")
Back to Journals » Infection and Drug Resistance » Volume 12
Prevalence Of mcr-1 Among Cefotaxime-Resistant Commensal Escherichia coli In Residents Of Vietnam
Authors Kawahara R , Khong DT, Le HV, Phan QN , Nguyen TN, Yamaguchi T, Kumeda Y, Yamamoto Y
Received 24 July 2019
Accepted for publication 9 October 2019
Published 23 October 2019 Volume 2019:12 Pages 3317—3325
DOI https://doi.org/10.2147/IDR.S224545
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Joachim Wink
Ryuji Kawahara,1,* Diep Thi Khong,2,* Ha Viet Le,2 Quang Ngoc Phan,2 Thang Nam Nguyen,2 Takahiro Yamaguchi,1 Yuko Kumeda,3 Yoshimasa Yamamoto4,5
1Department of Microbiology, Osaka Institute of Public Health, Osaka, Japan; 2Center of Medical, Pharmaceutical Science and Technology Services, Thai Binh University of Medicine and Pharmacy, Thai Binh, Vietnam; 3Research Center for the 21st Century, Osaka Prefecture University, Osaka, Japan; 4Life Science Research Center, Gifu University, Gifu, Japan; 5Graduate School of Pharmaceutical Sciences, Osaka University, Osaka, Japan
*These authors contributed equally to this work
Correspondence: Yoshimasa Yamamoto
Life Science Research Center, Gifu University, 1-1 Yanagido, Gifu 501-1194, Japan
Tel/fax +81 58 230 6239
Email [email protected]
Purpose: The dissemination of colistin-resistant bacteria harboring the colistin-resistance gene mcr-1 in developing countries has recently entered the spotlight as an emerging public health threat, which is attributed to the abuse of colistin use in these countries. However, the prevalence of these bacteria in developing countries has not been extensively investigated. Therefore, in the present study, we examined the prevalence of cefotaxime-resistant commensal Escherichia coli harboring mcr-1 among residents of a representative Vietnamese village and assessed the characteristics of these isolates.
Materials and methods: The stool samples, one stool sample per resident, of 612 residents were cultured on MacConkey agar with cefotaxime. Resulting E. coli-like colonies were isolated and examined further for the presence of colistin-resistant extended-spectrum β-lactamase (ESBL)-producing E. coli with mcr-1. Antibiotic susceptibility tests were performed, and clonal relationship among colistin-resistant isolates was assessed.
Results: Thirty-one of the 451 cefotaxime-resistant E. coli isolates were resistant to colistin and the majority possessed mcr-1, blaCTX-M, and/or blaTEM, except for two isolates that produced the AmpC β-lactamase. All mcr-1 ESBL-producing E. coli isolates were multidrug-resistant (5–11 antibiotics). The isolates contained various plasmid replicon types, including the most prevalent types IncHI2 (54.8%), IncFIB (48.4%), and IncN (41.9%). In addition, 83.9% of the mcr-1 ESBL-E. coli isolates possessed a transposon ISApl1-mcr-1 segment. Furthermore, 77.4% of the mcr-1 ESBL-E. coli isolates belonged to phylogenetic group A. Pulsed-field gel electrophoresis analysis indicated limited clonal expansion of a specific strain.
Conclusion: These results demonstrate the wide dissemination of colistin-resistant ESBL-E. coli harboring mcr-1 among commensal bacteria of rural residents in Vietnam, suggesting possible mobilization of the mcr-1 gene among ESBL-producing microbiota, which is a great public health concern.
Keywords: mcr-1, residents, stool specimens, commensal bacteria, Vietnam
Introduction
Colistin has been recognized as a last-resort antibiotic for the treatment of intractable infections involving multidrug-resistant (MDR) gram-negative bacteria, especially carbapenem-resistant MDR bacteria.1 However, the increasing usage of colistin in food-producing animals has resulted in a rising prevalence of colistin-resistant bacteria.2 Moreover, the mobile colistin-resistant gene mcr-1 was recently discovered in Escherichia coli isolates from animals and humans in China.3 To date, MCR-1-producing E. coli strains have been reported in many countries throughout Europe, Asia, and South and North America.4–6 Therefore, the possible global dissemination of mcr-1-positive bacteria has become a serious public health concern.
