels compared to untreated obese rats. Treating diabetic rats with menhaden oil-enriched diet significantly increased resolvin D1 levels compared to untreated diabetic rats as well as untreated and treated obese rats. Treating control rats with menhaden oil did not lead to an increase in resolvin D1 levels.
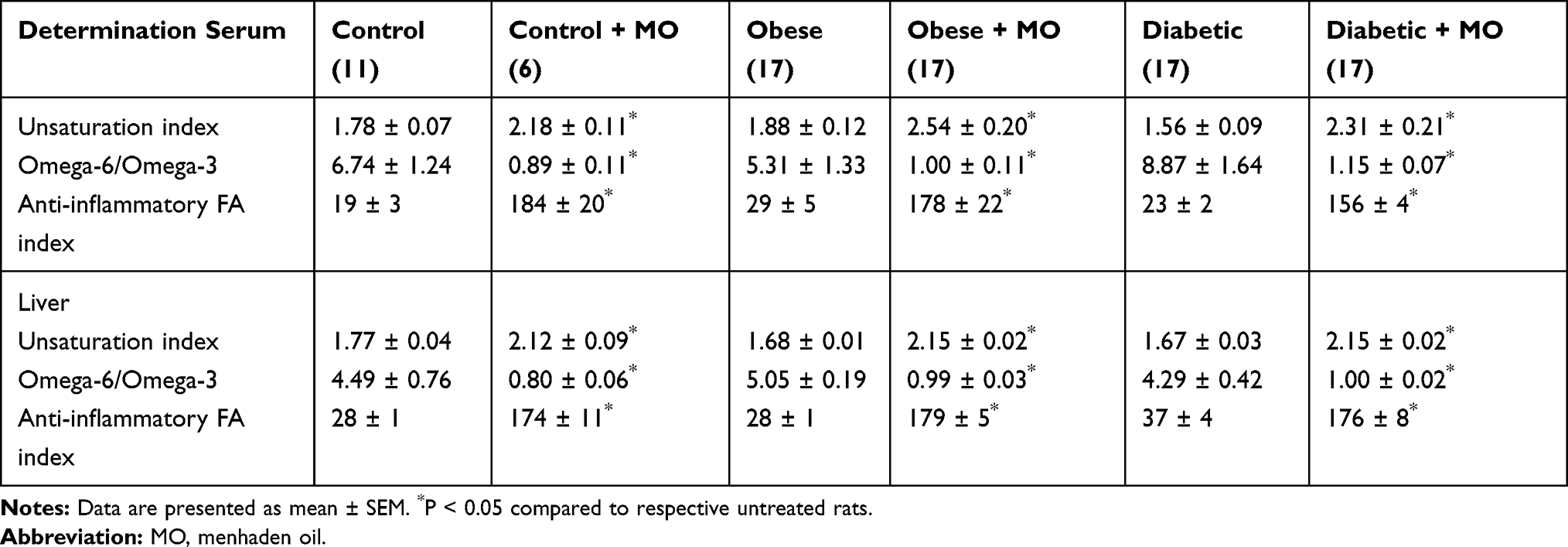 |
Table 2 Treatment with Menhaden Oil of Diet-Induced Obese or Type 2 Diabetic Rats Effect on Unsaturation Index, Omega-6 to Omega-3 Fatty Acid Ratio and Anti-Inflammatory Fatty Acid Index in Serum and Liver |
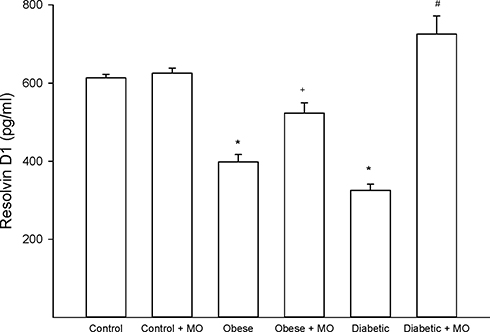 |
Figure 7 Effect of treating diet-induced obese or type 2 diabetic rats with menhaden oil on serum resolvin D1 levels. Serum resolvin D1 levels was determined as described in the 'Methods' section. Obese (12-week old rats fed a high-fat diet for 24 weeks) and diabetic (12-week old rats fed a high-fat diet for 24 weeks and hyperglycemic for 16 weeks) rats were treated with menhaden oil for 16 weeks by replacing 50% of the kcal in the high-fat diet derived from lard with menhaden oil. Control rats treated with or without menhaden oil were aged matched. Data are presented as the mean ± SEM. in pg/mL. The number of rats in each group was 12 except for control + menhaden oil the number of rats in this group was 6. *p < 0.05 compared to control rats; +p < 0.05 compared to obese rats; #p < 0.05 compared to diabetic rats. |
Table 3 presents data for multiple neural pathology-related endpoints and the effect of treating chronic obese and type 2 diabetic rats with menhaden oil for 16 weeks. These same endpoints were examined for the longitudinal study. Treating control rats with menhaden oil had no effect on any of the neurologic endpoints examined. After 40 weeks of a high-fat diet motor and sensory nerve conduction velocities were significantly decreased in obese rats. Treating obese rats for the final 16 weeks of the 40-week period prevented the decrease in motor nerve conduction velocity and reversed the slowing of sensory nerve conduction velocity. As shown above, motor and sensory nerve conduction velocities were significantly decreased in type 2 diabetic rats and treating these rats with menhaden oil significantly improves both.
