Back to Journals » Journal of Hepatocellular Carcinoma » Volume 12
Progress in the Study of Intratumoral Microorganisms in Hepatocellular Carcinoma
Authors Song Y, Tian S, Li Z, Miao J ![]() , Wu M
, Wu M ![]() , Xu T, Wu X
, Xu T, Wu X ![]() , Qiao J
, Qiao J ![]() , Zhang X, Zhao H, Kang L
, Zhang X, Zhao H, Kang L ![]() , Cao L, Zhu P, Miao M
, Cao L, Zhu P, Miao M
Received 19 September 2024
Accepted for publication 1 January 2025
Published 18 January 2025 Volume 2025:12 Pages 59—76
DOI https://doi.org/10.2147/JHC.S496964
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Mohamed Shaker
Yagang Song,1,2 Shuo Tian,1,2 Zhanzhan Li,1,2 Jinxin Miao,1,2 Mingming Wu,1,2 Tingli Xu,1,2 Xiangxiang Wu,1,2 Jingyi Qiao,1,2 Xialei Zhang,1,3 Hui Zhao,1,2 Le Kang,1,3 Lihua Cao,1,2 Pingsheng Zhu,4 Mingsan Miao1
1Academy of Chinese Medical Sciences, Henan University of Chinese Medicine, Zhengzhou, Henan, 450046, People’s Republic of China; 2Collaborative Innovation Center of Research and Development on the Whole Industry Chain of Yu-Yao, Henan University of Chinese Medicine, Zhengzhou, Henan, 450046, People’s Republic of China; 3School of Pharmacy, Henan University of Chinese Medicine, Zhengzhou, Henan, 450046, People’s Republic of China; 4College of Chinese Medicine, Henan University of Chinese Medicine, Zhengzhou, Henan, 450046, People’s Republic of China
Correspondence: Mingsan Miao, Academy of Chinese Medical Sciences, Henan University of Chinese Medicine, Zhengzhou, Henan, 450046, People’s Republic of China, Tel/Fax +86-371-65962546, Email [email protected]
Abstract: The intratumoral microbiota, an integral part of liver tumors, has garnered significant attention from researchers due to its role in tumor development regulation and impact on cancer treatment. Intratumoral microorganism not only influences tumorigenesis and progression, but also serves as potential biomarkers and targets for tumor therapy. Targeted manipulation of these microorganisms holds great promise for personalized liver cancer treatment. However, there is a lack of systematic summaries and reports on the study of intratumoral microorganism in hepatocellular carcinoma. This comprehensive review aims to address this gap by summarizing research progress related to in the field of hepatocellular carcinoma intratumoral bacteria, including their sources, types, distribution characteristics within tumors, impact on tumor development, underlying mechanisms, and application prospects. Through the analysis, it is proposed that intratumor organisms can be used as markers for liver cancer diagnosis and treatment, drug carrier materials for targeting liver cancer tissues, and the research prospects of developing new combination therapies based on the in-depth understanding of the interactions between intratumor microorganisms and the tumor microenvironment, immune cells, liver cancer cells, etc. as well as exploring the prospects of developing new combination therapies based on these interactions. It is hoped that from the perspective of intratumoral microbiota, potential theoretical support can be provided for future research on targeted cancer therapy for liver cancer intratumoral microbiota, and new insights and ideas can be provided for targeting points and research methods in tumor research.
Keywords: liver cancer, intratumoral microorganisms, species and distribution, role and mechanism, development prospect
Introduction
Microorganisms are closely associated with the human body, and it has been reported that there are approximately 3 trillion bacteria in the human body, which are involved in the regulation of pathological and physiological processes of the human body.1 Human microorganisms mainly include eukaryotes, prokaryotes and viruses, and the number of genes in these microorganisms is more than 150 times that of the human genome.2 Microorganisms mainly inhabit the mucosal tissues of the human body (including the gastrointestinal tract, oral cavity, skin, etc), which are closely related to human health and disease.3 Research has confirmed that gut microbiota is involved in the occurrence and progression of various diseases such as diabetes, Alzheimer’s disease, and tumors, and is related to clinical efficacy.4 Recent studies have confirmed the presence of microorganisms in tumor tissues as well, and their possible involvement in cancer development.5 In 2020, Nejman et al compared the relationship between seven types of cancer (breast cancer, lung cancer, ovarian cancer, pancreatic cancer, melanoma, bone cancer and brain) and intra-tumor bacteria using more than 1500 tumor samples and adjacent normal samples. The results showed that most tumors and their adjacent normal tissues contained different types of bacteria, which were mainly present in cancer cells and immune cells.6 This finding has brought intratumor microbes to the forefront of cancer research, as there is growing evidence that organs and tissues previously thought to be sterile actually harbor diverse microbial populations,7,8 including liver tissue.
Intratumor microbiota play a key role in shaping the local immune response to the tumor microenvironment, thereby influencing tumor progression, and they may modulate the therapeutic efficacy of tumors by either enhancing or suppressing anti-tumor immune responses.9,10 The presence of bacteria in human tumors was first identified in the 19th century,11 but research on the relationship between tumor microbiota and cancer pathogenesis has been hindered by low microbial content within tumors and technical challenges in detection, leading to limited progress in this field.12 Currently, advances in detection technologies provide unprecedented opportunities to study the diversity and functional characterization of the intratumor microbiota. Importantly, recent studies have revealed the interaction of various tumor-associated microbes in cancer progression and the immune system,13,14 but whether the similar effects exists in Hepatocellular Carcinoma (HCC) tumor tissues remains unclear. What can be confirmed is that the presence of microorganisms within the tumor can impact the response to cancer treatment and may also serve as new biomarkers and targets for tumor therapy. Currently, there is a lack of systematic summaries and reports on the study of intratumoral microorganisms in hepatocellular carcinoma. In summary, the present study is intended to discuss the sources of intratumoral microorganisms in hepatocellular carcinoma, the microbial species and distribution characteristics, the regulatory role and mechanism of action on tumor development, the detection means of intratumoral microorganisms, and the focuses of the future research in anticipation of providing theoretical support for the future tumor therapeutic research targeting intratumoral microorganisms in hepatocellular carcinoma from the perspective of intratumoral microorganisms, and providing new insights and thoughts on the targets of tumor research as well as the methods of research.
Sources of Intratumoral Microorganisms in Hepatocellular Carcinoma
Source from the Oral Cavity
Researchers performed Fluorescence in situ hybridization (FISH) assay to verify the distribution of Streptococcus parasanguinis, Streptococcus mitis, Streptococcus salivarius, Delftia acidovorans, Parabacteroides distasonis, and Stenotrophomonas maltophilia. The results showed that the changes in the intratumoral microbiota were consistent with the changes in the intestinal and oral microbiota. Ultimately, they suggested that the different microorganisms in the liver could also originate from the oral cavity.15 However, the entry of oral microbiota into the liver remains an unsolved mystery. According to available reports, oral microorganisms may form the so-called oral-gut axis by transferring to the gut, which in turn enters the liver and colonizes the liver through the hepatic and intestinal blood circulation,16 which shows that microorganisms of oral origin are still in their essence hepatic via the intestines and then reach the liver, and belong to an indirect source of hepatic microorganisms, as shown in pathway 1 of Figure 1.