Although the characteristics of animal and human E. coli isolates harboring mcr-1 have been examined in detail,1,3,4,7,8 there is limited comprehensive information regarding the prevalence and characteristics of colistin-resistant bacteria harboring mcr-1 among human residents of developing countries. Because the human intestinal flora is an important reservoir of resistance genes,9,10 the prevalence and characteristics of colistin-resistance in human commensal E. coli should be taken into account in prevention and control efforts. Therefore, the aim of this study was to explore the prevalence of mcr-1-positive E. coli in the feces of residents of a local community in Thai Binh province, Vietnam, which is known to be widely contaminated with extended-spectrum β-lactamase (ESBL)-producing bacteria,11 and to investigate the molecular characteristics of these strains. These data should provide a useful resource for understanding current threats in the region to help initiate appropriate screening, treatment, and control.
Materials And Methods
Specimen Collection
The bacterial strains used in this study were originally isolated from residents for assessment of the dissemination of ESBL-producing bacteria in Nguyen Xa village, Thai Binh province, Vietnam, from September 2013 to February 2016. The village is a representative rural community in this province, with a reported population of 7730 residents in 2008 households at the time of the study. A total of 612 asymptomatic healthy resident volunteers who had not received any medical treatment for the last 3 months participated in the study. One stool specimen was obtained from each participant using a transport swab with Cary-Blair transport medium (Eiken Chemical, Tokyo, Japan). The study was approved by the ethics committees of Osaka University and Thai Binh University of Medicine and Pharmacy. All participants provided written informed consent and that this was conducted in accordance with the Declaration of Helsinki.
Isolation Of Cefotaxime-Resistant E. coli
Stool specimens were plated on MacConkey agar (Nissui, Tokyo, Japan) supplemented with 1 mg/L cefotaxime (CTX-MacConkey) and incubated at 37°C for 18–20 hrs as previously described.11 Resulting colonies exhibiting E. coli characteristics were isolated and confirmed as E. coli using biochemical tests with triple sugar iron slants, lysine indole motility medium (BD, New Jersey, USA), cellobiose lactose indole β-glucuronidase medium (Nissui, Tokyo, Japan), and API 20E (bioMerieux, Marcy l’Etoile, France).
Detection Of ESBL- And AmpC-Producing E. coli
The presence of ESBLs was confirmed using the double-disc synergy test using CTX and ceftazidime with and without clavulanic acid, as recommended by the Clinical and Laboratory Standards Institute (CLSI 2014, Wayne, PA, USA). Cefoxitin (FOX)-resistant strains were classified as putative AmpC-producing E. coli.
Genetic detection and genotyping of β-lactamases were performed by PCR using boiled bacterial suspensions in Tris-EDTA buffer as the template. PCR analysis was performed with universal primers specific to the TEM, SHV, CTX-M-1, CTX-M-2, CTX-M-9, and CTX-M-8/25 groups, as previously described.12 The PCR products were visualized by 2% agarose gel electrophoresis and stained with GelRed nucleic acid (Biotium, Hayward, CA, USA). The sequence analysis of CTX-M genes was performed as previously described.13
Phylogenetic Determination
Phylogenetic groups were determined by triplex PCR using a combination of two genes, chuA and yjaA, and the DNA fragment TSPE4.C2 as previously described.14
mcr-1 Detection And Colistin MIC
The presence of the colistin resistance gene mcr-1 was detected by PCR with bacterial DNA and sequencing of the resulting products.3 The colistin MIC of the mcr-1-positive isolates was evaluated by ETEST® (bioMerieux) according to the manufacturer’s protocol.
Antibiotics Susceptibility
The susceptibility of the ESBL- or AmpC-producing E. coli isolates harboring mcr-1 to 14 antimicrobial agents was tested using the disc diffusion method following the standard procedure of the CLSI, as previously described.15 The disc diffusion test included the following antibiotics: ampicillin (AMP), FOX, CTX, ceftazidime (CAZ), meropenem (MEM), streptomycin (STR), kanamycin (KAN), gentamicin (GEN), ciprofloxacin (CIP), nalidixic acid (NAC), tetracycline (TET), chloramphenicol (CHL), fosfomycin (FOF), and sulfamethoxazole-trimethoprim (SXT).