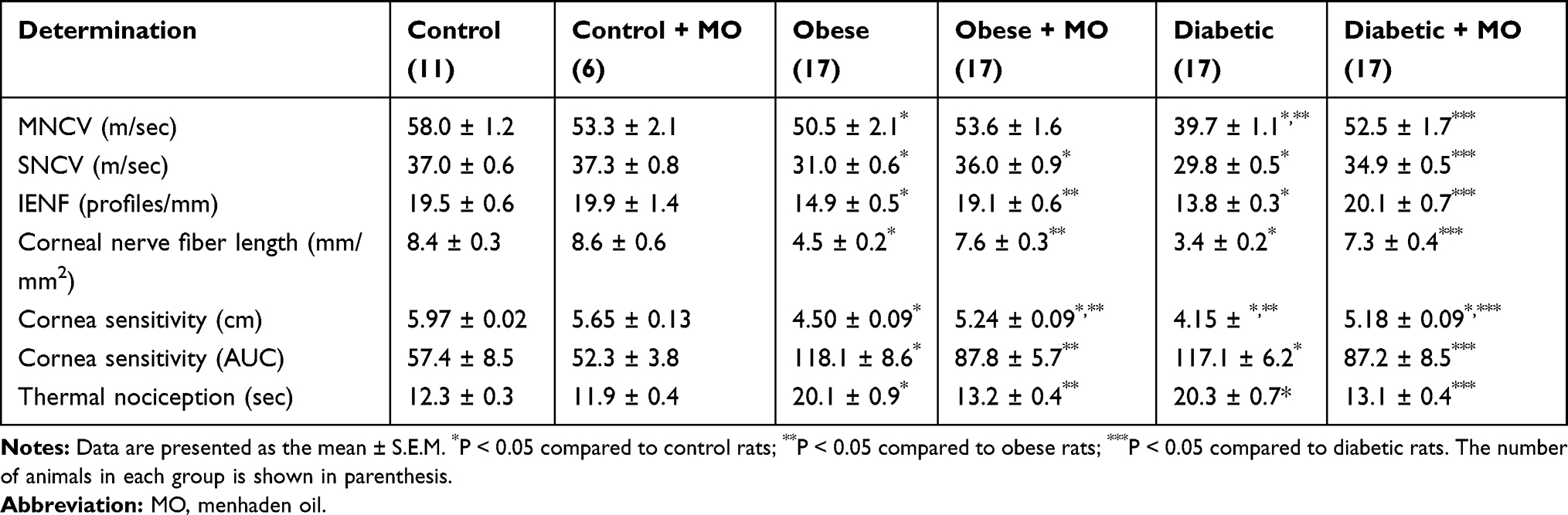 |
Table 3 Effect of Late Intervention with Menhaden Oil on Peripheral Neuropathy in Diet-Induced Obese and Type 2 Diabetic Sprague Dawley Rats |
Structure and function of sensory fibers in the skin and cornea are being promoted as a means of early detection and treatment for diabetic neuropathy.3,17,24,39 In this study, we demonstrate that treating chronic obese or diabetic rats with menhaden oil can reverse loss of small sensory nerve fibers in the skin or cornea. Data in Supplemental Figure 2 provide representative images of corneal nerves from control, obese and diabetic rats treated with or without menhaden oil. In addition, cornea sensitivity and thermal nociception of the hindpaw that were significantly impaired by long-term obesity and type 2 diabetes in rats were significantly improved by treating the rats with menhaden oil. As shown in Figures 5 and 6 vascular relaxations to acetylcholine and calcitonin gene-related peptide by epineurial arterioles of the sciatic nerve were impaired by chronic obesity and type 2 diabetes. Data in Figures 8 and 9 demonstrate that treating chronic obese and diabetic rats with menhaden oil significantly improved vasodilation to acetylcholine and calcitonin gene-related peptide, respectively.
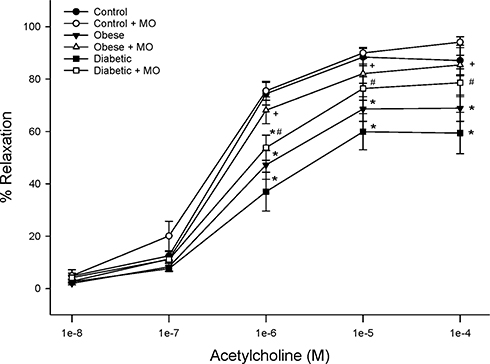 |
Figure 8 Effect of treating control, diet-induced obese or type 2 diabetic rats with menhaden oil on vascular reactivity to acetylcholine by epineurial arterioles of the sciatic nerve. Vascular reactivity was determined as described in the 'Methods' section. Control (12-week old rats fed a normal diet for 24 weeks), obese (12-week old rats fed a high-fat diet for 24 weeks) and diabetic (12-week old rats fed a high-fat diet for 24 weeks and hyperglycemic for 16 weeks) rats were treated with menhaden oil for 16 weeks by replacing 50% of the kcal in the high-fat diet derived from lard with menhaden oil. Data are presented as the mean ± SEM. in % relaxation. The number of rats in each group was 12. *p < 0.05 compared to control rats; +p < 0.05 compared to diet-induced obese rats; #p < 0.05 compared to diabetic rats. |
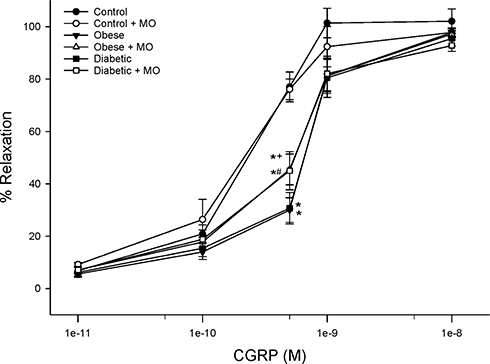 |
Figure 9 Effect of treating control, diet-induced obese or type 2 diabetic rats with menhaden oil on vascular reactivity to calcitonin gene-related peptide by epineurial arterioles of the sciatic nerve. Vascular reactivity was determined as described in the 'Methods' section. Control (12-week old rats fed a normal diet for 24 weeks), obese (12-week old rats fed a high-fat diet for 24 weeks) and diabetic (12-week old rats fed a high-fat diet for 24 weeks and hyperglycemic for 16 weeks) rats were treated with menhaden oil for 16 weeks by replacing 50% of the kcal in the high-fat diet derived from lard with menhaden oil. Data are presented as the mean ± SEM. in % relaxation. The number of rats in each group was 12.*p < 0.05 compared to control rats; +p < 0.05 compared to obese rats; #p < 0.05 compared to diabetic rats. |
Discussion
The primary findings of this study were that a decrease in cornea fiber length and sensitivity can be detected early in rat models of diet-induced obesity/prediabetes and type 2 diabetes and that intervention with menhaden oil after chronic obesity or type 2 diabetes reversed multiple endpoints associated with peripheral neuropathy including corneal nerve morphology and function. In addition, decrease in corneal nerve fiber length and cornea sensitivity as measured by corneal confocal microscopy and Cochet–Bonnet filament esthesiometer, respectively, was progressive in both animal models. There was a highly significant and early decrease in cornea sensitivity when measured by response to a hyperosmotic solution in both rat models, but this approach did not demonstrate progression. This is not surprising since this approach likely measures primarily the sensitivity of the nerve C-fibers that penetrate into the epithelial layer of the cornea. These nerve fibers respond to thermal and chemical stimuli.40,41 We have previously demonstrated that these nerve fibers are the first to decrease in diabetes.27 In contrast, the Cochet–Bonnet filament esthesiometer measures