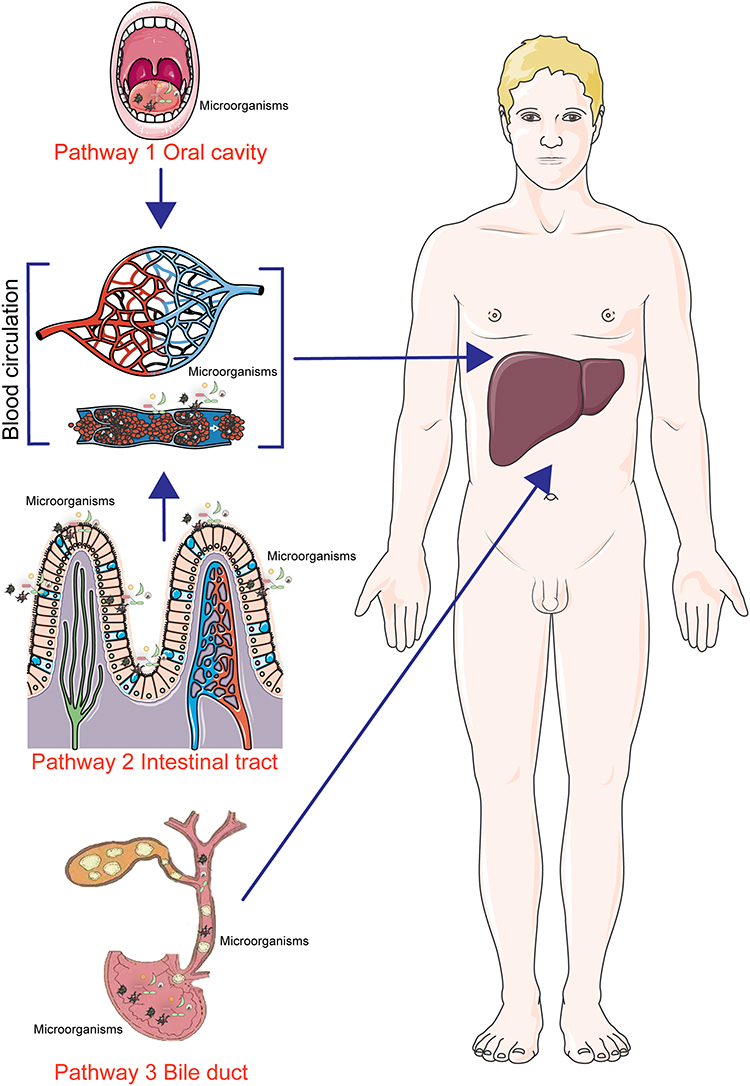 |
Figure 1 Intratumoral microbial sources of hepatocellular carcinoma. |
Source from Intestinal Tract and Blood Circulation
The liver-gut axis is currently considered to be the main source of intratumoral microorganisms in hepatocellular carcinoma. The anatomical relationship between the liver and the gut provides the basis for translocation of gut microbes to the liver. The gut and liver are closely related from embryonic origins and anatomy. Precursors of gut-associated lymphoid tissue originate in the developing liver, and there is mutual influence between the liver and intestines. The concept of gut-liver axis encompasses their major functions. Approximately 70% of the liver’s blood is supplied by the portal vein, which contains large amounts of metabolites from the digestive tract and intestinal microorganisms. When the intestinal barrier function is impaired, intestinal bacteria and their products enter and colonize the liver through the portal vein,17 thus modulating liver-related diseases,18,19 as shown in pathway 2 of Figure 1. In addition, another researcher examined whether bacteria can sustainably colonize the liver by intravenously injecting representative manY/lacE/ypdA and immR mutant strains in SPF mice and tracking their survival in the liver. The results showed that E. gallinarum was not detected in most animals’ livers two weeks later, indicating that hepatic E. gallinarum strains are not able to colonize the liver for a long period of time. Therefore, it is likely that hepatic microorganisms are constantly redistributed in the liver through intestinal translocation.20
Another study confirmed that blood is an important carrier of microorganisms, which was confirmed by Quantitative Polymerase Chain Reaction (qPCR) array analysis of plasma from cirrhotic patients.21 Huang et al performed FISH analysis using bacterial 16S library preparation and sequencing (16S rRNA) probes and found that liver erythrocytes were strongly stained.22 Further evidence was provided by Damgaard et al23 who found viable bacteria on plates inoculated with erythrocytes isolated from healthy individuals. Bacterial growth was observed in 21 out of 60 Red Blood Cells (RBCs) cultured and colonies were characterized by PCR and 16S rRNA gene sequencing. It was found that Streptococcus or Staphylococcus can inhabit the blood cells of healthy individuals.24 Researchers found that the relative abundance of bacteria in the genus differed significantly, with a marked increase in the bacterial abundance of Ruminococcaceae and Bacteroidaceae in the blood and liver tissue of patients with HCC and cirrhosis. The use of 16S rRNA and transcriptome sequencing confirmed a direct link between the abundance of gut bacteria genera in liver tissue and host response.25 These observations suggest that erythrocytes may be an important carrier for live bacteria to enter the liver.
Source from the Biliary Tract
Another reported route of bacterial entry into the liver may be the migration of microbiota in the biliary tract to the liver,26,27 as shown in pathway 3 of Figure 1, but this route is partially controversial and not reliably evidenced as researchers did not observe microbial fluorescence signals from the biliary tract in the FISH assay. Therefore, they concluded that circulating cells (eg, erythrocytes) may be the main carriers of bacterial material into the liver.
In summary, microorganisms in liver cancer tissues may originate from the gut, oral and biliary tracts, but the results may be more convincing when compared with the latter two, where the microorganisms originating from the intestinal tract of liver cancer tissues have been reported more frequently. Additionally, microorganisms in intratumoral tissues may also originate from pathways such as breakage of mucosal tissues such as skin, invasion and metastasis in adjacent tissues, as reported in studies of other tumors,28–30 and it was found that hepatocellular carcinoma and its paracancerous tissues have similar microbial species, but other sources of microorganisms in hepatocellular carcinoma tissues have not yet been confirmed.
Characteristics of Intratumoral Microorganisms and Distribution in Hepatocellular Carcinoma
It is well known that intestinal microorganisms can catalyze a variety of biochemical reactions and produce a variety of active factors, such as Bile Acids (BAs), Short-chain Fatty Acids, Choline, and Lipopolysaccharides (LPSs) that will be absorbed and circulated into the liver, and these metabolites play an important role in the process of hepatocellular carcinoma.31,32 Secondary Bile Acids, Lipoteichoic Acid, and Deoxycholic Acid promote cancer development by down-regulating the anti-tumor activity of the immune system in the liver, and also contribute to the progression of HCC by activating Toll-like receptors.33 In contrast to the gut microbiota, the intratumoral microbiota of hepatocellular carcinoma remains elusive. Although a recent study demonstrated that liver tissues from patients with Nonalcoholic Fatty Liver Disease (NAFLD) are characterized by a wide range of bacterial DNA classifications,34 the presence of viable bacteria in the liver and the type of cellular distribution of intratumoral bacteria remain unknown. In addition, the dysbiosis of the HCC microbiota, colony-specific markers, and the association with the clinicopathologic features of HCC remain to be further addressed. Based on this, the existing studies related to intratumoral microbiota in hepatocellular carcinoma will be summarized and analyzed through the species and distribution characteristics of intratumoral microorganisms and the mechanism of action of intratumoral microorganisms in this paper, the results are shown in Table 1.
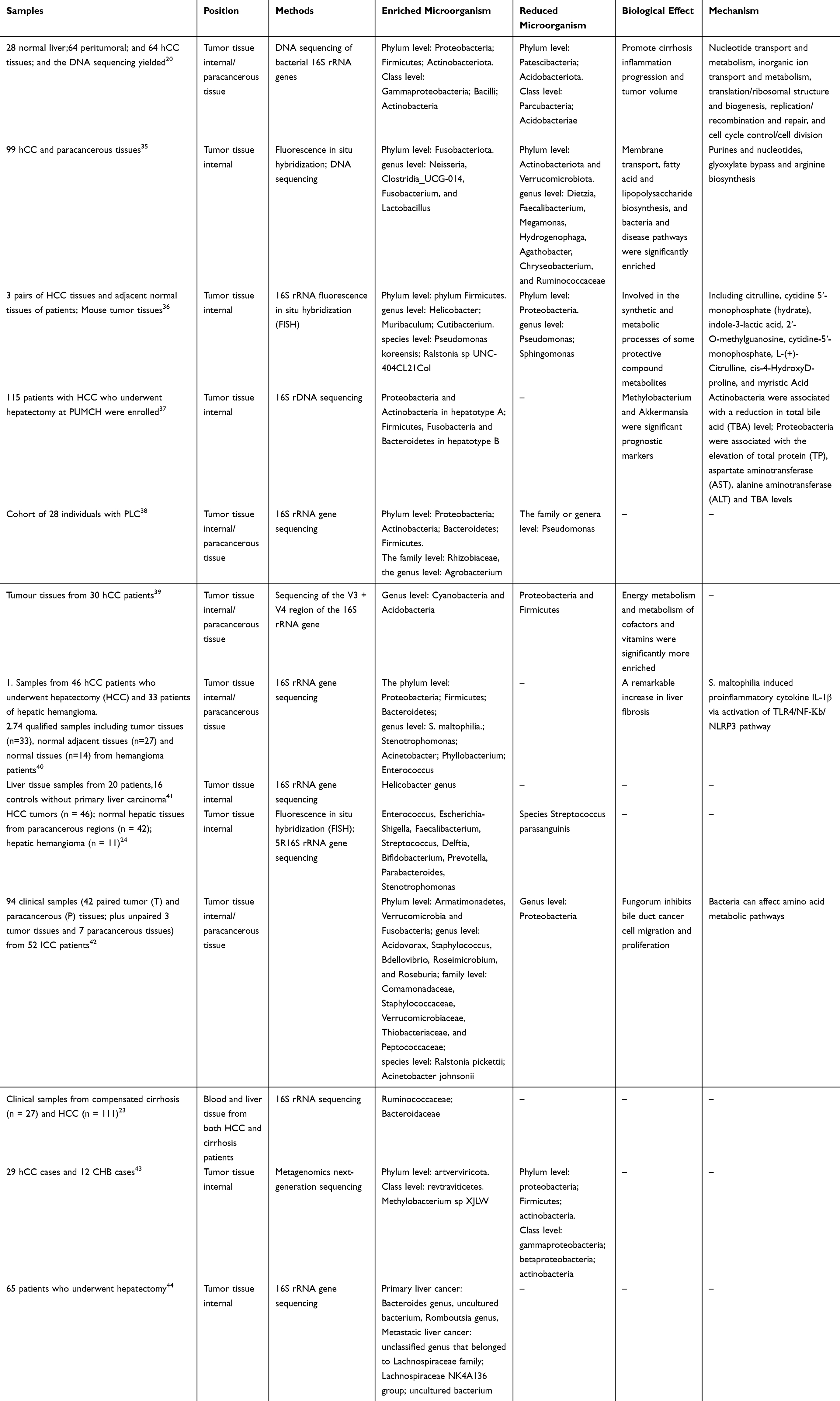 |
Table 1 Characteristics of Intratumoral Microorganisms in Hepatocellular Carcinoma |
Species and Distribution Characteristics of Intratumoral Microorganisms
Studies on intratumoral bacteria in hepatocellular carcinoma are still relatively few compared to other tumors. Huang Jianhang et al20 found that the most predominant phyla in tumor paracancerous tissues and HCC tissues were Patescibacteria, Proteobacteria, Bacillus-like Bacteroidota, Firmicutes, and Actinobacteriota. In their analysis, they concluded that Streptococcaceae and Lactococcus are the hallmark flora of cirrhotic HCC. It was found that the abundance of microorganisms such as Enterobacteriaceae, Clostridium, and Neisseria in HCC tissues was significantly increased, while the abundance of these antitumor bacteria such as Pseudomonas was significantly decreased,35 and processes such as fatty acid and lipid synthesis were significantly enhanced in HCC tissues, which was thought to be a possible key factor for the promotion of hepatocellular carcinoma progression by microorganisms within the tumor.