Pulsed-Field Gel Electrophoresis (PFGE) Analysis
XbaI-digested genomic DNA samples from the E. coli isolates were analyzed on a CHEF-DR III System (Bio-Rad, Hercules, CA, USA) following the detailed methods reported previously.16
Plasmid Replicon Typing
Plasmid replicon typing of the mcr-1-harboring E. coli isolates was assessed by multiplex PCR as previously described.17–20
Transposon ISApl1 Detection
The presence of ISApl1-mcr-1 was assessed by PCR with TaKara Gflex DNA polymerase (TaKaRa Bio Inc., Shiga, Japan) and the primers ISApl1-mcr1-F (5′-GCGCAAAATCGCAGTCG-3′) and ISApl1-mcr1-R (5′-TGTAGGGCATTTTGGAGCATG-3′) according to the manufacturer’s instructions. The cycle conditions were as follows: initial denaturation at 95°C for 5 mins; 30 cycles of denaturation for 10 s at 98°C, annealing for 15 s at 55°C, and extension for 1 min at 68°C; and a final extension at 68°C for 7 mins. The approximate 1000-bp PCR products were verified by direct sequencing.
Results
Prevalence Of Colistin-Resistant E. coli Harboring mcr-1 In Vietnamese Residents
The characteristics of the 612 participants are listed in Table 1. Bacterial growth on CTX-MacConkey was assessed for all participant stool specimens. One colony exhibiting E. coli characteristics from each participant specimen was obtained and subjected to further microbiological analysis. ESBL production was observed in 375 of the 451 E. coli isolates (83.1%). Moreover, 21 of the 451 isolates (4.7%) were AmpC producers.
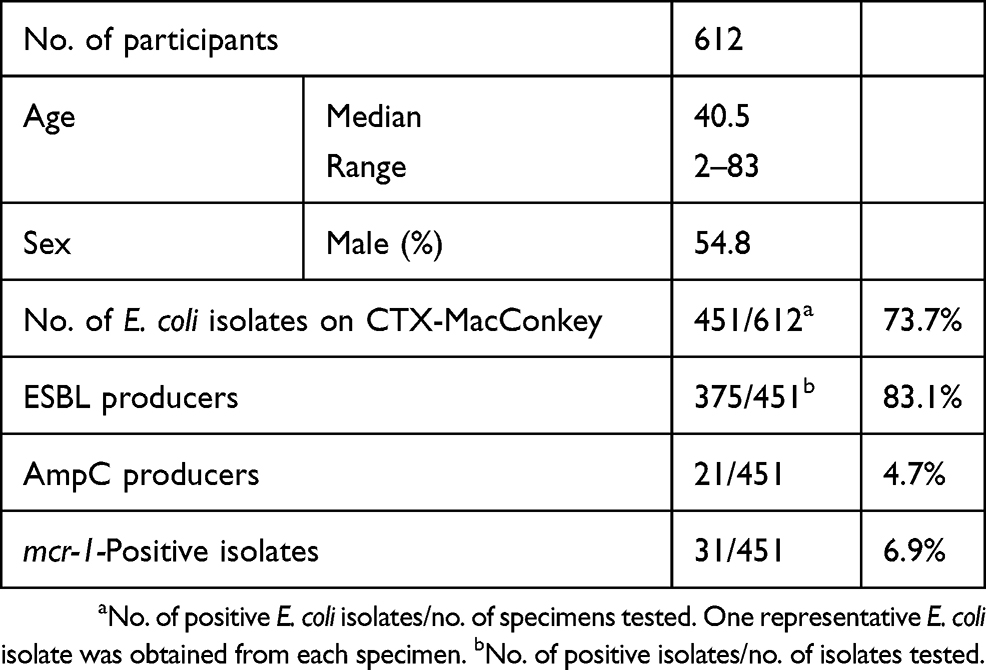 |
Table 1 Prevalence Of Colistin-Resistant E. Coli Harboring Mcr-1 In Residents In Vietnam |
PCR analysis revealed that 31 of the 451 isolates (6.9%) possessed the mcr-1 gene.
Characteristics Of Colistin-Resistant E. coli Isolates Harboring mcr-1
As shown in Table 2, the colistin MIC values of the mcr-1-positive E. coli isolates ranged from 3 to 6 µg/mL, as determined using ETEST. Phylogenetic typing of the isolates showed that group A was the most prevalent phenotype of the mcr-1-positive E. coli isolates (77.4%, 24/31), with other less common groups identified (B1, 6.5%; B2, 6.5%; D, 9.7%).