the sensitivity of primarily the Aδ nerve fibers located at the sub-epithelial layer of the cornea.40,41 The Aδ fibers are mechanosensory receptors that respond to mechanical stimuli.40,41 Loss of these fibers in diabetes occurs at a slower rate and thus the finding that cornea sensitivity is decreased at a slower rate and is progressive through use of the Cochet–Bonnet filament esthesiometer is reasonable.23 Our intervention study with menhaden oil treatment initiated at an advanced stage of obesity and type 2 diabetes clearly demonstrated that the decrease in cornea nerve fiber length and sensitivity was reversible as were the conventional neuropathology endpoints, motor and sensory nerve conduction velocity, as well as vascular reactivity to acetylcholine and calcitonin gene-related peptide of epineurial arterioles of the sciatic nerve. These findings support the premise that examination of corneal nerve fiber density and function may be an early marker of peripheral neuropathy and repair.7 Our longitudinal study also demonstrated that decrease in intraepidermal nerve fiber density and thermal nociception occurred early in both diet-induced obesity and type 2 diabetes. However, in our studies, the decrease in intraepidermal nerve fiber density did not decline further with the duration of the obese state or type 2 diabetes. This could be due to the location used to measure intraepidermal nerve fiber density in rodents is the hindpaw. This site is the most distal region for these fibers in the rodent and would be the initial site to detect decrease since axonal degeneration is believed to follow a distal-to-proximal pattern or “dying-back phenomenon”. In human subjects, detection of a decrease in intraepidermal nerve fiber density is dependent on the duration of diabetes and can be influenced by the location on the leg where the biopsy is collected.42 Sensitivity to thermal challenge of the hindpaw was progressive; however, a significant increase in latency was not detected until the intermediate study period. This could be due to signaling pathways that include action of calcitonin gene-related peptide and substance P are not sufficiently impaired early in obesity or type 2 diabetes to cause an increase in thermal latency. Thus, determining thermal nociception of the hindpaw was not as sensitive as corneal sensitivity for detecting early loss of function in obesity or type 2 diabetes. Our longitudinal studies of vascular reactivity of epineurial arterioles of the sciatic nerve also demonstrated that loss of functionality of these blood vessels, especially to acetylcholine, occurred early and was progressive in both diet-induced obesity and type 2 diabetes and was more severe in vessels derived from diabetic rats than in vessels derived from obese rats. This latter finding further supports our previous studies demonstrating that vascular dysfunction of blood vessels that provide circulation to peripheral nerves occurs early in metabolic-related diseases such as obesity and diabetes and is likely a contributing factor to peripheral neuropathy.23,29,43
In addition to early detection, a valid marker for diabetic peripheral neuropathy should also be useful for monitoring the success or failure of treatment. Previous studies have demonstrated early regeneration of corneal nerves detected by corneal confocal microscopy in patients with type 1 diabetes following pancreas transplant.44,45 In patients receiving a simultaneous pancreas and renal transplant besides improvement in corneal nerves 36 months following surgery, the Neuropathy Symptom Profile and peroneal nerve conduction velocity also improved significantly but not intraepidermal nerve fiber density, quantitative sensory testing or cardiac autonomic function assessments.45 It has also been reported that corneal confocal microscopy can detect improvement of corneal parameters in diabetic subjects following improved glycemic control and reduction of other risk factors for diabetic neuropathy.46,47 Whether interventions providing prolonged improvement of diabetes-induced corneal nerve fiber damage will eventually lead to improvement in multiple standard neuropathic endpoints, such as nerve conduction velocities, will require extensive investigation. However, previous studies from my laboratory and this current study provide preclinical evidence that interventions that improve corneal nerve fiber length and function also improved motor and sensory nerve conduction velocities and other parameters associated with diabetic peripheral neuropathy including intraepidermal nerve fiber density, thermal nociception and vascular reactivity of epineurial arterioles.11,27,34,35,37,38