Xue Chen et al36 found that at the species level Mycoplasma-sp-HU2014, Escherichia coli and Heterobacteria SKP4-8 were the most abundant flora in tumor tissues, and Pseudomonas and Streptococcus pyogenes were common flora in normal tissues. Sun Lejia et al37 utilized 16S rDNA sequencing to investigate alterations in microbial communities within the microenvironment of HCC tumors and adjacent normal tissues. The findings revealed a notable increase in the abundance of Actinobacteria and Firmicutes in tumor tissues, with the researchers suggesting that the rise in Firmicutes may be associated with decreased levels of total bile acid (TBA); Aspergillus was associated with total protein (TP), aspartate aminotransferase (AST), alanine (AST), alanine aminotransferase (ALT) and TBA levels. It was found38 that the abundance of Proteobacteria, Actinobacteria, Bacteroidetes and Firmicutes was significantly increased at the portal level in HCC tumor tissues and its paracancerous tissues. The abundance of Rhizobiaceae was increased at the family level, Agrobacterium was increased at the genus level, and the abundance of Pseudomonas was significantly decreased in the tumor tissues when compared with the paraneoplastic tissues. Sun Pengfei et al39 found that the abundance of Proteobacteria and Firmicutes decreased in hepatocellular carcinoma tumor tissues, while the abundance of Cyanobacteria and Acidobacteria increased; the abundance of Firmicutes, Proteobacteria, and Actinobacteria increased significantly in paraneoplastic tissues, and they believed that tumor growth factors and energy metabolism factors produced by intratumoral microorganisms were the main factors promoting tumor development. Firmicutes, Proteobacteria and Actinobacteria increased significantly in the paracancerous tissues. They concluded that tumor growth factors and energy metabolism factors produced by intratumoral microorganisms are the main factors promoting tumor development. It was found40 that the abundance of Stenotrophomonas, Acinetobacter, Phyllobacterium, Enterococcus were increased at the genus level in hepatocellular carcinoma tissues, and Stenotrophomonas maltophilia was present among the tumor microorganisms in cirrhotic HCC patients. Further studies revealed that Streptococcus maltophilia induced senescence-associated secretory phenotype (SASP) in hematopoietic stem cells through activation of the tlr-4-mediated Nuclear transcription factor-κB (NF-κB) signaling pathway, promoted the formation of the NLR Family, Pyrin Domain Containing Protein 3 (NLRP3) inflammatory vesicle complex and drove the progression of HCC in mice. Slender, curved microorganisms were found in the hepatic sinusoids or hilar region in 6 out of 20 hCC patients by specific H. pylori immunostaining, whereas no bacteria were found in the control group.41 Through FISH and 5R16S rRNA gene sequencing, the researchers confirmed that the abundance of Enterococcus, Escherichia-Shigella, Faecalibacterium, Streptococcus, Delftia, Bifidobacterium, Prevotella, Parabacteroides, Stenotrophomonas in liver tumors and non-tumor tissues increased, and the abundance of species Streptococcus parasanguinis decreased.24 They also found that the intratumor microbial species of hepatocellular carcinoma tissues were highly similar to oral and intestinal microbial species, and they provided a predictive tool for early detection of HCC by establishing a detection method based on oral-intestinal-tumor microbiota. Chai et al42 cultured a Gram-positive aerobic bacterium, identified as Staphylococcus capitis, from fresh cholangiocarcinoma tissue. Meanwhile, using 16S rRNA sequencing, the most abundant bacterial orders in tumor tissues were found to include Burkholderia, Pseudomonas, Xanthomonas, Mycobacterium, and Clostridium. In addition, they found antitumor activity of fungal bacilli from paraneoplastic tissues and concluded that fungal bacilli could inhibit tumor growth by regulating alanine, aspartic acid and glutamic acid metabolism. Patients with HCC and cirrhosis had a higher proportion of bacterial microorganisms in their blood and liver compared to NAFLD. Differential analysis of the relative abundance of bacterial genera revealed an increased abundance of Ruminococcaceae and Bacteroidaceae in the blood and liver tissues of patients with HCC and cirrhosis compared to NAFLD.23 In addition, tumor tissues were found to be enriched in Methylobacterium spp. XJLW, while paraneoplastic tissues were enriched in Klebsiella mutans.43 The abundance of Bacteroides genus, uncultured bacterium, and Romboutsia genus was significantly higher in primary HCC tumor tissues, while in metastatic hepatocellular carcinoma tissues Lachnospiraceae family, Lachnospiraceae NK4A136 and uncultured bacterium were significantly higher in abundance,44 and the researchers suggested that this might be the main difference between primary and metastatic hepatocellular carcinoma.
In summary, our inductive analysis suggests that Streptococcus and Lactococcus may be the signature flora of HCC in liver cirrhosis. Lachnospiraceae NK4A136 may be the main signature flora of metastatic hepatocellular carcinoma. Moreover, the main flora with increased abundance in hepatocellular carcinoma tissues were Enterobacteriaceae, Clostridium, Neisseria, Mycoplasma-sp-HU2014, Escherichia coli, Heterobacteria SKP4-8, Actinobacteria, Firmicutes, Rhizobiaceae, Agrobacterium, Cyanobacteria, Stenotrophomonas, Acinetobacter, Phyllobacterium, Enterococcus, Burkholderia, Pseudomonas, Xanthomonas, Mycobacterium, Clostridium, Methylobacterium spp. XJLW, Bacteroides genus, uncultured bacterium, Romboutsia genus and others. Romboutsia genus, etc., and these bacteria may be associated with the malignant development of hepatocellular carcinoma. The abundance of Streptococcus pyogenes, Pseudomonas, and Proteobacteria was down-regulated in hepatocellular carcinoma tissues, which may be associated with anti-hepatocellular carcinoma effects. Down-regulation of the abundance of Streptococcus pyogenes, Pseudomonas, and Proteobacteria in hepatocellular carcinoma tissues, which may be associated with anti-hepatocellular carcinoma effects.