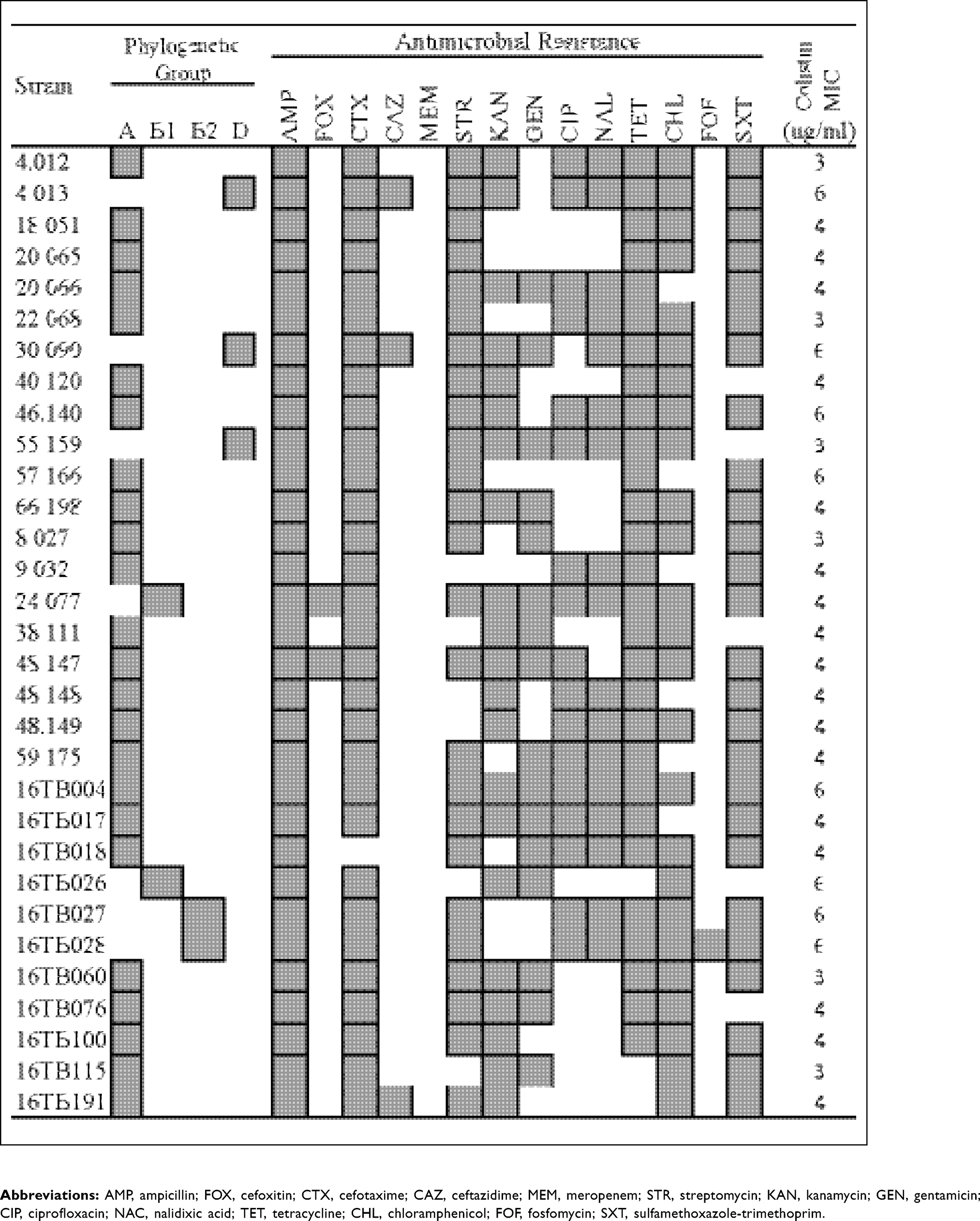 |
Table 2 Characteristics Of Colistin-Resistant E. Coli Isolates Harboring mcr-1 |
Assessment of antibiotic susceptibility showed nearly 100% resistance to AMP and CTX, while resistance to FOX, a second-generation cephamycin, and CAZ, a third-generation cephalosporin, was only 6.5% and 9.7%, respectively. Resistance levels to aminoglycosides, tetracycline, phenicols, and folic acid inhibitors were quite high (51.6–90.3%). Moreover, over half of the mcr-1-harboring E. coli isolates exhibited resistance (54.8%) to quinolones. However, no carbapenem-resistant isolates were identified.