In this study, we demonstrated that treating rats after chronic obesity (24 weeks of a high-fat diet) or type 2 diabetes (24 weeks of high-fat diet with 16 weeks of hyperglycemia) with menhaden (fish) oil for 16 weeks reversed multiple endpoints associated with peripheral neuropathy. Presently, the only recognized treatment for diabetic peripheral neuropathy is maintaining adequate glycemic control, but even this has been shown to be ineffective in thwarting the progression or development of peripheral neuropathy in subjects with type 2 diabetes.48–50 Many individuals supplement their diet with fish oil capsules. These capsules are enriched with long-chain omega-3 polyunsaturated fatty acids, primarily eicosapentaenoic and docosahexaenoic acids. Our preclinical studies suggest that fish oil and these fatty acids may be an effective treatment for diabetic peripheral neuropathy.34,35,38,51 Furthermore, our studies have demonstrated that E and D series resolvins (resolution-phase interaction products), metabolites of eicosapentaenoic and docosahexaenoic acids, respectively, which are known to have anti-inflammatory and neuro-protective properties, may be the active mediators responsible for the effect of fish oil on peripheral neuropathy and nerve regeneration.51–53 Zhang et al performed a systematic review and meta-analysis titled omega-3 polyunsaturated fatty acid oral supplements for improving peripheral nerve health.54 Most of the randomized trials cited in this review focused on chemotherapy-induced peripheral neuropathy and the authors concluded that there was little support of a therapeutic benefit for omega-3 fatty acid oral supplementation in this patient cohort. In regard to progressive peripheral neuropathy caused by diabetes, the authors stated that there was not enough evidence available to determine whether omega-3 fatty acid supplementation is beneficial.54 Importantly, the authors of this article state the need for appropriately powered, well-designed clinical trials to investigate the potential therapeutic effects of omega-3 polyunsaturated fatty acid oral supplements on peripheral nerve integrity that includes analyses of small and large fiber neuropathies. Most previous studies examining the effect of omega-3 fatty acid oral supplements for many different diseases including cardiovascular disease have failed to measure the omega-3 index or production of metabolites of omega-3 polyunsaturated fatty acids. For example, in the recent ASCEND study, 15,480 subjects with diabetes received 1 g capsules daily containing omega-3 fatty acids or placebo. In this study, the primary endpoint was a first serious vascular event. After a mean follow-up period of 7.4 years, the study found no significant difference in the risk of serious vascular events between those assigned to receive omega-3 fatty acids or placebo.55 In contrast, the REDUCE-IT trial enrolled 8179 subjects that included subjects with diabetes to receive a 4 g daily dose of icosapent ethyl, a highly enriched form of EPA, or placebo. After a follow-up period of 4.9 years, icosapent ethyl was found to have a statistical benefit on reducing ischemic events in subjects with hypertriglyceridemia.56 Was the apparent success of the REDUCE-IT trial due to increased dosing of the omega-3 polyunsaturated fatty acid derivative, study design or other factors? This question is difficult to answer because neither trial analyzed circulation levels of omega-3 polyunsaturated fatty acid or their metabolites before and after treatment. Future trials need to thoroughly address issues of dosage, source and type of omega-3 polyunsaturated fatty acid administered and ratio of eicosapentaenoic and docosahexaenoic acids in the blood.