Role of Intratumor Microorganisms in Hepatocellular Carcinoma
Promotion of Tumorigenesis and Development
In recent years, with the discovery of intratumoral microorganisms, researchers have begun to reveal the impact of intratumoral microbiota on tumors. Researchers found22 that the quantity of fungi correlated with cirrhotic inflammation grading, and Actinobacteriota was significantly more prevalent in female patients and was associated with tumor volume growth. At the level of microbial species, Saccharimonadia was associated with both the cirrhotic inflammation grading and tumor volume growth. These findings suggested that intratumoral microorganisms are closely related to the clinicopathological features of HCC. Recent studies have suggested that helicobacter species may be novel infectious agents associated with HCC.41 Colibactin producing Escherichia coli can synthesize and secrete colibactin, which directly leads to cross-linking of host cells during DNA replication, resulting in cell cycle abnormalities and cancerous transformation, thus inducing tumorigenesis.45
Inhibition of Hepatocellular Carcinoma Progression
However, it has been reported that intratumoral microorganisms do not always promote tumor progression. Recent experiments have shown that Pseudomonas was significantly reduced in tumor tissues at the species level, which researchers believe may be due to the antitumor effect of Pseudomonas, while Rhizobiaceae and Agrobacterium were significantly increased in tumor tissues at the family level.38 The intratumoral bacterium P. fungorum has antitumor activity, and metabolomics and transcriptomics show that P. fungorum inhibits tumor growth through the metabolism of alanine, aspartate, and glutamate. Researchers determined the characteristic profile of intratumoral microbiota and the anti-tumor effect of fungi in ICC.42 Peptides from intratumoral bacteria can be recognized and presented by tumor cells, which in turn activate the immune response, suggesting that intratumoral bacteria may activate the immune system by influencing antigen presentation, thereby affecting the therapeutic efficacy of tumor therapy.46
Intratumor Microbial Changes May Serve as Prognostic Markers for Hepatocellular Carcinoma
Intratumoral microorganisms, as a significant constituent of tumors, have the potential to serve as novel diagnostic or prognostic indicators for tumors. Sun et al. Concluded37 that Proteobacteria and Actinobacteria were more abundant in type A hepatocellular carcinoma tissues, while some phyla, such as Firmicutes, Fusobacteria and Bacteroidetes, were more abundant in type B hepatocellular carcinoma tissues. Firmicutes, Fusobacteria and Bacteroidetes were more abundant in type B hepatocellular carcinoma tissues. Actinobacteria were found to be correlated with reduced levels of total bile acids (TBA), while Proteobacteria showed a correlation with increased levels of total protein (TP), aspartate aminotransferase (AST), alanine aminotransferase (ALT), and Total Bile Acids (TBA). These findings may indicate the pathophysiological condition of the liver, which can serve as an important prognostic indicator for the overall survival rate and recurrence rate of hepatocellular carcinoma. Cani et al study found that high levels of Akkermansia in HCC tissues were associated with a favourable prognosis, suggesting that Akkermansia may also play a beneficial role in the HCC TME.47 In conclusion, the identification of potential intratumoral microbial markers in hepatocellular carcinoma may be one of the main directions for future prognostic studies in hepatocellular carcinoma.
Intratumoral Microorganisms Influence Tumor Therapy
Studies have shown that the metabolic activities of intratumoral microorganisms are one of the main reasons for the development of chemoresistance in tumor tissues.48 Researchers observed a notable increase in the presence of Akkermansia and Ruminococcus in the tumor tissues of HCC patients who exhibited resistance to anti-PD-1 therapy.49 Geller et al reported the potential role of intratumoral bacteria in mediating tumor resistance to the chemotherapeutic agent gemcitabine.7 Another study on Esophageal Squamous Cell Carcinoma (ESCC) found that higher levels of C. nucleatum were associated with negative effects of neoadjuvant chemotherapy, and antibiotic treatments targeting this bacterium improved chemotherapy outcomes in patients.50 On the contrary, intratumoral microbes are not all contributing to chemoresistance, and it has been shown that Akkermansia is a beneficial bacterium that improves anti-tumor immunity and controls tumor growth in vivo more effectively.51 In conclusion, intratumoral microorganisms have different effects on antitumor therapy, and distinguishing the role of different microorganisms in antitumor therapy is important for future research.
The Process Through Which Intratumoral Microbiota Impacts the Progression of Primary Liver Cancer
The impact of intratumoral microbes on tumors has been summarized in previous studies to encompass the regulation of oncogenes through microbial metabolism, promotion of DNA damage and gene mutations, and modulation of immune responses within the microenvironment.52,53 Nejman et al demonstrated that the intratumoral microbiota is tumor-specific, which may imply that the metabolic pathways involved in microorganisms in different environments will have different effects on tumor cells.54 Therefore, in order to further elucidate the mechanism of intratumoral microbial effects on hepatocellular carcinoma, it is necessary to do an in-depth excavation and discussion on the mechanisms of intratumoral microbial effects.
Promotion of DNA Damage and Gene Mutation
It was found that virulence factors released by E. coli, such as cytolethal distending toxin (CDT), worsen the inflammatory response and directly cause DNA damage.55 Inflammation is a recognized risk factor for cancer, and inflammation can lead to DNA damage, which may be a potential mechanism for the involvement of Enterobacteriaceae in the progression of HCC.56 Similarly, Fusobacterium is involved in pro-inflammatory processes,57 and further research has revealed its unique ability to potentially contribute to HCC progression by non-invasively shuttling into the cytoplasm of host cells.58 Researchers established two microbiome-based HCC molecular subtypes, namely bacterial and viral subtypes, and correlated them with different clinicopathological features, which resulted in a higher degree of infiltration of M2 macrophages in the bacterial-dominant subtype, and they also screened for a multiple genetic risk profile encompassing CSAG family, member 4 (CSAG4), Phosphatidylinositol-4,5-bisphosphate 4-phosphatase 2 (PIP4P2) and Translocase of Outer Mitochondrial Membrane 5 homolog (TOMM5).37 In summary, as shown in Figure 2, it is evident that intratumoral microbes may intervene in hepatocellular carcinoma progression through DNA damage and gene mutations, but more in-depth studies are needed to confirm which bacteria intervene in hepatocellular carcinoma progression through these pathways.
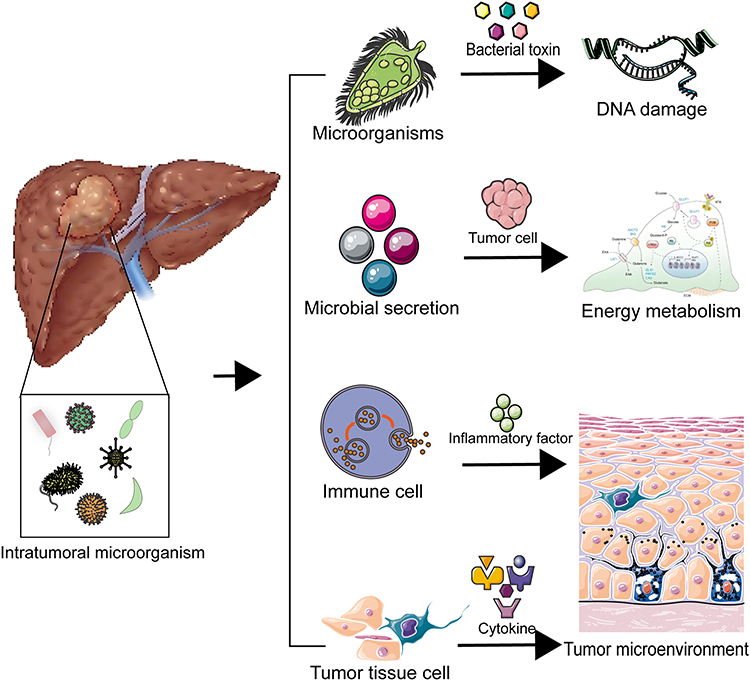 |
Figure 2 Mechanisms of action of intratumoral microorganisms in hepatocellular carcinoma. |
Intratumor Microbes May Regulate Energy Metabolism and Conversion Processes in Cancer Cells Affecting Tumor Progression
A study on microbial changes within tumors found20 that there were three Clusters of Orthologous Groups of proteins (COGs) that were reduced in tumors and adjacent tissues, including energy production and conversion, amino acid transport and metabolism, and signaling mechanisms, and the researchers concluded that the main pathways by which the microbiota affects the tumor cells in HCC include fatty acid and lipid biosynthesis, small molecule metabolism inhibition, and amino acid imbalance. Furthermore, in tumor tissues of hepatocellular carcinoma patients with HBV, the thiamin diphosphate biosynthesis pathway was inhibited, whereas the degradation of glutamate and the biosynthesis of diglycerides were significantly enhanced.35 Allobacillus sp SKP48 and Ralstonia sp UNC404CL21Col bacteria were found to have a significant and positive correlation with the metabolism of most host cells, particularly guanosine, cytidine 5ʹ-monophosphate, indole-3-lactic acid, and 2ʹ-O-methylguanosine. Conversely, there was a significant negative correlation with α-lactose and N-acetyl-D aminoglucose. They suggested that down-regulated microorganisms in HCC tissues correlate with reduced metabolites, and that reduced microorganisms in tumor tissues may play a protective role, and they are also involved in the synthesis and metabolic processes of protective metabolites.36 Acidobacteria, Cyanobacteria and Chloroflexi are the dominant microorganisms in tumor tissues and Kyoto Encyclopedia of Genes and Genomes (KEGG) predictions identified carbohydrate metabolism, energy metabolism and vitamin metabolism as well as membrane translocation as the main pathways in which they play their roles and suggested that these microorganisms are involved in energy metabolism in tumor tissues.39
Functional enrichment analysis performed by researchers found that microorganisms associated with HCC were involved in fatty acid and lipid biosynthesis and confirmed that lipid metabolism was altered in rapidly proliferating cancer cells.59 Microorganisms in tumor tissues may provide fatty acids and lipids to cancer cells, promoting proliferation and invasion. Various metabolites such as choline, bile acids (BA), short-chain fatty acids (SCFA) and amino acids have been reported to be key signaling molecules affecting liver function.60 Streptococcus and Lactobacillus species are found in higher abundance in HCC, whereas they are reduced in Akkermansia, Prevotella-2, Subdoligranulum, and Faecalibacterium, which are all associated with BAs synthesis, especially with lithocarbonic acid (LCA) and deoxycholic acid (DCA).61,62 In summary, intratumoral microorganisms are closely related to the metabolism of tumor tissues, and they may provide nutrients for the growth of tumor cells through their own secreted metabolites, or they may influence the proliferation of cancer cells by regulating the metabolism of tumor cells through the secretion of growth factors, as shown in Figure 2. It is worth noting that which intratumoral microorganisms promote the metabolism of hepatocellular carcinoma tumor cells and which inhibit the metabolism of tumor cells still need to be further investigated in depth, so as to provide new targets or pathways for the treatment of hepatocellular carcinoma.