Genetic Features Of Colistin-Resistant E. coli Isolates Harboring mcr-1
Genetic analysis showed that 74.2% (23/31) of the mcr-1-harboring E. coli isolates belonged to the CTX-M-9 group (Table 3). The blaCTX-M-9 gene was identified in 11 of 23 CTX-M-9 group isolates. Twelve (40%, 12/30) ESBL-producing isolates possessed both the CTX-M and TEM genes.
 |
Table 3 Genetic Features Of Colistin-Resistant E. Coli Isolates Harboring mcr-1 |
A wide variety of plasmid replicon types were present in the mcr-1-harboring E. coli isolates. The most frequently detected replicon type was HI2 (54.8%), followed by FIB (48.4%) and N (41.9%), whereas B/C, FIC, HI1, U, X2, and X3 were not detected. Replicon types A/C, FIB, and P were detected in the E. coli isolates harboring mcr-1 as a single replicon type.
The presence of the transposon, ISApl1, a key component contributing to the mobilization of mcr-1,21 in the mcr-1 E. coli isolates was also investigated. Because the PCR used in this study targeted only the ISApl1-mcr-1 segment, only the upstream portion of ISApl1 that is directly attached to mcr-1 was detected. The majority of mcr-1-harboring E. coli isolates (83.9%, 26/31) had the ISApl1-mcr-1 segment.
Clonal Relationship Among Colistin-Resistant E. coli Isolates Harboring mcr-1
PFGE was conducted to assess the clonal relationship among the 31 mcr-1 E. coli isolates, which included 24 phylogenetic group A isolates. Representative dendrograms of PFGE of the isolates are shown in Figure 1. A common PFGE pattern was only observed in strains 16TB017/16TB018 and 16TB027/16TB028. Strains 16TB017 and 16TB018 were isolated from residents belonging to the same household, and strains 16TB027 and 16TB028 were also isolated from the same, albeit different, households. The remaining isolates did not demonstrate any similarity.
 |
Figure 1 Representative dendrogram of PFGE patterns of colistin-resistant E. coli isolates harboring mcr-1. |
Discussion
This study revealed that the prevalence of colistin/CTX-resistant commensal E. coli harboring mcr-1 among residents of a rural community in northern Vietnam is as high as 5.1% (31 positives/612 residents tested), although the isolates assessed were selected based only on CTX resistance. Trung et al7 also reported a relatively high prevalence (17.9%) of mcr-1-carrying bacteria among residents of a rural area in South Vietnam. We have also detected a very high prevalence (69.4%) of colistin-resistant E. coli with mcr in stool specimens of healthy residents in Vietnam using selective medium for colistin-resistant bacteria, such as CHROMagarTM COL-APSE (CHROMagar, Paris, France), as primary selection for isolation of bacteria.22 Therefore, the prevalence of colistin-resistant bacteria seems to be dependent on the medium used for isolation of bacteria. Nevertheless, the results of these recent studies, including the present study, indicate a wide dissemination of mcr-1-positive commensal E. coli in Vietnam. In addition, the finding of such non-negligible numbers of isolates possessing both ESBL and colistin-resistance genes highlights a high risk of extensively drug-resistant bacteria circulating in the area.
Dissemination studies conducted in other countries demonstrated the absence of mcr-1 isolates in healthy people of Switzerland23 and the Netherlands.24 In contrast, 4.9% of the Chinese resident samples25 were found to be carriers of mcr-1-positive bacteria. Therefore, at the moment, the degree of the spread of E. coli harboring mcr-1 in healthy individuals has only been demonstrated in Asian countries, although there have been several reports of bacteria harboring mcr-1 in food and food-producing animals worldwide.26
Phylogenetic analysis revealed that the majority of the mcr-1-harboring E. coli isolates belonged to phylogenetic group A, with a few group B1, B2, and D isolates detected. These results are similar to the prevalence of phylogenetic groups in the normal human intestinal microbiota; A is the predominant group, followed by B2, whereas B1 and D are less common.27 Thus, the frequency of mcr-1-harboring E. coli clone phylogenetic groups in the residents of this Vietnamese village may reflect the normal intestinal microbiota in humans.
Analysis of the antibiotic susceptibility of the mcr-1-harboring E. coli isolates revealed that all isolates were MDR, ie, resistant to at least one antibiotic drug from three or more antibiotic classes.28 However, resistance to FOX and CAZ was less common among these isolates. In addition, the overall antibiotic resistance profile of the mcr-1-harboring E. coli isolates was similar to that reported from ESBL-producing E. coli isolated from food in Vietnam.12 Therefore, it is possible that the mcr-1 gene may have been transmitted to ESBL-producing E. coli.