In this study, we demonstrated that treating chronic obese or type 2 diabetic rats with menhaden oil reduced serum thiobarbituric acid substances, significantly reduced serum and liver omega-6 to omega-3 ratio and increased the anti-inflammatory fatty acid index indicating improvement in oxidative and inflammatory stress following treatment with menhaden oil. In addition to improving oxidative and inflammatory stress, omega-3 polyunsaturated fatty acids have been shown to affect a myriad of molecular pathways, including alteration of physical properties of cellular membranes, direct interactions with and modulation of membrane channels and proteins, regulation of gene expression via nuclear receptors and transcription factors, changes in eicosanoid profiles, and conversion to bioactive metabolites.57,58 As discussed above, unlike studies related to cardiovascular disease, in diabetes the role of omega-3 polyunsaturated fatty acids/fish oil as a potential treatment for complications including diabetic peripheral neuropathy, have not been adequately studied.54,59-61 Data in Table 2 indicate that the levels of omega-3 polyunsaturated fatty acids appearing in the serum of obese and diabetic rats with diet supplementation of menhaden oil can be achieved in human subjects taking 4 g of fish oil capsules per day.62,63 In a study by Laidlaw and Holub, the omega-3 polyunsaturated fatty acid levels in serum increased from 5.6% to 14.4% after taking 4 g of fish oil (each 1 g capsule contained 35% EPA and 25% DHA and capsule supplementation was adjusted to provide the intended average daily intake of 4 g) for 4 weeks and the eicosapentaenoic acid to arachidonic acid ratio increased from 0.12 to 0.88.63 However, this study also reported that 16% of the enrolled subjects failed to achieve the target EPA/arachidonic acid ratio in the blood.63 This further stresses the importance of measuring the responsiveness of each subject to omega-3 polyunsaturated fatty acid supplementation. In our study of diabetic rats treated with menhaden oil, the omega-3 polyunsaturated fatty acid levels in serum increased from 3.9% to 15.4% and the eicosapentaenoic acid to arachidonic acid ratio increased from 0.04 to 0.54.34
Lewis et al have demonstrated that treating subjects with type 1 diabetes for 12 months daily with 2330 mg of omega-3 polyunsaturated fatty acids derived from seal oil increased corneal nerve fiber length by 29%. In this study, 1 year of treatment was found to be insufficient to improve nerve conduction or sensory function.60 In a randomized, double-masked, placebo-controlled clinical trial it was found that krill oil (945 mg eicosapentaenoic acid and 510 mg of docosahexaenoic acid/day) or fish oil (1000 mg eicosapentaenoic acid and 500 mg of docosahexaenoic acid/day) improved dry eye disease compared to placebo (olive oil 1500 mg/day).64
In this and our other studies, we have found that treating rats with type 1 or type 2 diabetes with menhaden oil improves corneal nerve sensitivity and density as well as improves motor and sensory nerve conduction velocities and intraepidermal nerve fiber density and thermal nociception.11,34,35,38 He et al have shown that treatment with docosahexaenoic acid and pigment epithelium-derived factor of diabetic mice increased corneal nerve regeneration, improved corneal sensitivity and accelerated corneal wound healing.65 Zhang et al have shown that topical application of resolvin D1, a metabolite of docosahexaenoic acid, to the eye of diabetic mice following corneal injury promoted regeneration of corneal epithelium and nerves and restored mechanical sensation.66 In our studies, we have shown that resolvin D1 increases neurite growth by primary cultures of dorsal ganglion neurons and improves corneal nerve density and neural function of diabetic mice.51–53
Conclusion
The primary findings from the longitudinal studies were that examination of corneal nerve sensitivity and morphometry are potentially useful outcome measures for the early detection and treatment of peripheral neuropathy. Our studies demonstrated that loss of corneal nerve sensitivity and structure occurs not only in a rat model of type 2 diabetes but also in a rat model of diet-induced obesity, modeling prediabetes. The discovery of a reliable early marker for peripheral neuropathy could revolutionize design of clinical trials for this complication that affects about 50% of the diabetes population as well as earlier treatment. In addition, our late intervention studies support advancing omega-3 polyunsaturated fatty acids/fish oil to clinical trials for the treatment of diabetic peripheral neuropathy. Comparisons between human studies of fish oil supplementation have demonstrated that the level of omega-3 polyunsaturated fatty acids achieved in the rodent serum through diet modification that has been found to improve diabetic peripheral neuropathy in our studies can be achieved in human serum through taking fish oil supplements. The excellent safety profile makes fish oil supplementation an attractive choice for treatment of diabetic peripheral neuropathy.
Acknowledgments
Research reported in this publication was supported by the Department of Veterans Affairs, Veterans Health Administration, Office of Research and Development Merit award [RX000889-06] and by the National Institute of Diabetes and Digestive and Kidney Diseases of the National Institutes of Health award [DK107339-04]. The content of this manuscript are new and solely the responsibility of the authors, and do not necessarily represent the official views of the granting agencies. Since this work was done in part through the support of a grant from the Veterans Affairs, the original data are only available upon request. Interested parties can gain access to the data supporting conclusions of this study by contacting the senior author. Please email Dr. Mark Yorek at [email protected] or [email protected].
Disclosure
Dr Mark A Yorek reports grants from NIH and Department of Veterans Affairs, during the conduct of the study. The authors declare that there are no conflicts of interest to disclose.
References
1. Malik RA. Diabetic neuropathy: A focus on small fibres. Diab Metab Res Rev. 2019;e3255. doi:10.1002/dmrr.3255
2. Ziegler D, Papanas N, Zhivov A, et al. Early detection of nerve fiber loss by corneal confocal microscopy and skin biopsy in recently diagnosed type 2 diabetes. Diabetes. 2014;63(7):2454–2463. doi:10.2337/db13-1819
3. Hossain P, Sachdev A, Malik RA. Early detection of diabetic peripheral neuropathy with corneal confocal microscopy. Lancet. 2005;366(9494):1340–1343. doi:10.1016/S0140-6736(05)67546-0