Regulation of the Tumor Microenvironment
Metabolites generated by microorganisms within the tumor have a regulatory effect on the tumor microenvironment.63 New studies have shown that intratumoral microbes can induce inflammatory and immune responses to remodel the tumor microenvironment, which in turn affects tumorigenesis and progression.64,65 Certain specific bacteria can break the intestinal vascular barrier to colonize the liver and recruit immune cells such as macrophages, neutrophils and monocytes in the liver, thus creating a suitable microenvironment for tumor metastasis.66 The metabolic interactions between intratumoral microbes and the immune microenvironment plays a significant role in shaping tumor progression.67 Bacterial-mediated bile acid metabolism regulates HCC tumorigenesis by interfering with NKT cells.68 In addition, short-chain fatty acids produced by bacterial metabolism may attenuate liver injury by reducing lipopolysaccharide transport and inhibiting macrophage activation, pro-inflammatory cytokine production and neutrophil infiltration.69 Cancer tissues with intratumoral bacteria were accompanied by more M2 macrophage infiltration and upregulation of multiple metabolic pathways, suggesting that metabolism plays a regulatory role between the intratumoral microbiome and the immune microenvironment, as shown in Figure 2. Researchers have hypothesized that IMH in HCC may affect the immune microenvironment by regulating metabolic reprogramming, which may be the main reason for the development and progression of HBV-associated HCC.37
Intratumoral microbes have been reported to be recognized by immune receptors on hepatocytes, such as Kupffer cells and hepatic stellate cells, which further initiates and sustains the inflammatory cascade response, leading to hepatic fibrosis,70,71 which can ultimately progress from cirrhosis to hepatocellular carcinoma as the injury worsens.72 The abundance of Stenotrophomonas maltophilia was found to be significantly elevated in the tumor microbiota of cirrhotic HCC patients, and Stenotrophomonas maltophilia may expedite the advancement of HCC in mice through the induction of NLRP3 inflammasome complex formation, which promotes hepatic secretion of various inflammatory factors 40. Of interest, Faecalibacterium is involved in butyrate production and acts as a SCFAs-associated bacterium to regulate the onset of inflammatory responses,73 In summary, as shown in Figure 2, intratumoral microbes drive tumor progression by promoting inflammation. In summary, modulation of intratumoral microorganisms may achieve regulation of the tumor microenvironment, providing another direction for antitumor therapies based on the tumor microenvironment.
Means of Detecting Intratumor Microorganisms in Hepatocellular Carcinoma
Currently, the predominant approach employed by researchers for detecting intratumor microorganisms is 16S rRNA sequencing. This method primarily focuses on obtaining taxonomic information about various bacteria through the analysis of the variable region within the bacterial 16S rRNA gene sequence.74 The method mainly consists of PCR amplification, purification and sequencing, and finally clustering the sequences into operational taxonomic units (OTUs) based on similarity and comparing them with existing database entries based on the representative sequences in each OTU. This in turn enables the taxonomic identification and quantitative description of all bacteria in the tumor, but it is difficult to distinguish bacteria with small interspecies differences.75 In addition, the disadvantage of this method is that only bacteria can be identified, and microorganisms other than bacteria cannot be characterized by this method.
Macrogenome sequencing is another major tool for intratumoral microbial detection; the macrogenome is the sum of all microbial genomes in the environment. Unlike 16S rRNA sequencing, macrogenomes are not singularly targeted to a specific microbiome (fungal, bacterial, or viral), but rather, the sum of all microbial genomes is sequenced, which gives it an advantage in the precise identification of species and allows for the speculation of functional properties of the microbiome.76 The shortcomings of this method are that the assembly and comparison of sequences are limited by reference sequence entries in microbial sequence databases, and the vast majority of genetic information extracted from tumor tissues is human DNA, whereas access to data from low-content microorganisms is more limited.77
Secondly, immunohistochemistry (IHC) can also be utilized for the characterization of intratumoral microorganisms. Immunohistochemistry is based on the specific binding of antigen to antibody. This method is more similar to traditional HE staining in that the sample is first fixed and then bound to a blocking solution. The samples are combined with primary and secondary antibodies, then stained, and finally the samples are analyzed by microscopy.78 IHC is a qualitative and quantitative technique that labels specific locations of intratumoral microorganisms and enables localization of intratumoral microorganisms. Researchers usually use lipopolysaccharide (LPS) from Gram-negative bacteria or lipophosphatidic acid from Gram-positive bacteria to label the sample’s flora and demonstrate the presence of intratumoral microorganisms.79
FISH can also be used to detect the presence of bacteria in tissues, which is based on the principle of hybridization of fluorescein-labeled nucleic acid probes with bacterial nucleic acid sequences in the samples to be tested in accordance with the principle of base complementary pairing, which are washed and then directly observed under a fluorescence microscope.80 In addition, FISH combined with transmission electron microscopy imaging can accurately identify bacterial sublocalization in cancer cells or infiltrating immune cells.81
D-alanine is an important component of peptidoglycan for cell wall production by bacteria, whereas mammalian cells do not have cell walls and do not have recognizable D-alanine.82 Therefore, researchers usually use fluorescently labeled D-alanine to label live bacteria in situ, which can be used as the main means of live bacteria detection in tumor tissues.
Prospects of Intratumoral Microorganisms in the Prevention and Treatment of Hepatocellular Carcinoma
For a long time, researchers have been searching for biomarkers for different tumors with a view to achieving early identification and diagnosis of tumors, which offers the possibility of biomarker discovery in hepatocellular carcinoma due to the significant differences between intratumoral microorganisms and normal tissues. In addition, humans have found that intratumoral microorganisms may affect the progression of hepatocellular carcinoma by promoting or inhibiting the growth of tumor cells, making it difficult to meet the clinical needs of anti-tumor therapeutic efficacy by influencing the existing therapeutic measures for tumors. Although the study of intratumor microorganisms is still in the preliminary stage and belongs to the emerging field. However, its broad application prospects have attracted the attention of most researchers. With the continuous progress of molecular biology technology, the mystery of intratumoral microorganisms will be gradually revealed, and the corresponding tumor therapeutic strategies will be designed and applied to cancer treatment. In summary, intratumor microbes have a broad development prospect in the diagnosis and treatment of hepatocellular carcinoma.
Biomarkers as Diagnostic and Therapeutic Agents for Hepatocellular Carcinoma
It is well known that intratumoral microorganisms, as part of the main components of liver cancer, have the potential to become new diagnostic or prognostic markers. Currently, researchers have found that intratumoral microbial results in hepatocellular carcinoma can indicate the pathophysiological state of the liver and the development of the tumor.37 Other studies have also found similar results in different types of tumors, such as the identification of Aspergillus phylum as a significant prognostic marker in pancreatic ductal adenocarcinoma through cancer genome mapping.50 Four bacteria in non-small cell lung cancer samples were effective in predicting 2-year survival of patients.83 It was shown that the intratumoral microbiota can predict the prognosis of papillary thyroid carcinoma in patients of different sexes and subtypes.84 The phylum Aspergillus was significantly enriched in bronchoalveolar lavage fluid from non-small cell lung cancer, and further subdivision of the bacterial community was associated with discrimination between adenocarcinoma and squamous cell carcinoma.85 There is increasing evidence indicating that tumor-associated circulating microbial DNA holds potential as a biomarker in cancer liquid biopsies.86 In conclusion, the identification of potential biomarkers for tumor prognosis and their clinical application are major directions for future research, especially in hepatocellular carcinoma, which is characterized by high recurrence and metastasis rates.