Indeed, nearly all of the mcr-1-harboring E. coli isolates were ESBL producers except for two isolates, which were class C β-lactamase producers, as determined by both phenotypic and genetic assays. The majority of these ESBL-mcr-1 isolates possessed CTX-M-9 group genes; the blaCTX-M-14 gene was identified in nearly one-third of the CTX-M-9 group and mcr-1 isolates tested. In contrast, a recent paper on MDR E. coli harboring mcr-1 reported that most isolates from pig were co-harboring CTX-M-1 group genes.29 The difference between this previous study and our study may be due to the difference in specimen source.
Although it remains unclear whether the mcr-1 and CTX-M genes are located on the same plasmid, the present results demonstrate that ESBL-producing E. coli isolates with CTX-M-9 group genes are dominant among the colistin-resistant commensal E. coli in residents of the community. A previous study showed that mcr-1 and CTX-M-9 group genes are co-transferred by one or more plasmid types.30 Moreover, the mcr-1-carrying plasmid identified in the first-ever discovered mcr-1 isolate is known to harbor the CTX-M-9 group, blaCTX-M-14.31
Plasmid type plays an important role in the spread of mcr-1, not only in the human intestinal microbiota but also in community settings. Therefore, characterization of the plasmids in mcr-1-harboring E. coli is important for understanding their dissemination. Replicon typing of the present isolates revealed a high diversity of plasmid backbones. The major replicon types of the isolates were HI2 (54.8%), FIB (48.4%), and N (41.9%), which have been reported as mcr-1-associated replicon types.3,32,33 Replicon type I2, which is known as an mcr-1 disseminator,34 was detected in 22.6% (7/31) of the isolates. Moreover, plasmid replicon type X4 was identified in one of our isolates, which was recently identified as a common mcr-1-carrying plasmid replicon type in Enterobacteriaceae from food, animal, and human samples recovered in different countries.5 However, since we assessed the replicon type of all plasmids in the isolates, the replicon types of the plasmid(s) harboring the mcr-1 gene remain unclear. In spite of this limitation, these results indicate that the diversity of plasmid types among mcr-1 isolates may contribute to the successful spread of the mcr-1 gene among different E. coli clones.
Besides class 1 integrons,35 the transposon, ISApl1, is also known to be involved in horizontal gene transfer and maybe a key factor contributing to the widespread dissemination of the mcr-1 gene.36 In this study, transposon ISApl1 with mcr-1 was detected in the majority of the isolates, except for six. As ISApl1 can be lost through improper recombination,36 the ISApl1-mcr-1 segment may not have been detected in all of the mcr-1 harboring isolates. Zurfluh et al33 reported that the mcr-1 gene may be mobilized independently. The possible occurrence of the mcr-1 gene or gene module mobilization among plasmids in the E. coli clones is also supported by the PFGE analysis, showing the limited clonal expansion of mcr-1-harboring E. coli.
Overall, this study suggests that the wide dissemination of colistin-resistant ESBL-producing E. coli harboring mcr-1 in commensal bacteria in Vietnamese residents involves diverse clones with a variety of plasmid replicon types. Mobilization of the mcr-1 gene module among microbiota under certain circumstances such as exposure to colistin including suboptimal treatment during therapy is speculated as one of the likely causes of this dissemination.37 Such wide dissemination of the mcr-1 gene in the commensal bacteria of healthy people constitutes a great public health concern in regard to the prevention, monitoring, and treatment of colistin-resistant bacteria.
Disclosure
Dr Yoshimasa Yamamoto reports grants from Japan AMED/JICA, and Japan Society for Promotion of Science, during the conduct of the study. The authors report no other conflicts of interest in this work.