4. Yang AY, Chow J, Liu J. Corneal innervation and sensation: the eye and beyond. Yale J Biol Med. 2018;91(1):13–21.
5. Cruzat A, Qazi Y, Hamrah P. In vivo confocal microscopy of corneal nerves in health and disease. Ocul Surf. 2017;15(1):15–47. doi:10.1016/j.jtos.2016.09.004
6. Wang EF, Misra SL, Patel DV. In vivo confocal microscopy of the human cornea in the assessment of peripheral neuropathy and systemic diseases. Biomed Res Int. 2015;2015:951081. doi:10.1155/2015/951081
7. Petropoulos IN, Ponirakis G, Khan A, et al. Corneal confocal microscopy: ready for prime time. Clin Exp Optom. 2019. doi:10.1111/CXO.12887
8. Golebiowski B, Papas E, Stapleton F. Assessing the sensory function of the ocular surface: implications of use of a non-contact air jet aesthesiometer versus the Cochet-Bonnet aesthesiometer. Exp Eye Res. 2011;92(5):408–413. doi:10.1016/j.exer.2011.02.016
9. Feng Y, Simpson TL. Nociceptive sensation and sensitivity evoked from human cornea and conjunctiva stimulated by CO2. Invest Ophthalmol Vis Sci. 2003;44(2):529–532. doi:10.1167/iovs.02-0003
10. Yorek MS, Davidson EP, Poolman P, et al. Corneal sensitivity to hyperosmolar eye drops: A novel behavioral assay to assess diabetic peripheral neuropathy. Invest Ophthalmol Vis Sci. 2016;57(6):2412–2419. doi:10.1167/iovs.16-19435
11. Davidson EP, Coppey LJ, Shevalye H, Obrosov A, Kardon RH, Yorek MA. Impaired corneal sensation and nerve loss in a type 2 rat model of chronic diabetes is reversible with combination therapy of menhaden oil, α-lipoic acid, and enalapril. Cornea. 2017;36(6):725–731. doi:10.1097/ICO.0000000000001182
12. Bitirgen G, Tinkir Kayitmazbatir E, Satirtav G, Malik RA, Ozkagnici A. In vivo confocal microscopic evaluation of corneal nerve fibers and dendritic cells in patients with Behcet’s disease. Front Neurol. 2018;9:204. doi:10.3389/fneur.2018.00204
13. Khan A, Petropoulos IN, Ponirakis G, et al. Corneal confocal microscopy detects severe small fiber neuropathy in diabetic patients with Charcot neuroarthropathy. J Diabetes Investig. 2018;9(5):1167–1172. doi:10.1111/jdi.12806
14. Kass-Iliyya L, Leung M, Marshall A, et al. The perception of affective touch in Parkinson’s disease and its relation to small fibre neuropathy. Eur J Neurosci. 2017;45(2):232–237. doi:10.1111/ejn.13481
15. Podgorny PJ, Suchowersky O, Romanchuk KG, Teasby TE. Evidence for small fiber neuropathy in early Parkinson’s disease. Parkinsonism Relat Disord. 2016;28:94–99. doi:10.1016/j.parkreldis.2016.04.033
16. Tavakoli M, Marshall A, Banka S, et al. Corneal confocal microscopy detects small-fiber neuropathy in Charcot-Marie-Tooth disease type 1A patients. Muscle Nerve. 2012;46(5):698–704. doi:10.1022/mus.23377
17. Papanas N, Ziegler D. Corneal confocal microscopy: A new technique for early detection of diabetic neuropathy. Curr Diab Rep. 2013;13(4):488–499. doi:10.1007/s11892-013-0390
18. Tavakoli M, Quattrini C, Abbott C, et al. Corneal confocal microscopy: A novel noninvasive test to diagnose and stratify the severity of human diabetic neuropathy. Diab Care. 2010;33(8):1792–1797. doi:10.2337/dc.10-0253
19. Jiang MS, Yuan Y, Gu ZX, et al. Corneal confocal microscopy for assessment of diabetic peripheral neuropathy: a meta-analysis. Br J Ophthalmol. 2016;100(1):9–14. doi:10.1136/bjophthalmol-2014-306038
20. Asghar O, Petropoulos IN, Alam U, et al. Corneal confocal microscopy detects neuropathy in subjects with impaired glucose tolerance. Diab Care. 2014;37(9):2643–2646. doi:10.2337/dc14-0279
21. Azmi S, Ferdousi M, Petropoulos IN, et al. Corneal confocal microscopy identifies small-fiber neuropathy in subjects with impaired glucose tolerance who develop type 2 diabetes. Diab Care. 2015;38(8):1502–1508. doi:10.2337/dc14-2733
22. Papanas N, Vinik AI, Ziegler D. Neuropathy in prediabetes: does the clock start ticking early? Nat Rev Endocrinol. 2011;7(11):682–690. doi:10.1038/nrendo.2011.113
23. Davidson EP, Coppey LJ, Kardon RH, Yorek MA. Differences and similarities in development of corneal nerve damage and peripheral neuropathy and in diet-induced obesity and type 2 diabetic rats. Invest Ophthalmol Vis Sci. 2014;55(3):1222–1230. doi:10.1167/ivos.13-13794
24. Beiswenger KK. Calcutt NA and Mizisin AP. Epidermal nerve fiber quantification in the assessment of diabetic neuropathy. Acta Histochem. 2008;110(5):351–362. doi:10.1016/j.acthis.2007.12.004
25. Divisova S, Vickova E, Srotova I, et al. Intraepidermal nerve-fibre density as a biomarker of the course of neuropathy in patients with type 2 diabetes mellitus. Diabet Med. 2016;33(5):650–654. doi:10.1111/dme.12890