Microbial Vectors That Can Be Used to Target Tumor Tissue
With the advent of synthetic biology, a variety of bacteria have been modified to have the ability to target tumor tissues with low toxicity and high accuracy. These tumor-targeting bacteria exert important antitumor effects through a variety of payload delivery and effector systems, such as production of prodrug converting enzymes, control of cytotoxic drug expression, stimulation of immune responses, and targeting of tumor mesenchyme.87 In mouse experiments, researchers found that the expression of cytolysin A in E. coli or S. typhimurium was regulated by an inducible promoter, which enabled the targeting of tumor tissues.88,89 A Bifidobacterium infantis-mediated prodrug enzyme delivery system for herpes simplex virus type I thymidine kinase/ganciclovir (HSV1-TK/GCV) was found to reduce bladder cancer progression in rats.90 In addition, Staphylococcus epidermidis was found to produce 6-N-hydroxyaminopurine, which could selectively inhibit the proliferation and progression of skin tumors in mice by inhibiting DNA polymerase activity.91 The discovery of specific microorganisms from hepatocellular carcinoma tissues with tumor-targeting properties, detoxification through modification, and use as targeted drug carrier materials are of great significance for achieving targeted therapies for hepatocellular carcinoma.
Improvement of Hepatocellular Carcinoma Treatment
As mentioned in the previous section, different microbiota in hepatocellular carcinoma tissues play multiple roles in the process of tumorigenesis and development. Actinobacteriota was correlated with tumor volume growth, while Lactococcus showed a significant association with cirrhosis and HBcAb status. Additionally, Pseudomonas may exhibit antitumor effects.38 Helicobacter species may be a novel infectious agent associated with HCC.41 The intratumoral bacterium P. fungorum exerts antitumor activity through alanine, aspartate and glutamate metabolism.42 In addition, intratumoral bacteria may activate the immune system by influencing antigen presentation, thereby affecting the therapeutic efficacy of tumors.46 The metabolic activities of intratumoral microorganisms are the main reason for the development of chemoresistance in tumors.48 Potential role of intratumoral bacteria in mediating tumor resistance to the chemotherapeutic agent gemcitabine.7 Clarifying the role of intratumor bacteria and their mechanisms by imposing appropriate interventions for the different roles played by intratumor bacteria in the treatment of hepatocellular carcinoma would be expected to provide therapeutic efficacy in hepatocellular carcinoma.
Conclusion and Prospects
The intratumoral microbiota is one of the main components of liver cancer, has garnered significant attention from researchers due to its role in tumor development regulation and impact on cancer treatment. A variety of tumors are thought to have different microbial communities, and understanding the complex relationship between intratumoral microbes and tumors can provide valuable insights into potential and existing therapeutic strategies for hepatocellular carcinoma. In this study, we comprehensively reviewed the relevant research progress in the field of intratumoral bacteria in hepatocellular carcinoma, and summarized the research results in terms of the sources of intratumoral microorganisms in hepatocellular carcinoma, the species and distributional characteristics of intratumoral microorganisms, the influence of intratumoral microorganisms on the development of hepatocellular carcinoma, as well as the mechanism of their action and the prospect of their application. It is proposed that intratumoral organisms can be used as markers for liver cancer diagnosis and treatment, and can be used as drug carrier materials targeting liver cancer tissues. Through in-depth research on the interactions between intratumoral microorganisms and the tumor microenvironment, immune cells, and hepatocellular carcinoma cells, the roles and mechanisms of different microorganisms in intratumoral tumors will be clarified, and then, in combination with other interventions, it is expected that the therapeutic efficacy of hepatocellular carcinoma will be improved.
In addition, the study of intratumoral microbiota in tumorigenesis and progression is only the beginning. In the future, more rigorous animal models are needed to track tumor cells affected by microbes to provide more experimental evidence for the clinic. A combined multidisciplinary approach is needed to quantify the correlation between microorganisms within tumors and the development and advancement of tumors. By conducting comparative analysis of healthy tissues and tumor tissues, as well as healthy individuals and those with tumors, tumor tissues and paraneoplastic tissues, a comprehensive information base of intratumor microorganisms in hepatocellular carcinoma can be established. This will enable large-scale analysis of intratumor microorganisms in hepatocellular carcinoma, identification of potential microorganisms and risk factors for tumor therapy, and provide a reliable direction for subsequent in-depth research. However, this work is complicated by ethical and accessibility challenges associated with normal human tissues. In the context of antitumor therapy, linking identified microbial profiles to liver cancer response modulation may lead to new ways of intervention.
As the intratumoral microbiota belongs to an emerging field, relatively few clinical samples have been reported, especially the low abundance of intratumoral microorganisms, leading to discrepancies between the results of different experiments. In addition, the corresponding detection means and statistical methods for intratumoral microorganisms need to be further improved. As a result, the field is also subject to a lot of skepticism. However, it is certain that intratumoral microorganisms are present in hepatocellular carcinoma tissues,37,42 and intratumoral microorganisms may have a certain influence on the development of hepatocellular carcinoma.92–94 In conclusion, the field needs to further expand the analysis of results from clinical samples to improve the reliability of the results, and to develop more reliable tests and statistical methods in order to better support the development of the field. It is believed that this field can provide breakthroughs in the treatment of tumors in the future.
Funding
The authors are grateful for the financial support from the National Natural Science Foundation of China (No.82204679), the Joint Fund of Science and Technology R&D Program of Henan Province (Cultivation Category of Advantageous Disciplines) Project (No.232301420081), the Project of Young Talents Support Project of Henan Province (No.2023HYTP037), the National International Cooperation Base of China (No.2016-151), the Key scientific research projects of colleges and universities in Henan Province (No.22A360013), the Major Special Project of Henan Province (221100310400), and the Science and technology project of Henan Province (242102310505).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Liu J, Zhang Y. Intratumor microbiome in cancer progression: current developments, challenges and future trends. Biomark Res. 2022;10(1):37. doi:10.1186/s40364-022-00381-5
2. Gilbert J, Blaser M, Caporaso J, et al. Current understanding of the human microbiome. Nat Med. 2018;24:392–400. doi:10.1038/nm.4517
3. Campbell C, Mckenney P, Konstantinovsky D, et al. Bacterial metabolism of bile acids promotes generation of peripheral regulatory T cells. Nature. 2020;581(7809):475. doi:10.1038/s41586-020-2193-0
4. Davar D, Dzutsev A, Mcculloch J, et al. Fecal microbiota transplant overcomes resistance to anti-PD-1 therapy in melanoma patients. Science. 2021;371(6529):595. doi:10.1126/science.abf3363
5. Helmink B, Khan M, Hermann A, et al. The microbiome, cancer, and cancer therapy. Nat Med. 2019;25(3):377. doi:10.1038/s41591-019-0377-7
6. Nejman D, Livyatan I, Fuks G, et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science. 2020;368:973–980. doi:10.1126/science.aay9189
7. Geller L, Barzily-Rokni M, Danino T, et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science. 2017;357:1156–1160. doi:10.1126/science.aah5043
8. Parhi L, Alon-Maimon T, Sol A, et al. Breast cancer colonization by Fusobacterium nucleatum accelerates tumor growth and metastatic progression. Nat Commun. 2020;11:3259. doi:10.1038/s41467-020-16967-2
9. Siegel R, Miller K, Jemal A. Cancer statistics, 2020. CA Cancer J Clin. 2020;70:7–30. doi:10.3322/caac.21590
10. Cao Y, Xia H, Tan X, et al. Intratumoural microbiota: a new frontier in cancer development and therapy. Signal Transduct Target Ther. 2024;9(1):15. doi:10.1038/s41392-023-01693-0
11. Cogdill A, Gaudreau P, Arora R, et al. The impact of intratumoral and gastrointestinal microbiota on systemic cancer therapy. Trends Immunol. 2018;39(11):900–920. doi:10.1016/j.it.2018.09.007
12. Dart A. Bacterial residents. Nat Rev Cancer. 2020;20(8):413. doi:10.1038/s41568-020-0284-8
13. Gagliani N, Hu B, Huber S, et al. The fire within: microbes inflame tumors. Cell. 2014;157(4):776–783. doi:10.1016/j.cell.2014.03.006
14. Pushalkar S, Hundeyin M, Daley D, et al. The pancreatic cancer microbiome promotes oncogenesis by induction of innate and adaptive immune suppression. Cancer Discov. 2018;8(4):403–416. doi:10.1158/2159-8290.CD-17-1134
15. Yang J, He Q, Lu F, et al. A distinct microbiota signature precedes the clinical diagnosis of hepatocellular carcinoma. Gut Microbes. 2023;15(1):2201159. doi:10.1080/19490976.2023.2201159
16. Elghannam M, Hassanien M, Ameen Y, et al. Oral microbiota and liver diseases. Clin Nutr ESPEN. 2023;54:68–72. doi:10.1016/j.clnesp.2022.12.030
17. Jiang J, Chen X, Ren Z, et al. Gut microbial dysbiosis associates hepatocellular carcinoma via the gut-liver axis. Hepatobiliary Pancreat Dis Int. 2019;18(1):19–27. doi:10.1016/j.hbpd.2018.11.002
18. Bawa M, Saraswat V. Gut-liver axis: role of inflammasomes. J Clin Exp Hepatol. 2013;3:141–149. doi:10.1016/j.jceh.2013.03.225
19. Ponziani F, Bhoori S, Castelli C, et al. Hepatocellular carcinoma is associated with gut microbiota profile and inflammation in nonalcoholic fatty liver disease. Hepatology. 2019;69:107–120. doi:10.1002/hep.30036
20. Yang Y, Nguyen M, Khetrapal V, et al. Within-host evolution of a gut pathobiont facilitates liver translocation. Nature. 2022;607(7919):563–570. doi:10.1038/s41586-022-04949-x
21. Traykova D, Schneider B, Chojkier M, et al. Blood microbiome quantity and the hyperdynamic circulation in decompensated cirrhotic patients. PLoS One. 2017;12:e0169310. doi:10.1371/journal.pone.0169310
22. Huang J, Wang J, Chai X, et al. The intratumoral bacterial metataxonomic signature of hepatocellular carcinoma. Microbiol Spectr. 2022;10(5):e0098322. doi:10.1128/spectrum.00983-22
23. Damgaard C, Magnussen K, Enevold C, et al. Viable bacteria associated with red blood cells and plasma in freshly drawn blood donations. PLoS One. 2015;10:e0120826. doi:10.1371/journal.pone.0120826
24. Yamaguchi H, Yamada M, Uruma T, et al. Prevalence of viable Chlamydia pneumoniae in peripheral blood mononuclear cells of healthy blood donors. Transfusion. 2004;44:1072–1078. doi:10.1111/j.1537-2995.2004.04005.x
25. Effenberger M, Waschina S, Bronowski C, et al. A gut bacterial signature in blood and liver tissue characterizes cirrhosis and hepatocellular carcinoma. Hepatol Commun. 2023;7(7):e00182. doi:10.1097/HC9.0000000000000182
26. Molinero N, Ruiz L, Milani C, et al. The human gallbladder microbiome is related to the physiological state and the biliary metabolic profile. Microbiome. 2019;7:100. doi:10.1186/s40168-019-0712-8
27. Kose S, Grice K, Orsi W, et al. Metagenomics of pigmented and cholesterol gallstones: the putative role of bacteria. Sci Rep. 2018;8:11218. doi:10.1038/s41598-018-29571-8
28. Leinwand J, Miller G. Regulation and modulation of antitumor immunity in pancreatic cancer. Nat Immunol. 2020;21:1152–1159.