References
1. Poirel L, Jayol A, Nordmann P. Polymyxins: antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin Microbiol Rev. 2017;30:557–596. doi:10.1128/CMR.00064-16
2. Olaitan AO, Diene SM, Kempf M, et al. Worldwide emergence of colistin resistance in Klebsiella pneumoniae from healthy humans and patients in Lao PDR, Thailand, Israel, Nigeria and France owing to inactivation of the PhoP/PhoQ regulator mgrB: an epidemiological and molecular study. Int J Antimicrob Agents. 2014;44:500–507. doi:10.1016/j.ijantimicag.2014.07.020
3. Liu YY, Wang Y, Walsh TR, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16:161–168. doi:10.1016/S1473-3099(15)00424-7
4. Poirel L, Kieffer N, Brink A, et al. Genetic features of MCR-1-producing colistin-resistant escherichia coli isolates in South Africa. Antimicrob Agents Chemother. 2016;60:4394–4397. doi:10.1128/AAC.00444-16
5. Fernandes MR, McCulloch JA, Vianello MA, et al. First report of the globally disseminated IncX4 plasmid carrying the mcr-1 gene in a colistin-resistant escherichia coli sequence Type 101 isolate from a human infection in Brazil. Antimicrob Agents Chemother. 2016;60:6415–6417. doi:10.1128/AAC.01325-16
6. Fernandes MR, Moura Q, Sartori L, et al. Silent dissemination of colistin-resistant Escherichia coli in South America could contribute to the global spread of the mcr-1 gene. Euro Surveill. 2016;21. doi:10.2807/1560-7917.ES.2016.21.17.30214
7. Trung NV, Matamoros S, Carrique-Mas JJ, et al. Zoonotic transmission of mcr-1 colistin resistance gene from small-scale poultry farms, Vietnam. Emerg Infect Dis. 2017;23:529–532. doi:10.3201/eid2303.161553
8. Zurfluh K, Kieffer N, Poirel L, et al. Features of the mcr-1 cassette related to colistin resistance. Antimicrob Agents Chemother. 2016;60:6438–6439. doi:10.1128/AAC.01519-16
9. Huddleston JR. Horizontal gene transfer in the human gastrointestinal tract: potential spread of antibiotic resistance genes. Infect Drug Resist. 2014;7:167–176. doi:10.2147/IDR.S48820
10. Rolain JM. Food and human gut as reservoirs of transferable antibiotic resistance encoding genes. Front Microbiol. 2013;4:173. doi:10.3389/fmicb.2013.00077
11. Nakayama T, Ueda S, Huong BT, et al. Wide dissemination of extended-spectrum beta-lactamase-producing Escherichia coli in community residents in the Indochinese peninsula. Infect Drug Resist. 2015;8:1–5. doi:10.2147/IDR.S74934
12. Le QP, Ueda S, Nguyen TN, et al. Characteristics of extended-spectrum beta-lactamase-producing escherichia coli in retail meats and shrimp at a local market in Vietnam. Foodborne Pathog Dis. 2015;12:719–725. doi:10.1089/fpd.2015.1954
13. Harada T, Yamane R, Dang VC. et al. Prevalence and antimicrobial susceptibility of enterobacteriaceae isolated from retail pepper in Vietnam. J Food Prot;2017. 716–724. doi:10.4315/0362-028X.JFP-16-501
14. Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl Environ Microbiol. 2000;66:4555–4558. doi:10.1128/aem.66.10.4555-4558.2000
15. Ueda S, Ngan BT, Huong BT, et al. Limited transmission of bla(CTX-M-9)-type-positive Escherichia coli between humans and poultry in Vietnam. Antimicrob Agents Chemother. 2015;59:3574–3577. doi:10.1128/AAC.00517-15
16. Ribot EM, Fair MA, Gautom R, et al. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157: H7,Salmonella, and Shigella for PulseNet. Foodborne Pathog Dis. 2006;3:59–67. doi:10.1089/fpd.2006.3.59
17. Johnson TJ, Wannemuehler YM, Johnson SJ, et al. Plasmid replicon typing of commensal and pathogenic Escherichia coli isolates. Appl Environ Microbiol. 2007;73:1976–1983. doi:10.1128/AEM.02171-06
18. Garcia-Fernandez A, Fortini D, Veldman K, et al. Characterization of plasmids harbouring qnrS1, qnrB2 and qnrB19 genes in Salmonella. J Antimicrob Chemother. 2009;63:274–281. doi:10.1093/jac/dkn470
19. Johnson TJ, Bielak EM, Fortini D, et al. Expansion of the IncX plasmid family for improved identification and typing of novel plasmids in drug-resistant Enterobacteriaceae. Plasmid. 2012;68:43–50. doi:10.1016/j.plasmid.2012.03.001
20. Chen L, Chavda KD, Al Laham N, et al. Complete nucleotide sequence of a blaKPC-harboring IncI2 plasmid and its dissemination in New Jersey and New York hospitals. Antimicrob Agents Chemother. 2013;57:5019–5025. doi:10.1128/AAC.01397-13
21. Snesrud E, Ong AC, Corey B, et al. Analysis of serial isolates of mcr-1-positive escherichia coli reveals a highly active ISApl1 transposon. Antimicrob Agents Chemother. 2017;61. doi:10.1128/AAC.00056-17
22. Yamamoto Y, Kawahara R, Fujiya Y, et al. Wide dissemination of colistin-resistant Escherichia coli with the mobile resistance gene mcr in healthy residents in Vietnam. J Antimicrob Chemother. 2018;74:523–524.