26. Davidson EP, Coppey LJ, Holmes A, et al. Effect of inhibition of angiotensin converting enzyme and/or neutral endopeptidase on vascular and neural complications in high fat fed/low dose streptozotocin-diabetic rats. Eur J Pharmacol. 2012;677(1–3):180–187. doi:10.1016/j.ejphar.2011.12.003
27. Davidson EP, Coppey LJ, Yorek MA. Early loss of innervation of cornea epithelium in streptozotocin-induced type 1 diabetic rats: improvement with ilepatril treatment. Invest Ophthalmol Vis Sci. 2012;53(13):8067–8074. doi:10.1167/iovs.12-10826
28. Vagenas D, Pritchard N, Edwards K, et al. Optimal image sample size for corneal nerve morphometry. Optom Vis Sci. 2012;89(5):812–817. doi:10.1097/OPX.0b013e31824ee8c9
29. Coppey LJ, Davidson EP, Dunlap JA, Lund DD, Yorek MA. Slowing of motor nerve conduction velocity in streptozotocin-induced diabetic rats is preceded by impaired vasodilation in arterioles that overlie the sciatic nerve. Int J Exp Diab Res. 2000;1(2):131–143. doi:10.1155/EDR.2000.131
30. Coppey LJ, Gellett JS, Davidson EP, Dunlap J, Lund DD, Yorek MA. Effect of antioxidant treatment of streptozotocin-induced diabetic rats on endoneurial blood flow, motor nerve conduction velocity, and vascular reactivity of epineurial arterioles of the sciatic nerve. Diabetes. 2001;50(8):1927–1937. doi:10.2337/diabetes.50.8.1927
31. Coppey LJ, Davidson EP, Rinehart TW, et al. ACE inhibitor or angiotensin II receptor antagonist attenuates diabetic neuropathy in streptozotocin-induced diabetic rats. Diabetes. 2006;55(2):341–348. doi:10.2337/diabetes.55.02.06.db05-0885
32. Yorek MA, Bohnker RR, Dudley DT, Spector AA. Comparative utilization of n-3 polyunsaturated fatty acids by cultured human Y-79 retinoblastoma cells. Biochim Biophys Acta. 1984;795(2):277–285. doi:10.1016/0005-2760(84)90076-6
33. Mihara M, Uchiyama M, Fukuzawa K. Thiobarbituric acid value on fresh homogenate of rat as a parameter of lipid peroxidation in aging, CCl4 intoxication, and vitamin E deficiency. Biochem Med. 1980;23(3):302–311. doi:10.1016/0006-2944(80)90040-X
34. Coppey LJ, Holmes A, Davidson EP, Yorek MA. Partial replacement with menhaden oil improves peripheral neuropathy in high fat fed low dose streptozotocin type 2 diabetic rat. J Nutr Metab. 2012;2012:950517. doi:10.1155/2012/950517
35. Coppey LJ, Davidson EP, Obrosov A, Yorek MA. Enriching the diet with menhaden oil improves peripheral neuropathy in streptozotocin-induced type 1 diabetic rats. J Neurophysiol. 2015;113(3):701–708. doi:10.1152/jn.00718.2014
36. Yorek MA, Coppey LJ, Gellett JS, et al. Sensory nerve innervation of epineurial arterioles of the sciatic nerve containing calcitonin gene-related peptide: effect of streptozotocin-induced diabetes. Exp Diab Res. 2004;5(3):187–193. doi:10.1080/15438600490486732
37. Coppey L, Davidson E, Shevalye H, Torres ME, Yorek MA. Effect of dietary oils on peripheral neuropathy-related endpoints in dietary obese rats. Diab Metab Syndr Obes. 2018;11:117–127. doi:10.2147/DMSO.s159071
38. Coppey L, Davidson E, Shevalye H, Torres ME, Yorek MA. Effect of early and late interventions with dietary oils on vascular and neural complications in a type 2 diabetic rat model. J Diab Res. 2019;2019:5020465. doi:10.1155/2019/5020465
39. Tavakoli M, Mitu-Pretorian M, Petropoulos IN, et al. Corneal confocal microscopy detects early nerve regeneration in diabetic neuropathy after simultaneous pancreas and kidney transplantation. Diabetes. 2013;62(1):254–260. doi:10.2337/db12-0574
40. MacIver MB, Tanelian DL. Structural and functional specialization of Adelta and C fiber free nerve endings innervating rabbit corneal epithelium. J Neurosci. 1993;13(10):4511–4524. doi:10.1523/JNEUROSCI.13-10-04511.1993
41. Murphy PJ, Patel S, Kong N, et al. Noninvasive assessment of corneal sensitivity in young and elderly diabetic and nondiabetic subjects. Invest Ophthalmol Vis Sci. 2004;45(6):1737–1742. doi:10.1167/iovs.03-0689
42. Smith AG, Howard JR, Kroll R, et al. The reliability of skin biopsy with measurement of intraepidermal nerve fiber density. J Neurol Sci. 2005;228(1):65–69. doi:10.1016/j.jns.2004.09.032
43. Davidson EP, Coppey LJ, Calcutt NA, Oltman CL, Yorek MA. Diet-induced obesity in Sprague-Dawley rats causes microvascular and neural dysfunction. Diab Metab Res Rev. 2010;26(4):306–318. doi:10.1002/dmrr.1088
44. Mehra S, Tavakoli M, Kallinikos PA, et al. Corneal confocal microscopy detects early nerve regeneration after pancreas transplantation in patients with type 1 diabetes. Diab Care. 2007;30(10):2608–2612. doi:10.2337/dc07-0870
45. Azmi S, Jeziorska M, Ferdousi M, et al. Early nerve fibre regeneration in individuals with type 1 diabetes after simultaneous pancreas and kidney transplantation. Diabetologia. 2019;62(8):1478–1487. doi:10.1007/s00125-019-4897-y