29. Desai S, Dharavath B, Manavalan S, et al. Fusobacterium nucleatum is associated with inflammation and poor survival in early-stage HPV-negative tongue cancer. NAR Cancer. 2022;4(1):zcac006. doi:10.1093/narcan/zcac006
30. Qiao H, Tan X, Li H, et al. Association of intratumoral microbiota with prognosis in patients with nasopharyngeal carcinoma from 2 hospitals in China. JAMA Oncol. 2022;8(9):1301–1309.
31. Jia B, Jeon C. Promotion and induction of liver cancer by gut microbiome-mediated modulation of bile acids. PLoS Pathog. 2019;15(9):e1007954. doi:10.1371/journal.ppat.1007954
32. Sinha R, Dey A, Agarwal S. Initial presentation of hepatocellular carcinoma as soft tissue swelling of the forearm: a rare cytomorphological diagnosis. J Cancer Res Ther. 2015;11:1032. doi:10.4103/0973-1482.154052
33. Dapito D, Mencin A, Gwak G, et al. Promotion of hepatocellular carcinoma by the intestinal microbiota and TLR4. Cancer Cell. 2012;21:504–516. doi:10.1016/j.ccr.2012.02.007
34. Sookoian S, Salatino A, Castano GO, et al. Intrahepatic bacterial metataxonomic signature in non-alcoholic fatty liver disease. Gut. 2020;69:1483–1491. doi:10.1136/gutjnl-2019-318811
35. He Y, Zhang Q, Yu X, et al. Overview of microbial profiles in human hepatocellular carcinoma and adjacent nontumor tissues. J Transl Med. 2023;21(1):68. doi:10.1186/s12967-023-03938-6
36. Xue C, Gu X, Shi Q, et al. The interaction between intratumoral bacteria and metabolic distortion in hepatocellular carcinoma. J Transl Med. 2024;22(1):237. doi:10.1186/s12967-024-05036-7
37. Sun L, Ke X, Guan A, et al. Intratumoural microbiome can predict the prognosis of hepatocellular carcinoma after surgery. Clin Transl Med. 2023;13(7):e1331. doi:10.1002/ctm2.1331
38. Qu D, Wang Y, Xia Q, et al. Intratumoral microbiome of human primary liver cancer. Hepatol Commun. 2022;6(7):1741–1752. doi:10.1002/hep4.1908
39. Sun P, Li Z, Zhang B. Characterization of disease-associated microbiota in hepatocellular carcinoma. J Cancer Res Ther. 2023;19(4):881–891. doi:10.4103/jcrt.jcrt_139_22
40. Liu B, Zhou Z, Jin Y, et al. Hepatic stellate cell activation and senescence induced by intrahepatic microbiota disturbances drive progression of liver cirrhosis toward hepatocellular carcinoma. J Immunother Cancer. 2022;10(1):e003069. doi:10.1136/jitc-2021-003069
41. Yao H, Ma S, Huang J, et al. Trojan-horse strategy targeting the gut-liver axis modulates gut microbiome and reshapes microenvironment for orthotopic hepatocellular carcinoma therapy. Adv Sci. 2024;11(44):e2310002. doi:10.1002/advs.202310002
42. Chai X, Wang J, Li H, et al. Intratumor microbiome features reveal antitumor potentials of intrahepatic cholangiocarcinoma. Gut Microbes. 2023;15(1):2156255. doi:10.1080/19490976.2022.2156255
43. Li S, Xia H, Wang Z, et al. Intratumoral microbial heterogeneity affected tumor immune microenvironment and determined clinical outcome of HBV-related HCC. Hepatology. 2023;78(4):1079–1091. doi:10.1097/HEP.0000000000000427
44. Seiga K, Takahiro Y, Nobuyuki T, et al. Profiling of tumour-associated microbiota in human hepatocellular carcinoma. Sci Rep. 2021;11:10589. doi:10.1038/s41598-021-89963-1
45. Pleguezuelos-Manzano C, Puschhof J, Rosendahl H, et al. Mutational signature in colorectal cancer caused by genotoxic pks+ E. Nature. 2020;580(7802):269–273. doi:10.1038/s41586-020-2080-8
46. Kalaora S, Nagler A, Nejman D, et al. Identification of bacteria-derived HLA-bound peptides in melanoma. Nature. 2021;592(7852):138–143. doi:10.1038/s41586-021-03368-8
47. Cani P, Depommier C, Derrien M, et al. Akkermansia muciniphila: paradigm for next‐generation beneficial microorganisms. Nat Rev Gastroenterol Hepatol. 2022;19(10):625–637. doi:10.1038/s41575-022-00631-9
48. Geller L, Straussman R. Intratumoral bacteria may elicit chemoresistance by metabolizing anticancer agents. Mol Cell Oncol. 2018;5(1):e1405139. doi:10.1080/23723556.2017.1405139
49. Zheng Y, Wang T, Tu X, et al. Gut microbiome affects the response to anti-PD-1 immunotherapy in patients with hepatocellular carcinoma. J ImmunoTher Cancer. 2019;7:193. doi:10.1186/s40425-019-0650-9
50. Yamamura K, Izumi D, Kandimalla R, et al. Intratumoral Fusobacterium nucleatum levels predict therapeutic response to neoadjuvant chemotherapy in esophageal squamous cell carcinoma. Clin Cancer Res. 2019;25:6170–6179. doi:10.1158/1078-0432.CCR-19-0318
51. Li W, Deng Y, Chu Q, et al. Gut microbiome and cancer immunotherapy. Cancer Lett. 2019;447:41–47. doi:10.1016/j.canlet.2019.01.015
52. Garrett W. Cancer and the microbiota. Science. 2015;348:80–86. doi:10.1126/science.aaa4972
53. Ramirez-Labrada A, Isla D, Artal A, et al. The influence of lung microbiota on lung carcinogenesis, immunity, and immunotherapy. Trends Cancer. 2020;6:86–97. doi:10.1016/j.trecan.2019.12.007
54. Dzutsev A, Badger J, Perez-Chanona E, et al. Microbes and cancer. Annu Rev Immunol. 2017;35:199–228. doi:10.1146/annurev-immunol-051116-052133
55. Nesic D, Hsu Y, Stebbins C. Assembly and function of a bacterial genotoxin. Nature. 2004;429:429–433. doi:10.1038/nature02532
56. Elinav E, Nowarski R, Thaiss CA, et al. Inflammation-induced cancer: crosstalk between tumours, immune cells and microorganisms. Nat Rev Cancer. 2013;13:759–771. doi:10.1038/nrc3611
57. Dharmani P, Strauss J, Ambrose C, et al. Fusobacterium nucleatum infection of colonic cells stimulates MUC2 mucin and tumor necrosis factor alpha. Infect Immun. 2011;79:2597–2607. doi:10.1128/IAI.05118-11
58. Xu M, Yamada M, Li M, et al. FadA from Fusobacterium nucleatum utilizes both secreted and nonsecreted forms for functional oligomerization for attachment and invasion of host cells. J Biol Chem. 2007;282:25000–25009. doi:10.1074/jbc.M611567200
59. Gedgaudas R, Bajaj J, Skieceviciene J, et al. Circulating microbiome in patients with portal hypertension. Gut Microbes. 2022;14:2029674. doi:10.1080/19490976.2022.2029674
60. Di Ciaula A, Baj J, Garruti G, et al. Liver steatosis, gut-liver axis, microbiome and environmental factors. A never-ending bidirectional cross-talk. J Clin Med. 2020;9(8):2648. doi:10.3390/jcm9082648
61. Yang N, Xu J, Wang X, et al. The spatial landscape of the bacterial community and bile acids in the digestive tract of patients with bile reflux. Front Microbiol. 2022;13:835310. doi:10.3389/fmicb.2022.835310
62. Yan H, Wei W, Hu L, et al. Reduced feeding frequency improves feed efficiency associated with altered fecal microbiota and bile acid composition in pigs. Front Microbiol. 2021;12:761210. doi:10.3389/fmicb.2021.761210