23. Zurfluh K, Stephan R, Widmer A, et al. Screening for fecal carriage of MCR-producing Enterobacteriaceae in healthy humans and primary care patients. Antimicrob Resist Infect Control. 2017;6:28. doi:10.1186/s13756-017-0186-z
24. Kluytmans-van Den Bergh MF, Huizinga P, Bonten MJ, et al. Presence of mcr-1-positive Enterobacteriaceae in retail chicken meat but not in humans in the Netherlands since 2009. Euro Surveill. 2016;21:30149. doi:10.2807/1560-7917.ES.2016.21.9.30149
25. Bi Z, Berglund B, Sun Q, et al. Prevalence of the mcr-1 colistin resistance gene in extended-spectrum beta-lactamase-producing Escherichia coli from human faecal samples collected in 2012 in rural villages in Shandong Province, China. Int J Antimicrob Agents. 2017;49:493–497. doi:10.1016/j.ijantimicag.2016.12.018
26. Schwarz S, Johnson AP. Transferable resistance to colistin: a new but old threat. J Antimicrob Chemother. 2016;71:2066–2070. doi:10.1093/jac/dkw274
27. Tenaillon O, Skurnik D, Picard B, et al. The population genetics of commensal Escherichia coli. Nat Rev Microbiol. 2010;8:207–217. doi:10.1038/nrmicro2298
28. Magiorakos AP, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18:268–281. doi:10.1111/j.1469-0691.2011.03570.x
29. Shafiq M, Huang J, Ur Rahman S, et al. High incidence of multidrug-resistant Escherichia coli coharboring mcr-1 and bla CTX-M-15 recovered from pigs. Infect Drug Resist. 2019;12:2135–2149. doi:10.2147/IDR.S209473
30. Liu X, Li R, Zheng Z, et al. Molecular characterization of escherichia coli isolates carrying mcr-1, fosA3, and extended-spectrum-beta-lactamase genes from food samples in China. Antimicrob Agents Chemother. 2017;61. doi:10.1128/AAC.00064-17
31. Zhi C, Lv L, Yu LF, et al. Dissemination of the mcr-1 colistin resistance gene. Lancet Infect Dis. 2016;16:292–293. doi:10.1016/S1473-3099(16)00063-3
32. Nordmann P, Lienhard R, Kieffer N, et al. Plasmid-mediated colistin-resistant escherichia coli in bacteremia in Switzerland. Clin Infect Dis. 2016;62:1322–1323. doi:10.1093/cid/ciw124
33. Zurfluh K, Klumpp J, Nuesch-Inderbinen M, et al. Full-length nucleotide sequences of mcr-1-harboring plasmids isolated from extended-spectrum-beta-lactamase-producing escherichia coli isolates of different origins. Antimicrob Agents Chemother. 2016;60:S5589–91. doi:10.1128/AAC.00935-16
34. Tijet N, Faccone D, Rapoport M, et al. Molecular characteristics of mcr-1-carrying plasmids and new mcr-1 variant recovered from polyclonal clinical Escherichia coli from Argentina and Canada. PLoS One. 2017;12:e0180347. doi:10.1371/journal.pone.0180347
35. Bagheri-Nesami M, Rezai MS, Ahangarkani F, et al. Multidrug and co-resistance patterns of non-fermenting Gram-negative bacilli involved in ventilator-associated pneumonia carrying class 1 integron in the North of Iran. Germs. 2017;7:123–131. doi:10.18683/germs.2017.1117
36. Snesrud E, He S, Chandler M, et al. A model for transposition of the colistin resistance gene mcr-1 by ISApl1. Antimicrob Agents Chemother. 2016;60:6973–6976. doi:10.1128/AAC.01457-16
37. Streinu-Cercel A. Colistin in the management of severe infections with multidrug resistant Gram-negative bacilli. Germs. 2014;4:7–8. doi:10.11599/germs.2014.1048
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.