46. Tavakoli M, Kallinikos P, Iqbal A, et al. Corneal confocal microscopy detects improvement in corneal nerve morphology with an improvement in risk factors for diabetic neuropathy. Diab Med. 2011;28(10):1261–1267. doi:10.1111/j.1464-5491.2011
47. Jia X, Wang X, Wang X, et al. In vivo corneal confocal microscopy detects improvement of corneal nerve parameters following glycemic control in patients with type 2 diabetes. J Diab Res. 2018;2018:8516276. doi:10.1155/2018/8516276
48. Pop-Busui R, Boulton AJ, Feldman EL, et al. Diabetic neuropathy: A position statement by the American Diabetes Association. Diab Care. 2017;40(1):136–154. doi:10.2337/dc16-2042
49. Ang L, Jaiswal M, Martin C, Pop-Busui R. Glucose control and diabetic neuropathy: lessons from recent large clinical trials. Curr Diab Rep. 2014;14(9):528. doi:10.1007/s11892-014-0528-7
50. Callaghan BC, Hur J, Feldman EL. Diabetic neuropathy: one disease or two? Curr Opin Neurol. 2012;25(5):536–541. doi:10.1097/WCO.ob013e328357a797
51. Shevalye H, Yorek MS, Coppey LJ, et al. Effect of enriching the diet with menhaden oil or daily treatment with resolvin D1 on neuropathy in a mouse model of type 2 diabetes. J Neurophysiol. 2015;114(1):199–208. doi:10.1152/jn.00224.2015
52. Obrosov A, Coppey LJ, Shevalye H, Yorek MA. Effect of fish oil vs. resolvin D1, E1, methyl esters of resolvins D1 or D2 on diabetic peripheral neuropathy. J Neurol Neurophysiol. 2017;8(6):453. doi:10.4172/2155-9562.10000453
53. Yorek MS, Coppey LJ, Shevalye H, Obrosov A, Kardon BH, Yorek MA. Effect of treatment with salsalate, menhaden oil, combination of salsalate and menhaden oil, or resolvin D1 of C57Bl/6J type 1 diabetic mouse on neuropathic endpoints. J Nutr Metab. 2016;2016:5905891. doi:10.1155/2016/5905891
54. Zhang AC, De Silva MEH, MacIsaac RJ, et al. Omega-3 polyunsaturated fatty acid oral supplements for improving peripheral nerve health: a systematic review and meta-analysis. Nutr Rev. 2020;78(4):323-341.
55. Bowman L, Mafham M, Wallendszus K, the ASCEND Study Collaborative Group. Effects of n-3 fatty acids supplements in diabetes mellitus. N Engl J Med. 2018;379(16):1540–1550. doi:10.1161/CIRCULATIONAHA.119.044165.
56. Bhatt DL, Steg PG, Miller M, et al. Effects of icosapent ethyl on total ischemic events: from REDUCE-IT. J Am Coll Cardiol. 2019;73(22):2791–2802. doi:10.1016/j.jacc.2019.02.032
57. Shaikh SR, Kinnun JJ, Leng X, Williams JA, Wassall SR. How polyunsaturated fatty acids modify molecular organization in membranes: insight from NMR studies of model systems. Biochim Biophys Acta. 2015;1848:211–219. doi:10.1016/j.bbamem.2014.04.020
58. Mozaffarian D, Wu JHY. Omega-3 fatty acids and cardiovascular disease. J Am Coll Cardiol. 2011;58(20):2047–2067. doi:10.1016/j.jacc.2011.06.063
59. Soleimani Z, Hashemdokht F, Bahmani F, Tagizadeh M, Memarzadeh MR, Asemi Z. Clinical and metabolic response to flaxseed oil omega-3 fatty acids supplementation in patients with diabetic foot ulcer: a randomized, double-blind, placebo-controlled trial. J Diab Complications. 2017;31(9):1394–1400. doi:10.1016/j.jdiacomp.2017.06.010
60. Lewis EJH, Perkins BA, Lovblom LE, Bazinet RP, Wolever TMS, Bril V. Effect of omega-3 supplementation on neuropathy in type 1 diabetes. Neurology. 2017;88(24):2294–2301. doi:10.1212/WNL.120s.4033
61. Duran AM, Salto LM, Camara J, et al. Effects of omega-3 polyunsaturated fatty-acid supplementation on neuropathic pain symptoms and sphingosine levels in Mexican-Americans with type 2 diabetes. Diab Metab Syndr Obes. 2019;12:109–120. doi:10.2147/DMSO.S187268
62. Superko HR, Superko SM, Nasir K, Agatston A, Garrett BC. Omega-3 fatty acid blood levels: clinical significance and controversy. Circulation. 2013;128(19):2154–2161. doi:10.1161/CIRCULATION113.002731
63. Laidlaw M, Holub BJ. Effects of supplementation with fish oil-derived n-3 fatty acids and gamma-linolenic acid on circulating plasma lipids and fatty acid profiles in women. Am J Clin Nutr. 2003;77(1):37–42. doi:10.1093/ajcn/77.1.37
64. Deinema LA, Vingrys AJ, Wong CY, et al. Double-masked, placebo-controlled clinical trial of two forms of omega-3 supplements for treating dry eye disease. Ophthalmology. 2017;124(1):43–52. doi:10.1016/j.ophtha.2016.09.023
65. He J, Pham TL, Kakazu A, Bazan HEP. Recovery of corneal sensitivity and increase in nerve density and wound healing in diabetic mice after PEDF plus DHA treatment. Diabetes. 2017;66(9):2511–2520. doi:10.2337/db17-0249
66. Zhang Z, Hu X, Di G, et al. Resolvin D1 promotes corneal epithelial wound healing and restoration of mechanical sensation in diabetic mice. Mol Vis. 2018;24:274–285.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.