63. Jiang S, Xie Y, Xiao X, et al. Fusobacterium nucleatum-derived succinic acid induces tumor resistance to immunotherapy in colorectal cancer. Cell Host Microbe. 2023;31(5):781–797.e9. doi:10.1016/j.chom.2023.04.010
64. Baruch E, Wang J, Wargo J. Gut microbiota and antitumor immunity: potential mechanisms for clinical effect. Cancer Immunol Res. 2021;9:365–370. doi:10.1158/2326-6066.CIR-20-0877
65. Sepich-Poore G, Zitvogel L, Straussman R, et al. The microbiome and human cancer. Science. 2021;371:6536. doi:10.1126/science.abc4552
66. Bertocchi A, Carloni S, Ravenda P, et al. Gut vascular barrier impairment leads to intestinal bacteria dissemination and colorectal cancer metastasis to liver. Cancer Cell. 2021;39:708–24e11. doi:10.1016/j.ccell.2021.03.004
67. Bidkhori G, Benfeitas R, Klevstig M, et al. Metabolic network-based stratification of hepatocellular carcinoma reveals three distinct tumor subtypes. Proc Natl Acad Sci U S A. 2018;115:e11874–83. doi:10.1073/pnas.1807305115
68. Ma C, Han M, Heinrich B, et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science. 2018;360:eaan5931. doi:10.1126/science.aan5931
69. Wang H, Wang P, Wang X, et al. Butyrate enhances intestinal epithelial barrier function via up-regulation of tight junction protein Claudin-1 transcription. Dig Dis Sci. 2012;57:3126–3135. doi:10.1007/s10620-012-2259-4
70. Csak T, Ganz M, Pespisa J, et al. Fatty acid and endotoxin activate inflammasomes in mouse hepatocytes that release danger signals to stimulate immune cells. Hepatology. 2011;54(1):133–144. doi:10.1002/hep.24341
71. Anand G, Zarrinpar A, Loomba R. Targeting dysbiosis for the treatment of liver disease. Semin Liver Dis. 2016;36:37–47. doi:10.1055/s-0035-1571276
72. Seki E, Schnabl B. Role of innate immunity and the microbiota in liver fibrosis: crosstalk between the liver and gut. J Physiol. 2012;590(3):447–458. doi:10.1113/jphysiol.2011.219691
73. Louis P, Young P, Holtrop G, et al. Diversity of human colonic butyrate-producing bacteria revealed by analysis of the butyryl-CoA: acetate CoA-transferase gene. Environ Microbiol. 2010;12(2):304–314. doi:10.1111/j.1462-2920.2009.02066.x
74. Yang L, Li A, Wang Y, et al. Intratumoral microbiota: roles in cancer initiation, development and therapeutic efficacy. Signal Transduct Target Ther. 2023;8(1):35. doi:10.1038/s41392-022-01304-4
75. Liu N, Yi C, Wei L, et al. The intratumor mycobiome promotes lung cancer progression via myeloid-derived suppressor cells. Cancer Cell. 2023;41(11):1927–1944.e9. doi:10.1016/j.ccell.2023.08.012
76. Purushothaman S, Meola M, Egli A. Combination of whole genome sequencing and metagenomics for microbiological diagnostics. Int J Mol Sci. 2022;23(17):9834. doi:10.3390/ijms23179834
77. Ngara TR, Zhang H. Recent advances in function-based metagenomic screening. Genomics Proteomics Bioinf. 2018;16(6):405–415. doi:10.1016/j.gpb.2018.01.002
78. Xue C, Chu Q, Zheng Q, et al. Current understanding of the intratumoral microbiome in various tumors. Cell Rep Med. 2023;4(1):100884. doi:10.1016/j.xcrm.2022.100884
79. Haghighi M, Bryce C, Paulsen JD, et al. Digital pathology in the detection of infectious microorganisms: an evaluation of its strengths and weaknesses across a panel of immunohistochemical and histochemical stains routinely used in diagnostic surgical pathology. Arch Pathol Lab Med. 2024;2:22.
80. Prudent E, Raoult D. Fluorescence in situ hybridization, a complementary molecular tool for the clinical diagnosis of infectious diseases by intracellular and fastidious bacteria. FEMS Microbiol Rev. 2019;43(1):88–107. doi:10.1093/femsre/fuy040
81. Massironi S, Facciotti F, Cavalcoli F, et al. Intratumor microbiome in neuroendocrine neoplasms: a new partner of tumor microenvironment? A pilot study. Cells. 2022;11(4):692. doi:10.3390/cells11040692
82. Hu F, Qi G, Kenry, et al. Visualization and in situ ablation of intracellular bacterial pathogens through metabolic labeling. Angew Chem Int Ed Engl. 2020;59(24):9288–9292. doi:10.1002/anie.201910187
83. Chakladar J, Kuo S, Castaneda G, et al. The pancreatic microbiome is associated with carcinogenesis and worse prognosis in males and smokers. Cancers. 2020;12(9):2672. doi:10.3390/cancers12092672
84. Zhang M, Zhang Y, Sun Y, et al. Intratumoral microbiota impacts the first-line treatment efficacy and survival in non-small cell lung cancer patients free of lung infection. J Healthc Eng. 2022;2022:5466853. doi:10.1155/2022/5466853
85. Gomes S, Cavadas B, Ferreira J, et al. Profiling of lung microbiota discloses differences in adenocarcinoma and squamous cell carcinoma. Sci Rep. 2019;9(1):12838. doi:10.1038/s41598-019-49195-w
86. Chen H, Ma Y, Liu Z, et al. Circulating microbiome DNA: an emerging paradigm for cancer liquid biopsy. Cancer Lett. 2021;521:82–87. doi:10.1016/j.canlet.2021.08.036
87. Riglar D, Silver P. Engineering bacteria for diagnostic and therapeutic applications. Nat Rev Microbiol. 2018;16(4):214–225. doi:10.1038/nrmicro.2017.172
88. Tang W, He Y, Zhou S, et al. A novel Bifidobacterium infantis-mediated TK/GCV suicide gene therapy system exhibits antitumor activity in a rat model of bladder cancer. J Exp Clin Cancer Res. 2009;28(1):155. doi:10.1186/1756-9966-28-155
89. Jiang S, Phan T, Nam T, et al. Inhibition of tumor growth and metastasis by a combination of Escherichia coli-mediated cytolytic therapy and radiotherapy. Mol Ther. 2010;18(3):635–642. doi:10.1038/mt.2009.295
90. Jiang S, Park S, Lee H, et al. Engineering of bacteria for the visualization of targeted delivery of a cytolytic anticancer agent. Mol Ther. 2013;21(11):1985–1995. doi:10.1038/mt.2013.183
91. Nakatsuji T, Chen T, Butcher A, et al. A commensal strain of Staphylococcus epidermidis protects against skin neoplasia. Sci Adv. 2018;4(2):eaao4502. doi:10.1126/sciadv.aao4502
92. Battaglia T, Mimpen I, Traets J, et al. A pan-cancer analysis of the microbiome in metastatic cancer. Cell. 2024;187(9):2324–2335.e19. doi:10.1016/j.cell.2024.03.021
93. Galeano N, Wu H, LaCourse K, et al. Effect of the intratumoral microbiota on spatial and cellular heterogeneity in cancer. Nature. 2022;611(7937):810–817. doi:10.1038/s41586-022-05435-0
94. Fu A, Yao B, Dong T, et al. Tumor-resident intracellular microbiota promotes metastatic colonization in breast cancer. Cell. 2022;185(8):1356–1372.e26. doi:10.1016/j.cell.2022.02.027
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.